D-Aspartate Upregulates DAAM1 Protein Levels in the Rat Testis and Induces Its Localization in Spermatogonia Nucleus

 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experiments
2.2. Cell Culture and Treatments
2.3. Preparation of Total Protein Extracts from Rat Testis and GC-1 Cells and Western Blotting Analysis
2.4. Preparation of Nuclear Protein Extracts from Rat Testis and Western Blotting Analysis
2.5. Immunofluorescence Analysis on Rat Testis
2.6. Immunofluorescence Analysis on GC-1
2.7. Statistical Analysis
3. Results
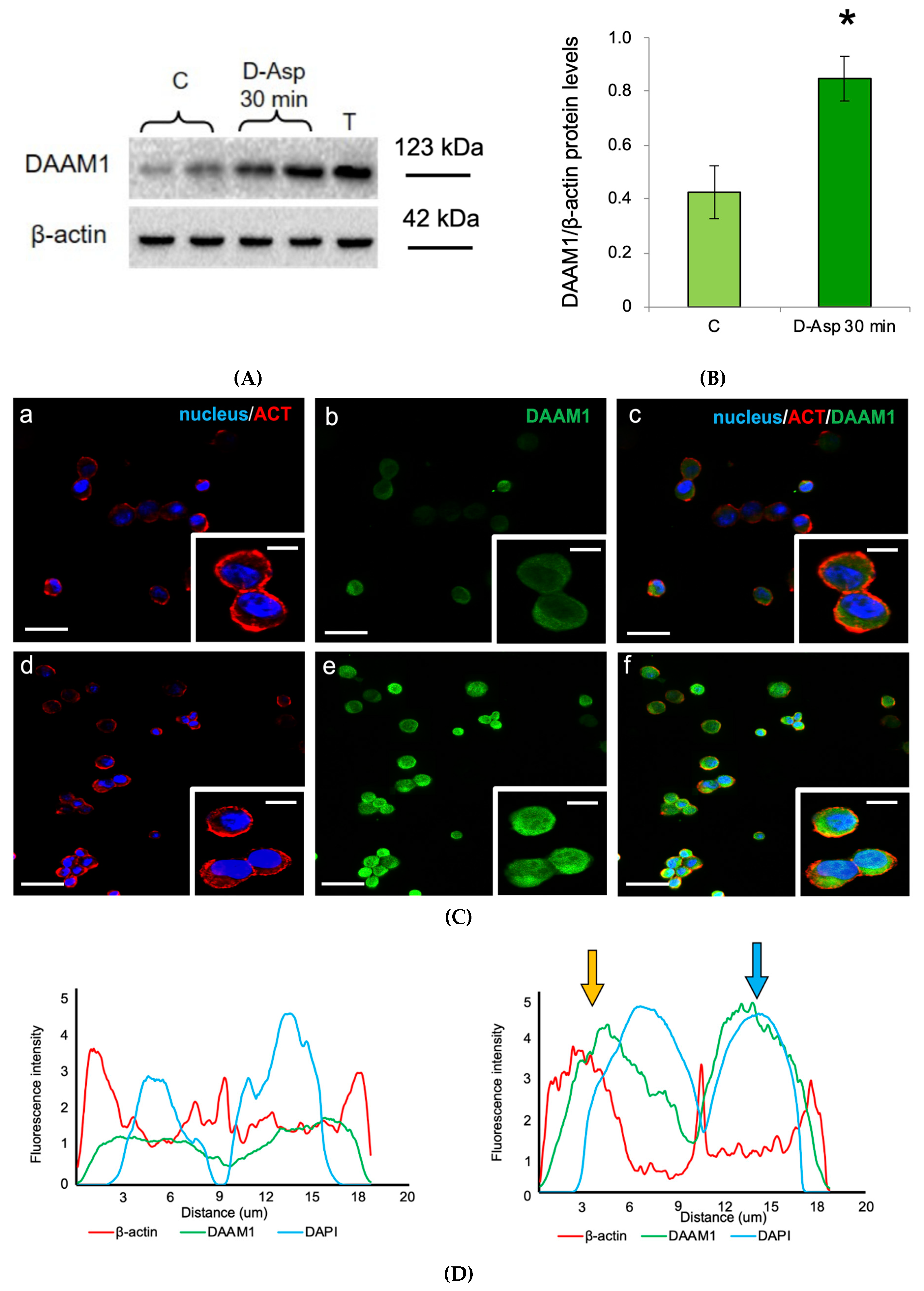
3.1. Effects of Oral Administration of D-Asp on Testicular DAAM1 Protein Levels and Localization
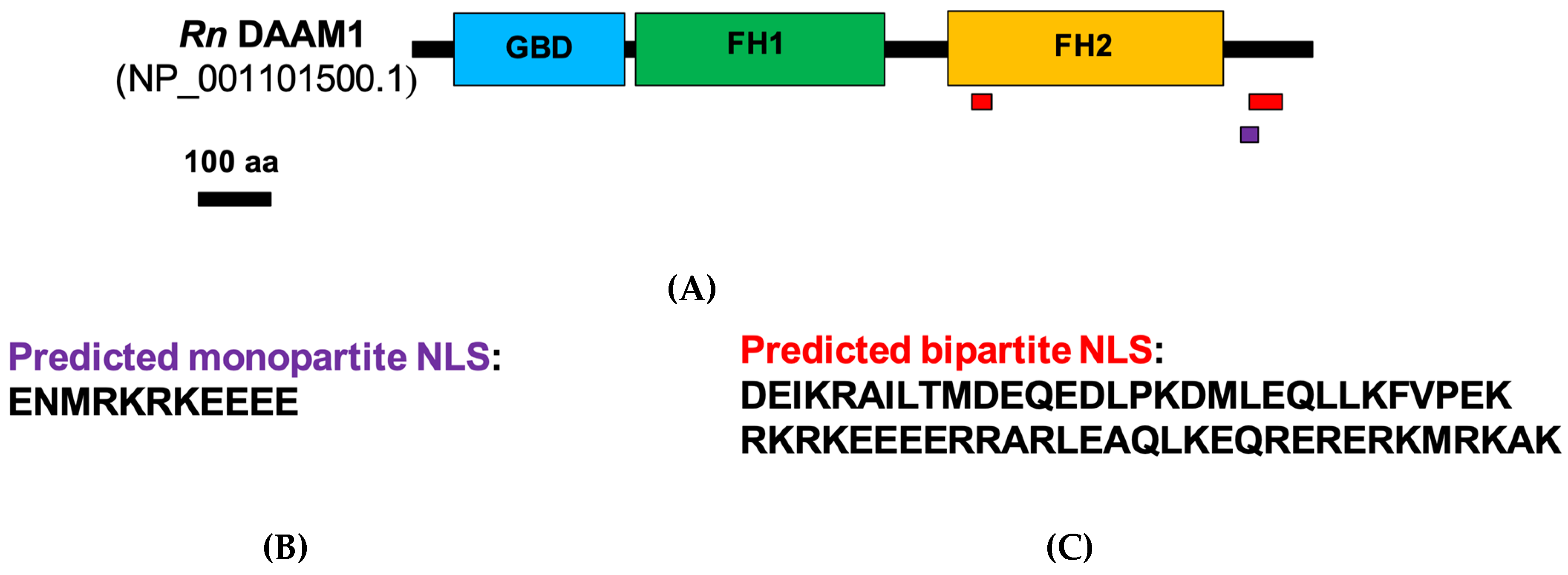
3.2. DAAM1 Nuclear Localization: Bioinformatics and Biochemical Analysis
3.3. Effects of D-Asp on DAAM1 Protein Levels and Localization in Mouse GC-1 Cells
3.4. Effects of Oral Administration of D-Asp on Testicular PCNA and p-H3 Protein Levels
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- McLean, D.J.; Friel, P.J.; Johnston, D.S.; Griswold, M.D. Characterization of Spermatogonial Stem Cell Maturation and Differentiation in Neonatal Mice1. Biol. Reprod. 2003, 69, 2085–2091. [Google Scholar] [CrossRef] [PubMed]
- Venditti, M.; Fasano, C.; Minucci, S.; Serino, I.; Sinisi, A.A.; Dale, B.; Di Matteo, L. DAAM1 and PREP are involved in human spermatogenesis. Reprod. Fertil. Dev. 2020, 32, 484. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.S.; Shakes, D.C. Spermatogenesis. In Germ Cell Development in C. elegans; Springer: New York, NY, USA, 2012; Volume 757, pp. 171–203. [Google Scholar]
- Pariante, P.; Dotolo, R.; Venditti, M.; Ferrara, D.; Donizetti, A.; Aniello, F.; Minucci, S. Prothymosin alpha expression and localization during the spermatogenesis of Danio rerio. Zygote 2015, 24, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Venditti, M.; Minucci, S. Prothymosin alpha expression in the vertebrate testis: A comparative review. Zygote 2017, 25, 760–770. [Google Scholar] [CrossRef] [PubMed]
- Ergoli, M.; Venditti, M.; Piccillo, E.; Minucci, S.; Politano, L. Study of Expression of Genes Potentially Responsible for Reduced Fitness in Patients With Myotonic Dystrophy Type 1 and Identification of New Biomarkers of Testicular Function. Mol. Reprod. Dev. 2020, 87, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.Q.; Garbers, D.L. Male Germ Cell Specification and Differentiation. Dev. Cell 2002, 2, 537–547. [Google Scholar] [CrossRef]
- Flück, C.E.; Pandey, A.V. Steroidogenesis of the testis—New genes and pathways. In Annales d’endocrinologie; Elsevier Masson: Paris, France, 2014; Volume 75, pp. 40–47. [Google Scholar] [CrossRef]
- Smith, L.B.; Walker, W.H. The regulation of spermatogenesis by androgens. Semin. Cell Dev. Biol. 2014, 30, 2–13. [Google Scholar] [CrossRef]
- Tremblay, J.J. Molecular regulation of steroidogenesis in endocrine Leydig cells. Steroids 2015, 103, 3–10. [Google Scholar] [CrossRef]
- D’Aniello, G. d-Aspartic acid: An endogenous amino acid with an important neuroendocrine role. Brain Res. Rev. 2007, 53, 215–234. [Google Scholar] [CrossRef]
- Di Fiore, M.M.; Santillo, A.; Baccari, G.C. Current knowledge of d-aspartate in glandular tissues. Amino Acids 2014, 46, 1805–1818. [Google Scholar] [CrossRef]
- Furuchi, T.; Homma, H. Free D-Aspartate in Mammals. Biol. Pharm. Bull. 2005, 28, 1566–1570. [Google Scholar] [CrossRef] [PubMed]
- Ota, N.; Shi, T.; Sweedler, J.V. D-Aspartate acts as a signaling molecule in nervous and neuroendocrine systems. Amino Acids 2012, 43, 1873–1886. [Google Scholar] [CrossRef] [PubMed]
- Burrone, L.; Di Giovanni, M.; Di Fiore, M.M.; Baccari, G.C.; Santillo, A. Effects of D-Aspartate Treatment on D-Aspartate Oxidase, Superoxide Dismutase, and Caspase 3 Activities in Frog (Rana esculenta) Tissues. Chem. Biodivers. 2010, 7, 1459–1466. [Google Scholar] [CrossRef] [PubMed]
- Lamanna, C.; Assisi, L.; Botte, V.; Di Fiore, M.M. Involvement of D-Asp in P450 aromatase activity and estrogen receptors in boar testis. Amino Acids 2006, 32, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Lamanna, C.; Assisi, L.; Vittoria, A.; Botte, V.; Di Fiore, M.M. d-Aspartic acid and nitric oxide as regulators of androgen production in boar testis. Theriogenology 2007, 67, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Santillo, A.; Falvo, S.; Chieffi, P.; Burrone, L.; Baccari, G.C.; Longobardi, S.; Di Fiore, M.M. d-aspartate affects NMDA receptor-extracellular signal–regulated kinase pathway and upregulates androgen receptor expression in the rat testis. Theriogenology 2014, 81, 744–751. [Google Scholar] [CrossRef]
- Santillo, A.; Chieffi Baccari, G.; Falvo, S.; Di Giacomo Russo, F.; Venditti, M.; Di Fiore, M.M. Effects of D-aspartate on sex hormone-dependent tissues in Pelophylax esculentus. In Amphibians: Biology, Ecology and Conservation; Cannon, L., Ed.; Nova Science Publishers: New York, NY, USA, 2018; pp. 21–37. [Google Scholar]
- Santillo, A.; Falvo, S.; Chieffi Baccari, G.; Di Giacomo Russo, F.; Rosati, L.; Di Fiore, M.M. Testis and brain steroidogenesis during the reproductive cycle of Pelophylax esculentus. In Amphibians: Biology, Ecology and Conservation; Cannon, L., Ed.; Nova Science Publishers: New York, NY, USA, 2018; pp. 39–54. [Google Scholar]
- Di Giovanni, M.; Burrone, L.; Baccari, G.C.; Topo, E.; Santillo, A. Distribution of free D-aspartic acid and D-aspartate oxidase in frogRana esculentatissues. J. Exp. Zoöl. Part A: Ecol. Genet. Physiol. 2010, 313, 137–143. [Google Scholar] [CrossRef]
- D’Aniello, A.; Di Fiore, M.M.; Fisher, G.; Milone, A.; Seleni, A.; D’Aniello, S.; Perna, A.F.; Ingrosso, D. Occurrence of D-aspartic acid and N-methyl-D-aspartic acid in rat neuroendocrine tissues and their role in the modulation of luteinizing hormone and growth hormone release. FASEB J. 2000, 14, 699–714. [Google Scholar] [CrossRef]
- Topo, E.; Soricelli, A.; D’Aniello, A.; Ronsini, S.; D’Aniello, G. The role and molecular mechanism of D-aspartic acid in the release and synthesis of LH and testosterone in humans and rats. Reprod. Biol. Endocrinol. 2009, 7, 120. [Google Scholar] [CrossRef]
- Di Fiore, M.M.; Santillo, A.; Falvo, S.; Longobardi, S.; Baccari, G.C. Molecular Mechanisms Elicited by d-Aspartate in Leydig Cells and Spermatogonia. Int. J. Mol. Sci. 2016, 17, 1127. [Google Scholar] [CrossRef]
- Di Fiore, M.M.; Burrone, L.; Santillo, A.; Chieffi Baccari, G. Endocrine activity of D-aspartate in nonmammalian animals. In D-Amino Acids: Physiology, Metabolism, and Application; Yoshimura, T., Nishikawa, T., Homma, H., Eds.; Springer: Tokyo, Japan, 2016; pp. 157–172. [Google Scholar]
- Di Fiore, M.M.; Boni, R.; Santillo, A.; Falvo, S.; Gallo, A.; Esposito, S.; Baccari, G.C. D-Aspartic Acid in Vertebrate Reproduction: Animal Models and Experimental Designs. Biomolecules 2019, 9, 445. [Google Scholar] [CrossRef] [PubMed]
- Nagata, Y.; Homma, H.; Lee, J.-A.; Imai, K. D-Aspartate stimulation of testosterone synthesis in rat Leydig cells. FEBS Lett. 1999, 444, 160–164. [Google Scholar] [CrossRef]
- Nagata, Y.; Homma, H.; Matsumoto, M.; Imai, K. Stimulation of steroidogenic acute regulatory protein (STAR) gene expression by D-aspartate in rat Leydig cells. FEBS Lett. 1999, 454, 317–320. [Google Scholar] [CrossRef] [PubMed]
- Santillo, A.; Falvo, S.; Chieffi, P.; Di Fiore, M.M.; Senese, R.; Baccari, G.C. D-Aspartate Induces Proliferative Pathways in Spermatogonial GC-1 Cells. J. Cell. Physiol. 2015, 231, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Santillo, A.; Falvo, S.; Di Fiore, M.M.; Russo, F.D.G.; Chieffi, P.; Usiello, A.; Pinelli, C.; Baccari, G.C. AMPA receptor expression in mouse testis and spermatogonial GC-1 cells: A study on its regulation by excitatory amino acids. J. Cell. Biochem. 2019, 120, 11044–11055. [Google Scholar] [CrossRef] [PubMed]
- Sakai, K.; Homma, H.; Lee, J.-A.; Fukushima, T.; Santa, T.; Tashiro, K.; Iwatsubo, T.; Imai, K. Localization ofd-Aspartic Acid in Elongate Spermatids in Rat Testis. Arch. Biochem. Biophys. 1998, 351, 96–105. [Google Scholar] [CrossRef]
- Tomita, K.; Tanaka, H.; Kageyama, S.; Nagasawa, M.; Wada, A.; Murai, R.; Kobayashi, K.; Hanada, E.; Agata, Y.; Kawauchi, A. The Effect of d-Aspartate on Spermatogenesis in Mouse Testis1. Biol. Reprod. 2016, 94, 30. [Google Scholar] [CrossRef]
- D’Aniello, G.; Ronsini, S.; Guida, F.; Spinelli, P.; D’Aniello, A. Occurrence of D-aspartic acid in human seminal plasma and spermatozoa: Possible role in reproduction. Fertil. Steril. 2005, 84, 1444–1449. [Google Scholar] [CrossRef]
- D’Aniello, A.; Di Cosmo, A.; Di Cristo, C.; Annunziato, L.; Petrucelli, L.; Fisher, G. Involvement of D-Aspartic acid in the synthesis of testosterone in rat testes. Life Sci. 1996, 59, 97–104. [Google Scholar] [CrossRef]
- Santillo, A.; Venditti, M.; Minucci, S.; Baccari, G.C.; Falvo, S.; Rosati, L.; Di Fiore, M.M. D-Asp upregulates PREP and GluA2/3 expressions and induces p-ERK1/2 and p-Akt in rat testis. Reproduction 2019, 158, 357–367. [Google Scholar] [CrossRef]
- Dotolo, R.; Kim, J.D.; Pariante, P.; Minucci, S.; Diano, S. Prolyl Endopeptidase (PREP) is Associated With Male Reproductive Functions and Gamete Physiology in Mice. J. Cell. Physiol. 2015, 231, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Venditti, M.; Minucci, S. Subcellular Localization of Prolyl Endopeptidase during the First Wave of Rat Spermatogenesis and in Rat and Human Sperm. J. Histochem. Cytochem. 2018, 67, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Venditti, M.; Chemek, M.; Minucci, S.; Messaoudi, I. Cadmium-induced toxicity increases prolyl endopeptidase (PREP) expression in the rat testis. Mol. Reprod. Dev. 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- Schulz, I.; Zeitschel, U.; Rudolph, T.; Rahfeld, J.-U.; Gerhartz, B.; Bigl, V.; DeMuth, H.-U.; Roßner, S.; Ruiz-Carrillo, D. Subcellular localization suggests novel functions for prolyl endopeptidase in protein secretion. J. Neurochem. 2005, 94, 970–979. [Google Scholar] [CrossRef] [PubMed]
- Venditti, M.; Aniello, F.; Santillo, A.; Minucci, S. Study on PREP localization in mouse seminal vesicles and its possible involvement during regulated exocytosis. Zygote 2019, 27, 160–165. [Google Scholar] [CrossRef]
- Li, L.; Mao, B.; Wu, S.; Lian, Q.; Ge, R.-S.; Silvestrini, B.; Cheng, C.Y. Regulation of spermatid polarity by the actin- and microtubule (MT)-based cytoskeletons. Semin. Cell Dev. Biol. 2018, 81, 88–96. [Google Scholar] [CrossRef]
- Pariante, P.; Dotolo, R.; Venditti, M.; Ferrara, D.; Donizetti, A.; Aniello, F.; Minucci, S. First Evidence of DAAM1 Localization during the Post-Natal Development of Rat Testis and in Mammalian Sperm. J. Cell. Physiol. 2016, 231, 2172–2184. [Google Scholar] [CrossRef]
- Kovar, D.R. Molecular details of formin-mediated actin assembly. Curr. Opin. Cell Biol. 2006, 18, 11–17. [Google Scholar] [CrossRef]
- Falvo, S.; Baccaria, G.C.; Spaziano, G.; Rosati, L.; Venditti, M.; Di Fiore, M.M.; Santillo, A.; Di Flore, M.M. StAR protein and steroidogenic enzyme expressions in the rat Harderian gland. Comptes Rendus Biol. 2018, 341, 160–166. [Google Scholar] [CrossRef]
- Hofmann, M.-C.; Narisawa, S.; Hess, R.; Millán, J.L. Immortalization of germ cells and somatic testicular cells using the SV40 large T antigen. Exp. Cell Res. 1992, 201, 417–435. [Google Scholar] [CrossRef]
- Santillo, A.; Pinelli, C.; Burrone, L.; Baccari, G.C.; Di Fiore, M.M. d-Aspartic acid implication in the modulation of frog brain sex steroid levels. Gen. Comp. Endocrinol. 2013, 181, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Chemek, M.; Venditti, M.; Boughamoura, S.; Mimouna, S.B.; Messaoudi, I.; Minucci, S. Involvement of testicular DAAM1 expression in zinc protection against cadmium-induced male rat reproductive toxicity. J. Cell. Physiol. 2017, 233, 630–640. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Sato, A.; Khadka, D.; Bharti, R.; Diaz, H.; Runnels, L.; Habas, R. Mechanism of activation of the Formin protein Daam1. Proc. Natl. Acad. Sci. USA 2007, 105, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Kosugi, S.; Hasebe, M.; Tomita, M.; Yanagawa, H. Systematic identification of cell cycle-dependent yeast nucleocytoplasmic shuttling proteins by prediction of composite motifs. Proc. Natl. Acad. Sci. USA 2009, 106, 10171–10176. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Duan, F.; Zhou, X.; Pan, H.; Li, R. Differential responses of GC-1 spermatogonia cells to high and low doses of bisphenol A. Mol. Med. Rep. 2018, 18, 3034–3040. [Google Scholar] [CrossRef]
- Li, C.; Zhu, X.; Chen, S.; Chen, L.; Zhao, Y.; Jiang, Y.; Gao, S.; Wang, F.; Liu, Z.; Fan, R.; et al. Melatonin promotes the proliferation of GC-1 spg cells by inducing metallothionein-2 expression through ERK1/2 signaling pathway activation. Oncotarget 2017, 8, 65627–65641. [Google Scholar] [CrossRef]
- Boehm, E.M.; Gildenberg, M.S.; Washington, T. The Many Roles of PCNA in Eukaryotic DNA Replication. Enzymes 2016, 39, 231–254. [Google Scholar] [CrossRef]
- Nowak, S.; Corces, V.G. Phosphorylation of histone H3: A balancing act between chromosome condensation and transcriptional activation. Trends Genet. 2004, 20, 214–220. [Google Scholar] [CrossRef]
- Moujaber, O.; Stochaj, U. The Cytoskeleton as Regulator of Cell Signaling Pathways. Trends Biochem. Sci. 2020, 45, 96–107. [Google Scholar] [CrossRef]
- Oda, T.; Iwasa, M.; Aihara, T.; Maéda, Y.; Narita, A. The nature of the globular- to fibrous-actin transition. Nature 2009, 457, 441–445. [Google Scholar] [CrossRef]
- Pollard, T.D. Actin and Actin-Binding Proteins. Cold Spring Harb. Perspect. Biol. 2016, 8, a018226. [Google Scholar] [CrossRef] [PubMed]
- Chesarone, M.A.; DuPage, A.G.; Goode, B.L. Unleashing formins to remodel the actin and microtubule cytoskeletons. Nat. Rev. Mol. Cell Biol. 2009, 11, 62–74. [Google Scholar] [CrossRef]
- Schönichen, A.; Geyer, M. Fifteen formins for an actin filament: A molecular view on the regulation of human formins. Biochim. Biophys. Acta (BBA) Bioenerg. 2010, 1803, 152–163. [Google Scholar] [CrossRef]
- Chianese, R.; Cobellis, G.; Chioccarelli, T.; Ciaramella, V.; Migliaccio, M.; Fasano, S.; Pierantoni, R.; Meccariello, R. Kisspeptins, Estrogens and Male Fertility. Curr. Med. Chem. 2016, 23, 4070–4091. [Google Scholar] [CrossRef]
- Di Fiore, M.M.; Santillo, A.; Falvo, S.; Baccari, G.C.; Venditti, M.; Russo, F.D.G.; Lispi, M.; D’Aniello, G. Sex hormone levels in the brain of d -aspartate-treated rats. C. R. Biol. 2018, 341, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Dunleavy, J.; O’Bryan, M.K.; Stanton, P.G.; O’Donnell, L. The cytoskeleton in spermatogenesis. Reproduction 2019, 157, R53–R72. [Google Scholar] [CrossRef]
- Tanegashima, K.; Zhao, H.; Dawid, I.B. WGEF activates Rho in the Wnt–PCP pathway and controls convergent extension in Xenopus gastrulation. EMBO J. 2008, 27, 606–617. [Google Scholar] [CrossRef] [PubMed]
- Marlow, F.; Topczewski, J.; Sepich, D.; Solnica-Krezel, L. Zebrafish Rho Kinase 2 Acts Downstream of Wnt11 to Mediate Cell Polarity and Effective Convergence and Extension Movements. Curr. Biol. 2002, 12, 876–884. [Google Scholar] [CrossRef]
- Habas, R.; Kato, Y.; He, X. Wnt/Frizzled Activation of Rho Regulates Vertebrate Gastrulation and Requires a Novel Formin Homology Protein Daam1. Cell 2001, 107, 843–854. [Google Scholar] [CrossRef]
- Cingolani, L.A.; Goda, Y. Actin in action: The interplay between the actin cytoskeleton and synaptic efficacy. Nat. Rev. Neurosci. 2008, 9, 344–356. [Google Scholar] [CrossRef]
- Salomon, S.N.; Haber, M.; Murai, K.K.; Dunn, R.J. Localization of the Diaphanous-related formin Daam1 to neuronal dendrites. Neurosci. Lett. 2008, 447, 62–67. [Google Scholar] [CrossRef] [PubMed]
- McLeod, F.; Salinas, P.C. Wnt proteins as modulators of synaptic plasticity. Curr. Opin. Neurobiol. 2018, 53, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Asrar, S.; Jia, Z. Molecular mechanisms coordinating functional and morphological plasticity at the synapse: Role of GluA2/N-cadherin interaction-mediated actin signaling in mGluR-dependent LTD. Cell. Signal. 2013, 25, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Di Giovanni, M.; Topo, E.; Santillo, A.; D’Aniello, A.; Baccari, G.C. d-Aspartate binding sites in rat Harderian gland. Amino Acids 2009, 38, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Percipalle, P. An actin-based nucleoskeleton involved in gene regulation and genome organization. Biochem. Biophys. Res. Commun. 2018, 506, 378–386. [Google Scholar] [CrossRef]
- Gonsior, S.M.; Platz, S.; Buchmeier, S.; Scheer, U.; Jockusch, B.M.; Hinssen, H. Conformational difference between nuclear and cytoplasmic actin as detected by a monoclonal anti-body. J. Cell. Sci. 1999, 112, 797–809. [Google Scholar]
- Grzanka, A.; Grzanka, A.; Orlikowska, M. Fluorescence and ultrastructural localization of actin distribution patterns in the nucleus of HL-60 and K-562 cell lines treated with cytostatic drugs. Oncol. Rep. 2004, 11, 765–770. [Google Scholar] [CrossRef][Green Version]
- Schoenenberger, C.-A.; Buchmeier, S.; Boerries, M.; Sütterlin, R.; Aebi, U.; Jockusch, B. Conformation-specific antibodies reveal distinct actin structures in the nucleus and the cytoplasm. J. Struct. Biol. 2005, 152, 157–168. [Google Scholar] [CrossRef]
- Kelpsch, D.J.; Tootle, T. Nuclear Actin: From Discovery to Function. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2018, 301, 1999–2013. [Google Scholar] [CrossRef]
- Stüven, T.; Hartmann, E.; Görlich, D. Exportin 6: A novel nuclear export receptor that is specific for profilin·actin complexes. EMBO J. 2003, 22, 5928–5940. [Google Scholar] [CrossRef]
- Dopie, J.; Skarp, K.-P.; Rajakylä, E.K.; Tanhuanpää, K.; Vartiainen, M.K. Active maintenance of nuclear actin by importin 9 supports transcription. Proc. Natl. Acad. Sci. USA 2012, 109, E544–E552. [Google Scholar] [CrossRef] [PubMed]
- Grosse, R.; Vartiainen, M.K. To be or not to be assembled: Progressing into nuclear actin filaments. Nat. Rev. Mol. Cell Biol. 2013, 14, 693–697. [Google Scholar] [CrossRef] [PubMed]
- Isogai, T.; Innocenti, M. New nuclear and perinuclear functions of formins. Biochem. Soc. Trans. 2016, 44, 1701–1708. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.C.; Leder, P. Genetic Evidence That Formins Function within the Nucleus. J. Biol. Chem. 1996, 271, 23472–23477. [Google Scholar] [CrossRef] [PubMed]
- Johnston, R.J.; Copeland, J.W.; Fasnacht, M.; Etchberger, J.F.; Liu, J.; Honig, B.; Hobert, O. An unusual Zn-finger/FH2 domain protein controls a left/right asymmetric neuronal fate decision in C. elegans. Development 2006, 133, 3317–3328. [Google Scholar] [CrossRef] [PubMed]
- Copeland, S.J.; Green, B.J.; Burchat, S.; Papalia, G.A.; Banner, D.; Copeland, J.W. The Diaphanous Inhibitory Domain/Diaphanous Autoregulatory Domain Interaction Is Able to Mediate Heterodimerization between mDia1 and mDia2. J. Biol. Chem. 2007, 282, 30120–30130. [Google Scholar] [CrossRef]
- O’Rourke, D.; Liu, Z.X.; Sellin, L.; Spokes, K.; Zeller, R.; Cantley, L.G. Hepatocyte growth factor induces MAPK-dependent formin IV translocation in renal epithelial cells. J. Am. Soc. Nephrol. 2000, 11, 2212–2221. [Google Scholar]
- Ménard, I.; Gervais, F.G.; Nicholson, D.W.; Roy, S. Caspase-3 cleaves the formin-homology-domain-containing protein FHOD1 during apoptosis to generate a C-terminal fragment that is targeted to the nucleolus. Apoptosis 2006, 11, 1863–1876. [Google Scholar] [CrossRef]
- Baarlink, C.; Wang, H.; Grosse, R. Nuclear Actin Network Assembly by Formins Regulates the SRF Coactivator MAL. Science 2013, 340, 864–867. [Google Scholar] [CrossRef]
- Kyheröinen, S.; Vartiainen, M.K. Nuclear actin dynamics in gene expression and genome organization. Semin. Cell Dev. Biol. 2019. [Google Scholar] [CrossRef]
- Parisis, N.; Krasinska, L.; Harker, B.; Urbach, S.; Rossignol, M.; Camasses, A.; Dewar, J.; Morin, N.; Fisher, D. Initiation of DNA replication requires actin dynamics and formin activity. EMBO J. 2017, 36, 3212–3231. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Venditti, M.; Santillo, A.; Falvo, S.; Di Fiore, M.M.; Chieffi Baccari, G.; Minucci, S. D-Aspartate Upregulates DAAM1 Protein Levels in the Rat Testis and Induces Its Localization in Spermatogonia Nucleus. Biomolecules 2020, 10, 677. https://doi.org/10.3390/biom10050677
Venditti M, Santillo A, Falvo S, Di Fiore MM, Chieffi Baccari G, Minucci S. D-Aspartate Upregulates DAAM1 Protein Levels in the Rat Testis and Induces Its Localization in Spermatogonia Nucleus. Biomolecules. 2020; 10(5):677. https://doi.org/10.3390/biom10050677
Chicago/Turabian StyleVenditti, Massimo, Alessandra Santillo, Sara Falvo, Maria Maddalena Di Fiore, Gabriella Chieffi Baccari, and Sergio Minucci. 2020. "D-Aspartate Upregulates DAAM1 Protein Levels in the Rat Testis and Induces Its Localization in Spermatogonia Nucleus" Biomolecules 10, no. 5: 677. https://doi.org/10.3390/biom10050677
APA StyleVenditti, M., Santillo, A., Falvo, S., Di Fiore, M. M., Chieffi Baccari, G., & Minucci, S. (2020). D-Aspartate Upregulates DAAM1 Protein Levels in the Rat Testis and Induces Its Localization in Spermatogonia Nucleus. Biomolecules, 10(5), 677. https://doi.org/10.3390/biom10050677