Untargeted Metabolomics Profiling of an 80.5 km Simulated Treadmill Ultramarathon


 and
and 
Abstract
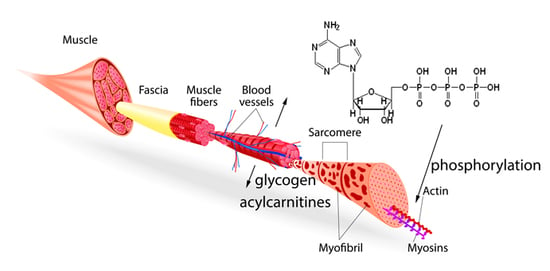
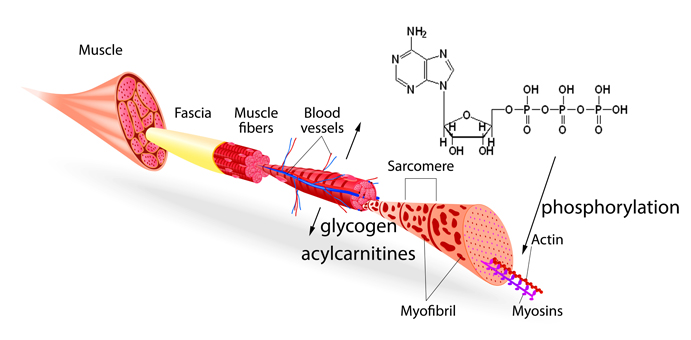
1. Introduction
2. Results
2.1. Physiological Response to the Ultramarathon
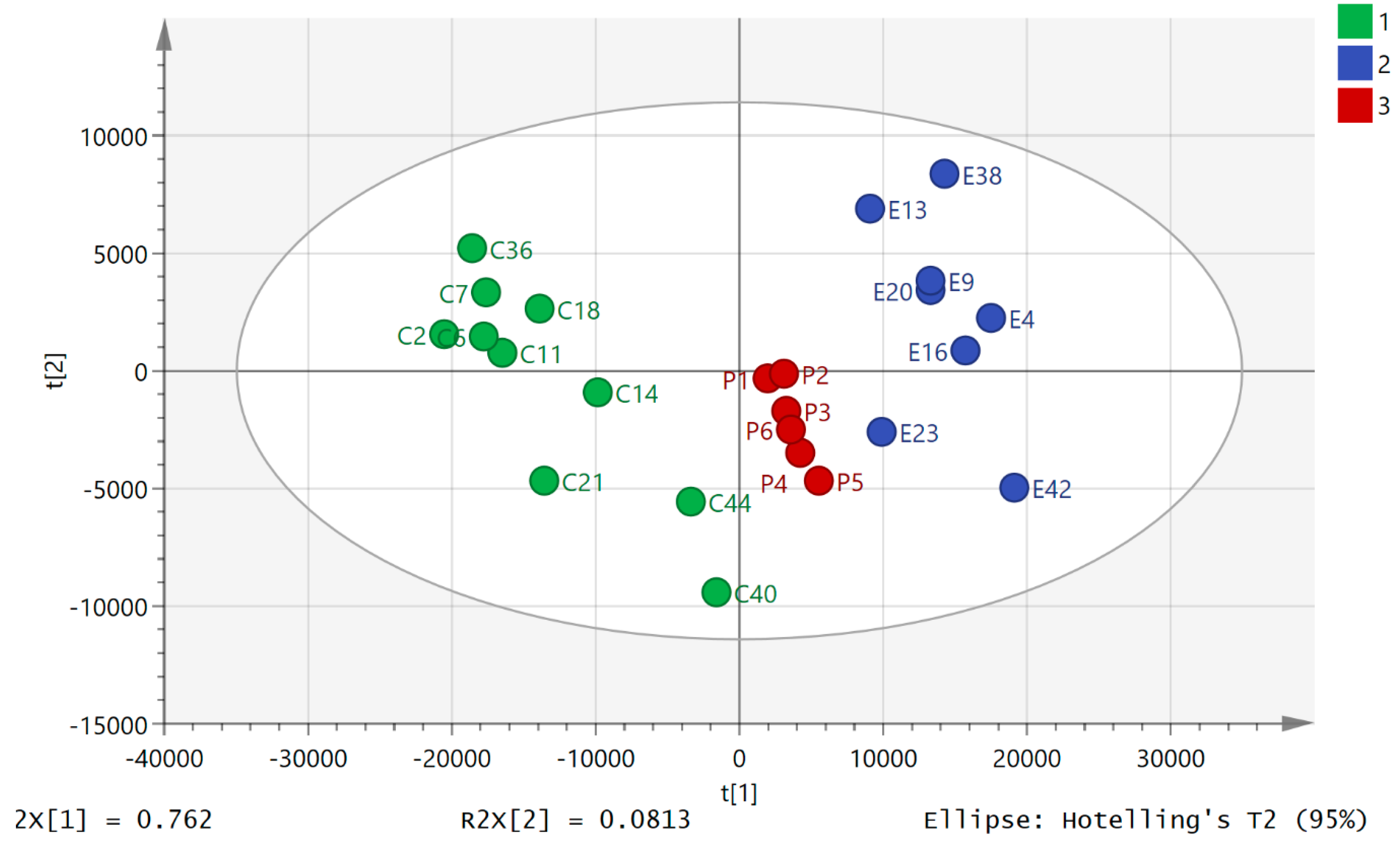
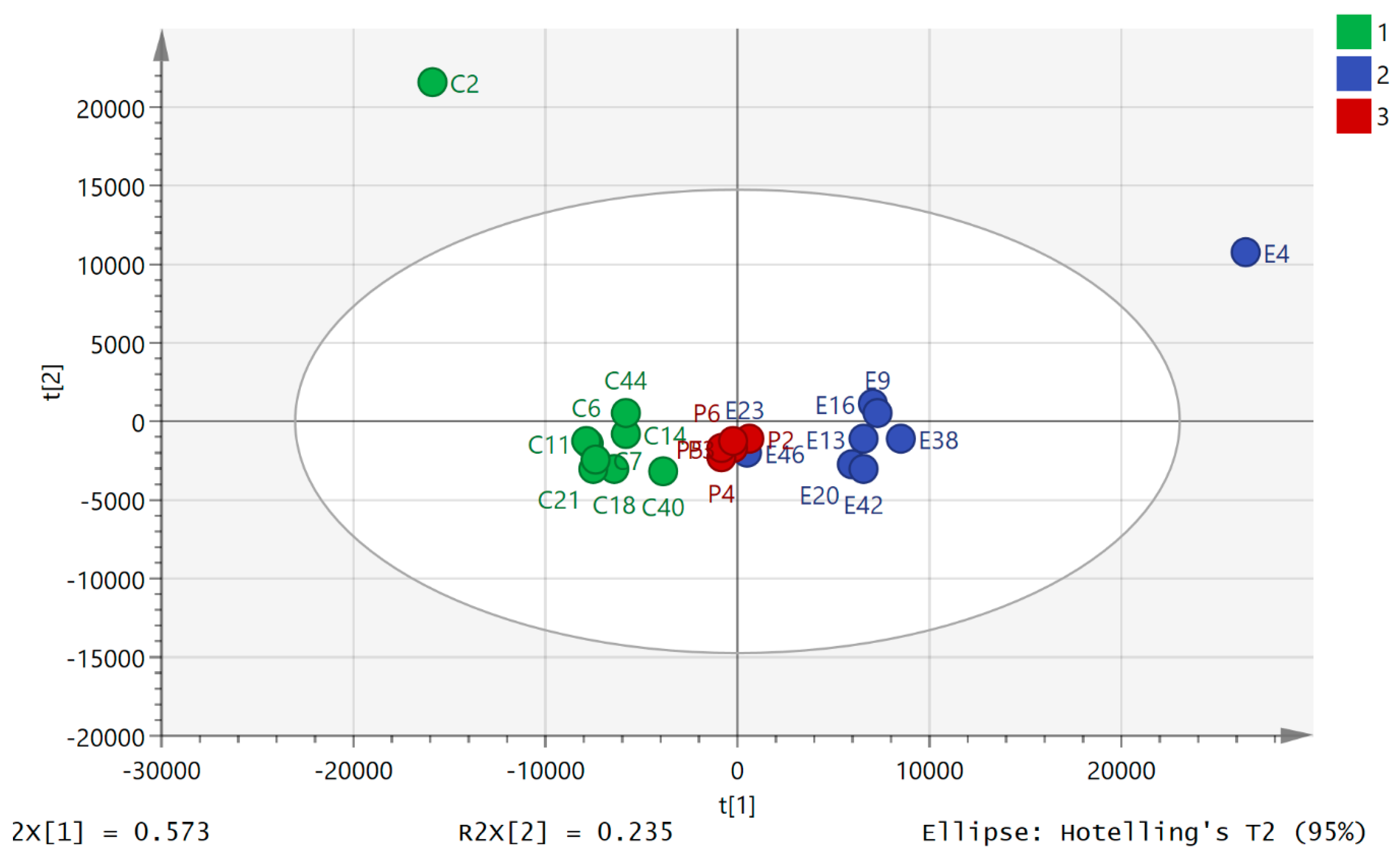
2.2. Variation of Metabolic Profile with Exercise
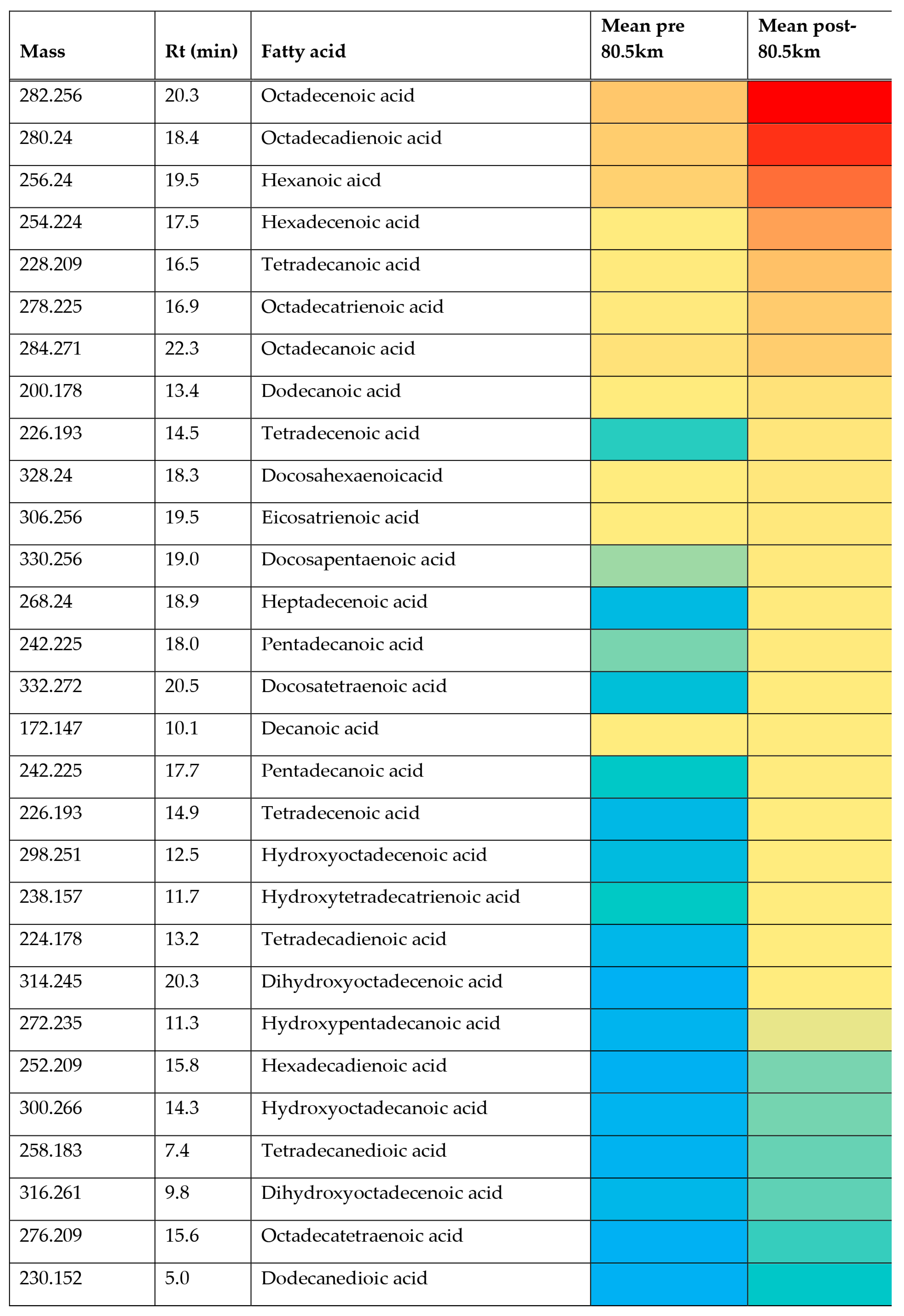
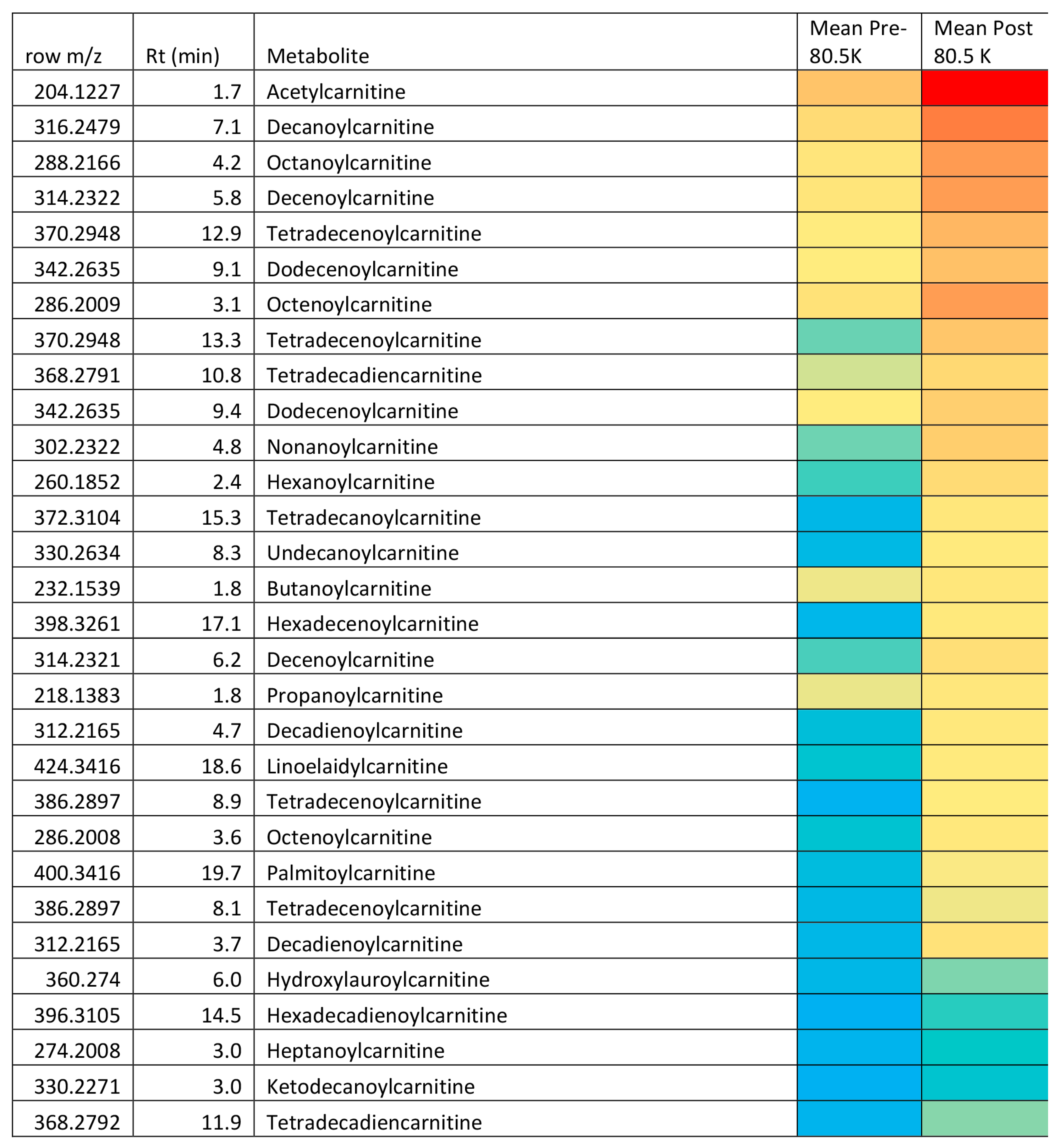
2.3. Univariate Comparisons
3. Discussion
4. Materials and Methods
4.1. Chemicals and Solvents
4.2. Participants
4.3. Experimental Design
4.4. Blood Sampling
4.5. Sample Preparation
4.6. LC-MS Conditions
4.7. Data Extraction and Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Harper, S.; Howse, K. An upper limit to human longevity? J. Popul. Ageing 2008, 1, 99–106. [Google Scholar] [CrossRef]
- Williams, P.T. Relationship of distance run per week to coronary heart disease risk factors in 8283 male runners: The national runners’ health study. Arch. Intern. Med. 1997, 157, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Trappe, S. Marathon runners. Sports Med. 2007, 37, 302–305. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, J.A.; Davis, C.S.; Mendias, C.L.; Brooks, S.V. The aging of elite male athletes: Age-related changes in performance and skeletal muscle structure and function. Clin. J. Sport Med. 2008, 18, 501–507. [Google Scholar] [CrossRef] [PubMed]
- Sarris, J.; O’Neil, A.; Coulson, C.E.; Schweitzer, I.; Berk, M. Lifestyle medicine for depression. BMC Psychiatry 2014, 14, 107. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Dengue: Guidelines for Diagnosis, Treatment, Prevention and Control; World Health Organization: Geneva, Switzerland, 2009. [Google Scholar]
- Williams, P.T. Lower prevalence of hypertension, hypercholesterolemia, and diabetes in marathoners. Med. Sci. Sports Exerc. 2009, 41, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, M.D.; Krishnan, E. Exercise behavior of ultramarathon runners: Baseline findings from the ultrastudy. J. Strength Cond. Res. 2013, 27, 2939–2945. [Google Scholar] [CrossRef] [PubMed]
- O’keefe, J.H.; Patil, H.R.; Lavie, C.J.; Magalski, A.; Vogel, R.A.; McCullough, P.A. Potential adverse cardiovascular effects from excessive endurance exercise. Mayo Clin. Proc. 2012, 87, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Denham, J.; Nelson, C.P.; O’Brien, B.J.; Nankervis, S.A.; Denniff, M.; Harvey, J.T.; Marques, F.Z.; Codd, V.; Zukowska-Szczechowska, E.; Samani, N.J. Longer leukocyte telomeres are associated with ultra-endurance exercise independent of cardiovascular risk factors. PLoS ONE 2013, 8, e69377. [Google Scholar] [CrossRef] [PubMed]
- Borghini, A.; Giardini, G.; Tonacci, A.; Mastorci, F.; Mercuri, A.; Sposta, S.M.; Moretti, S.; Andreassi, M.G.; Pratali, L. Chronic and acute effects of endurance training on telomere length. Mutagenesis 2015, 30, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, M.D.; Ong, J.C.; Wang, G. Historical analysis of participation in 161 km ultramarathons in North America. Int. J. Hist. Sport 2010, 27, 1877–1891. [Google Scholar] [CrossRef] [PubMed]
- Cejka, N.; Rüst, C.A.; Lepers, R.; Onywera, V.; Rosemann, T.; Knechtle, B. Participation and performance trends in 100-km ultra-marathons worldwide. J. Sports Sci. 2014, 32, 354–366. [Google Scholar] [CrossRef] [PubMed]
- Krouse, R.Z.; Ransdell, L.B.; Lucas, S.M.; Pritchard, M.E. Motivation, goal orientation, coaching, and training habits of women ultrarunners. J. Strength Cond. Res. 2011, 25, 2835–2842. [Google Scholar] [CrossRef] [PubMed]
- Rochat, N.; Hauw, D.; Philippe, R.A.; von Roten, F.C.; Seifert, L. Comparison of vitality states of finishers and withdrawers in trail running: An enactive and phenomenological perspective. PLoS ONE 2017, 12, e0173667. [Google Scholar] [CrossRef] [PubMed]
- Knechtle, B.; Rüst, C.A.; Rosemann, T.; Lepers, R. Age-related changes in 100-km ultra-marathon running performance. Age 2012, 34, 1033–1045. [Google Scholar] [CrossRef] [PubMed]
- Khodaee, M.; Spittler, J.; VanBaak, K.; Changstrom, B.; Hill, J. Effects of running an ultramarathon on cardiac, hematologic, and metabolic biomarkers. Int. J. Sports Med. 2015, 94, 867–871. [Google Scholar] [CrossRef] [PubMed]
- Shave, R.; Oxborough, D. Exercise-induced cardiac injury: Evidence from novel imaging techniques and highly sensitive cardiac troponin assays. Prog. Cardiovasc. Dis. 2012, 54, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Simpson, D.; Post, P.G.; Young, G.; Jensen, P.R. “It’s not about taking the easy road”: The experiences of ultramarathon runners. Sport Psychol. 2014, 28, 176–185. [Google Scholar] [CrossRef]
- Pearson, H. Physiology: Freaks of nature? Nature 2006, 444, 1000–1001. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Seals, D.R. Endurance exercise performance in masters athletes: Age-associated changes and underlying physiological mechanisms. J. Physiol. 2008, 586, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Noakes, T.D.; Lambert, E.V.; Lambert, M.I.; McArthur, P.S.; Myburgh, K.H.; Benade, A.S. Carbohydrate ingestion and muscle glycogen depletion during marathon and ultramarathon racing. Eur. J. Appl. Physiol. Occup. Physiol. 1988, 57, 482–489. [Google Scholar] [CrossRef] [PubMed]
- Ivy, J.L.; Goforth, H.W., Jr.; Damon, B.M.; McCauley, T.R.; Parsons, E.C.; Price, T.B. Early postexercise muscle glycogen recovery is enhanced with a carbohydrate-protein supplement. J. Appl. Physiol. 2002, 93, 1337–1344. [Google Scholar] [CrossRef] [PubMed]
- Waśkiewicz, Z.; Kłapcińska, B.; Sadowska-Krępa, E.; Czuba, M.; Kempa, K.; Kimsa, E.; Gerasimuk, D. Acute metabolic responses to a 24-h ultra-marathon race in male amateur runners. Eur. J. Appl. Physiol. 2012, 112, 1679–1688. [Google Scholar] [CrossRef] [PubMed]
- Muhsen Ali, A.; Burleigh, M.; Daskalaki, E.; Zhang, T.; Easton, C.; Watson, D.G. Metabolomic profiling of submaximal exercise at a standardised relative intensity in healthy adults. Metabolites 2016, 6, 9. [Google Scholar] [CrossRef] [PubMed]
- Sparling, P.B.; O’donnell, E.M.; Snow, T.K. The gender difference in distance running performance has plateaued: An analysis of world rankings from 1980 to 1996. Med. Sci. Sports Exerc. 1998, 30, 1725–1729. [Google Scholar] [CrossRef] [PubMed]
- Baker, A.B.; Tang, Y.Q. Aging performance for masters records in athletics, swimming, rowing, cycling, triathlon, and weightlifting. Exp. Aging Res. 2010, 36, 453–477. [Google Scholar] [CrossRef] [PubMed]
- Hunter, S.K.; Stevens, A.A.; Magennis, K.; Skelton, K.W.; Fauth, M. Is there a sex difference in the age of elite marathon runners? Med. Sci. Sports Exerc. 2011, 43, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Ransdell, L.B.; Vener, J.; Huberty, J. Masters athletes: An analysis of running, swimming and cycling performance by age and gender. J. Exerc. Sci. Fit. 2009, 7, S61–S73. [Google Scholar] [CrossRef]
- Knechtle, B.; Wirth, A.; Knechtle, P.; Zimmermann, K.; Kohler, G. Personal best marathon performance is associated with performance in a 24-h run and not anthropometry or training volume. Br. J. Sports Med. 2009, 43, 836–839. [Google Scholar] [CrossRef] [PubMed]
- Lepers, R.; Cattagni, T. Do older athletes reach limits in their performance during marathon running? Age 2012, 34, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Daskalaki, E.; Easton, C.; Watson, D.G. The application of metabolomic profiling to the effects of physical activity. Curr. Metab. 2014, 2, 233–263. [Google Scholar] [CrossRef]
- Carnethon, M.R.; Gulati, M.; Greenland, P. Prevalence and cardiovascular disease correlates of low cardiorespiratory fitness in adolescents and adults. JAMA 2005, 294, 2981–2988. [Google Scholar] [CrossRef] [PubMed]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.-M.; Fiehn, O.; Goodacre, R.; Griffin, J.L. Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar]
- Neal, C.M.; Hunter, A.M.; Brennan, L.; O’Sullivan, A.; Hamilton, D.L.; DeVito, G.; Galloway, S.D. Six weeks of a polarized training-intensity distribution leads to greater physiological and performance adaptations than a threshold model in trained cyclists. J. Appl. Physiol. 2013, 114, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Lustgarten, M.S.; Price, L.L.; Logvinenko, T.; Hatzis, C.; Padukone, N.; Reo, N.V.; Phillips, E.M.; Kirn, D.; Mills, J.; Fielding, R.A. Identification of serum analytes and metabolites associated with aerobic capacity. Eur. J. Appl. Physiol. 2013, 113, 1311–1320. [Google Scholar] [CrossRef] [PubMed]
- Huffman, K.M.; Koves, T.R.; Hubal, M.J.; Abouassi, H.; Beri, N.; Bateman, L.A.; Stevens, R.D.; Ilkayeva, O.R.; Hoffman, E.P.; Muoio, D.M. Metabolite signatures of exercise training in human skeletal muscle relate to mitochondrial remodelling and cardiometabolic fitness. Diabetologia 2014, 57, 2282–2295. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Hansen, J.; Zhao, X.; Chen, S.; Hoene, M.; Wang, X.-L.; Clemmesen, J.O.; Secher, N.H.; Häring, H.; Pedersen, B. Liver and muscle contribute differently to the plasma acylcarnitine pool during fasting and exercise in humans. J. Clin. Endocrinol. Metab. 2016, 101, 5044–5052. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Light, A.R.; Hoppel, C.L.; Campbell, C.; Chandler, C.J.; Burnett, D.J.; Souza, E.C.; Casazza, G.A.; Hughen, R.W.; Keim, N.L. Acylcarnitines as markers of exercise-associated fuel partitioning, xenometabolism, and potential signals to muscle afferent neurons. Exp. Physiol. 2017, 102, 48–69. [Google Scholar] [CrossRef] [PubMed]
- Volek, J.S.; Freidenreich, D.J.; Saenz, C.; Kunces, L.J.; Creighton, B.C.; Bartley, J.M.; Davitt, P.M.; Munoz, C.X.; Anderson, J.M.; Maresh, C.M. Metabolic characteristics of keto-adapted ultra-endurance runners. Metabolism 2016, 65, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Ginsburg, G.S.; Agil, A.; O’toole, M.; Rimm, E.; Douglas, P.S.; Rifai, N. Effects of a single bout of ultraendurance exercise on lipid levels and susceptibility of lipids to peroxidation in triathletes. JAMA 1996, 276, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Nieman, D.C.; Shanely, R.A.; Luo, B.; Meaney, M.P.; Dew, D.A.; Pappan, K.L. Metabolomics approach to assessing plasma 13-and 9-hydroxy-octadecadienoic acid and linoleic acid metabolite responses to 75-km cycling. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307, R68–R74. [Google Scholar] [CrossRef] [PubMed]
- Nieman, D.C.; Meaney, M.P.; John, C.S.; Knagge, K.J.; Chen, H. 9-and 13-hydroxy-octadecadienoic acids (9 + 13 hode) are inversely related to granulocyte colony stimulating factor and il-6 in runners after 2 h running. Brain Behav. Immun. 2016, 56, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Abeywardena, M.Y.; Head, R.J. Longchain n − 3 polyunsaturated fatty acids and blood vessel function. Cardiovasc. Res. 2001, 52, 361–371. [Google Scholar] [CrossRef]
- Turner, J.E.; Bennett, S.J.; Bosch, J.A.; Griffiths, H.R.; Aldred, S. Ultra-Endurance Exercise: Unanswered Questions in Redox Biology and Immunology; Portland Press Limited: London, UK, 2014. [Google Scholar]
- Yamada, H.; Umemoto, T.; Kakei, M.; Momomura, S.-I.; Kawakami, M.; Ishikawa, S.-E.; Hara, K. Eicosapentaenoic acid shows anti-inflammatory effect via gpr120 in 3t3-l1 adipocytes and attenuates adipose tissue inflammation in diet-induced obese mice. Nutr. Metab. 2017, 14, 33. [Google Scholar] [CrossRef] [PubMed]
- Mrakic-Sposta, S.; Gussoni, M.; Moretti, S.; Pratali, L.; Giardini, G.; Tacchini, P.; Dellanoce, C.; Tonacci, A.; Mastorci, F.; Borghini, A. Effects of mountain ultra-marathon running on ros production and oxidative damage by micro-invasive analytic techniques. PLoS ONE 2015, 10, e0141780. [Google Scholar] [CrossRef] [PubMed]
- Wanders, R.J.; Waterham, H.R. Biochemistry of mammalian peroxisomes revisited. Annu. Rev. Biochem. 2006, 75, 295–332. [Google Scholar] [CrossRef] [PubMed]
- Zipper, J. Proliferation of myocardial peroxisomes caused by several agents and conditions. J. Mol. Cell. Cardiol. 1997, 29, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, R.R.; Zammit, V.A. Carnitine acyltransferases and their influence on CoA pools in health and disease. Mol. Asp. Med. 2004, 25, 475–493. [Google Scholar] [CrossRef] [PubMed]
- Reddy, J.K.; Mannaerts, G.P. Peroxisomal lipid metabolism. Annu. Rev. Nutr. 1994, 14, 343–370. [Google Scholar] [CrossRef] [PubMed]
- Thomas, A.W.; Davies, N.A.; Moir, H.; Watkeys, L.; Ruffino, J.S.; Isa, S.A.; Butcher, L.R.; Hughes, M.G.; Morris, K.; Webb, R. Exercise-associated generation of pparγ ligands activates pparγ signaling events and upregulates genes related to lipid metabolism. J. Appl. Physiol. 2012, 112, 806–815. [Google Scholar] [CrossRef] [PubMed]
- Harber, M.P.; Crane, J.D.; Dickinson, J.M.; Jemiolo, B.; Raue, U.; Trappe, T.A.; Trappe, S.W. Protein synthesis and the expression of growth-related genes are altered by running in human vastus lateralis and soleus muscles. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R708–R714. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.K.; Dickinson, J.M.; Timmerman, K.L.; Drummond, M.J.; Reidy, P.T.; Fry, C.S.; Gundermann, D.M.; Rasmussen, B.B. Exercise, amino acids and aging in the control of human muscle protein synthesis. Med. Sci. Sports Exerc. 2011, 43, 2249–2258. [Google Scholar] [CrossRef] [PubMed]
- Gatti, R.; Cappellin, E.; Zecchin, B.; Antonelli, G.; Spinella, P.; Mantero, F.; De Palo, E.F. Urinary high performance reverse phase chromatography cortisol and cortisone analyses before and at the end of a race in elite cyclists. J. Chromatogr. B 2005, 824, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Dovio, A.; Roveda, E.; Sciolla, C.; Montaruli, A.; Raffaelli, A.; Saba, A.; Calogiuri, G.; De Francia, S.; Borrione, P.; Salvadori, P. Intense physical exercise increases systemic 11β-hydroxysteroid dehydrogenase type 1 activity in healthy adult subjects. Eur. J. Appl. Physiol. 2010, 108, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Zieliński, J.; Krasińska, B.; Kusy, K. Hypoxanthine as a predictor of performance in highly trained athletes. Int. J. Sports Med. 2013, 34, 1079–1086. [Google Scholar] [CrossRef] [PubMed]
- Stathis, C.G.; Carey, M.F.; Hayes, A.; Garnham, A.P.; Snow, R.J. Sprint training reduces urinary purine loss following intense exercise in humans. Appl. Physiol. Nutr. Metab. 2006, 31, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Subudhi, A.W.; Davis, S.L.; Kipp, R.W.; Askew, E.W. Antioxidant status and oxidative stress in elite alpine ski racers. Int. J. Sport Nutr. Exerc. Metab. 2001, 11, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Watson, D.G.; Wang, L.; Westrop, G.D.; Coombs, G.H.; Zhang, T. Evaluation of mobile phase characteristics on three zwitterionic columns in hydrophilic interaction liquid chromatography mode for liquid chromatography-high resolution mass spectrometry based untargeted metabolite profiling of leishmania parasites. J. Chromatogr. A 2014, 1362, 168–179. [Google Scholar] [CrossRef] [PubMed]
- British Pharamacopoeia 2013; The Stationary Office: London, UK, 2012.
- Scheltema, R.A.; Jankevics, A.; Jansen, R.C.; Swertz, M.A.; Breitling, R. Peakml/mzmatch: A file format, Java library, R library, and tool-chain for mass spectrometry data analysis. Anal. Chem. 2011, 83, 2786–2793. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mode | Mass | RT (min) | Metabolite | Ratio [Post-80.5 km/Pre-80.5 km] | p-Value |
|---|---|---|---|---|---|
| Amino acids and their metabolites | |||||
| + | 75.032 | 15.4 | * Glycine | 0.510 | <0.001 |
| + | 89.048 | 14.4 | * Alanine | 0.603 | 0.012 |
| + | 103.063 | 13.4 | * 3-Amino-isobutanoate | 0.392 | <0.001 |
| + | 105.043 | 15.7 | * Serine | 0.512 | <0.001 |
| + | 111.032 | 9.5 | Pyrrole-2-carboxylate | 0.413 | <0.001 |
| + | 115.063 | 12.4 | * Proline | 0.420 | <0.001 |
| + | 116.047 | 1.7 | Oxopentanoic acid | 0.819 | <0.001 |
| + | 117.054 | 15.5 | Guanidinoacetate | 0.627 | 0.001 |
| + | 117.079 | 12.1 | * Valine | 0.447 | <0.001 |
| + | 117.079 | 10.8 | * Betaine | 0.505 | <0.001 |
| - | 118.063 | 1.8 | Hydroxypentanoate | 1.393 | <0.001 |
| + | 119.058 | 14.4 | * Threonine | 0.217 | <0.001 |
| + | 125.015 | 15.4 | * Taurine | 0.565 | 0.001 |
| + | 129.043 | 14.1 | 5-Oxoproline | 0.352 | <0.001 |
| + | 131.058 | 14.1 | Hydroxyproline | 0.361 | <0.005 |
| + | 131.095 | 10.3 | * Leucine | 0.455 | <0.001 |
| + | 131.095 | 10.8 | * Isoleucine | 0.430 | <0.001 |
| + | 132.079 | 2.1 | Hydroxyhexanoic acid ‡ | 2.237 | 0.004 |
| + | 132.053 | 15.2 | * Asparagine | 0.465 | <0.001 |
| + | 132.090 | 22.4 | * Ornithine | 0.545 | 0.003 |
| + | 138.043 | 8.8 | * Urocanate | 0.626 | 0.019 |
| + | 146.069 | 14.8 | * Glutamine | 0.710 | <0.001 |
| + | 146.106 | 23.8 | * Lysine | 0.369 | <0.006 |
| + | 147.053 | 11.2 | * Glutamate | 0.528 | <0.001 |
| + | 149.051 | 11.2 | * Methionine | 0.609 | <0.003 |
| - | 154.038 | 11.7 | Imidazol-5-yl-pyruvate | 0.469 | <0.001 |
| - | 159.068 | 8.1 | Indole-3-acetaldehyde | 0.432 | 0.001 |
| + | 161.069 | 9.9 | O-Acetylhomoserine | 0.524 | <0.001 |
| + | 174.112 | 25.4 | * Arginine | 0.387 | <0.003 |
| + | 175.096 | 15.6 | * Citrulline | 0.673 | 0.047 |
| + | 181.074 | 12.8 | * Tyrosine | 0.761 | 0.016 |
| - | 182.058 | 9.4 | Hydroxyphenyllactate | 0.541 | 0.002 |
| + | 188.116 | 16.2 | N6-Acetyl-l-lysine | 0.233 | 0.054 |
| + | 189.043 | 6.4 | Kynurenate | 2.322 | 0.001 |
| + | 204.090 | 11.1 | * l-Tryptophan | 0.539 | <0.001 |
| + | 208.085 | 10.2 | Formylhydroxykynurenamine | 0.668 | 0.004 |
| - | 219.053 | 4.9 | Hydroxyindolepyruvate | 5.131 | 0.010 |
| Acylcarnitines | |||||
| + | 204.123 | 10.3 | * Acetylcarnitine | 3.353 | <0.001 |
| 218.138 | 9.1 | Propanoylcarnitine | 1.420 | 0.042 | |
| + | 232.154 | 7.9 | Butanoylcarnitine | 1.775 | 0.010 |
| + | 258.170 | 2.1 | Hexenoylcarnitine ‡ | 6.350 | 0.002 |
| + | 260.185 | 2.4 | * Hexanoylcarnitine isomer ‡ | 9.640 | 0.011 |
| 260.185 | 2.9 | Hexanoylcarnitine isomer ‡ | 13.091 | 0.045 | |
| + | 274.201 | 3.0 | Heptanoylcarnitine ‡ | 5.685 | 0.013 |
| + | 286.201 | 3.6 | Octenoylcarnitine ‡ | 6.009 | 0.003 |
| + | 286.201 | 3.1 | Octenoylcarnitine ‡ | 5.184 | 0.001 |
| + | 288.217 | 4.2 | * Octanoylcarnitine ‡ | 7.119 | 0.004 |
| 302.232 | 4.8 | Nonanoykcarnitine ‡ | 14.587 | 0.001 | |
| + | 312.217 | 4.7 | Decadienoylcarnitine ‡ | 7.016 | 0.001 |
| + | 312.217 | 3.7 | Decadienoylcarnitine ‡ | 16.727 | 0.102 |
| + | 314.232 | 6.2 | Decenoylcarnitine ‡ | 7.186 | 0.039 |
| + | 314.232 | 5.8 | Decenoylcarnitine ‡ | 6.285 | 0.004 |
| + | 316.248 | 7.1 | * Decanoylcarnitine ‡ | 5.017 | 0.005 |
| 330.227 | 4.4 | Keto-decanoylcarnitine ‡ | 13.121 | 0.000 | |
| + | 330.227 | 3.0 | Keto-decanoylcarnitine ‡ | 7.719 | 0.003 |
| 330.263 | 8.3 | Dimethylnonanoylcarnitine ‡ | 11.088 | 0.002 | |
| + | 342.264 | 9.4 | Dodecenoylcarnitine ‡ | 6.439 | 0.089 |
| + | 342.264 | 9.1 | * Dodecenoylcarnitine ‡ | 8.849 | 0.004 |
| + | 360.274 | 6.0 | Hydroxydodecanoylcarnitine ‡ | 4.825 | 0.003 |
| + | 368.279 | 10.8 | Tetradecadiencarnitine isomer ‡ | 5.659 | 0.022 |
| + | 368.279 | 9.5 | Tetradecadiencarnitine isomer ‡ | 24.743 | 0.012 |
| + | 368.279 | 9.9 | Tetradecadiencarnitine isomer ‡ | 19.098 | 0.055 |
| + | 368.279 | 11.9 | Tetradecadiencarnitine isomer ‡ | 9.195 | 0.031 |
| + | 370.295 | 13.3 | Tetradecenoylcarnitine isomer ‡ | 16.422 | 0.070 |
| + | 370.295 | 12.9 | Tetradecenoylcarnitine isomer ‡ | 9.253 | 0.004 |
| + | 372.310 | 15.3 | * Tetradecanoylcarnitine ‡ | 18.265 | 0.007 |
| + | 384.274 | 6.6 | Hydroxytetradecadiencarnitine ‡ | 11.908 | 0.001 |
| + | 386.290 | 8.1 | Hydroxytetradecenoylcarnitine ‡ | 6.193 | 0.007 |
| + | 386.290 | 8.9 | Hydroxytetradecenoylcarnitine ‡ | 27.813 | 0.003 |
| + | 388.305 | 9.4 | Hydroxymyristoylcarnitine ‡ | 4.245 | 0.006 |
| + | 396.310 | 15.2 | Hexadecadienoylcarnitine ‡ | 90.958 | 0.149 |
| + | 396.311 | 14.5 | Hexadecadienoylcarnitine ‡ | 17.816 | 0.016 |
| + | 398.326 | 17.1 | Hexadecenoylcarnitine ‡ | 14.097 | 0.011 |
| + | 400.342 | 19.7 | Palmitoylcarnitine ‡ | 4.618 | 0.089 |
| + | 412.305 | 9.3 | Hydroxyhexadecadienoylcarnitine ‡ | 6.590 | 0.003 |
| + | 414.321 | 11.1 | Hydroxyhexadecenoylcarnitine ‡ | 35.292 | 0.003 |
| + | 424.342 | 18.6 | Octadecadienoylcarnitine ‡ | 3.955 | 0.048 |
| + | 424.342 | 19.3 | Octadecadienoylcarnitine ‡ | 6.043 | 0.121 |
| + | 430.316 | 8.1 | Hexadecanedioicacidmonocarnitineester ‡ | 114,475.436 | 0.015 |
| Fatty acids | |||||
| - | 172.147 | 10.0 | Decanoic acid ‡ | 1.909 | 0.034 |
| - | 196.146 | 10.1 | Dodecadienoic acid ‡ | 5.989 | 0.001 |
| - | 200.178 | 13.4 | Dodecanoic acid | 4.342 | 0.009 |
| - | 202.120 | 3.2 | Decanedioic acid ‡ | 6.045 | 0.004 |
| - | 210.126 | 9.3 | Hydroxydodecatrienoic acid ‡ | 4.709 | 0.001 |
| - | 212.178 | 13.3 | Tridecenoic acid ‡ | 13.224 | 0.006 |
| - | 224.178 | 13.2 | Tetradecadienoic acid ‡ | 10.003 | 0.013 |
| - | 226.193 | 14.5 | Tetradecenoic acid isomer ‡ | 25.065 | 0.004 |
| - | 226.193 | 14.9 | Tetradecenoic acid isomer ‡ | 14.409 | 0.020 |
| - | 230.152 | 5.0 | Dodecanedioic acid ‡ | 9.432 | 0.014 |
| - | 240.173 | 8.2 | Hydroxytetradecadienoic acid ‡ | 11.109 | 0.002 |
| - | 240.209 | 16.5 | Pentadecenoic acid ‡ | 3.192 | 0.007 |
| - | 242.188 | 11.4 | Hydroxytetradecadienoic acid ‡ | 4.066 | 0.001 |
| - | 244.204 | 8.2 | Hydroxytetradecanoic acid isomer ‡ | 11.109 | 0.002 |
| - | 244.204 | 9.3 | Hydroxytetradecanoic acid isomer ‡ | 3.581 | 0.000 |
| - | 252.209 | 15.8 | * Hexadecadienoicacid isomer ‡ | 10.174 | 0.059 |
| - | 252.209 | 16.3 | * Hexadecadienoicacid isomer ‡ | 13.108 | 0.041 |
| - | 254.224 | 17.5 | * Palmitoleic acid ‡ | 38.719 | 0.006 |
| - | 258.183 | 7.4 | Tetradecanedioic acid ‡ | 7.206 | 0.006 |
| - | 266.188 | 13.6 | Hydroxyhexadecatrienoic acid ‡ | 8.977 | 0.014 |
| - | 268.204 | 11.3 | Hydroxyhexadecadienoic acid | 3.355 | 0.007 |
| - | 268.240 | 18.9 | Heptadecenoic acid ‡ | 29.923 | 0.004 |
| - | 270.220 | 12.1 | Hydroxyhexadecenoic acid isomer ‡ | 3.553 | 0.004 |
| - | 270.220 | 17.5 | Hydroxyhexadecenoic acid ‡ | 8.969 | 0.001 |
| - | 272.235 | 11.3 | Hydroxyhexadecanoic acid ‡ | 8.969 | 0.001 |
| - | 276.209 | 15.6 | * Octadecatetraenoic acid ‡ | 10.190 | 0.067 |
| - | 278.225 | 16.9 | * Linolenic acid ‡ | 8.511 | 0.003 |
| - | 280.240 | 18.4 | * Linoleic acid ‡ | 5.769 | 0.008 |
| - | 282.256 | 20.3 | * Oleic acid ‡ | 6.231 | 0.000 |
| - | 284.199 | 9.1 | Dihydroxyhexadecadienoic acid ‡ | 2.897 | 0.001 |
| - | 286.214 | 10.5 | Dihydroxyhexdecenoic acid ‡ | 16.426 | 0.001 |
| - | 296.235 | 14.0 | Hydroxyoctadecadienoic acid ‡ | 3.145 | 0.024 |
| - | 300.266 | 14.3 | Hydroxyoctdecanoic acid ‡ | 6.618 | 0.015 |
| - | 316.261 | 9.8 | Dihydroxyoctadecanoic acid ‡ | 4.040 | 0.002 |
| - | 327.241 | 7.0 | Nitrooctadecenoic acid ‡ | 10.453 | <0.001 |
| - | 328.240 | 18.3 | * Docosahexaenoicacid ‡ | 4.266 | 0.022 |
| - | 330.256 | 19.0 | * Docosapentaenoic acid ‡ | 9.179 | 0.003 |
| - | 332.272 | 20.5 | Docosatetraenoic acid ‡ | 14.588 | 0.002 |
| Steroids | |||||
| - | 362.209 | 4.5 | Hydrocortisone | 1.787 | 0.014 |
| - | 364.225 | 5.0 | Urocortisone | 3.243 | 0.003 |
| - | 376.298 | 3.9 | Hydroxycholanate | 0.315 | 0.004 |
| - | 392.293 | 4.3 | Deoxycholanoic acid | 0.361 | 0.026 |
| - | 449.314 | 4.3 | Chenodeoxyglycocholate | 0.162 | <0.001 |
| - | 465.309 | 4.9 | * Glycocholate | 0.174 | 0.003 |
| - | 515.291 | 4.5 | Taurocholate | 0.275 | 0.039 |
| - | 568.324 | 7.3 | Chenodeoxycholic acid glucuronide | 0.311 | <0.001 |
| - | 612.387 | 4.5 | Cholestane—tetrol-glucuronide | 0.443 | 0.001 |
| Miscellaneous | |||||
| + | 136.039 | 9.8 | * Hypoxanthine | 1.917 | 0.003 |
| - | 244.069 | 9.5 | * Uridine | 0.420 | <0.001 |
| - | 244.070 | 11.7 | Pseudouridine | 0.416 | <0.001 |
| + | 136.064 | 23.7 | * 1-Methylnicotinamide | 0.226 | 0.090 |
| - | 164.069 | 11.8 | Rhamnose | 0.348 | <0.001 |
| + | 179.079 | 10.8 | Galactosamine | 0.181 | <0.001 |
| - | 180.064 | 14.1 | Hexose | 0.447 | <0.001 |
| + | 214.132 | 9.4 | Dethiobiotin | 1.517 | 0.002 |
| - | 416.366 | 3.4 | gamma-Tocopherol | 0.529 | <0.001 |
| - | 430.381 | 3.4 | Alpha-Tocopherol | 0.509 | <0.001 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Howe, C.C.F.; Alshehri, A.; Muggeridge, D.; Mullen, A.B.; Boyd, M.; Spendiff, O.; Moir, H.J.; Watson, D.G. Untargeted Metabolomics Profiling of an 80.5 km Simulated Treadmill Ultramarathon. Metabolites 2018, 8, 14. https://doi.org/10.3390/metabo8010014
Howe CCF, Alshehri A, Muggeridge D, Mullen AB, Boyd M, Spendiff O, Moir HJ, Watson DG. Untargeted Metabolomics Profiling of an 80.5 km Simulated Treadmill Ultramarathon. Metabolites. 2018; 8(1):14. https://doi.org/10.3390/metabo8010014
Chicago/Turabian StyleHowe, Christopher C. F., Ahmed Alshehri, David Muggeridge, Alexander B. Mullen, Marie Boyd, Owen Spendiff, Hannah J. Moir, and David G. Watson. 2018. "Untargeted Metabolomics Profiling of an 80.5 km Simulated Treadmill Ultramarathon" Metabolites 8, no. 1: 14. https://doi.org/10.3390/metabo8010014
APA StyleHowe, C. C. F., Alshehri, A., Muggeridge, D., Mullen, A. B., Boyd, M., Spendiff, O., Moir, H. J., & Watson, D. G. (2018). Untargeted Metabolomics Profiling of an 80.5 km Simulated Treadmill Ultramarathon. Metabolites, 8(1), 14. https://doi.org/10.3390/metabo8010014