Temporal Effects on Radiation Responses in Nonhuman Primates: Identification of Biofluid Small Molecule Signatures by Gas Chromatography–Mass Spectrometry Metabolomics


 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. NHP System
2.2. Sample Preparation and GC-MS Instrumentation
2.2.1. Chemicals
2.2.2. Sample Preparation
2.2.3. GC/MS Instrumentation
2.2.4. Data Processing and Statistical Analysis
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Ionizing Radiation | (IR) |
| Radiological Dispersal Device | (RDD) |
| Improvised Nuclear Device | (IND) |
| National Institute of Allergy and Infectious Diseases | (NIAID) |
| Centers for Medical Countermeasures Against Radiation Consortium | (CMCRC) |
| Mass Spectrometry | (MS) |
| Acute Radiation Syndrome | (ARS) |
| Nuclear Magnetic Resonance | (NMR) |
| Tandem Quadrupole | (TQ) |
| Fourier-transform Ion Cyclotron Resonance | (FTICR) |
| Desorption Electrospray Ionization | (DESI) |
| Ultra-performance Liquid Chromatography | (UPLC) |
| Gas Chromatography | (GC) |
| Capillary Electrophoresis | (CE) |
| Nonhuman Primate | (NHP) |
| Total Body Irradiation | (TBI) |
| Time-of-Flight | (TOF) |
| Tricarboxylic Acid | (TCA) |
| Multiple Reaction Monitoring | (MRM) |
| Electron Ionization | (EI) |
| Partial Least Squares-Discriminant Analysis | (PLS-DA) |
| Cross-validation | (CV) |
| Differential Mobility Spectrometry | (DMS) |
References
- Czarwinski, R.; Crick, M.J. Occupational exposures worldwide and revision of international standards for protection. Radiat. Prot. Dosim. 2011, 144, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Izrael, Y.A. Chernobyl radionuclide distribution and migration. Health Phys. 2007, 93, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Mc Laughlin, J.P. Some characteristics and effects of natural radiation. Radiat. Prot. Dosim. 2015, 167, 2–7. [Google Scholar] [CrossRef]
- Ozasa, K.; Grant, E.J.; Kodama, K. Japanese legacy cohorts: The life span study atomic bomb survivor cohort and survivors’ offspring. J. Epidemiol. 2018, 28, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Ozasa, K.; Cullings, H.M.; Ohishi, W.; Hida, A.; Grant, E.J. Epidemiological studies of atomic bomb radiation at the Radiation Effects Research Foundation. Int. J. Radiat. Biol. 2019, 1–13. [Google Scholar] [CrossRef] [PubMed]
- DiCarlo, A.L.; Ramakrishnan, N.; Hatchett, R.J. Radiation combined injury: Overview of NIAID research. Health Phys. 2010, 98, 863–867. [Google Scholar] [CrossRef] [PubMed]
- Pannkuk, E.L.; Fornace, A.J., Jr.; Laiakis, E.C. Metabolomic applications in radiation biodosimetry: Exploring radiation effects through small molecules. Int. J. Radiat. Biol. 2017, 93, 1151–1176. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.H.; Ivanisevic, J.; Siuzdak, G. Metabolomics: Beyond biomarkers and towards mechanisms. Nat. Rev. Mol. Cell Biol. 2016, 17, 451–459. [Google Scholar] [CrossRef]
- Gowda, G.A.; Djukovic, D. Overview of mass spectrometry-based metabolomics: Opportunities and challenges. Methods Mol. Biol. 2014, 1198, 3–12. [Google Scholar] [PubMed]
- Markley, J.L.; Brüschweiler, R.; Edison, A.S.; Eghbalnia, H.R.; Powers, R.; Raftery, D.; Wishart, D.S. The future of NMR-based metabolomics. Curr. Opin. Biotechnol. 2017, 43, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.; Xiang, X.; Ong, P.; Mitchell, E.; Syn, N.; Wee, I.; Kumar, A.P.; Yong, W.P.; Sethi, G.; Goh, B.C.; et al. A review on liquid chromatography-tandem mass spectrometry methods for rapid quantification of oncology drugs. Pharmaceutics 2018, 10, 221. [Google Scholar] [CrossRef]
- Clendinen, C.S.; Monge, M.E.; Fernández, F.M. Ambient mass spectrometry in metabolomics. Analyst 2017, 142, 3101–3117. [Google Scholar] [CrossRef] [PubMed]
- Fuhrer, T.; Zamboni, N. High-throughput discovery metabolomics. Curr. Opin. Biotechnol. 2015, 31, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Horning, E.C.; Horning, M.G. Metabolic profiles: Chromatographic methods for isolation and characterization of a variety of metabolites in man. Methods Med. Res. 1970, 12, 369–371. [Google Scholar] [PubMed]
- Horning, E.C.; Horning, M.G. Metabolic profiles: Gas-phase methods for analysis of metabolites. Clin. Chem. 1971, 17, 802–809. [Google Scholar]
- Fiehn, O. Metabolomics by gas chromatography-mass spectrometry: Combined targeted and untargeted profiling. Curr. Protoc. Mol. Biol. 2016, 114, 30.4.1–30.4.32. [Google Scholar] [PubMed]
- Beale, D.J.; Pinu, F.R.; Kouremenos, K.A.; Poojary, M.M.; Narayana, V.K.; Boughton, B.A.; Kanojia, K.; Dayalan, S.; Jones, O.A.H.; Dias, D.A. Review of recent developments in GC-MS approaches to metabolomics-based research. Metabolomics 2018, 14, 152. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Bowen, B.P.; Nguyen, D.H.; Parsa, S.; Huang, Y.; Mao, J.H.; Northen, T.R. Low-dose ionizing radiation-induced blood plasma metabolic response in a diverse genetic mouse population. Radiat. Res. 2012, 178, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Lanz, C.; Patterson, A.D.; Slavik, J.; Krausz, K.W.; Ledermann, M.; Gonzalez, F.J.; Idle, J.R. Radiation metabolomics. 3. Biomarker discovery in the urine of gamma-irradiated rats using a simplified metabolomics protocol of gas chromatography-mass spectrometry combined with random forests machine learning algorithm. Radiat. Res. 2009, 172, 198–212. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Lau, K.K.; Zhou, X.; Wu, J.; Yang, J.; Wang, C. Urinary metabolic signatures and early triage of acute radiation exposure in rat model. Mol. Biosyst. 2017, 13, 756–766. [Google Scholar] [CrossRef]
- Liu, H.; Wang, Z.; Zhang, X.; Qiao, Y.; Wu, S.; Dong, F.; Chen, Y. Selection of candidate radiation biomarkers in the serum of rats exposed to gamma-rays by GC/TOFMS-based metabolomics. Radiat. Prot. Dosim. 2013, 154, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Pannkuk, E.L.; Laiakis, E.C.; Authier, S.; Wong, K.; Fornace, A.J., Jr. Gas chromatography/mass spectrometry metabolomics of urine and serum from nonhuman primates exposed to ionizing radiation: Impacts on the tricarboxylic acid cycle and protein metabolism. J. Proteome Res. 2017, 16, 2091–2100. [Google Scholar] [CrossRef]
- Laiakis, E.C.; Pannkuk, E.L.; Chauthe, S.K.; Wang, Y.W.; Lian, M.; Mak, T.D.; Barker, C.A.; Astarita, G.; Fornace, A.J., Jr. A serum small molecule biosignature of radiation exposure from total body irradiated patients. J. Proteome Res. 2017, 16, 3805–3815. [Google Scholar] [CrossRef] [PubMed]
- Mörén, L.; Wibom, C.; Bergström, P.; Johansson, M.; Antti, H.; Bergenheim, A.T. Characterization of the serum metabolome following radiation treatment in patients with high-grade gliomas. Radiat. Oncol. 2016, 11, 51. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.K.; Newman, V.L.; Berg, A.N.; MacVittie, T.J. Animal models for acute radiation syndrome drug discovery. Expert Opin. Drug Discov. 2015, 10, 497–517. [Google Scholar] [CrossRef] [PubMed]
- Azzam, E.I.; Jay-Gerin, J.P.; Pain, D. Ionizing radiation-induced metabolic oxidative stress and prolonged cell injury. Cancer Lett. 2012, 327, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Pannkuk, E.L.; Laiakis, E.C.; Gill, K.; Jain, S.; Mehta, K.; Nishita, D.; Bujold, K.; Bakke, J.; Gahagen, J.; Authier, S.; et al. Liquid chromatography—Mass spectrometry based metabolomics of nonhuman primates after 4 Gy total body radiation exposure: Global effects and targeted panels. J. Proteome Res. 2019, 18, 2260–2269. [Google Scholar] [CrossRef] [PubMed]
- Pannkuk, E.L.; Laiakis, E.C.; Authier, S.; Wong, K.; Fornace, A.J., Jr. Global metabolomic identification of longer-term dose dependent urinary biomarkers in nonhuman primates exposed to ionizing radiation. Radiat. Res. 2015, 184, 121–131. [Google Scholar]
- Pannkuk, E.L.; Laiakis, E.C.; Mak, T.D.; Astarita, G.; Authier, S.; Wong, K.; Fornace, A.J., Jr. A lipidomic and metabolomic serum signature from nonhuman primates exposed to ionizing radiation. Metabolomics 2016, 12, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Khamis, M.M.; Holt, T.; Awad, H.; El-Aneed, A.; Adamko, D.J. Comparative analysis of creatinine and osmolality as urine normalization strategies in targeted metabolomics for the differential diagnosis of asthma and COPD. Metabolomics 2018, 14, 115. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [PubMed]
- Bijlsma, S.; Bobeldijk, I.; Verheij, E.R.; Ramaker, R.; Kochhar, S.; Macdonald, I.A.; van Ommen, B.; Smilde, A.K. Large-scale human metabolomics studies: A strategy for data (pre-) processing and validation. Anal. Chem. 2006, 78, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Ramazzina, I.; Folli, C.; Secchi, A.; Berni, R.; Percudani, R. Completing the uric acid degradation pathway through phylogenetic comparison of whole genomes. Nat. Chem. Biol. 2006, 2, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Usuda, N.; Hayashi, S.; Fujiwara, S.; Noguchi, T.; Nagata, T.; Rao, M.S.; Alvares, K.; Reddy, J.K.; Yeldandi, A.V. Uric acid degrading enzymes, urate oxidase and allantoinase, are associated with different subcellular organelles in frog liver and kidney. J. Cell Sci. 1994, 107, 1073–1081. [Google Scholar] [PubMed]
- Kratzer, J.T.; Lanaspa, M.A.; Murphy, M.N.; Cicerchi, C.; Graves, C.L.; Tipton, P.A.; Ortlund, E.A.; Johnson, R.J.; Gaucher, E.A. Evolutionary history and metabolic insights of ancient mammalian uricases. Proc. Natl. Acad. Sci. USA 2014, 111, 3763–3768. [Google Scholar] [CrossRef] [PubMed]
- Yano, J.M.; Yu, K.; Donaldson, G.P.; Shastri, G.G.; Ann, P.; Ma, L.; Nagler, C.R.; Ismagilov, R.F.; Mazmanian, S.K.; Hsiao, E.Y. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell 2015, 161, 264–276. [Google Scholar] [CrossRef] [PubMed]
- Christenson, J.G.; Dairman, W.; Udenfriend, S. On the identity of DOPA decarboxylase and 5-hydroxytryptophan decarboxylase (immunological titration-aromatic L-amino acid decarboxylase-serotonin-dopamine-norepinephrine). Proc. Natl. Acad. Sci. USA 1972, 69, 343–347. [Google Scholar] [CrossRef]
- Kobayashi, S.; Nakamura, W.; Eto, H. Protective effect of 5-hydroxytryptophan against lethal doses of x-radiation in mice. Int. J. Radiat. Biol. Relat. Stud. Phys. Chem. Med. 1966, 11, 505–508. [Google Scholar] [CrossRef]
- Barnes, J.H.; Lowman, D.M. Relative radioprotective abilities of 5-hydroxytryptophan and 5-hydroxytryptamine. Int. J. Radiat. Biol. Relat. Stud. Phys. Chem. Med. 1968, 14, 87–88. [Google Scholar] [CrossRef] [PubMed]
- Randic, M.; Supek, Z. Urinary excretion of 5-hydroxyindolacetic acid after a single whole-body x-irradiation in normal and adrenalectomized rats. Int. J. Radiat. Biol. Relat. Stud. Phys. Chem. Med. 1961, 4, 151–153. [Google Scholar] [CrossRef] [PubMed]
- Grison, S.; Favé, G.; Maillot, M.; Manens, L.; Delissen, O.; Blanchardon, E.; Banzet, N.; Defoort, C.; Bott, R.; Dublineau, I.; et al. Metabolomics identifies a biological response to chronic low-dose natural uranium contamination in urine samples. Metabolomics 2013, 9, 1168–1180. [Google Scholar] [CrossRef]
- Deanovic, Z.; Supek, Z.; Randic, M. Relationship between the dose of whole-body x-irradiation and the urinary excretion of 5-hydroxyindoleacetic acid in rats. Int. J. Radiat. Biol. Relat. Stud. Phys. Chem. Med. 1963, 7, 569–574. [Google Scholar] [CrossRef]
- Ó Broin, P.; Vaitheesvaran, B.; Saha, S.; Hartil, K.; Chen, E.I.; Goldman, D.; Fleming, W.H.; Kurland, I.J.; Guha, C.; Golden, A. Intestinal microbiota-derived metabolomic blood plasma markers for prior radiation injury. Int. J. Radiat. Oncol. Biol. Phys. 2015, 91, 360–367. [Google Scholar] [CrossRef]
- Johnson, C.H.; Patterson, A.D.; Krausz, K.W.; Kalinich, J.F.; Tyburski, J.B.; Kang, D.W.; Luecke, H.; Gonzalez, F.J.; Blakely, W.F.; Idle, J.R. Radiation metabolomics. 5. Identification of urinary biomarkers of ionizing radiation exposure in nonhuman primates by mass spectrometry-based metabolomics. Radiat. Res. 2012, 178, 328–340. [Google Scholar] [CrossRef]
- Jones, J.W.; Tudor, G.; Bennett, A.; Farese, A.M.; Moroni, M.; Booth, C.; MacVittie, T.J.; Kane, M.A. Development and validation of a LC-MS/MS assay for quantitation of plasma citrulline for application to animal models of the acute radiation syndrome across multiple species. Anal. Bioanal. Chem. 2014, 406, 4663–4675. [Google Scholar] [CrossRef] [PubMed]
- Pannkuk, E.L.; Laiakis, E.C.; Authier, S.; Wong, K.; Fornace, A.J., Jr. Targeted metabolomics of nonhuman primate serum after exposure to ionizing radiation: Potential tools for high-throughput biodosimetry. RSC Adv. 2016, 6, 51192–51202. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Zheng, M.; Zhang, Y.; Fan, S.; Wang, C. Estimation value of plasma amino acid target analysis to the acute radiation injury early triage in the rat model. Metabolomics 2013, 9, 853–863. [Google Scholar] [CrossRef]
- Bujold, K.; Hauer-Jensen, M.; Donini, O.; Rumage, A.; Hartman, D.; Hendrickson, H.P.; Stamatopoulos, J.; Naraghi, H.; Pouliot, M.; Ascah, A.; et al. Citrulline as a biomarker for gastrointestinal-acute radiation syndrome: Species differences and experimental condition effects. Radiat. Res. 2016, 186, 71–78. [Google Scholar] [CrossRef]
- Cui, W.; Bennett, A.W.; Zhang, P.; Barrow, K.R.; Kearney, S.R.; Hankey, K.G.; Taylor-Howell, C.; Gibbs, A.M.; Smith, C.P.; MacVittie, T.J. A non-human primate model of radiation-induced cachexia. Sci. Rep. 2016, 6, 23612. [Google Scholar] [CrossRef] [PubMed]
- Laiakis, E.C.; Mak, T.D.; Anizan, S.; Amundson, S.A.; Barker, C.A.; Wolden, S.L.; Brenner, D.J.; Fornace, A.J., Jr. Development of a metabolomic radiation signature in urine from patients undergoing total body irradiation. Radiat. Res. 2014, 181, 350–361. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.W.; Alloush, J.; Sellamuthu, R.; Chua, H.L.; MacVittie, T.J.; Orschell, C.M.; Kane, M.A. Effect of sex on biomarker response in a mouse model of the hematopoietic acute radiation syndrome. Health Phys. 2019, 116, 484–502. [Google Scholar] [CrossRef] [PubMed]
- Simoni, R.E.; Gomes, L.N.; Scalco, F.B.; Oliveira, C.P.; Aquino Neto, F.R.; de Oliveira, M.L. Uric acid changes in urine and plasma: An effective tool in screening for purine inborn errors of metabolism and other pathological conditions. J. Inherit. Metab. Dis. 2007, 30, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Tyburski, J.B.; Patterson, A.D.; Krausz, K.W.; Slavik, J.; Fornace, A.J., Jr.; Gonzalez, F.J.; Idle, J.R. Radiation metabolomics. 2. Dose- and time-dependent urinary excretion of deaminated purines and pyrimidines after sublethal gamma-radiation exposure in mice. Radiat. Res. 2009, 172, 42–57. [Google Scholar] [CrossRef] [PubMed]
- Niwa, T.; Takeda, N.; Yoshizumi, H. RNA metabolism in uremic patients: Accumulation of modified ribonucleosides in uremic serum. Technical note. Kidney Int. 1998, 53, 1801–1806. [Google Scholar] [CrossRef] [PubMed]
- Jabs, C.M.; Sigurdsson, G.H.; Neglen, P. Plasma levels of high-energy compounds compared with severity of illness in critically ill patients in the intensive care unit. Surgery 1998, 124, 65–72. [Google Scholar] [CrossRef]
- Yin, H.; Xu, L.; Porter, N.A. Free radical lipid peroxidation: Mechanisms and analysis. Chem. Rev. 2011, 111, 5944–5972. [Google Scholar] [CrossRef] [PubMed]
- Tyurina, Y.Y.; Tyurin, V.A.; Kapralova, V.I.; Wasserloos, K.; Mosher, M.; Epperly, M.W.; Greenberger, J.S.; Pitt, B.R.; Kagan, V.E. Oxidative lipidomics of γ-radiation-induced lung injury: Mass spectrometric characterization of cardiolipin and phosphatidylserine peroxidation. Radiat. Res. 2011, 175, 610–621. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.L.; Jones, J.W.; Barrow, K.; Kieta, K.; Taylor-Howell, C.; Kearney, S.; Smith, C.P.; Gibbs, A.; Farese, A.M.; MacVittie, T.J.; et al. A MALDI-MSI approach to the characterization of radiation-induced lung injury and medical countermeasure development. Health Phys. 2015, 109, 466–478. [Google Scholar] [CrossRef]
- Goudarzi, M.; Weber, W.M.; Chung, J.; Doyle-Eisele, M.; Melo, D.R.; Mak, T.D.; Strawn, S.J.; Brenner, D.J.; Guilmette, R.; Fornace, A.J., Jr. Serum dyslipidemia is induced by internal exposure to strontium-90 in mice, lipidomic profiling using a data-independent liquid chromatography-mass spectrometry approach. J. Proteome Res. 2015, 14, 4039–4049. [Google Scholar] [CrossRef] [PubMed]
- Laiakis, E.C.; Strassburg, K.; Bogumil, R.; Lai, S.; Vreeken, R.J.; Hankemeier, T.; Langridge, J.; Plumb, R.S.; Fornace, A.J., Jr.; Astarita, G. Metabolic phenotyping reveals a lipid mediator response to ionizing radiation. J. Proteome Res. 2014, 13, 4143–4154. [Google Scholar] [CrossRef] [PubMed]
- Williamson, J.R.; Cooper, R.H. Regulation of the citric acid cycle in mammalian systems. FEBS Lett. 1980, 117, K73–K85. [Google Scholar] [CrossRef]
- Chen, C.; Brenner, D.J.; Brown, T.R. Identification of urinary biomarkers from x-irradiated mice using NMR spectroscopy. Radiat. Res. 2011, 175, 622–630. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.K.; Kulkarni, S.; Fatanmi, O.O.; Wise, S.Y.; Newman, V.L.; Romaine, P.L.; Hendrickson, H.; Gulani, J.; Ghosh, S.P.; Kumar, K.S.; et al. Radioprotective efficacy of gamma-tocotrienol in nonhuman primates. Radiat. Res. 2016, 185, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Farese, A.M.; Cohen, M.V.; Katz, B.P.; Smith, C.P.; Jackson, W.; Cohen, D.M.; MacVittie, T.J. A nonhuman primate model of the hematopoietic acute radiation syndrome plus medical management. Health Phys. 2012, 103, 367–382. [Google Scholar] [CrossRef] [PubMed]
- López, M.; Martín, M. Medical management of the acute radiation syndrome. Rep. Pract. Oncol. Radiother. 2011, 16, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Macià I Garau, M.; Lucas Calduch, A.; López, E.C. Radiobiology of the acute radiation syndrome. Rep. Pract. Oncol. Radiother. 2011, 16, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Vera, N.B.; Chen, Z.; Pannkuk, E.L.; Laiakis, E.C.; Fornace, A.J., Jr.; Erion, D.M.; Coy, S.L.; Pfefferkorn, J.A.; Vouros, P. Differential mobility spectrometry (DMS) reveals the elevation of urinary acetylcarnitine in non-human primates (NHPs) exposed to radiation. J. Mass Spectrom. 2018, 53, 548–559. [Google Scholar] [CrossRef]
- Chen, Z.; Coy, S.L.; Pannkuk, E.L.; Laiakis, E.C.; Fornace, A.J., Jr.; Vouros, P. Differential mobility spectrometry-mass spectrometry (DMS-MS) in radiation biodosimetry: Rapid and high-throughput quantitation of multiple radiation biomarkers in nonhuman primate urine. J. Am. Soc. Mass Spectrom. 2018, 29, 1650–1664. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhou, X.; Li, C.; Wu, J.; Kuo, J.E.; Wang, C. Assessment of early triage for acute radiation injury in rat model based on urinary amino acid target analysis. Mol. Biosyst. 2014, 10, 1441–1449. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
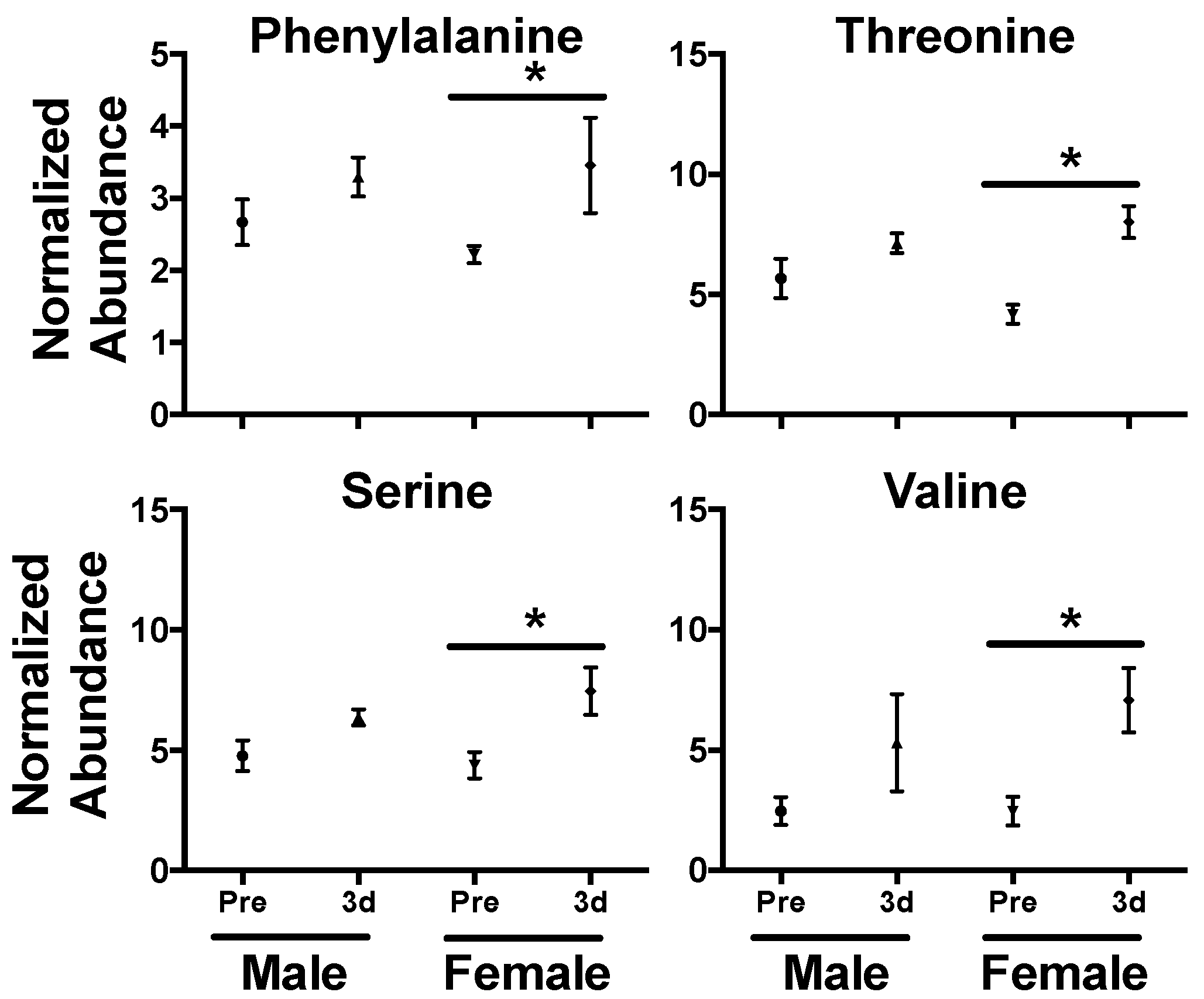{kind=link}
| Metabolite. | Parent ion (m/z) | Daughter ion (m/z) | Collision (eV) | Retention Time (min) |
|---|---|---|---|---|
| Pyruvic acid | 174 | 74.1 | 20 | 4.69 |
| Citric acid | 273 | 73.1 | 15 | 14.27 |
| Citric acid-d4 | 276 | 185 | 15 | 14.25 |
| Isocitric acid | 245 | 73.1 | 20 | 14.28 |
| cis-Aconitic acid | 229 | 147.1 | 15 | 13.41 |
| α-Ketoglutaric acid | 198 | 73.1 | 20 | 11.48 |
| Malic acid | 233 | 73.1 | 15 | 10.46 |
| Succinic acid | 247 | 147 | 15 | 8.15 |
| Fumaric acid | 245 | 73.1 | 20 | 8.56 |
| Biofluid | Metabolite | Retention Index | Unique Mass | P Value | No. TMS | HMDB ID |
|---|---|---|---|---|---|---|
| Urine | Allantoic acid | 662,102 | 331 | 0.020 | 1 | HMDB 01209 |
| 5-Hydroxyindoleacetic acid | 786,849 | 290 | 0.003 | 3 | HMDB 00763 | |
| Serum | Oleic acid | 787,976 | 117 | <0.001 | 1 | HMDB 00207 |
| Inosine | 906,197 | 73 | 0.002 | 4 | HMDB 00195 | |
| Leucine | 353,596 | 158 | 0.001 | 2 | HMDB 00687 | |
| Isoleucine | 366,901 | 158 | 0.005 | 2 | HMDB 00172 | |
| Valine | 320,280 | 144 | 0.002 | 2 | HMDB 00883 | |
| Serine | 405,133 | 73 | 0.012 | 3 | HMDB 00187 | |
| Threonine | 420,194 | 73 | 0.003 | 3 | HMDB 00167 | |
| Phenylalanine | 542,089 | 73 | 0.004 | 2 | HMDB 00159 |
| Biofluid | Metabolite | HMDB ID | 1 d LogFC | 3 d LogFC | 5 d LogFC | 7 d LogFC |
|---|---|---|---|---|---|---|
| Urine | Pyruvic acid | HMDB 00243 | −0.55 | −0.39 | −0.48 | −0.35 |
| Citric acid | HMDB 00094 | −0.02 | −0.58 | −0.24 | −0.27 | |
| Isocitric acid | HMDB 00193 | 0.12 | −0.49 | −0.25 | −0.20 | |
| cis-Aconitic acid | HMDB 00072 | −0.30 | −0.90 | −0.66 | −0.08 | |
| α-Ketoglutaric acid | HMDB 00208 | −0.17 | −1.02 | −0.65 | −0.10 | |
| Malic acid | HMDB 00744 | −0.28 | −0.60 | −0.43 | −0.28 | |
| Succinic acid | HMDB 00254 | −0.24 | −0.79 | −0.38 | −0.21 | |
| Fumaric acid | HMDB 00134 | −0.27 | −0.70 | −0.57 | −0.11 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pannkuk, E.L.; Laiakis, E.C.; Girgis, M.; Dowd, S.E.; Dhungana, S.; Nishita, D.; Bujold, K.; Bakke, J.; Gahagen, J.; Authier, S.; et al. Temporal Effects on Radiation Responses in Nonhuman Primates: Identification of Biofluid Small Molecule Signatures by Gas Chromatography–Mass Spectrometry Metabolomics. Metabolites 2019, 9, 98. https://doi.org/10.3390/metabo9050098
Pannkuk EL, Laiakis EC, Girgis M, Dowd SE, Dhungana S, Nishita D, Bujold K, Bakke J, Gahagen J, Authier S, et al. Temporal Effects on Radiation Responses in Nonhuman Primates: Identification of Biofluid Small Molecule Signatures by Gas Chromatography–Mass Spectrometry Metabolomics. Metabolites. 2019; 9(5):98. https://doi.org/10.3390/metabo9050098
Chicago/Turabian StylePannkuk, Evan L., Evagelia C. Laiakis, Michael Girgis, Sarah E. Dowd, Suraj Dhungana, Denise Nishita, Kim Bujold, James Bakke, Janet Gahagen, Simon Authier, and et al. 2019. "Temporal Effects on Radiation Responses in Nonhuman Primates: Identification of Biofluid Small Molecule Signatures by Gas Chromatography–Mass Spectrometry Metabolomics" Metabolites 9, no. 5: 98. https://doi.org/10.3390/metabo9050098
APA StylePannkuk, E. L., Laiakis, E. C., Girgis, M., Dowd, S. E., Dhungana, S., Nishita, D., Bujold, K., Bakke, J., Gahagen, J., Authier, S., Chang, P. Y., & Fornace, Jr., A. J. (2019). Temporal Effects on Radiation Responses in Nonhuman Primates: Identification of Biofluid Small Molecule Signatures by Gas Chromatography–Mass Spectrometry Metabolomics. Metabolites, 9(5), 98. https://doi.org/10.3390/metabo9050098