
Profiles of Secondary Metabolites (Phenolic Acids, Carotenoids, Anthocyanins, and Galantamine) and Primary Metabolites (Carbohydrates, Amino Acids, and Organic Acids) during Flower Development in Lycoris radiata

 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Carotenoid HPLC Analysis
2.3. Phenolic Acid HPLC Analysis
2.4. Galantamine HPLC Analysis
2.5. Anthocyanin HPLC Analysis
2.6. QRT-PCR Analysis
2.7. GC-TOFMS Analysis for Carbohydrate, Amino Acid, Organic Acid, and Phenolic Acid
2.8. Statistical Analysis
3. Results and Discussion
3.1. Carotenoids in Flowers at Different Flower Developmental Stages
3.2. Galantamine in Flowers at the Different Flower Developmental Stages
3.3. Phenolic Acids and Anthocyanins in Flowers at Different Flower Developmental Stages
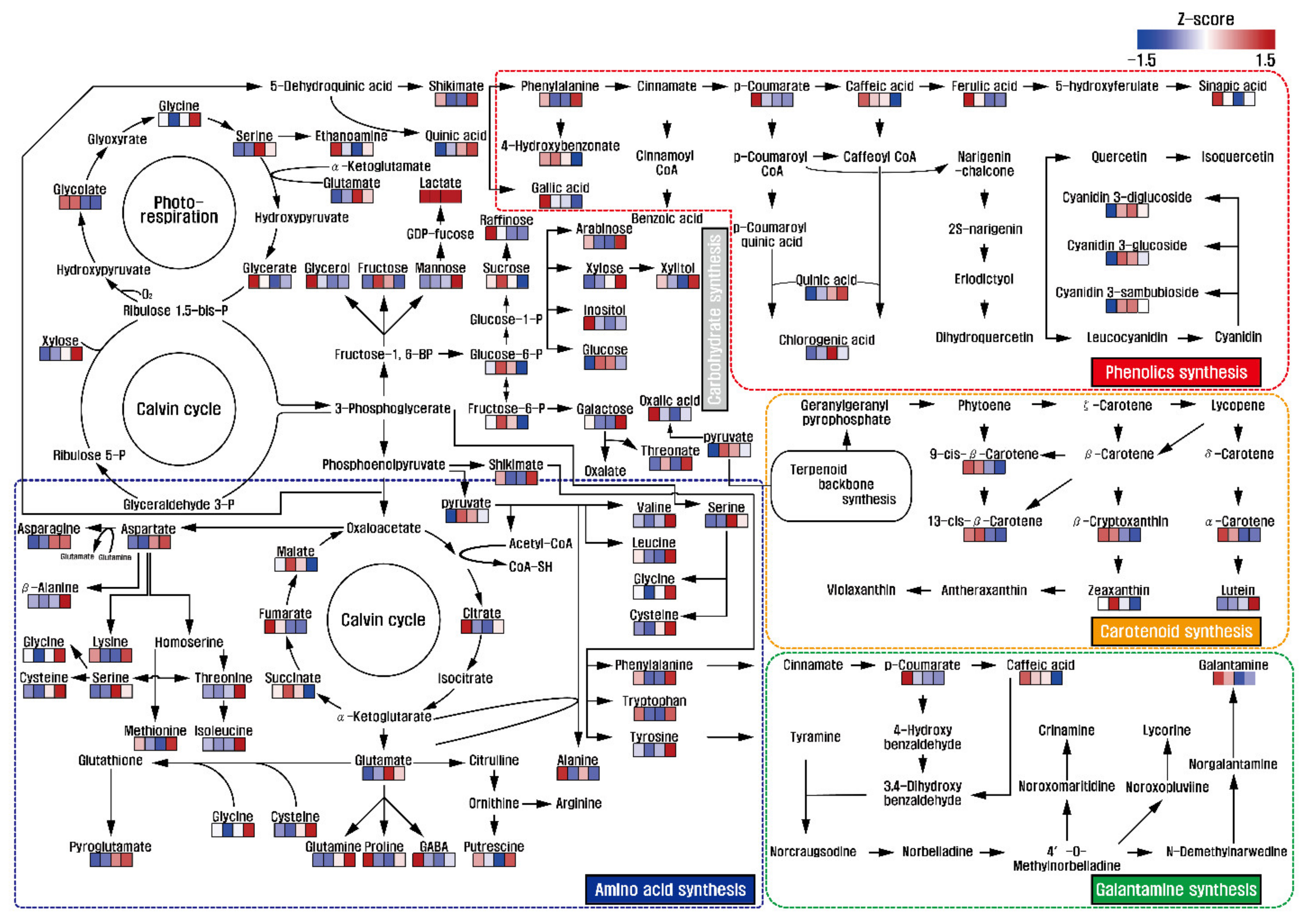
3.4. Metabolic Profiling and Multivariate Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Hayashi, A.; Saito, T.; Mukai, Y.; Kurita, S.; Hori, T.A. Genetic variations in Lycoris radiata var. radiata in Japan. Genes Genet. Syst. 2005, 80, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Yeo, H.J.; Park, Y.E.; Baek, S.A.; Kim, J.K.; Park, S.U. Transcriptome analysis and metabolic profiling of Lycoris radiata. Biology 2019, 8, 63. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.M.; Huang, X.Y.; Cui, M.R.; Zhang, X.D.; Chen, Z.; Yang, B.S.; Zhao, X.K. Amaryllidaceae alkaloids from the bulbs of Lycoris radiata with cytotoxic and anti-inflammatory activities. Fitoterapia 2015, 101, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Sener, B.; Orhan, I.; Satayavivad, J. Antimalarial activity screening of some alkaloids and the plant extracts from Amaryllidaceae. Phytother. Res. 2003, 17, 1220–1223. [Google Scholar] [CrossRef] [PubMed]
- Hanes, W.M.; Olofsson, P.S.; Kwan, K.; Hudson, L.K.; Chavan, S.S.; Pavlov, V.A.; Tracey, K.J. Galantamine attenuates type 1 diabetes and inhibits anti-insulin antibodies in nonobese diabetic mice. Mol. Med. 2015, 21, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Gantait, S.S.; Pal, P. Anthocyanin content of spray Chrysanthemum cultivars under polyhouse and open field conditions. IJNPR 2010, 1, 236–242. [Google Scholar]
- Huang, H.; Hu, K.; Han, K.T.; Xiang, Q.Y.; Dai, S.L. Flower colour modification of Chrysanthemum by suppression of F3′H and overexpression of the exogenous Senecio cruentus F3′5′H gene. PLoS ONE 2013, 8, e74395. [Google Scholar]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef]
- Park, C.H.; Park, S.Y.; Park, Y.J.; Kim, J.K.; Park, S.U. Metabolite Profiling and Comparative Analysis of Secondary Metabolites in Chinese Cabbage, Radish, and Hybrid ×Brassicoraphanus. J. Agric. Food Chem. 2020, 68, 13711–13719. [Google Scholar] [CrossRef]
- Sathasivam, R.; Radhakrishnan, R.; Kim, J.K.; Park, S.U. An update on biosynthesis and regulation of carotenoids in plants. S. Afr. J. Bot. 2020, 1–13. [Google Scholar] [CrossRef]
- Domonkos, I.; Kis, M.; Gombos, Z.; Ughy, B. Carotenoids, versatile components of oxygenic photosynthesis. Prog. Lipid Res. 2013, 52, 539–561. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.G.; Zhang, C.; Dong, B.; Fu, J.X.; Hu, S.Q.; Zhao, H.B. Carotenoid accumulation and its contribution to flower coloration of Osmanthus fragrans. Front. Plant Sci. 2018, 9, 1499. [Google Scholar] [CrossRef] [PubMed]
- Ohmiya, A. Qualitative and quantitative control of carotenoid accumulation in flower petals. Sci. Hortic. 2013, 163, 10–19. [Google Scholar] [CrossRef]
- Yuan, H.; Zhang, J.X.; Nageswaran, D.; Li, L. Carotenoid metabolism and regulation in horticultural crops. Hortic. Res. 2015, 2, 15036. [Google Scholar] [CrossRef] [PubMed]
- Castaneda-Ovando, A.; Pacheco-Hernandez, M.D.; Paez-Hernandez, M.E.; Rodriguez, J.A.; Galan-Vidal, C.A. Chemical studies of anthocyanins: A review. Food Chem. 2009, 113, 859–871. [Google Scholar] [CrossRef]
- Clifford, M.N. Anthocyanins—Nature, occurrence and dietary burden. J. Sci. Food Agric. 2000, 80, 1063–1072. [Google Scholar] [CrossRef]
- Park, C.H.; Park, S.Y.; Lee, S.Y.; Kim, J.K.; Park, S.U. Analysis of metabolites in white lowers of Magnolia denudata Desr. and violet flowers of Magnolia liliiflora Desr. Molecules 2018, 23, 1558. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.H.; Chen, H.H. Orchid Biotechnology II, 1st ed.; World Scientific Publishing Co.: Singapore, 2011. [Google Scholar]
- Chun, J.H.; Jang, I.H.; Arasu, M.V.; Al-Dhabi, N.A.; Duraipandiyan, V.; Lee, D.H.; Lee, S.; Kim, S.J. Isolation and identification of alkaloids and anthocyanins from flower and bulb of Lycoris radiata using HPLC and LC-ESI-MS J. Agric. Chem. Environ. 2013, 2, 22–26. [Google Scholar] [CrossRef]
- Ma, N.; Ma, C.; Liu, Y.; Shahid, M.O.; Wang, C.P.; Gao, J.P. Petal senescence: A hormone view. J. Exp. Bot. 2018, 69, 719–732. [Google Scholar] [CrossRef]
- Arrom, L.; Munne-Bosch, S. Sucrose accelerates flower opening and delays senescence through a hormonal effect in cut lily flowers. Plant Sci. 2012, 188, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Bong, S.J.; Lim, C.J.; Kim, J.K.; Park, S.U. Transcriptome analysis and metabolic profiling of green and red mizuna (Brassica rapa L. var. japonica). Foods 2020, 9, 1079. [Google Scholar] [CrossRef]
- Park, C.H.; Sathasivam, R.; Nguyen, B.V.; Baek, S.A.; Yeo, H.J.; Park, Y.E.; Kim, H.H.; Kim, J.K. Metabolic Profiling of Primary Metabolites and Galantamine Biosynthesis in Wounded Lycoris radiata Callus. Plants 2020, 9, 1616. [Google Scholar] [CrossRef]
- Zhao, S.C.; Park, C.H.; Yang, J.L.; Yeo, H.J.; Kim, T.J.; Kim, J.K.; Park, S.U. Molecular characterization of anthocyanin and betulinic acid biosynthesis in red and white mulberry fruits using high-throughput sequencing. Food Chem. 2019, 279, 364–372. [Google Scholar] [CrossRef]
- Yamamizo, C.; Kishimoto, S.; Ohmiya, A. Carotenoid composition and carotenogenic gene expression during Ipomoea petal development. J. Exp. Bot. 2010, 61, 709–719. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, M.; Kishimoto, S.; Nakayama, M. Carotenoid composition and changes in expression of carotenoid biosynthetic genes in tepals of Asiatic hybrid lily. Plant Breed. 2010, 129, 100–107. [Google Scholar] [CrossRef]
- Wan, H.H.; Yu, C.; Han, Y.; Guo, X.L.; Luo, L.; Pan, H.T.; Zheng, T.C.; Wang, J.; Cheng, T.R.; Zhang, Q.X. Determination of flavonoids and carotenoids and their contributions to various colors of rose cultivars (Rosa spp.). Front. Plant Sci. 2019, 10, 123. [Google Scholar] [CrossRef] [PubMed]
- Schmitzer, V.; Veberic, R.; Osterc, G.; Stampar, F. Changes in the phenolic concentration during flower development of rose ‘KORcrisett’. J. Am. Soc. Hortic. Sci. 2009, 134, 491–496. [Google Scholar] [CrossRef]
- Schmitzer, V.; Veberic, R.; Osterc, G.; Stampar, F. Color and phenolic content changes during flower development in groundcover rose. J. Am. Soc. Hortic. Sci. 2010, 135, 195–202. [Google Scholar] [CrossRef]
- Nisar, S.; Tahir, I.; Ahmad, S.S.; Dar, R.A. Physiological and biochemical aspects of flower development and senescence in Nicotiana plumbaginifolia Viv. Folia Hortic. 2017, 29, 25–31. [Google Scholar] [CrossRef][Green Version]
- Ferri, D.; Ubaldi, C.; Marcozzi, G.; Fasciani, P.; Bacchetta, L.; Pace, L. Chemical characterization of Narcissus poeticus from sirente—Velino (Apennines—Italy): Galantamine accumulation and distribution of allergenic compounds in the flower. Nat. Prod. Commun. 2017, 12, 15–18. [Google Scholar] [CrossRef]
- Lopez, S.; Bastida, J.; Viladomat, F.; Codina, C. Galanthamine pattern in Narcissus confusus plants. Planta Med. 2003, 69, 1166–1168. [Google Scholar]
- Niu, X.X.; Chen, X.W.; Se, H.; Eneji, A.E.; Guo, Y.H.; Dong, X.H. Changes of secondary metabolites and trace elements in Gentiana macrophylla flowers: A potential medicinal plant part. Chinese Herb. Med. 2014, 6, 145–151. [Google Scholar] [CrossRef]
- Gul, F.; Tahir, I.; Shahri, W. Flower development and senescence in Narcissus tazetta ‘Kashmir Local’. Folia Hortic. 2015, 27, 115–121. [Google Scholar] [CrossRef]
- Waseem, S.; Inayatullah, T. Physiological and biochemical changes associated with flower development and senescence in Consolida ajacis Nieuwl cv. Violet blue. Front. Agric. China 2011, 5, 201–208. [Google Scholar] [CrossRef]
- Shahri, W.; Tahir, I. Flower development and senescence In Ranunculus asiaticus L. J. Fruit Ornam. Plant Res. 2011, 19, 123–131. [Google Scholar]
- El Tahchy, A.; Bordage, S.; Ptak, A.; Dupire, F.; Barre, E.; Guillou, C.; Henry, M.; Chapleur, Y.; Laurain-Mattar, D. Effects of sucrose and plant growth regulators on acetylcholinesterase inhibitory activity of alkaloids accumulated in shoot cultures of Amaryllidaceae. Plant Cell Tiss. Org. 2011, 106, 381–390. [Google Scholar] [CrossRef]
- Georgiev, V.; Berkov, S.; Georgiev, M.; Burrus, M.; Codina, C.; Bastida, J.; Ilieva, M.; Pavlov, A. Optimized nutrient medium for galanthamine production in Leucojum aestivum L. In Vitro shoot system. Z. Naturforsch. C. 2009, 64, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Selles, M.; Bergonon, S.; Viladomat, F.; Bastida, J.; Codina, C. Effect of sucrose on growth and galanthamine production in shoot-clump cultures of Narcissus confusus in liquid-shake medium. Plant Cell Tiss. Org. 1997, 49, 129–136. [Google Scholar] [CrossRef]
- Guo, S.; Liu, J.; Zheng, Y.; Huang, M.; Zhang, H.; Gong, G.; He, H.; Ren, Y.; Zhong, S.; Fei, Z.; et al. Characterization of transcriptome dynamics during watermelon fruit development: Sequencing, assembly, annotation and gene expression profiles. BMC Genom. 2011, 12, 454. [Google Scholar] [CrossRef] [PubMed]
- Telef, N.; Stammitti-Bert, L.; Mortain-Bertrand, A.; Maucourt, M.; Carde, J.P.; Rolin, D.; Gallusci, P. Sucrose deficiency delays lycopene accumulation in tomato fruit pericarp discs. Plant Mol. Biol. 2006, 62, 453–469. [Google Scholar] [CrossRef]
- Zhang, L.C.; Ma, G.; Kato, M.; Yamawaki, K.; Takagi, T.; Kiriiwa, Y.; Ikoma, Y.; Matsumoto, H.; Yoshioka, T.; Nesumi, H. Regulation of carotenoid accumulation and the expression of carotenoid metabolic genes in citrus juice sacs In Vitro. J. Exp. Bot. 2012, 63, 871–886. [Google Scholar] [CrossRef]
- Yun, J.W.; Kim, J.H.; Yoo, Y.J. Optimizations of carotenoid biosynthesis by controlling sucrose concentration. Biotechnol. Lett. 1990, 12, 905–910. [Google Scholar] [CrossRef]
- Shin, D.H.; Choi, M.G.; Lee, H.K.; Cho, M.; Choi, S.B.; Choi, G.; Park, Y.I. Calcium dependent sucrose uptake links sugar signaling to anthocyanin biosynthesis in Arabidopsis. Biochem. Bioph. Res. Commun. 2013, 430, 634–639. [Google Scholar] [CrossRef] [PubMed]
- Zakhleniuk, O.V.; Raines, C.A.; Lloyd, J.C. pho3: A phosphorus-deficient mutant of Arabidopsis thaliana (L.) Heynh. Planta 2001, 212, 529–534. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Xu, H.; Yeo, H.J.; Park, Y.E.; Hwang, G.S.; Park, N.I.; Park, S.U. Enhancement of the flavone contents of Scutellaria baicalensis hairy roots via metabolic engineering using maize Lc and Arabidopsis PAP1 transcription factors. Metab. Eng. 2021, 64, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Park, Y.E.; Yeo, H.J.; Yoon, J.S.; Park, S.Y.; Kim, J.K.; Park, S.U. Comparative Analysis of Secondary Metabolites and Metabolic Profiling between Diploid and Tetraploid Morus alba L. J. Agric. Food Chem. 2021, 69, 1300–1307. [Google Scholar] [CrossRef] [PubMed]
- Le, B.H.; Cheng, C.; Bui, A.Q.; Wagmaister, J.A.; Henry, K.F.; Pelletier, J.; Kwong, L.; Belmonte, M.; Kirkbride, R.; Horvath, S.; et al. Global analysis of gene activity during Arabidopsis seed development and identification of seed-specific transcription factors. Proc. Natl. Acad. Sci. USA 2010, 107, 8063–8070. [Google Scholar] [CrossRef] [PubMed]
- Gaufichon, L.; Marmagne, A.; Belcram, K.; Yoneyama, T.; Sakakibara, Y.; Hase, T.; Grandjean, O.; Clement, G.; Citerne, S.; Boutet-Mercey, S.; et al. ASN1-encoded asparagine synthetase in floral organs contributes to nitrogen filling in Arabidopsis seeds. Plant J. 2017, 91, 371–393. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; Wang, Y.; Hu, J.; Ding, Z.; Liang, Q.; Zhang, Y.; Wang, H. Mineral and metabolic profiles in tea leaves and flowers during flower development. Plant Physiol. Biochem. 2016, 106, 316–326. [Google Scholar] [CrossRef]
- Jayanthi, D.; Martin, P.A.; Thamizhseran, N. Biochemical changes during flowering of Spathodea campanulata P. Beauv. Pharmacogn. J. 2019, 11, 842–847. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound (µg/g) | Stage 1 | Stage 2 | Stage 3 | Stage 4 |
|---|---|---|---|---|
| Lutein | 16.12 ± 1.50c 1 | 18.05 ± 1.71c | 22.37 ± 3.30b | 47.99 ± 1.00a |
| Zeaxanthin | 7.63 ± 0.26ab | 8.15 ± 0.44a | 7.59 ± 0.26ab | 7.19 ± 0.07b |
| β-Cryptoxanthin | 17.43 ± 0.58a | 16.58 ± 0.44a | 10.93 ± 1.15b | 9.27 ± 0.22c |
| 13Z-β-Carotene | 20.69 ± 1.25a | 22.25 ± 0.54a | 11.43 ± 2.25b | 10.22 ± 0.49b |
| α-Carotene | 2.37 ± 0.18a | 1.97 ± 0.30b | 0.93 ± 0.17c | 1.04 ± 0.05c |
| β-Carotene | 89.15 ± 4.85a | 80.75 ± 2.08a | 42.10 ± 7.36b | 50.31 ± 0.70b |
| 9Z-β-Carotene | 77.60 ± 4.15a | 68.87 ± 2.31b | 28.81 ± 6.26c | 12.26 ± 0.80d |
| Flower Developmental Stages | Galantamine (mg/g DW) |
|---|---|
| Stage 1 | 2.71 ± 0.13a 1 |
| Stage 2 | 2.39 ± 0.17b |
| Stage 3 | 1.65 ± 0.01d |
| Stage 4 | 1.89 ± 0.08c |
| Equipment | Compound | Stage 1 | Stage 2 | Stage 3 | Stage 4 |
|---|---|---|---|---|---|
| HPLC (µg/g) | Gallic acid | 74.51 ± 5.50a 1 | 36.99 ± 4.55b | 34.63 ± 6.74b | 13.38 ± 0.46b |
| 4-Hydroxybenzoic acid | 60.08 ± 4.64ab | 65.40 ± 26.23a | 51.82 ± 34.68ab | 17.86 ± 11.92b | |
| Chlorogenic acid | 83.95 ± 15.74b | 108.48 ± 27.79ab | 222.52 ± 121.59a | 125.98 ± 12.83ab | |
| Caffeic acid | 143.96 ± 17.31a | 110.98 ± 6.95b | 104.00 ± 5.94b | 24.66 ± 0.00c | |
| p-Coumaric acid | 26.24 ± 2.36a | 4.12 ± 2.82b | 1.10 ± 0.81bc | N.D | |
| GC-TOFMS (Intensity/g) | Ferulic acid | 0.06 ± 0.01a | 0.04 ± 0.00b | 0.02 ± 0.00c | 0.02 ± 0.00c |
| Sinapinic acid | 0.08 ± 0.01a | 0.07 ± 0.01ab | 0.05 ± 0.00c | 0.06 ± 0.01bc |
| Compound (mg/g) | Stage 1 | Stage 2 | Stage 3 | Stage 4 |
|---|---|---|---|---|
| Cyanidin 3-diglucoside | 0.04 ± 0.03b 1 | 0.14 ± 0.04a | 0.16 ± 0.06a | 0.12 ± 0.04ab |
| Cyanidin 3-sambubioside | 3.94 ± 2.04c | 11.69 ± 0.22ab | 12.25 ± 0.26a | 9.27 ±1.55b |
| Cyanidin 3-glucoside | 0.04 ± 0.02c | 0.17 ± 0.04a | 0.15 ± 0.03ab | 0.11 ± 0.01b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, C.H.; Yeo, H.J.; Kim, Y.J.; Nguyen, B.V.; Park, Y.E.; Sathasivam, R.; Kim, J.K.; Park, S.U. Profiles of Secondary Metabolites (Phenolic Acids, Carotenoids, Anthocyanins, and Galantamine) and Primary Metabolites (Carbohydrates, Amino Acids, and Organic Acids) during Flower Development in Lycoris radiata. Biomolecules 2021, 11, 248. https://doi.org/10.3390/biom11020248
Park CH, Yeo HJ, Kim YJ, Nguyen BV, Park YE, Sathasivam R, Kim JK, Park SU. Profiles of Secondary Metabolites (Phenolic Acids, Carotenoids, Anthocyanins, and Galantamine) and Primary Metabolites (Carbohydrates, Amino Acids, and Organic Acids) during Flower Development in Lycoris radiata. Biomolecules. 2021; 11(2):248. https://doi.org/10.3390/biom11020248
Chicago/Turabian StylePark, Chang Ha, Hyeon Ji Yeo, Ye Jin Kim, Bao Van Nguyen, Ye Eun Park, Ramaraj Sathasivam, Jae Kwang Kim, and Sang Un Park. 2021. "Profiles of Secondary Metabolites (Phenolic Acids, Carotenoids, Anthocyanins, and Galantamine) and Primary Metabolites (Carbohydrates, Amino Acids, and Organic Acids) during Flower Development in Lycoris radiata" Biomolecules 11, no. 2: 248. https://doi.org/10.3390/biom11020248
APA StylePark, C. H., Yeo, H. J., Kim, Y. J., Nguyen, B. V., Park, Y. E., Sathasivam, R., Kim, J. K., & Park, S. U. (2021). Profiles of Secondary Metabolites (Phenolic Acids, Carotenoids, Anthocyanins, and Galantamine) and Primary Metabolites (Carbohydrates, Amino Acids, and Organic Acids) during Flower Development in Lycoris radiata. Biomolecules, 11(2), 248. https://doi.org/10.3390/biom11020248