
Effects of Diethylstilbestrol on Zebrafish Gonad Development and Endocrine Disruption Mechanism


Abstract
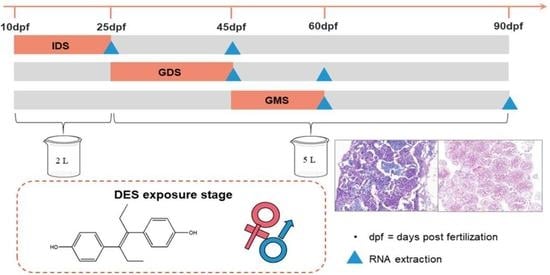
1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Chemicals
2.1.2. Zebrafish Maintenance
2.2. Methods
2.2.1. Eighty-Day Exposure Experiments
2.2.2. Observation of Gonadal Tissue
2.2.3. Analysis of Genes and Proteins
2.2.4. Statistical Analysis
3. Results
3.1. Morphology Parameters
3.1.1. 25,45 and 60 dpf Body Length
3.1.2. Body Length, Body Weight, and K at 90 dpf
3.1.3. GSI
3.1.4. Sex Ratio
3.2. Gonad Tissues Observation
3.3. Gene Expression Changes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Appendix A

References
- Hayes, T.; Haston, K.; Tsui, M.; Hoang, A.; Haeffele, C.; Vonk, A. Herbicides: Feminization of Male Frogs in the Wild. Nature 2002, 419, 895–896. [Google Scholar] [CrossRef] [PubMed]
- Hunt, P.A.; Koehler, K.E.; Susiarjo, M.; Hodges, C.A.; Ilagan, A.; Voigt, R.C.; Thomas, S.; Thomas, B.F.; Hassold, T.J. Bisphenol A Exposure Causes Meiotic Aneuploidy in the Female Mouse. Curr. Biol. 2003, 13, 546–553. [Google Scholar] [CrossRef]
- Witorsch, R.J. Low-Dose in Utero Effects of Xenoestrogens in Mice and Their Relevance to Humans: An Analytical Review of the Literature. Food Chem. Toxicol. 2002, 40, 905–912. [Google Scholar] [CrossRef]
- Herbst, A.L.; Ulfelder, H.; Poskanzer, D.C. Adenocarcinoma of the Vagina: Association of Maternal Stilbestrol Therapy with Tumor Appearance in Young Women. N. Engl. J. Med. 1971, 284, 878–881. [Google Scholar] [CrossRef]
- Adedeji, O.B.; Durhan, E.J.; Garcia-Reyero, N.; Kahl, M.D.; Jensen, K.M.; LaLone, C.A.; Makynen, E.A.; Perkins, E.J.; Thomas, L.; Villeneuve, D.L.; et al. Short-Term Study Investigating the Estrogenic Potency of Diethylstilbesterol in the Fathead Minnow (Pimephales Promelas). Environ. Sci. Technol. 2012, 46, 7826–7835. [Google Scholar] [CrossRef]
- Chen, T.-S.; Chen, T.-C.; Yeh, K.-J.C.; Chao, H.-R.; Liaw, E.-T.; Hsieh, C.-Y.; Chen, K.-C.; Hsieh, L.-T.; Yeh, Y.-L. High Estrogen Concentrations in Receiving River Discharge from a Concentrated Livestock Feedlot. Sci. Total Environ. 2010, 408, 3223–3230. [Google Scholar] [CrossRef]
- Potts, A.C.; Phelps, R.P. Use of Diethylstilbestrol and Ethynylestradiol to Feminize Nile Tilapia Oreochromis niloticus (L.) in an Outdoor Environment. J. Appl. Ichthyol. 1995, 11, 111–117. [Google Scholar] [CrossRef]
- Lei, B.; Peng, W.; Li, W.; Yu, Y.; Xu, J.; Wang, Y. Diethylstilbestrol at Environmental Levels Affects the Development of Early Life Stage and Target Gene Expression in Japanese Medaka (Oryzias latipes). Ecotoxicology 2016, 25, 563–573. [Google Scholar] [CrossRef]
- Lei, B.; Huang, S.; Zhou, Y.; Wang, D.; Wang, Z. Levels of Six Estrogens in Water and Sediment from Three Rivers in Tianjin Area, China. Chemosphere 2009, 76, 36–42. [Google Scholar] [CrossRef]
- Solé, M.; López de Alda, M.J.; Castillo, M.; Porte, C.; Ladegaard-Pedersen, K.; Barceló, D. Estrogenicity Determination in Sewage Treatment Plants and Surface Waters from the Catalonian Area (NE Spain). Environ. Sci. Technol. 2000, 34, 5076–5083. [Google Scholar] [CrossRef]
- He, Y.; Chen, W.; Zheng, X.; Wang, X.; Huang, X. Fate and Removal of Typical Pharmaceuticals and Personal Care Products by Three Different Treatment Processes. Sci. Total Environ. 2013, 447, 248–254. [Google Scholar] [CrossRef]
- Rahman, M.M.; Wie, J.; Cho, J.-H.; Tae, H.-J.; Ahn, D.; Lee, S.W.; Kim, I.-S.; Park, B.-Y. Diethylstilbestrol Induces Morphological Changes in the Spermatogonia, Sertoli Cells and Leydig Cells of Adult Rat. Res. Vet. Sci. 2019, 124, 433–438. [Google Scholar] [CrossRef]
- Goyal, H.O.; Robateau, A.; Braden, T.D.; Williams, C.S.; Srivastava, K.K.; Ali, K. Neonatal Estrogen Exposure of Male Rats Alters Reproductive Functions at Adulthood1. Biol. Reprod. 2003, 68, 2081–2091. [Google Scholar] [CrossRef]
- Hosoi, I.; Toyama, Y.; Maekawa, M.; Ito, H.; Yuasa, S. Development of the Blood-Testis Barrier in the Mouse Is Delayed by Neonatally Administered Diethylstilbestrol but Not by Beta-Estradiol 3-Benzoate. Andrologia 2002, 34, 255–262. [Google Scholar] [CrossRef]
- Odum, J. Comparison of the Developmental and Reproductive Toxicity of Diethylstilbestrol Administered to Rats in Utero, Lactationally, Preweaning, or Postweaning. Toxicol. Sci. 2002, 68, 147–163. [Google Scholar] [CrossRef][Green Version]
- Shen, Y.; Li, Y.; Zhu, M.; Li, J.; Qin, Z. Transcriptional Changes Caused by Estrogenic Endocrine Disrupting Chemicals in Gonad-Mesonephros Complexes of Genetic Male Xenopus Laevis: Multiple Biomarkers for Early Detection of Testis Differentiation Disruption. Sci. Total Environ. 2020, 726, 138522. [Google Scholar] [CrossRef]
- Sridevi, P.; Chaitanya, R.K.; Prathibha, Y.; Balakrishna, S.L.; Dutta-Gupta, A.; Senthilokumaran, B. Early Exposure of 17α-Ethynylestradiol and Diethylstilbestrol Induces Morphological Changes and Alters Ovarian Steroidogenic Pathway Enzyme Gene Expression in Catfish, Clarias gariepinus: EE 2 and Des Affect Ovarian Steroidogenesis. Environ. Toxicol. 2015, 30, 439–451. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.; Chen, Q.; Liu, Z. Diethylstilbestrol Impaired Oogenesis of Yellow Catfish Juveniles through Disrupting Hypothalamic-Pituitary-Gonadal Axis and Germ Cell Development. J. Appl. Toxicol. 2018, 38, 308–317. [Google Scholar] [CrossRef]
- Yin, P.; Li, Y.-W.; Chen, Q.-L.; Liu, Z.-H. Diethylstilbestrol, Flutamide and Their Combination Impaired the Spermatogenesis of Male Adult Zebrafish through Disrupting HPG Axis, Meiosis and Apoptosis. Aquat. Toxicol. 2017, 185, 129–137. [Google Scholar] [CrossRef]
- Dai, Y.-J.; Jia, Y.-F.; Chen, N.; Bian, W.-P.; Li, Q.-K.; Ma, Y.-B.; Chen, Y.-L.; Pei, D.-S. Zebrafish as a Model System to Study Toxicology: Zebrafish Toxicology Monitoring. Environ. Toxicol. Chem. 2014, 33, 11–17. [Google Scholar] [CrossRef]
- Santos, D.; Luzio, A.; Coimbra, A.M. Zebrafish Sex Differentiation and Gonad Development: A Review on the Impact of Environmental Factors. Aquat. Toxicol. 2017, 191, 141–163. [Google Scholar] [CrossRef]
- Maack, G.; Segner, H. Morphological Development of the Gonads in Zebrafish: Zebrafish. J. Fish Biol. 2003, 62, 895–906. [Google Scholar] [CrossRef]
- Wang, X.G.; Bartfai, R.; Sleptsova-Freidrich, I.; Orban, L. The Timing and Extent of “juvenile Ovary” Phase Are Highly Variable during Zebrafish Testis Differentiation. J. Fish Biol. 2007, 70, 33–44. [Google Scholar] [CrossRef]
- Chen, W.; Ge, W. Gonad Differentiation and Puberty Onset in the Zebrafish: Evidence for the Dependence of Puberty Onset on Body Growth but Not Age in Females: G Rowth and P Uberty in the Z Ebrafish. Mol. Reprod. Dev. 2013, 80, 384–392. [Google Scholar] [CrossRef]
- Ma, Z.; Tang, S.; Su, G.; Miao, Y.; Liu, H.; Xie, Y. Effects of Tris (2-Butoxyethyl) Phosphate (TBOEP) on Endocrine Axes during Development of Early Life Stages of Zebrafish (Danio Rerio). Chemosphere 2016, 144, 1920–1927. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(T)(-Delta Delta C) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Kennedy, G.C.; Mitra, J. Body Weight and Food Intake as Initiating Factors for Puberty in the Rat. J. Physiol. 1963, 166, 408–418. [Google Scholar] [CrossRef]
- Ramaley, J.A.; Phares, C.K. Delay of Puberty Onset in Females Due to Suppression of Growth Hormone. Endocrinology 1980, 106, 1989–1993. [Google Scholar] [CrossRef]
- Taranger, G.L.; Carrillo, M.; Schulz, R.W.; Fontaine, P.; Zanuy, S.; Felip, A.; Weltzien, F.-A.; Dufour, S.; Karlsen, Ø.; Norberg, B.; et al. Control of Puberty in Farmed Fish. Gen. Comp. Endocrinol. 2010, 165, 483–515. [Google Scholar] [CrossRef]
- Hazlerigg, C.R.E.; Tyler, C.R.; Lorenzen, K.; Wheeler, J.R.; Thorbek, P. Population Relevance of Toxicant Mediated Changes in Sex Ratio in Fish: An Assessment Using an Individual-Based Zebrafish (Danio Rerio) Model. Ecol. Model. 2014, 280, 76–88. [Google Scholar] [CrossRef]
- White, J.W.; Cole, B.J.; Cherr, G.N.; Connon, R.E.; Brander, S.M. Scaling Up Endocrine Disruption Effects from Individuals to Populations: Outcomes Depend on How Many Males a Population Needs. Environ. Sci. Technol. 2017, 51, 1802–1810. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, E.J.; Baldwin, L.A. A Quantitatively-based Methodology for the Evaluation of Chemical Hormesis. Hum. Ecol. Risk Assess. Int. J. 1997, 3, 545–554. [Google Scholar] [CrossRef]
- Holbech, H.; Kinnberg, K.; Petersen, G.I.; Jackson, P.; Hylland, K.; Norrgren, L.; Bjerregaard, P. Detection of Endocrine Disrupters: Evaluation of a Fish Sexual Development Test (FSDT). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2006, 144, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Han, J.; Guo, Y.; Lam, P.K.S.; Wu, R.S.S.; Giesy, J.P.; Zhang, X.; Zhou, B. Disruption of Endocrine Function in in Vitro H295R Cell-Based and in in Vivo Assay in Zebrafish by 2,4-Dichlorophenol. Aquat. Toxicol. 2012, 106–107, 173–181. [Google Scholar] [CrossRef]
- Dang, Z.; Kienzler, A. Changes in Fish Sex Ratio as a Basis for Regulating Endocrine Disruptors. Environ. Int. 2019, 130, 104928. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Gene ID |
|---|---|---|---|
| ddx4 | ctcaaccacaagcatcca | agcgtccagttcttccta | ENSDARG00000014373 |
| casp9 | acgagatggacgctattc | ttgagtaggacaccaggat | ENSDARG00000004325 |
| caspb | ggacagagacgaggagaa | agttcacagagttcagatgg | ENSDARG00000052039 |
| casp3a | gcggatacggagactact | gattgaggcttggcatca | ENSDARG00000017905 |
| cyp26a1 | tgttctccttgccaatcg | tcacttcttctgctgttctc | ENSDARG00000033999 |
| aldh1a2 | tcttcaataacggtcaatgc | ttcctcctctgtgctctc | ENSDARG00000053493 |
| sox9a | gacaccagcagacaacaa | gcatcagacagacacttct | ENSDARG00000003293 |
| ghra | ccttcttcacaaccattctg | ctccaccacttctgattcc | ENSDARG00000054771 |
| gh1, | cagttggtggtggttagtt | gcgttcctcaggcataag | ENSDARG00000038185 |
| vtg5 | agctaatgctctgcccgtta | gttcagcctcaaacagcaca | ENSDARG00000092126 |
| vtg4 | ctacaaggtggaggctctgc | ggaggacaaatcaccagcat | ENSDARG00000078429 |
| vtg2 | tactttgggcactgatgcaa | agacttcgtgaagcccaaga | ENSDARG00000055809 |
| vtg1 | ctgcgtgaagttgtcatgct | gaccagcattgcccataact | ENSDARG00000092233 |
| ar | gtctattaagagccgcctat | caccgcaacaagttcatc | ENSDARG00000067976 |
| esr2b | ttgtgttctccagcatgagc | ccacatatggggaaggaatg | ENSDARG00000034181 |
| esr2a | agcattagccttgttagca | atgttgtcacggatgtcat | ENSDARG00000016454 |
| esr1 | ggtccagtgtggtgtcctct | cacacgaccagactccgtaa | ENSDARG00000004111 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Xie, X.; Liu, H. Effects of Diethylstilbestrol on Zebrafish Gonad Development and Endocrine Disruption Mechanism. Biomolecules 2021, 11, 941. https://doi.org/10.3390/biom11070941
Liu X, Xie X, Liu H. Effects of Diethylstilbestrol on Zebrafish Gonad Development and Endocrine Disruption Mechanism. Biomolecules. 2021; 11(7):941. https://doi.org/10.3390/biom11070941
Chicago/Turabian StyleLiu, Xuan, Xianyi Xie, and Hongling Liu. 2021. "Effects of Diethylstilbestrol on Zebrafish Gonad Development and Endocrine Disruption Mechanism" Biomolecules 11, no. 7: 941. https://doi.org/10.3390/biom11070941
APA StyleLiu, X., Xie, X., & Liu, H. (2021). Effects of Diethylstilbestrol on Zebrafish Gonad Development and Endocrine Disruption Mechanism. Biomolecules, 11(7), 941. https://doi.org/10.3390/biom11070941