Investigation of Phenolic Acids in Suspension Cultures of Vitis vinifera Stimulated with Indanoyl-Isoleucine, N-Linolenoyl-L-Glutamine, Malonyl Coenzyme A and Insect Saliva


Abstract
:1. Introduction
2. Results and Discussion
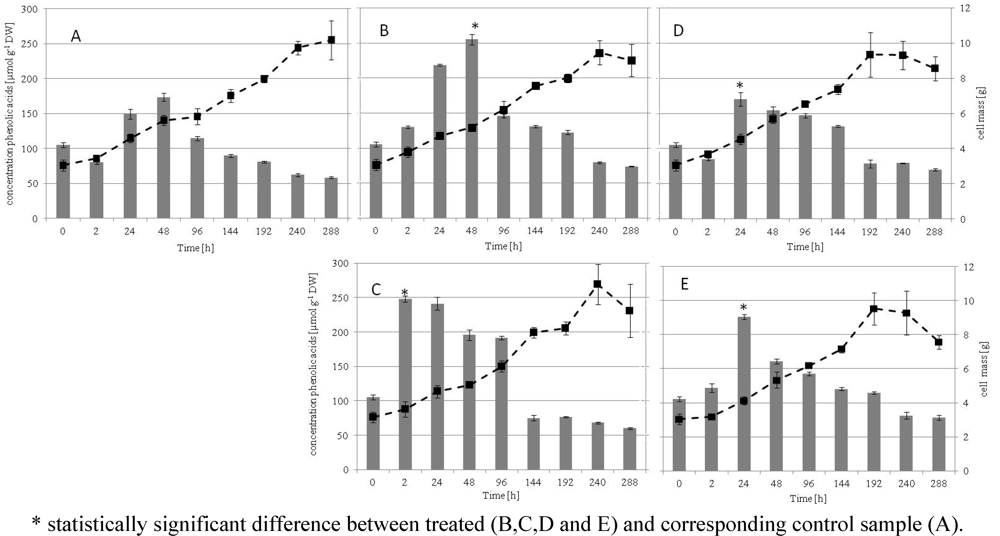
2.1. Growth Kinetics, Phenolic Acids of the Culture and HPLC Analysis
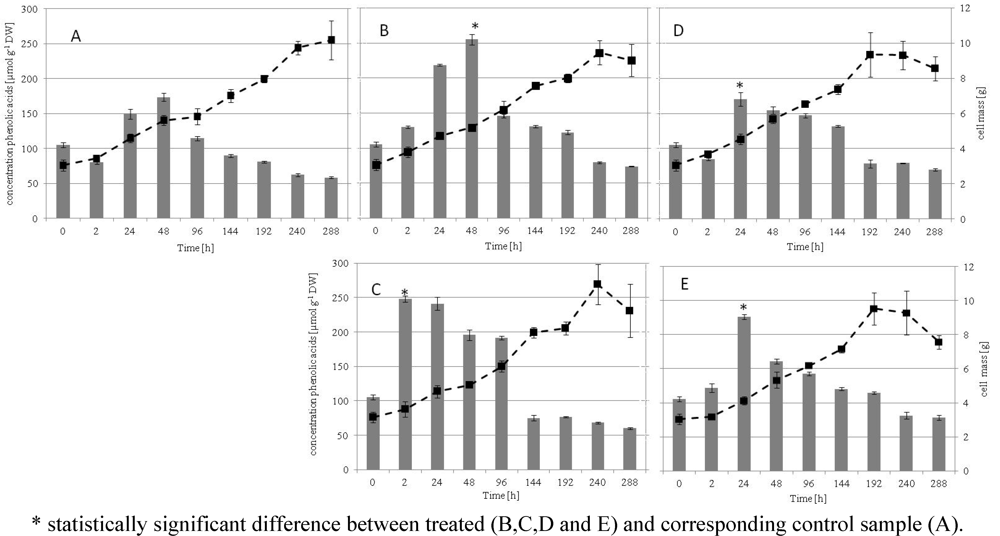
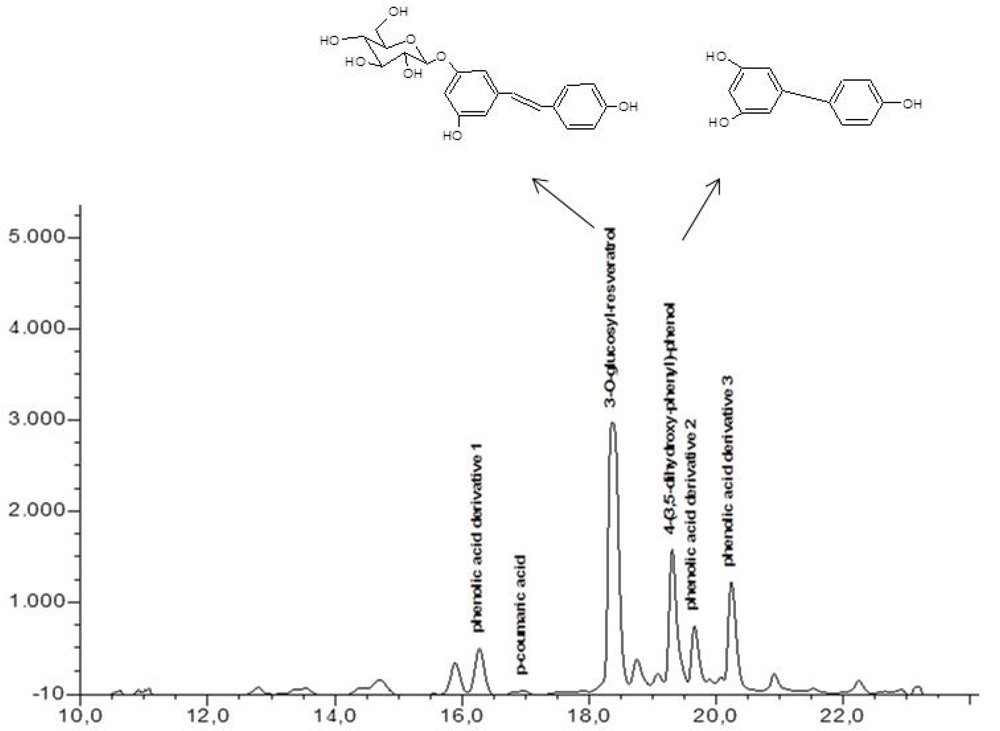
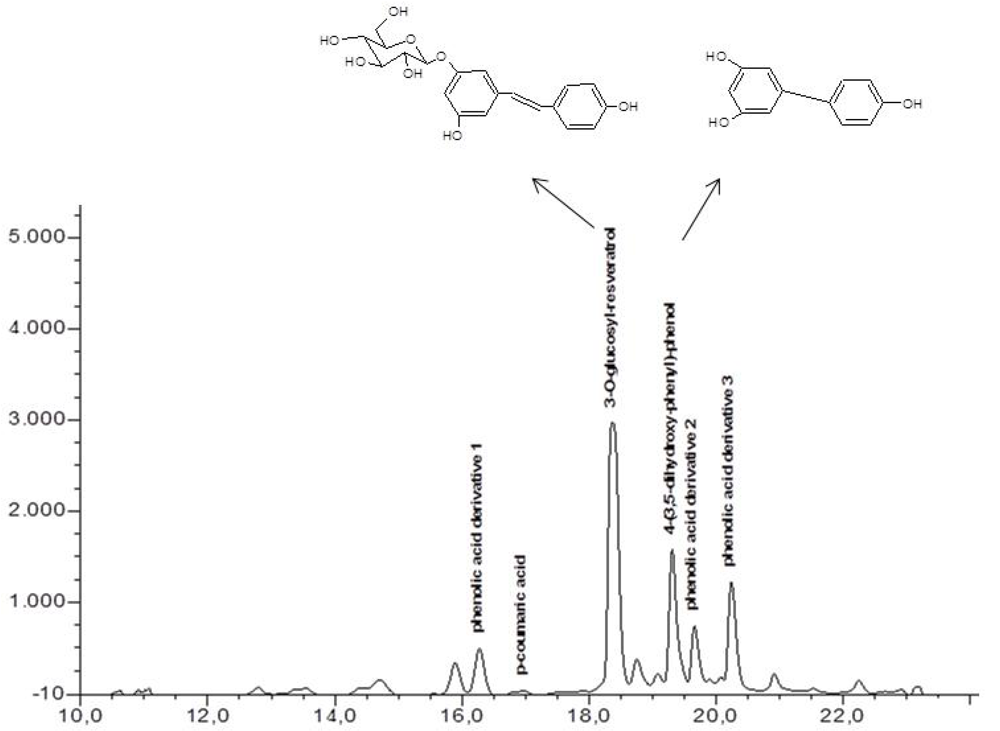
2.2. Chemical Analysis of in Vitro Grape Cells with HPLC
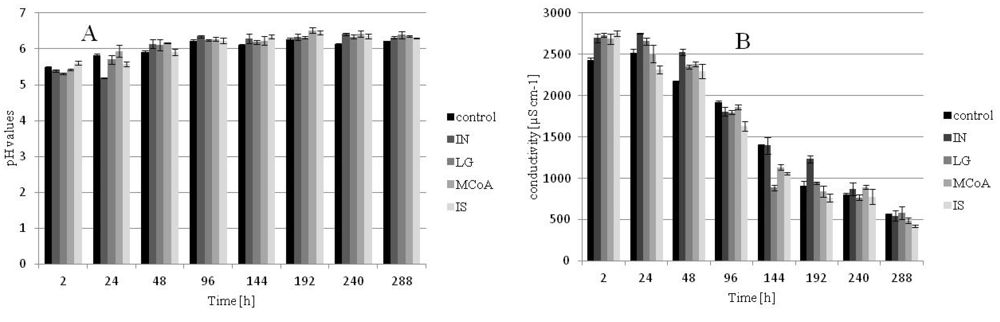
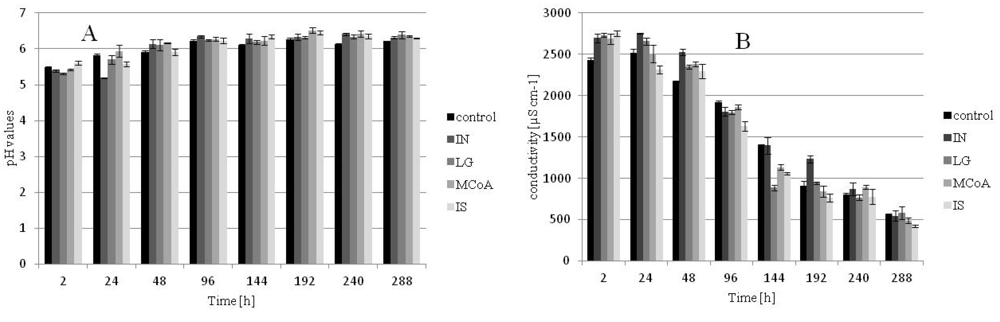
2.3. Effects on the pH and Conductivity
2.4. Influence of Experimental Parameters on Phenolic Acid Synthesis

{kind=link}
{kind=link}
{kind=link}
| Parameter | DF | Mean Square | F Value | p-value |
|---|---|---|---|---|
| Treatment | 5 | 7453.09 | 146.06 | <0.0001 |
| Time (harvesting time) | 7 | 35004.10 | 687.60 | <0.0001 |
| pH | 1 | 564.18 | 11.06 | 0.0013 |
| Fresh weight | 1 | 223.30 | 4.38 | 0.0396 |
| Treatment * Time | 28 | 2955.58 | 57.92 | <0.0001 |
3. Materials and methods
3.1. Cultivation and Maintenance of Vitis vinifera c.v. Muscat de Frontignan
3.2. Chemicals
3.3. Culture Preparation
3.4. Preparation and Treatment with Elicitors
3.6. Estimation of Experimental Parameters from Plant Cells and Medium
3.7. Chemical Analysis of Phenolic Acids with HPLC
3.8. Statistical Analysis
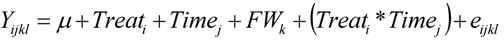
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Kammerer, D.; Claus, A.; Carle, R.; Schieber, A. Polyphenol screening of pomace from red and white grape varieties (Vitis vinifera L.) by HPLC-DAD-MS/MS. J. Agric. Food Chem. 2004, 52, 4360–4367. [Google Scholar]
- Thimothe, J.; Bonsi, I.A.; Padilla-Zakour, O.I.; Koo, H. Chemical characterization of red wine grape (Vitis vinifera and Vitis interspecific hybrids) and pomace phenolic extracts and their biological activity against Streptococcus mutans. J. Agric. Food Chem. 2007, 55, 10200–10207. [Google Scholar]
- Curtin, C.; Zhang, W.; Franco, C. Manipulating anthocyanin composition in Vitis vinifera suspension cultures by elicitation with jasmonic acid and light irradiation. Biotechnol. Lett. 2003, 25, 1131–1135. [Google Scholar] [CrossRef]
- Saw, N.M.M.T.; Riedel, H.; Cai, Z.; Kütük, O.; Smetanska, I. Stimulation of anthocyanin synthesis in grape (Vitis vinifera) cell cultures by pulsed electric fields and ehephon. Plant Cell Tissue and Organ Cult. (PCTOC) 2012, 108, 47–54. [Google Scholar]
- Zhong, J.J. Biochemical engineering of the production of plant-specific secondary metabolites by cell suspension cultures. Adv. Biochem. Engineer.Biotechnol. 2001, 72, 1–26. [Google Scholar] [CrossRef]
- Hammond-Kosack, K.E.; Rudd, J.J. Plant resistance signalling hijacked by a necrotrophic fungal pathogen. Plant Signal. Behvior 2008, 3, 993–995. [Google Scholar]
- Riedel, H.; Cai, Z.; Kütük, O.; Smetanska, I. Obtaining phenolic acids from cell cultures of various Artemisia species. Afr. J. Biotechnol. 2010, 9, 8805–8809. [Google Scholar]
- Mohr, T.J.; Mammarella, N.D.; Hoff, T.; Woffenden, B.J.; Jelesko, J.G.; McDowell, J.M. The Arabidopsis downy mildew resistance gene RPP8 is induced by pathogens and salicylic acid and is regulated by W box cis elements. Mol. Plant Microbe Interact. 2010, 23, 1303–1315. [Google Scholar] [CrossRef]
- Ferrari, S. Biological elicitors of plant secondary metabolites: Mode of action and use in the production of nutraceutics. Adv. Experiment. Med. Biol. 2010, 698, 152–166. [Google Scholar] [CrossRef]
- Snyder, M.J.; Glendinning, J.I. Causal connection between detoxification enzyme activity and consumption of a toxic plant compound. J. Comp. Physiol. 1996, 179, 255–261. [Google Scholar]
- Musser, R.O.; Hum-Musser, S.M.; Eichenseer, H.; Peiffer, M.; Ervin, G.; Murphy, J.B.; Felton, G.W. Herbivory: Caterpillar saliva beats plant defences. Nature 2002, 416, 599–600. [Google Scholar]
- Jwa, N.S.; Agrawal, G.K.; Tamogami, S.; Yonekura, M.; Han, O.; Iwahashi, H.; Rakwal, R. Role of defense/stress-related marker genes, proteins and secondary metabolites in defining rice self-defense mechanisms. Plant Physiol. Biochem. 2006, 44, 261–273. [Google Scholar] [CrossRef]
- Aguilar, O.; Rito-Palomares, M. Aqueous two-phase systems strategies for the recovery and characterization of biological products from plants. J. Sci. Food Agric. 2011, 90, 1385–1392. [Google Scholar]
- Schuler, G.; Mithofer, A.; Baldwin, I.T.; Berger, S.; Ebel, J.; Santos, J.G.; Herrmann, G.; Holscher, D.; Kramell, R.; Kutchan, T.M.; et al. Coronalon: A powerful tool in plant stress physiology. FEBS Lett. 2004, 563, 17–22. [Google Scholar] [CrossRef]
- Fujita, M.; Fujita, Y.; Noutoshi, Y.; Takahashi, F.; Narusaka, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Crosstalk between abiotic and biotic stress responses: A current view from the points of convergence in the stress signaling networks. Curr. Opin. Plant Biol. 2006, 9, 436–442. [Google Scholar]
- Berim, A.; Spring, O.; Conrad, J.; Maitrejean, M.; Boland, W.; Petersen, M. Enhancement of lignan biosynthesis in suspension cultures of Linum nodiflorum by coronalon, indanoyl-isoleucine and methyl jasmonate. Planta 2005, 222, 769–776. [Google Scholar]
- Turlings, T.C.; Tumlinson, J.H. Systemic release of chemical signals by herbivore-injured corn. Proc. Natl. Acad. Sci. USA 1992, 89, 8399–8402. [Google Scholar]
- Halitschke, R.; Schittko, U.; Pohnert, G.; Boland, W.; Baldwin, I.T. Molecular interactions between the specialist herbivore Manduca sexta (Lepidoptera, Sphingidae) and its natural host Nicotiana attenuata. III. Fatty acid-amino acid conjugates in herbivore oral secretions are necessary and sufficient for herbivore-specific plant responses. Plant Physiol. 2001, 125, 711–717. [Google Scholar]
- Turlings, T.C.; Ton, J. Exploiting scents of distress: The prospect of manipulating herbivore-induced plant odours to enhance the control of agricultural pests. Curr. Opin. Plant Biol. 2006, 9, 421–427. [Google Scholar]
- Lait, C.G.; Alborn, H.T.; Teal, P.E.; Tumlinson, J.H. 3rd Rapid biosynthesis of N-linolenoyl-L-glutamine, an elicitor of plant volatiles, by membrane-associated enzyme(s) in Manduca sexta. Proc. Natl. Acad. Sci. USA 2003, 100, 7027–7032. [Google Scholar]
- Schroder, J.; Schroder, G. Stilbene and chalcone synthases: Related enzymes with key functions in plant-specific pathways. Z. Naturforsch. 1990, 45, 1–8. [Google Scholar]
- Howe, G.A.; Jander, G. Plant immunity to insect herbivores. Annu. Rev. Plant Biol. 2008, 59, 41–66. [Google Scholar]
- Reymond, P.; Bodenhausen, N.; Van Poecke, R.M.; Krishnamurthy, V.; Dicke, M.; Farmer, E.E. A conserved transcript pattern in response to a specialist and a generalist herbivore. Plant Cell 2004, 16, 3132–3147. [Google Scholar] [CrossRef]
- Baldwin, I.T.; Halitschke, R.; Kessler, A.; Schittko, U. Merging molecular and ecological approaches in plant-insect interactions. Curr. Opin. Plant Biol. 2001, 4, 351–358. [Google Scholar] [CrossRef]
- Waffo Teguo, P.; Fauconneau, B.; Deffieux, G.; Huguet, F.; Vercauteren, J.; Merillon, J.M. Isolation, identification, and antioxidant activity of three stilbene glucosides newly extracted from vitis vinifera cell culture. J. Nat. Prod. 1998, 61, 655–657. [Google Scholar] [CrossRef]
- Decendit, A.; Merillon, J.M. Condensed tannin and anthocyanin production in Vitis vinifera cell suspension cultures. Plant Cell Rep. 1996, 15, 762–765. [Google Scholar] [CrossRef]
- Cai, Z.; Riedel, H.; Thaw Saw, N.M.; Kutuk, O.; Mewis, I.; Jager, H.; Knorr, D.; Smetanska, I. Effects of pulsed electric field on secondary metabolism of vitis vinifera L. cv. gamay freaux suspension culture and exudates. Appl. Biochem. Biotechnol. 2011, 164, 443–453. [Google Scholar]
- Mewis, I.; Smetanska, I.M.; Muller, C.T.; Ulrichs, C. Specific Poly-phenolic Compounds in Cell Culture of Vitis vinifera L. cv. Gamay Freaux. Appl. Biochem. Biotechnol. 2011, 164, 148–161. [Google Scholar]
- Righetti, L.; Marina, F.; Maura, F.; Annalisa, T.; Nello, B. Resveratrol production in Vitis vinifera cell suspensions treated with several elicitors. CARYOLOGIA 2007, 60, 169–171. [Google Scholar]
- Guerrero, R.F.; Garcia-Parrilla, M.C.; Puertas, B.; Cantos-Villar, E. Wine, resveratrol and health: A review. Nat. Prod. Commun. 2009, 4, 635–658. [Google Scholar]
- Schoppner, A.; Kindl, H. Purification and properties of a stilbene synthase from induced cell suspension cultures of peanut. J. Biol. Chem. 1984, 259, 6806–6811. [Google Scholar]
- Hain, R.; Reif, H.J.; Krause, E.; Langebartels, R.; Kindl, H.; Vornam, B.; Wiese, W.; Schmelzer, E.; Schreier, P.H.; Stocker, R.H.; et al. Disease resistance results from foreign phytoalexin expression in a novel plant. Nature 1993, 361, 153–156. [Google Scholar]
- Begley, T.P.; Kinsland, C.; Strauss, E. The biosynthesis of coenzyme A in bacteria. Vitamin. Hormone. 2001, 61, 157–171. [Google Scholar] [CrossRef]
- Jeandet, P.; Douillet-Breuil, A.C.; Bessis, R.; Debord, S.; Sbaghi, M.; Adrian, M. Phytoalexins from the Vitaceae: Biosynthesis, phytoalexin gene expression in transgenic plants, antifungal activity, and metabolism. J. Agric. Food Chem. 2002, 50, 2731–2741. [Google Scholar]
- Kuc, J. Phytoalexins, stress metabolism, and disease resistance in plant. Annu. Rev. Phytopathol. 1995, 33, 275–297. [Google Scholar] [CrossRef]
- Pare, P.W.; Alborn, H.T.; Tumlinson, J.H. Concerted biosynthesis of an insect elicitor of plant volatiles. Proc. Natl. Acad. Sci. USA 1998, 95, 13971–13975. [Google Scholar]
- Alborn, H.T.; Brennan, M.M.; Tumlinson, J.H. Differential activity and degradation of plant volatile elicitors in regurgitant of tobacco hornworm (Manduca sexta) larvae. J. Chem. Ecol. 2003, 29, 1357–1372. [Google Scholar] [CrossRef]
- Pare, P.W.; Farag, M.A.; Krishnamachari, V.; Zhang, H.; Ryu, C.M.; Kloepper, J.W. Elicitors and priming agents initiate plant defense responses. Photosynth. Res. 2005, 85, 149–159. [Google Scholar] [CrossRef]
- Guardiola, J.; Iborra, J.L.; Canovas, M. A model that links growth and secondary metabolite production in plant cell suspension cultures. Biotechnol. Bioeng. 1995, 46, 291–297. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Riedel, H.; Akumo, D.N.; Saw, N.M.M.T.; Smetanska, I.; Neubauer, P. Investigation of Phenolic Acids in Suspension Cultures of Vitis vinifera Stimulated with Indanoyl-Isoleucine, N-Linolenoyl-L-Glutamine, Malonyl Coenzyme A and Insect Saliva. Metabolites 2012, 2, 165-177. https://doi.org/10.3390/metabo2010165
Riedel H, Akumo DN, Saw NMMT, Smetanska I, Neubauer P. Investigation of Phenolic Acids in Suspension Cultures of Vitis vinifera Stimulated with Indanoyl-Isoleucine, N-Linolenoyl-L-Glutamine, Malonyl Coenzyme A and Insect Saliva. Metabolites. 2012; 2(1):165-177. https://doi.org/10.3390/metabo2010165
Chicago/Turabian StyleRiedel, Heidi, Divine N. Akumo, Nay Min Min Thaw Saw, Iryna Smetanska, and Peter Neubauer. 2012. "Investigation of Phenolic Acids in Suspension Cultures of Vitis vinifera Stimulated with Indanoyl-Isoleucine, N-Linolenoyl-L-Glutamine, Malonyl Coenzyme A and Insect Saliva" Metabolites 2, no. 1: 165-177. https://doi.org/10.3390/metabo2010165
APA StyleRiedel, H., Akumo, D. N., Saw, N. M. M. T., Smetanska, I., & Neubauer, P. (2012). Investigation of Phenolic Acids in Suspension Cultures of Vitis vinifera Stimulated with Indanoyl-Isoleucine, N-Linolenoyl-L-Glutamine, Malonyl Coenzyme A and Insect Saliva. Metabolites, 2(1), 165-177. https://doi.org/10.3390/metabo2010165