



Quantifying Coexistence Concentrations in Multi-Component Phase-Separating Systems Using Analytical HPLC

 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Rationale for the Proposed Approach
2.2. Details of the Approach
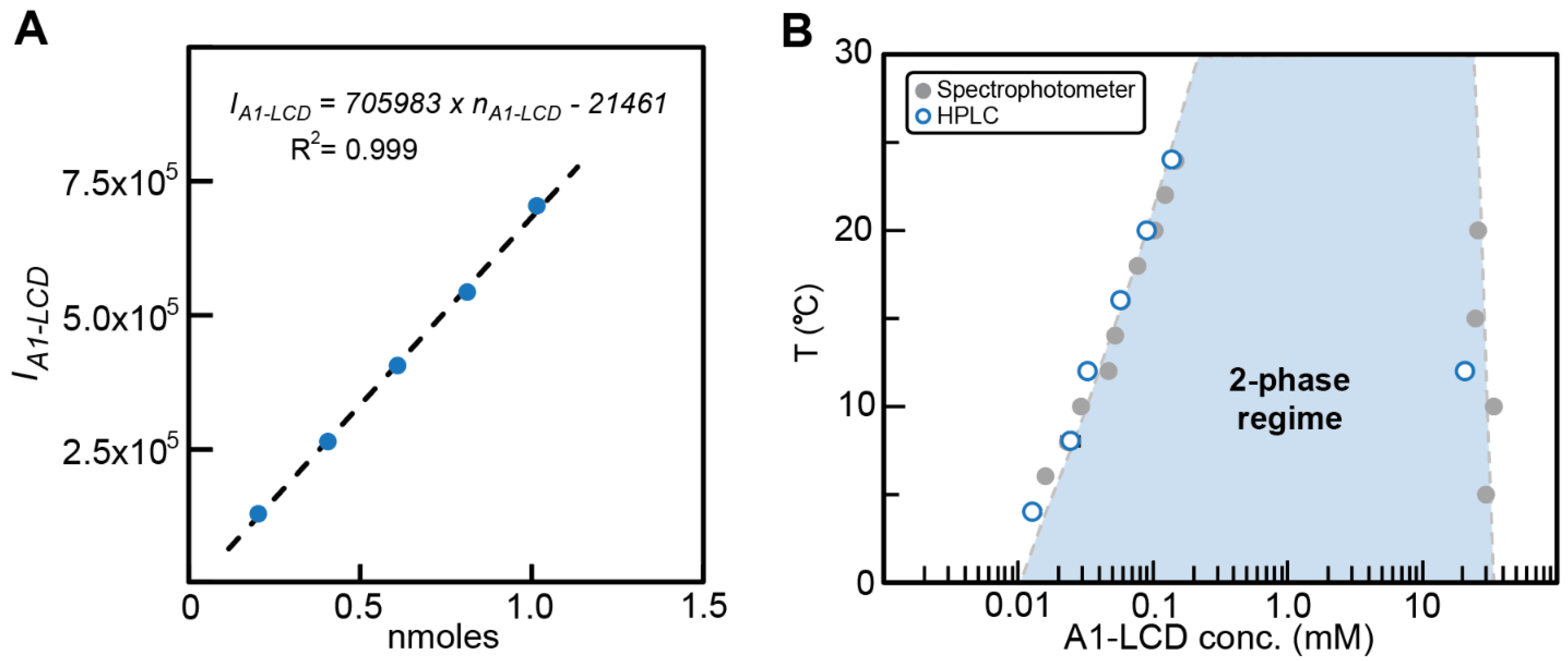
2.3. Validation of the Method with a Single-Component Phase-Separating System
2.4. Application to Multi-Component Phase Separation
2.5. General Considerations for Implementation of the Method
- (a)
- Column: Columns to achieve separation include normal-phase, reverse-phase, ion exchange and size exclusion columns, which are readily available for HPLC systems. The work presented here used C4 or C18 (ReproSil Gold 200; Dr. Maisch) reverse-phase columns.
- (b)
- Mobile phase: The mobile phase solvents used are primarily dictated by the column. The typical chromatographic buffers used for size exclusion and ion exchange are aqueous buffers, while RP-HPLC uses a gradient of organic solvents in water. However, also within the remit of RP-HPLC, there are different options possible for the organic solvent, including acetonitrile, methanol, and tetrahydrofuran. The solvents used must be miscible with water and of HPLC-grade quality to minimize their contribution to the absorbance signals measured. In this work, gradients used involved the mixing of H2O + 0.1% TFA (trifluoroacetic acid) with pure acetonitrile. 0.1% TFA yields a pH of 2.1 ensuring full ionization of analytes and acts as a weak ion-pairing agent thereby conferring more uniform binding of each analyte. This yields sharper peaks and more reproducible elution profiles. Use of TFA in just water, and not in the acetonitrile, is employed as it effectively adds an ion exchange component to the RP-HPLC separation and can result in better peak separation. Of note, the low pH results in the denaturation of protein structure. In cases where it is desirable to recover the components, reverse-phase columns are only suitable if the macromolecules readily refold.
- (c)
- Gradient: Optimization of the gradient is required to obtain sufficient separation between eluting species. It is important to consider sufficient equilibration time given the column volume if step changes are made at any point in the overall gradient run. The appropriate combination of points a, b and c is key to a successful use of the HPLC methodology and likely requires iteration for optimization based on the types of samples that are being studied.
- (d)
- Detection: HPLC systems may have different detection capabilities ranging from a single absorbance wavelength to setups with photodiode array detectors providing absorbance spectra rather than single wavelengths, or even fluorescence detectors. This work made use of an HPLC system with a dual-selectable wavelength detector. The selection of wavelengths to be monitored will depend on the macromolecule of interest. Typical choices include 280 nm for proteins containing aromatic residues, 260 nm for nucleic acids and 230 to 215 nm for proteins lacking aromatic residues. The monitored wavelength should also avoid interference from solvent components.
- (e)
- Column loading: The range of volumes that can be injected onto the column will depend on the system at hand. Injection of accurate volumes, a prerequisite for accurate determination of coexistence concentrations, is most easily achieved with an autoinjector. Further, the amount of macromolecule of interest in the sample should yield an absorbance signal in the linear range of the detector as confirmed through the standard curve. The amounts for which this can be achieved will vary based on the extinction coefficient of the molecule, the wavelength being monitored, and the width of the elution peak, which can be optimized by solvent choice and gradient properties. A further consideration is that loading of high concentrations of some buffer components such as glycerol or PEG can lead to contamination and ultimately damage the column.
- (f)
- Washing: It is good practice to perform wash programs/cycles between batches of samples to ensure that the column remains in good working order and is frequently cleaned. This avoids material or contamination from previous runs interfering with following measurements.
- (g)
- Tests: Routine running of blank injections using the method gradient is valuable to check that sample material has not been retained on the column. Retention in the column can lead to subsequent elution that interferes with the quantitation of components in injected samples.
- (h)
- Sample recovery: If the HPLC system is coupled to a fraction collector, the eluted peaks can be collected to recover sample components. In the case of RP-HPLC, these fractions are best dried on a speed-vac and then resuspended in the buffer of choice. Keep in mind however, that as RP-HPLC denatures the protein, structured proteins need to be refolded.Considerations regarding handling of dense phase:
- (i)
- Viscosity of dense phase: The dense phase is highly viscous and needs to be carefully pipetted. We recommend the use of a positive displacement pipette to minimize errors and achieve accurate volumes (see also [41]. The variability in the measured dense phase concentrations is higher than the measured dilute phase concentrations as can be seen in Figure 2B and Figure 3C,D, but the percentage errors are relatively small. Compared to error sources in other approaches for determining dense phase concentrations, e.g., microscopic determination of fluorescence intensity in the dense phase, the error contribution from pipetting the viscous dense phase is relatively small and manageable. Several replicate measurements should be performed to get a sense of their precision.
- (j)
- Sample requirements: The required biomolecule amounts to generate sufficient dilute phase for detection depend almost exclusively on the extinction coefficient of the biomolecule. Dense phase requirements can be more limiting. We typically remove 2 μL of dense phase for dilution and subsequent injection into the HPLC. The amount of protein needed to generate a slightly larger volume of dense phase depends on the dilute vs. dense phase concentrations and the concentration of the stock solution. If we, e.g., consider the hnRNPA1 LCD (Figure 2B) with dilute and dense phase concentrations at 20 °C of ~100 μM and ~20 mM, a stock solution used to generate a dense phase sample could be 100 μL of a 1 mM protein. Induction of phase separation (by addition of NaCl to 150 mM final concentration in this experiment) would result in approximately 95.5 μL of 100 μM dilute phase and 4.5 μL of 20 mM dense phase. Notably, the resulting dense phase volume is not only determined by the total amount of protein but also by how far above the saturation concentration the preparation starts, with higher concentrations capturing a larger fraction of protein in the dense phase. Less concentrated dense phases require substantially lower protein amounts.
3. Materials and Methods
3.1. Details of Protein Constructs
3.2. Phase Separation Assay
3.3. HPLC instrumentation, Columns, and Solvents
3.4. Calibration of Concentration Measurements by HPLC
3.5. Determination of the Dilute (csat) and Dense Phase (cdense) Concentration Using HPLC
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shin, Y.; Brangwynne, C.P. Liquid phase condensation in cell physiology and disease. Science 2017, 357. [Google Scholar] [CrossRef] [PubMed]
- Banani, S.F.; Lee, H.O.; Hyman, A.A.; Rosen, M.K. Biomolecular condensates: Organizers of cellular biochemistry. Nat. Rev. Mol. Cell Biol. 2017, 18, 285–298. [Google Scholar]
- Holehouse, A.S.; Pappu, R.V. Functional Implications of Intracellular Phase Transitions. Biochemistry 2018, 57, 2415–2423. [Google Scholar] [CrossRef] [PubMed]
- Alberti, S.; Gladfelter, A.; Mittag, T. Considerations and Challenges in Studying Liquid-Liquid Phase Separation and Biomolecular Condensates. Cell 2019, 176, 419–434. [Google Scholar] [CrossRef] [PubMed]
- Mittag, T.; Pappu, R.V. A conceptual framework for understanding phase separation and addressing open questions and challenges. Mol. Cell 2022, 82, 2201–2214. [Google Scholar] [CrossRef]
- Brangwynne, C.P.; Mitchison, T.J.; Hyman, A.A. Active liquid-like behavior of nucleoli determines their size and shape in Xenopus laevis oocytes. Proc. Natl. Acad. Sci. USA 2011, 108, 4334–4339. [Google Scholar] [CrossRef] [PubMed]
- Feric, M.; Vaidya, N.; Harmon, T.S.; Mitrea, D.M.; Zhu, L.; Richardson, T.M.; Kriwacki, R.W.; Pappu, R.V.; Brangwynne, C.P. Coexisting Liquid Phases Underlie Nucleolar Subcompartments. Cell 2016, 165, 1686–1697. [Google Scholar] [CrossRef]
- Sanders, D.W.; Kedersha, N.; Lee, D.S.W.; Strom, A.R.; Drake, V.; Riback, J.A.; Bracha, D.; Eeftens, J.M.; Iwanicki, A.; Wang, A.; et al. Competing Protein-RNA Interaction Networks Control Multiphase Intracellular Organization. Cell 2020, 181, 306–324.e28. [Google Scholar] [CrossRef]
- Yang, P.; Mathieu, C.; Kolaitis, R.M.; Zhang, P.; Messing, J.; Yurtsever, U.; Yang, Z.; Wu, J.; Li, Y.; Pan, Q.; et al. G3BP1 Is a Tunable Switch that Triggers Phase Separation to Assemble Stress Granules. Cell 2020, 181, 325–345.e28. [Google Scholar] [CrossRef]
- Guillen-Boixet, J.; Kopach, A.; Holehouse, A.S.; Wittmann, S.; Jahnel, M.; Schlussler, R.; Kim, K.; Trussina, I.; Wang, J.; Mateju, D.; et al. RNA-Induced Conformational Switching and Clustering of G3BP Drive Stress Granule Assembly by Condensation. Cell 2020, 181, 346–361.e17. [Google Scholar] [CrossRef] [PubMed]
- Xing, W.; Muhlrad, D.; Parker, R.; Rosen, M.K. A quantitative inventory of yeast P body proteins reveals principles of composition and specificity. Elife 2020, 9, e56525. [Google Scholar] [CrossRef] [PubMed]
- Kilic, S.; Lezaja, A.; Gatti, M.; Bianco, E.; Michelena, J.; Imhof, R.; Altmeyer, M. Phase separation of 53BP1 determines liquid-like behavior of DNA repair compartments. EMBO J. 2019, 38, e101379. [Google Scholar] [CrossRef]
- Levone, B.R.; Lenzken, S.C.; Antonaci, M.; Maiser, A.; Rapp, A.; Conte, F.; Reber, S.; Mechtersheimer, J.; Ronchi, A.E.; Muhlemann, O.; et al. FUS-dependent liquid-liquid phase separation is important for DNA repair initiation. J. Cell Biol. 2021, 220. [Google Scholar] [CrossRef] [PubMed]
- Boija, A.; Klein, I.A.; Sabari, B.R.; Dall’Agnese, A.; Coffey, E.L.; Zamudio, A.V.; Li, C.H.; Shrinivas, K.; Manteiga, J.C.; Hannett, N.M.; et al. Transcription Factors Activate Genes through the Phase-Separation Capacity of Their Activation Domains. Cell 2018, 175, 1842–1855.e16. [Google Scholar] [CrossRef] [PubMed]
- Cho, W.K.; Spille, J.H.; Hecht, M.; Lee, C.; Li, C.; Grube, V.; Cisse, I.I. Mediator and RNA polymerase II clusters associate in transcription-dependent condensates. Science 2018, 361, 412–415. [Google Scholar] [CrossRef]
- Boehning, M.; Dugast-Darzacq, C.; Rankovic, M.; Hansen, A.S.; Yu, T.; Marie-Nelly, H.; McSwiggen, D.T.; Kokic, G.; Dailey, G.M.; Cramer, P.; et al. RNA polymerase II clustering through carboxy-terminal domain phase separation. Nat. Struct. Mol. Biol. 2018, 25, 833–840. [Google Scholar] [CrossRef]
- Su, X.; Ditlev, J.A.; Hui, E.; Xing, W.; Banjade, S.; Okrut, J.; King, D.S.; Taunton, J.; Rosen, M.K.; Vale, R.D. Phase separation of signaling molecules promotes T cell receptor signal transduction. Science 2016, 352, 595–599. [Google Scholar] [CrossRef]
- Case, L.B.; Zhang, X.; Ditlev, J.A.; Rosen, M.K. Stoichiometry controls activity of phase-separated clusters of actin signaling proteins. Science 2019, 363, 1093–1097. [Google Scholar] [CrossRef]
- Alberti, S.; Dormann, D. Liquid-Liquid Phase Separation in Disease. Annu. Rev. Genet. 2019, 53, 171–194. [Google Scholar] [CrossRef]
- Boija, A.; Klein, I.A.; Young, R.A. Biomolecular Condensates and Cancer. Cancer Cell 2021, 39, 174–192. [Google Scholar] [CrossRef]
- Bouchard, J.J.; Otero, J.H.; Scott, D.C.; Szulc, E.; Martin, E.W.; Sabri, N.; Granata, D.; Marzahn, M.R.; Lindorff-Larsen, K.; Salvatella, X.; et al. Cancer Mutations of the Tumor Suppressor SPOP Disrupt the Formation of Active, Phase-Separated Compartments. Mol. Cell 2018, 72, 19–36.e18. [Google Scholar] [CrossRef] [PubMed]
- Chandra, B.; Michmerhuizen, N.L.; Shirnekhi, H.K.; Tripathi, S.; Pioso, B.J.; Baggett, D.W.; Mitrea, D.M.; Iacobucci, I.; White, M.R.; Chen, J.; et al. Phase Separation Mediates NUP98 Fusion Oncoprotein Leukemic Transformation. Cancer Discov. 2022, 12, 1152–1169. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.H.; Davis, E.S.; Daugird, T.A.; Zhao, S.; Quiroga, I.Y.; Uryu, H.; Li, J.; Storey, A.J.; Tsai, Y.H.; Keeley, D.P.; et al. Phase separation drives aberrant chromatin looping and cancer development. Nature 2021, 595, 591–595. [Google Scholar] [CrossRef] [PubMed]
- Tulpule, A.; Guan, J.; Neel, D.S.; Allegakoen, H.R.; Lin, Y.P.; Brown, D.; Chou, Y.T.; Heslin, A.; Chatterjee, N.; Perati, S.; et al. Kinase-mediated RAS signaling via membraneless cytoplasmic protein granules. Cell 2021, 184, 2649–2664.e18. [Google Scholar]
- Mackenzie, I.R.; Nicholson, A.M.; Sarkar, M.; Messing, J.; Purice, M.D.; Pottier, C.; Annu, K.; Baker, M.; Perkerson, R.B.; Kurti, A.; et al. TIA1 Mutations in Amyotrophic Lateral Sclerosis and Frontotemporal Dementia Promote Phase Separation and Alter Stress Granule Dynamics. Neuron 2017, 95, 808–816.e9. [Google Scholar] [CrossRef]
- White, M.R.; Mitrea, D.M.; Zhang, P.; Stanley, C.B.; Cassidy, D.E.; Nourse, A.; Phillips, A.H.; Tolbert, M.; Taylor, J.P.; Kriwacki, R.W. C9orf72 Poly(PR) Dipeptide Repeats Disturb Biomolecular Phase Separation and Disrupt Nucleolar Function. Mol. Cell 2019, 74, 713–728.e6. [Google Scholar] [CrossRef]
- Zhang, P.; Fan, B.; Yang, P.; Temirov, J.; Messing, J.; Kim, H.J.; Taylor, J.P. Chronic optogenetic induction of stress granules is cytotoxic and reveals the evolution of ALS-FTD pathology. eLife 2019, 8, e39578. [Google Scholar] [CrossRef]
- Fernandopulle, M.; Wang, G.; Nixon-Abell, J.; Qamar, S.; Balaji, V.; Morihara, R.; St George-Hyslop, P.H. Inherited and Sporadic Amyotrophic Lateral Sclerosis and Fronto-Temporal Lobar Degenerations arising from Pathological Condensates of Phase Separating Proteins. Hum. Mol. Genet. 2019, 28, R187–R196. [Google Scholar] [CrossRef]
- Nedelsky, N.B.; Taylor, J.P. Pathological phase transitions in ALS-FTD impair dynamic RNA-protein granules. Rna 2022, 28, 97–113. [Google Scholar] [CrossRef]
- Mathieu, C.; Pappu, R.V.; Taylor, J.P. Beyond aggregation: Pathological phase transitions in neurodegenerative disease. Science 2020, 370, 56–60. [Google Scholar] [CrossRef]
- Choi, J.M.; Holehouse, A.S.; Pappu, R.V. Physical Principles Underlying the Complex Biology of Intracellular Phase Transitions. Annu. Rev. Biophys. 2020, 49, 107–133. [Google Scholar] [CrossRef] [PubMed]
- Ruff, K.M.; Dar, F.; Pappu, R.V. Polyphasic linkage and the impact of ligand binding on the regulation of biomolecular condensates. Biophys. Rev. 2021, 2, 021302. [Google Scholar] [CrossRef] [PubMed]
- Ruff, K.M.; Dar, F.; Pappu, R.V. Ligand effects on phase separation of multivalent macromolecules. Proc. Natl. Acad. Sci. USA 2021, 118, e2017184118. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Banjade, S.; Cheng, H.C.; Kim, S.; Chen, B.; Guo, L.; Llaguno, M.; Hollingsworth, J.V.; King, D.S.; Banani, S.F.; et al. Phase transitions in the assembly of multivalent signalling proteins. Nature 2012, 483, 336–340. [Google Scholar] [CrossRef]
- Martin, E.W.; Holehouse, A.S.; Peran, I.; Farag, M.; Incicco, J.J.; Bremer, A.; Grace, C.R.; Soranno, A.; Pappu, R.V.; Mittag, T. Valence and patterning of aromatic residues determine the phase behavior of prion-like domains. Science 2020, 367, 694–699. [Google Scholar] [CrossRef]
- Bremer, A.; Farag, M.; Borcherds, W.M.; Peran, I.; Martin, E.W.; Pappu, R.V.; Mittag, T. Deciphering how naturally occurring sequence features impact the phase behaviours of disordered prion-like domains. Nat. Chem. 2022, 14, 196–207. [Google Scholar] [CrossRef]
- Brady, J.P.; Farber, P.J.; Sekhar, A.; Lin, Y.H.; Huang, R.; Bah, A.; Nott, T.J.; Chan, H.S.; Baldwin, A.J.; Forman-Kay, J.D.; et al. Structural and hydrodynamic properties of an intrinsically disordered region of a germ cell-specific protein on phase separation. Proc. Natl. Acad. Sci. USA 2017, 114, E8194–E8203. [Google Scholar] [CrossRef]
- Wei, M.T.; Elbaum-Garfinkle, S.; Holehouse, A.S.; Chen, C.C.; Feric, M.; Arnold, C.B.; Priestley, R.D.; Pappu, R.V.; Brangwynne, C.P. Phase behaviour of disordered proteins underlying low density and high permeability of liquid organelles. Nat. Chem. 2017, 9, 1118–1125. [Google Scholar] [CrossRef]
- Wang, J.; Choi, J.M.; Holehouse, A.S.; Lee, H.O.; Zhang, X.; Jahnel, M.; Maharana, S.; Lemaitre, R.; Pozniakovsky, A.; Drechsel, D.; et al. A Molecular Grammar Governing the Driving Forces for Phase Separation of Prion-like RNA Binding Proteins. Cell 2018, 174, 688–699.e16. [Google Scholar] [CrossRef]
- Choi, J.-M.; Dar, F.; Pappu, R.V. LASSI: A lattice model for simulating phase transitions of multivalent proteins. PLoS Ccomput. Biol. 2019, 15, e1007028. [Google Scholar] [CrossRef]
- Milkovic, N.M.; Mittag, T. Determination of Protein Phase Diagrams by Centrifugation. Methods Mol. Biol. 2020, 2141, 685–702. [Google Scholar] [PubMed]
- Peran, I.; Martin, E.W.; Mittag, T. Walking Along a Protein Phase Diagram to Determine Coexistence Points by Static Light Scattering. Methods Mol. Biol. 2020, 2141, 715–730. [Google Scholar] [PubMed]
- Farag, M.; Bremer, A.; Borcherds, W.M.; Mittag, T.; Pappu, R.V. Electrostatic interactions contribute to the cooperative interplay in co-phase separation of mixtures of prion-like low complexity domains. 2022; in preparation. [Google Scholar]
- Pak, C.W.; Kosno, M.; Holehouse, A.S.; Padrick, S.B.; Mittal, A.; Ali, R.; Yunus, A.A.; Liu, D.R.; Pappu, R.V.; Rosen, M.K. Sequence Determinants of Intracellular Phase Separation by Complex Coacervation of a Disordered Protein. Mol. Cell 2016, 63, 72–85. [Google Scholar] [CrossRef] [PubMed]
- Martin, E.W.; Thomasen, F.E.; Milkovic, N.M.; Cuneo, M.J.; Grace, C.R.; Nourse, A.; Lindorff-Larsen, K.; Mittag, T. Interplay of folded domains and the disordered low-complexity domain in mediating hnRNPA1 phase separation. Nucleic Acids Res. 2021, 49, 2931–2945. [Google Scholar] [CrossRef]
- Boeynaems, S.; Holehouse, A.S.; Weinhardt, V.; Kovacs, D.; Van Lindt, J.; Larabell, C.; Van Den Bosch, L.; Das, R.; Tompa, P.S.; Pappu, R.V.; et al. Spontaneous driving forces give rise to protein-RNA condensates with coexisting phases and complex material properties. Proc. Natl. Acad. Sci. USA 2019, 116, 7889–7898. [Google Scholar] [CrossRef]
- Kar, M.; Dar, F.; Welsh, T.J.; Vogel, L.T.; Kuhnemuth, R.; Majumdar, A.; Krainer, G.; Franzmann, T.M.; Alberti, S.; Seidel, C.A.M.; et al. Phase-separating RNA-binding proteins form heterogeneous distributions of clusters in subsaturated solutions. Proc. Natl. Acad. Sci. USA 2022, 119, e2202222119. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bremer, A.; Posey, A.E.; Borgia, M.B.; Borcherds, W.M.; Farag, M.; Pappu, R.V.; Mittag, T. Quantifying Coexistence Concentrations in Multi-Component Phase-Separating Systems Using Analytical HPLC. Biomolecules 2022, 12, 1480. https://doi.org/10.3390/biom12101480
Bremer A, Posey AE, Borgia MB, Borcherds WM, Farag M, Pappu RV, Mittag T. Quantifying Coexistence Concentrations in Multi-Component Phase-Separating Systems Using Analytical HPLC. Biomolecules. 2022; 12(10):1480. https://doi.org/10.3390/biom12101480
Chicago/Turabian StyleBremer, Anne, Ammon E. Posey, Madeleine B. Borgia, Wade M. Borcherds, Mina Farag, Rohit V. Pappu, and Tanja Mittag. 2022. "Quantifying Coexistence Concentrations in Multi-Component Phase-Separating Systems Using Analytical HPLC" Biomolecules 12, no. 10: 1480. https://doi.org/10.3390/biom12101480
APA StyleBremer, A., Posey, A. E., Borgia, M. B., Borcherds, W. M., Farag, M., Pappu, R. V., & Mittag, T. (2022). Quantifying Coexistence Concentrations in Multi-Component Phase-Separating Systems Using Analytical HPLC. Biomolecules, 12(10), 1480. https://doi.org/10.3390/biom12101480