
Activation of PKCε-ALDH2 Axis Prevents 4-HNE-Induced Pain in Mice

 ,
,  and
and
Abstract
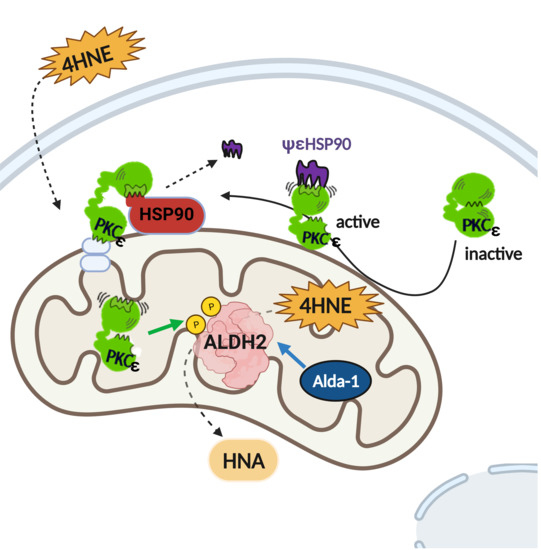
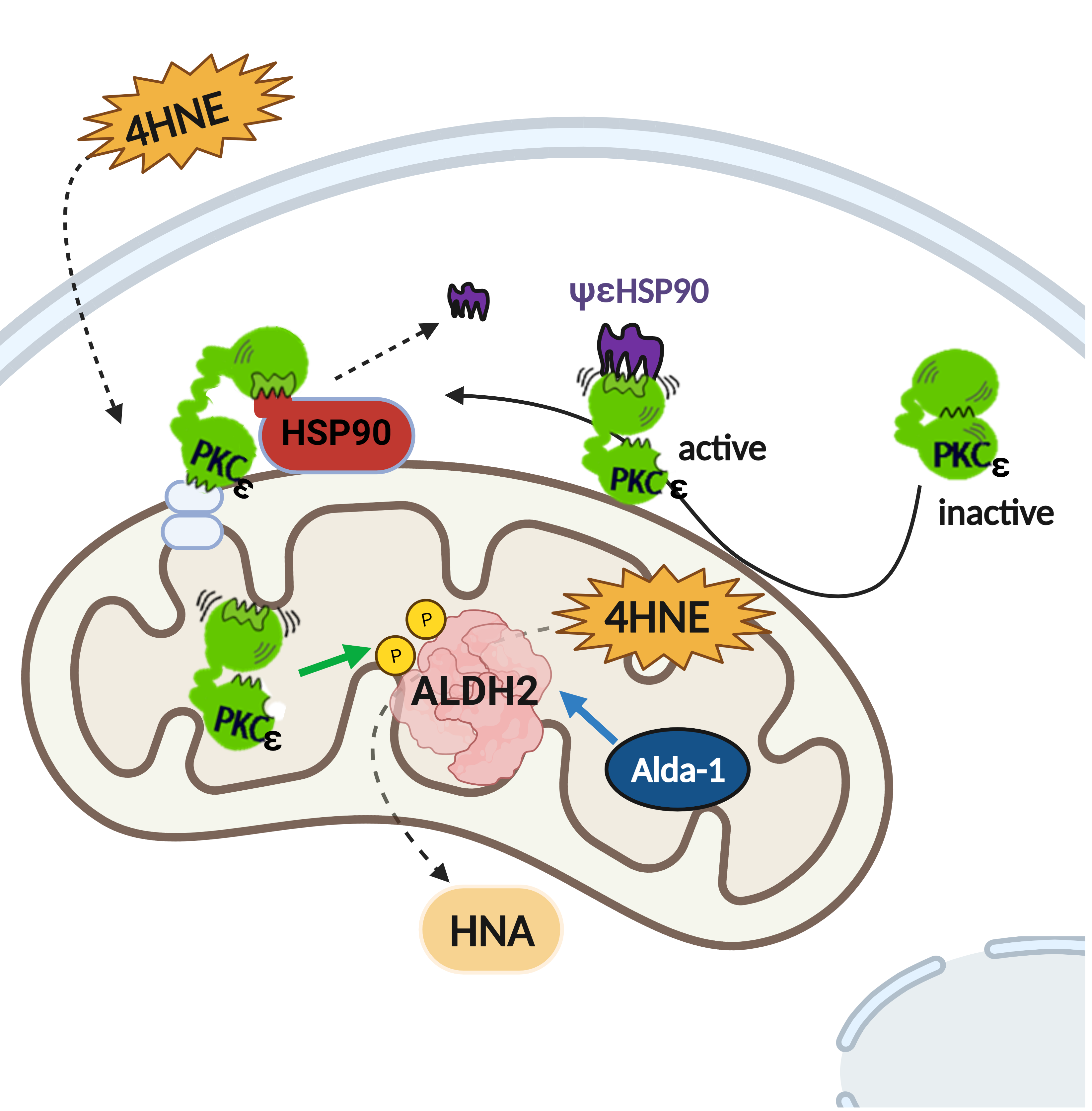{kind=link}
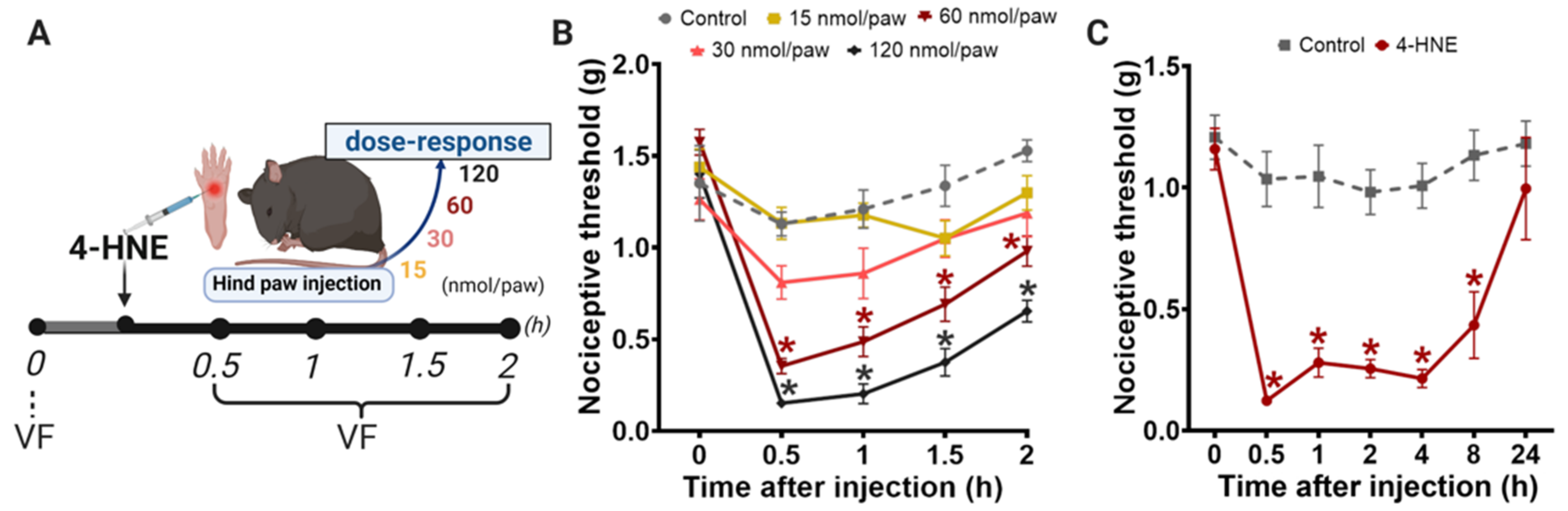{kind=link}
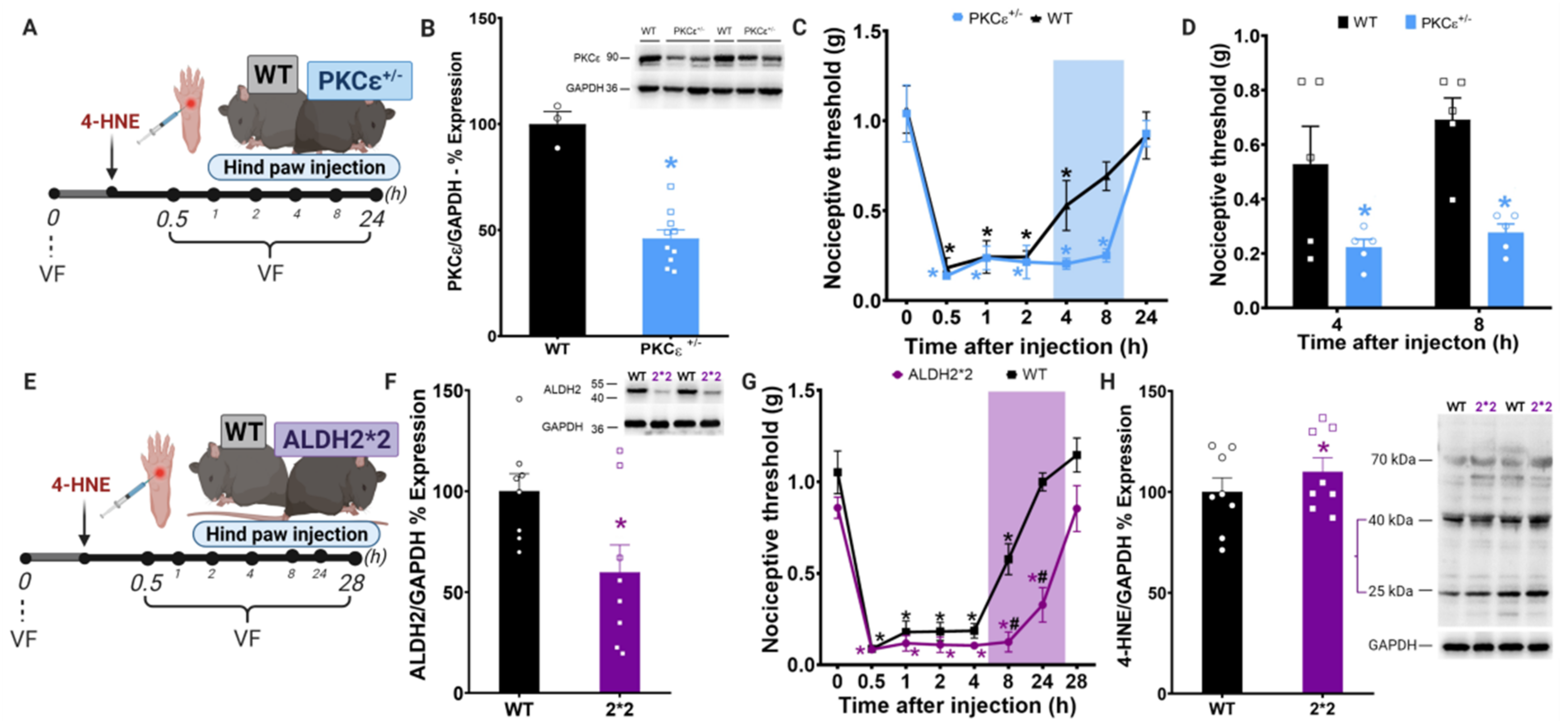{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Reagents
2.3. Drug Administration
2.4. Behavioral Testing
2.5. Western Blot
2.6. Statistical Analysis
3. Results
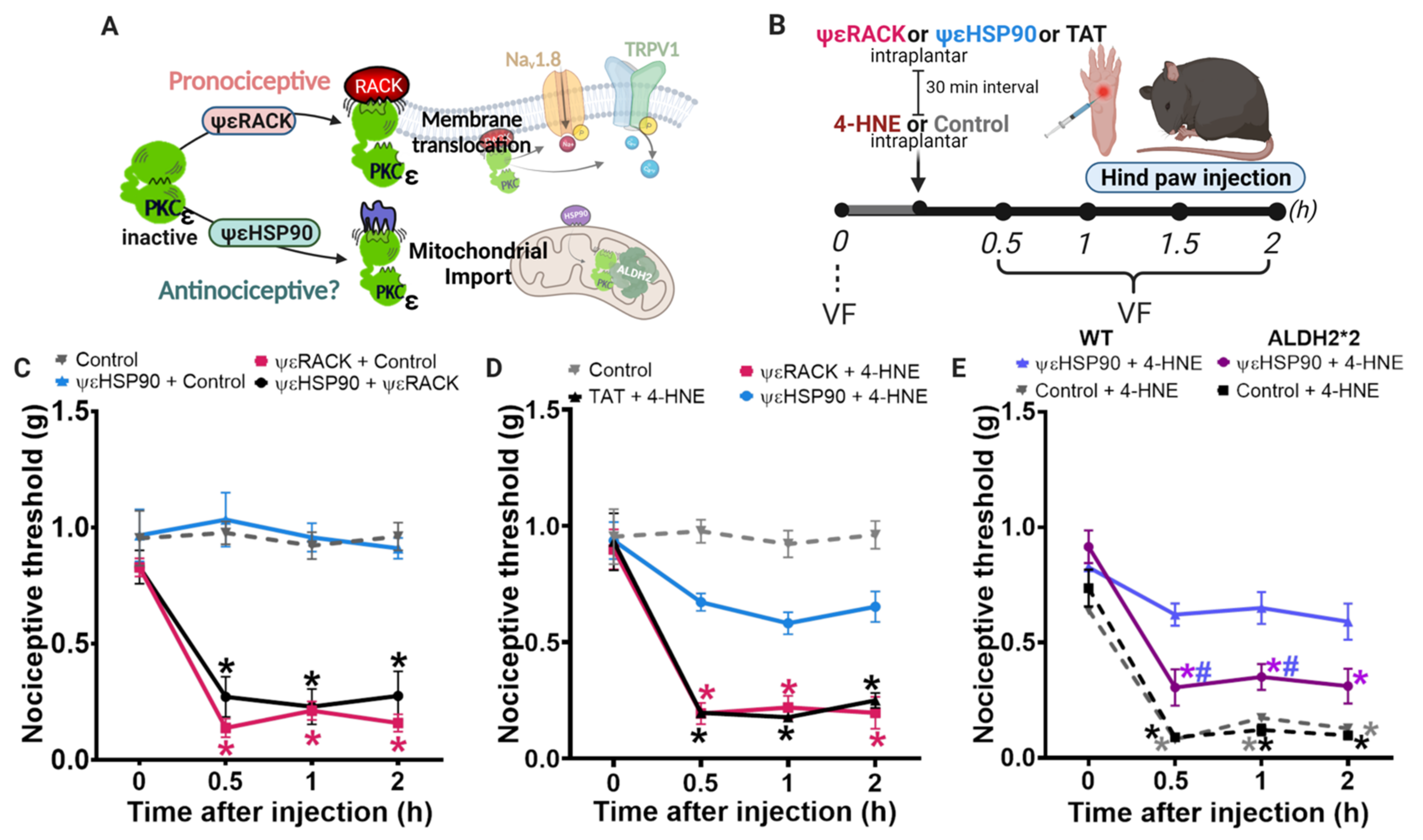
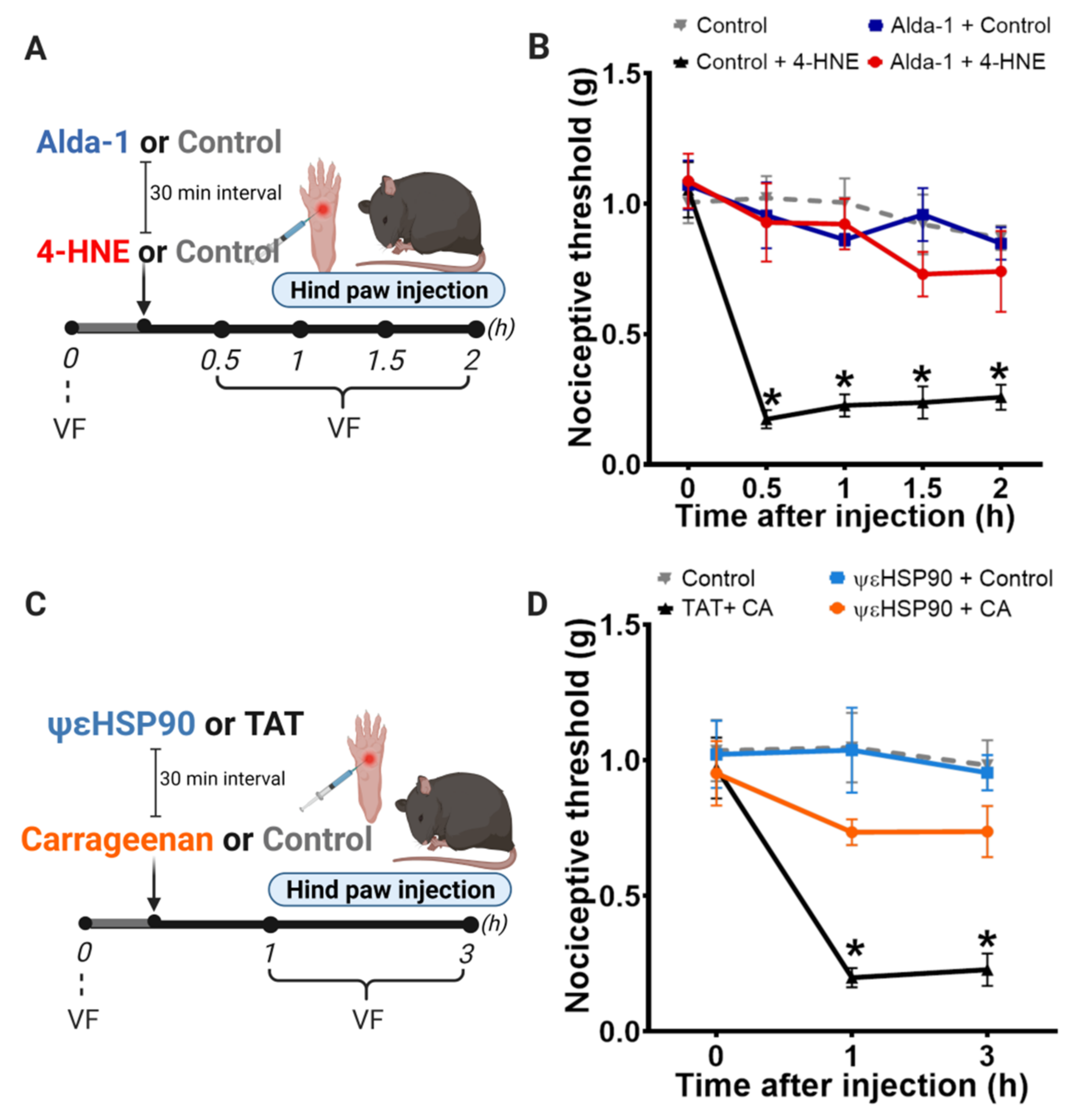
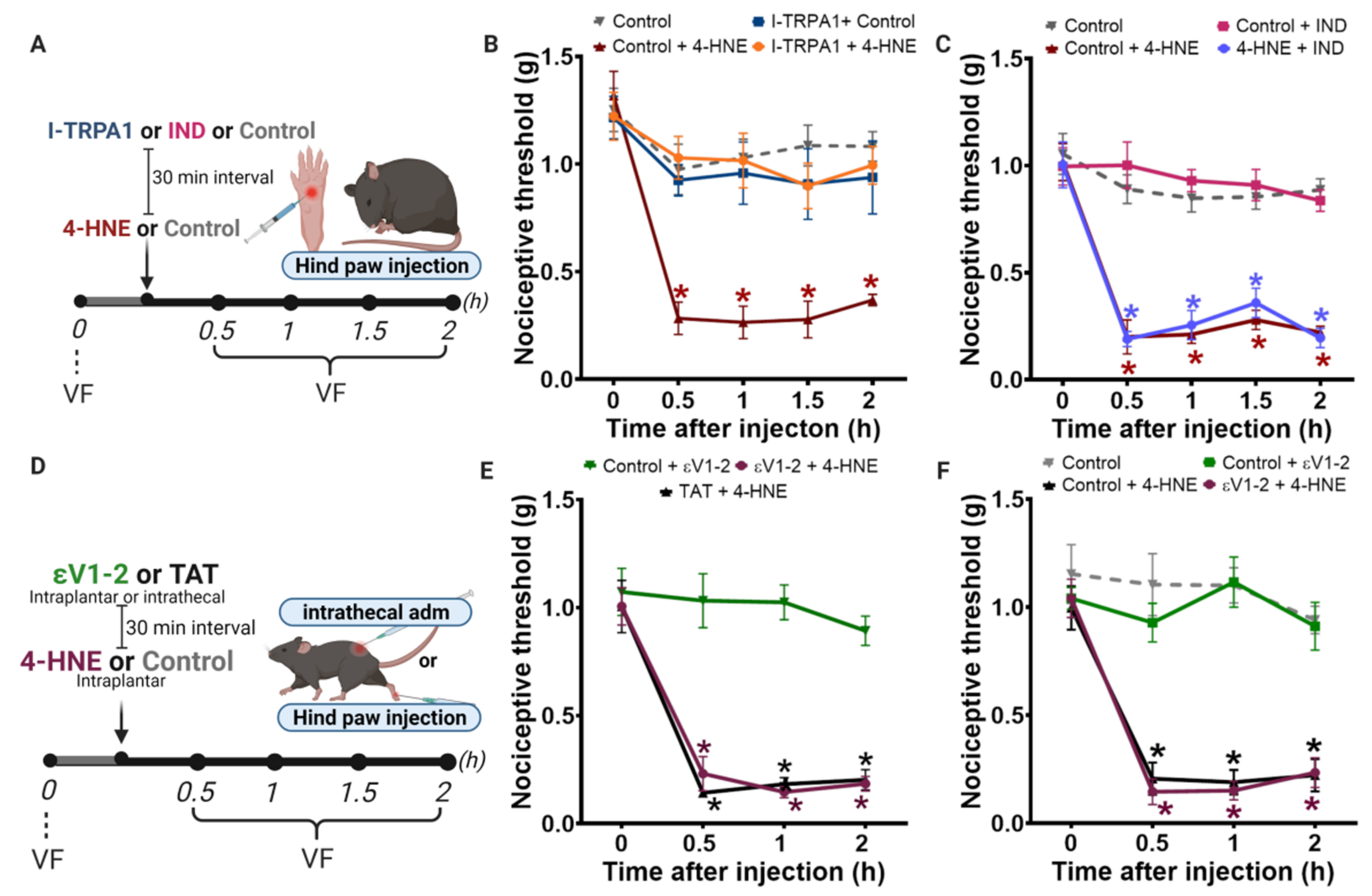
3.1. Disruption of PKCε-ALDH2 Axis Contributes to 4-HNE-Induced Mechanical Hypersensitivity
3.2. Selective Activation of PKCε in Mitochondria Blocks 4-HNE-Induced Hypersensitivity
3.3. Selective Activation of PKCε in Mitochondria Prevents Carrageenan-Induced Hypersensitivity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Goldberg, D.S.; McGee, S.J. Pain as a Global Public Health Priority. BMC Public Health 2011, 11, 770. [Google Scholar] [CrossRef] [PubMed]
- Holmes, D. The pain drain. Nature 2016, 535, S2–S3. [Google Scholar] [CrossRef] [PubMed]
- Disatnik, M.H.; Jones, S.N.; Mochly-Rosen, D. Stimulus-dependent subcellular localization of activated protein kinase C; a study with acidic fibroblast growth factor and transforming growth factor-β1 in cardiac myocytes. J. Mol. Cell. Cardiol. 1995, 27, 2473–2481. [Google Scholar] [CrossRef]
- Cesare, P.; Dekker, L.V.; Sardini, A.; Parker, P.J.; Mcnaughton, P.A. Specific Involvement of PKC-ε in Sensitization of the Neuronal Response to Painful Heat plaining the lowering of the threshold for heat pain that. Neuron 1999, 23, 617–624. [Google Scholar] [CrossRef]
- Khasar, S.G.; Lin, Y.-H.; Martin, A.; Dadgar, J.; McMahon, T.; Wang, D.; Hundle, B.; Aley, K.; Isenberg, W.; McCarter, G.; et al. A novel nociceptor signaling pathway revealed in protein kinase c ε mutant mice. Neuron 1999, 24, 253–260. [Google Scholar] [CrossRef]
- Wu, D.-F.; Chandra, D.; McMahon, T.; Wang, D.; Dadgar, J.; Kharazia, V.N.; Liang, Y.-J.; Waxman, S.G.; Dib-Hajj, S.D.; Messing, R. PKCε phosphorylation of the sodium channel Na V1.8 increases channel function and produces mechanical hyperalgesia in mice. J. Clin. Investig. 2012, 122, 1306–1315. [Google Scholar] [CrossRef]
- Cousins, M.J.; Pickthorn, K.; Huang, S.; Critchley, L.; Bell, G. The Safety and Efficacy of KAI-1678- An Inhibitor of Epsilon Protein Kinase C (εPKC)-Versus Lidocaine and Placebo for the Treatment of Postherpetic Neuralgia: A Crossover Study Design. Pain Med. 2013, 14, 533–540. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Moodie, J.E.; Bisley, E.J.; Huang, S.; Pickthorn, K.; Bell, G. A Single-Center, Randomized, Double-Blind, Active, and Placebo-Controlled Study of KAI-1678, a Novel PKC-Epsilon Inhibitor, in the Treatment of Acute Postoperative Orthopedic Pain. Pain Med. 2013, 916–924. [Google Scholar] [CrossRef]
- Zambelli, V.O.; Gross, E.R.; Chen, C.H.; Gutierrez, V.P.; Cury, Y.; Mochly-Rosen, D. Aldehyde dehydrogenase-2 regulates nociception in rodent models of acute inflammatory pain. Sci. Transl. Med. 2014, 6, 251ra118. [Google Scholar] [CrossRef]
- Chen, C.H.; Ferreira, J.C.B.; Gross, E.R.; Mochly-Rosen, D. Targeting aldehyde dehydrogenase 2: New therapeutic opportunities. Physiol. Rev. 2014, 94, 1–34. [Google Scholar] [CrossRef]
- Dalleau, S.; Baradat, M.; Guéraud, F.; Huc, L. Cell death and diseases related to oxidative stress:4-hydroxynonenal (HNE) in the balance. Cell Death Differ. 2013, 20, 1615–1630. [Google Scholar] [CrossRef] [PubMed]
- McAllister, S.L.; Sinharoy, P.; Vasu, M.; Gross, E.R. Aberrant reactive aldehyde detoxification by aldehyde dehydrogenase-2 influences endometriosis development and pain-associated behaviors. Pain 2021, 162, 71–83. [Google Scholar] [CrossRef]
- Khan, M.; Qiao, F.; Kumar, P.; Islam, S.M.T.; Singh, A.K.; Won, J.; Singh, I. Neuroprotective effects of Alda-1 mitigate spinal cord injury in mice: Involvement of Alda-1-induced ALDH2 activation-mediated suppression of reactive aldehyde mechanisms. Neural Regen. Res. 2022, 17, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Ritter, C.; Dalenogare, D.P.; de Almeida, A.S.; Pereira, V.L.; Pereira, G.C.; Fialho, M.F.P.; Lückemeyer, D.D.; Antoniazzi, C.; Kudsi, S.Q.; Ferreira, J.; et al. Nociception in a Progressive Multiple Sclerosis Model in Mice Is Dependent on Spinal TRPA1 Channel Activation. Mol. Neurobiol. 2020, 57, 2420–2435. [Google Scholar] [CrossRef] [PubMed]
- Marone, I.M.; De Logu, F.; Nassini, R.; Goncalves, M.D.C.; Benemei, S.; Ferreira, J.; Jain, P.; Puma, S.L.; Bunnett, N.W.; Geppetti, P.; et al. TRPA1/NOX in the soma of trigeminal ganglion neurons mediates migraine-related pain of glyceryl trinitrate in mice. Brain 2018, 141, 2312–2328. [Google Scholar] [CrossRef]
- Trevisani, M.; Siemens, J.; Materazzi, S.; Bautista, D.M.; Nassini, R.; Campi, B.; Imamachi, N.; Andre, E.; Patacchini, R.; Cottrell, G.S.; et al. 4-Hydroxynonenal, an endogenous aldehyde, causes pain and neurogenic inflammation through activation of the irritant receptor TRPA1. Anticancer Res. 2007, 104, 13519–13524. [Google Scholar] [CrossRef]
- Dalle-Donne, I.; Aldini, G.; Carini, M.; Colombo, R.; Rossi, R.; Milzani, A. Protein carbonylation, cellular dysfunction, and disease progression. J. Cell Mol. Med. 2006, 10, 389–406. [Google Scholar] [CrossRef]
- Castrillo, A.; Pennington, D.J.; Otto, F.; Parker, P.J.; Owen, M.J.; Boscá, L. Protein kinase Cε is required for macrophage activation and defense against bacterial infection. J. Exp. Med. 2001, 194, 1231–1242. [Google Scholar] [CrossRef]
- da Costa, D.S.M.; Meotti, F.C.; Andrade, E.L.; Leal, P.C.; Motta, E.M.; Emerson Marcelo Motta, J.B.C. The involvement of the transient receptor potential A1 (TRPA1) in the maintenance of mechanical and cold hyperalgesia in persistent inflammation. Pain 2010, 148, 431–437. [Google Scholar] [CrossRef]
- Trevisan, G.; Hoffmeisterc, C.; Rossatoa, M.F.; Oliveira, S.M.; Silva, M.A.; Silva, C.R.; Fusi, C.; Tonello, R.; Minocci, D.; Guerra, G.P. TRPA1 receptor stimulation by hydrogen peroxide is critical to triggerhyperalgesia and inflammation in a model of acute gout. Free Radic. Biol. Med. 2014, 72, 200–209. [Google Scholar] [CrossRef]
- Sant’Anna, M.; Kusuda, R.; Bozzo, T.A.; Bassi, G.S.; Alves-Filho, J.C.; Cunha, F.Q.; Ferreira, S.H.; Souza, G.R.; Cunha, T. Medial plantar nerve ligation as a novel model of neuropathic pain in mice: Pharmacological and molecular characterization. Sci. Rep. 2016, 6, 26955. [Google Scholar] [CrossRef]
- Sweitzer, S.; Wong, S. Protein Kinase C ϵ and γ Involvement in Formalin-Induced Nociception in Neonatal Rats. J. Pharmacol. Exp. Ther. 2004, 309, 616–625. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Wang, Z.J. Nociceptor beta II, delta, and epsilon isoforms of PKC differentially mediate paclitaxel-induced spontaneous and evoked pain. J. Neurosci. 2015, 35, 4614–4625. [Google Scholar] [CrossRef]
- Joseph, E.K.; Reichling, D.B.; Levine, J.D. Shared Mechanisms for Opioid Tolerance and a Transition to Chronic Pain. J. Neurosci. 2010, 30, 4660–4666. [Google Scholar] [CrossRef]
- Sun, X.; Budas, G.R.; Xu, L.; Barreto, G.E.; Mochly-Rosen, D.; Sun, X.; Budas, G.R.; Xu, L.; Barreto, G.E.; Daria Mochly-Rosen, R.G.G. Selective activation of protein kinase C∊ in mitochondria is neuroprotective in vitro and reduces focal ischemic brain injury in mice. J. Neurosci. Res. 2013, 91, 799–807. [Google Scholar] [CrossRef]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Dalenogare, D.P.; Theisen, M.C.; Peres, D.S.; Fialho, M.F.P.; Lückemeyer, D.D.; Antoniazzi, C.T.d.D.; Kudsi, S.Q.; Ferreira, M.D.A.; Ritter, C.D.S.; Ferreira, J.; et al. TRPA1 activation mediates nociception behaviors in a mouse model of relapsing-remitting experimental autoimmune encephalomyelitis. Exp. Neurol. 2020, 328, 113241. [Google Scholar] [CrossRef] [PubMed]
- Brooks, P.J.; Enoch, M.; Goldman, D.; Li, T.; Yokoyama, A. The Alcohol Flushing Response: An Unrecognized Risk Factor for Esophageal Cancer from Alcohol Consumption. PLoS Med. 2009, 6, 258–263. [Google Scholar] [CrossRef]
- Chen, C.-H.; Budas, G.R.; Churchill, E.N.; Disatnik, M.-H.; Hurley, T.D.; Mochly-Rosen, D. Activation of aldehyde dehydrogenase-2 reduces ischemic damage to the heart. Supporting Online Mater. Sci. 2008, 321, 1493–1495. [Google Scholar] [CrossRef] [PubMed]
- Budas, G.R.; Churchill, E.N.; Disatnik, M.H.; Sun, L.; Mochly-Rosen, D. Mitochondrial import of PKC is mediated by HSP90: A role in cardioprotection from ischaemia and reperfusion injury. Cardiovasc. Res. 2010, 88, 83–92. [Google Scholar] [CrossRef]
- Parada, C.A.; Yeh, J.J.; Reichling, D.B.; Levine, J.D. Transient attenuation of protein kinase Cε can terminate a chronic hyperalgesic state in the rat. Neuroscience 2003, 120, 219–226. [Google Scholar] [CrossRef]
- Numazaki, M.; Tominaga, T.; Toyooka, H.; Tominaga, M. Direct phosphorylation of capsaicin receptor VR1 by protein kinase Cε and identification of two target serine residues. J. Biol. Chem. 2002, 277, 13375–13378. [Google Scholar] [CrossRef]
- Srinivasan, R.; Wolfe, D.; Goss, J.; Watkins, S.; de Groat, W.C.; Sculptoreanu, A.; Glorioso, J.C. Protein kinase C epsilon contributes to basal and sensitizing responses of TRPV1 to capsaicin in rat dorsal root ganglion neurons. Eur. J. Neurosci. 2008, 28, 1241–1254. [Google Scholar] [CrossRef] [PubMed]
- Sachs, D.; Villarreal, C.; Cunha, F.; Parada, C.; Ferreira, S. The role of PKA and PKCe pathways in prostaglandin E2-mediated hypernociception. Br. J. Pharmacol. 2009, 156, 826–834. [Google Scholar] [CrossRef] [PubMed]
- Esterbauer, H.; Schaur, R.J.; Zollner, H. Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic. Biol. Med. 1991, 11, 81–128. [Google Scholar] [CrossRef]
- Strohmaier, H.; Highofer-Szalkay, H.; Schaur, R.J. Detection of 4-hydroxynonenal (HNE) as a physiological component in human plasma. J. Lipid Mediat. Cell Signal. 1995, 11, 51–61. [Google Scholar] [CrossRef]
- Zhong, H.; Yin, H. Role of lipid peroxidation derived 4-hydroxynonenal (4-HNE) in cancer: Focusing on mitochondria. Redox Biol. 2015, 4, 193–199. [Google Scholar] [CrossRef] [PubMed]
- El-Brolosy, M.A.; Stainier, D.Y.R. Genetic compensation: A phenomenon in search of mechanisms. PLoS Genet. 2017, 13, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Neupert, W.; Herrmann, J.M. Translocation of proteins into mitochondria. Annu. Rev. Biochem. 2007, 76, 723–749. [Google Scholar] [CrossRef] [PubMed]
- Perez-Miller, S.; Younus, H.; Vanam, R.; Chen, C.; Mochly-Rosen, D.; Hurley, T.D. Alda-1 is an agonist and chemical chaperone for the common human aldehyde dehydrogenase 2 variant. Nat. Struct Mol. Biol. 2010, 17, 159–164. [Google Scholar] [CrossRef]
- Dray, A.; Bettaney, J.; Forster, P.; Perkins, M.N. Bradykinin-induced stimulation of afferent fibres is mediated through protein kinase C. Neurosci. Lett. 1988, 91, 301–307. [Google Scholar] [CrossRef]
- Rang, H.P.; Ritchie, J.M. Depolarization of Nonmyelinated Fibers of the Rat Vagus Nerve Produced by Activation of Protein Kinase C. J. Neurosci. 1988, 8, 2606–2617. [Google Scholar] [CrossRef] [PubMed]
- Mefllinger, K.; Schmidt, R.F. The effects of phorbol ester on slowly conducting afferents of the cat’s knee joint. Exp. Brain Res. 1993, 92, 391–398. [Google Scholar]
- De Logu, F.; De Prá, S.D.-T.; Antoniazzi, C.T.D.D.; Kudsi, S.Q.; Ferro, P.R.; Landini, L.; Rigo, F.K.; Silveira, G.D.B.; Silveira, P.C.L.; Oliveira, S.M.; et al. Macrophages and Schwann cell TRPA1 mediate chronic allodynia in a mouse model of complex regional pain syndrome type I. Brain Behav. Immun. 2020, 88, 535–546. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Cinar, R.; Hu, X.; Lin, Y.; Luo, G.; Lovinger, D.M. Spinal astrocyte aldehyde dehydrogenase-2 mediates ethanol metabolism and analgesia in mice. Br. J. Anaesth. 2021, 1–14. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martins, B.B.; Hösch, N.G.; Alcantara, Q.A.; Budas, G.R.; Chen, C.-H.; Mochly-Rosen, D.; Ferreira, J.C.B.; Zambelli, V.O. Activation of PKCε-ALDH2 Axis Prevents 4-HNE-Induced Pain in Mice. Biomolecules 2021, 11, 1798. https://doi.org/10.3390/biom11121798
Martins BB, Hösch NG, Alcantara QA, Budas GR, Chen C-H, Mochly-Rosen D, Ferreira JCB, Zambelli VO. Activation of PKCε-ALDH2 Axis Prevents 4-HNE-Induced Pain in Mice. Biomolecules. 2021; 11(12):1798. https://doi.org/10.3390/biom11121798
Chicago/Turabian StyleMartins, Bárbara B., Natália G. Hösch, Queren A. Alcantara, Grant R. Budas, Che-Hong Chen, Daria Mochly-Rosen, Julio C. B. Ferreira, and Vanessa O. Zambelli. 2021. "Activation of PKCε-ALDH2 Axis Prevents 4-HNE-Induced Pain in Mice" Biomolecules 11, no. 12: 1798. https://doi.org/10.3390/biom11121798
APA StyleMartins, B. B., Hösch, N. G., Alcantara, Q. A., Budas, G. R., Chen, C.-H., Mochly-Rosen, D., Ferreira, J. C. B., & Zambelli, V. O. (2021). Activation of PKCε-ALDH2 Axis Prevents 4-HNE-Induced Pain in Mice. Biomolecules, 11(12), 1798. https://doi.org/10.3390/biom11121798