

Recent Advancements in Understanding the Role and Mechanisms of Angiopoietin-like Proteins in Diabetic Retinopathy

 ,
,
Abstract
1. Introduction
2. ANGPTL Family Protein Member
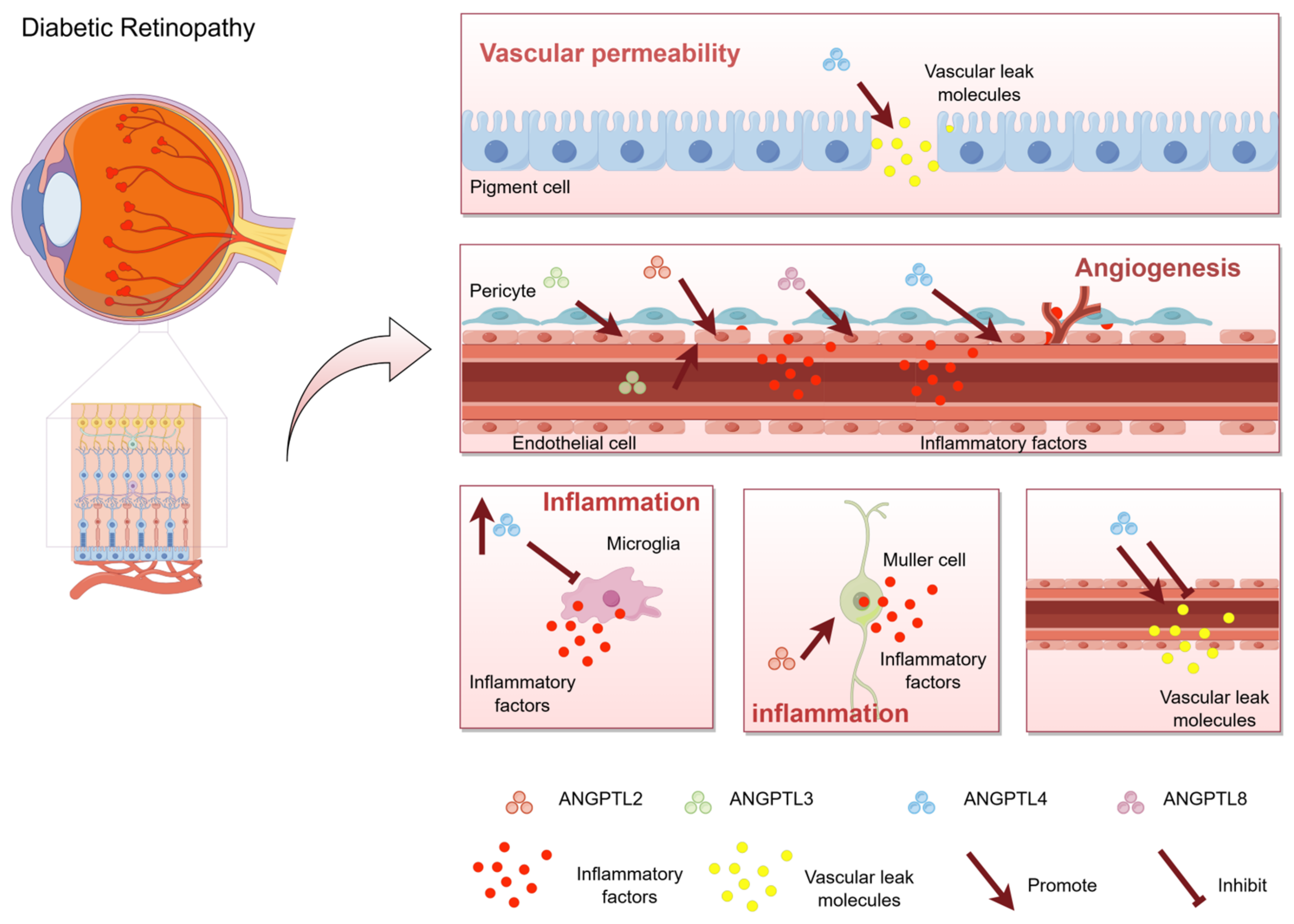
3. The Occurrence and Progression of DR Involve ANGPTL
3.1. ANGPTL2 and DR
3.2. ANGPTL3 and DR
3.3. ANGPTL4 and DR
3.4. ANGPTL8 and DR
4. Molecular Mechanisms of ANGPTL in DR
4.1. Glucose and Lipid Metabolism Involve ANGPTL
4.2. Vascular Permeability Promoted by ANGPTL
4.3. Pathological Angiogenesis Involves ANGPTL
4.4. Intraocular Inflammation Involves ANGPTL
5. Therapeutic Implications
6. Discussion
7. Summary and Prospect
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eisma, J.H.; Dulle, J.E.; Fort, P.E. Current knowledge on diabetic retinopathy from human donor tissues. World J. Diabetes 2015, 6, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Teo, Z.L.; Tham, Y.C.; Yu, M.; Chee, M.L.; Rim, T.H.; Cheung, N.; Bikbov, M.M.; Wang, Y.X.; Tang, Y.; Lu, Y.; et al. Global Prevalence of Diabetic Retinopathy and Projection of Burden through 2045: Systematic Review and Meta-analysis. Ophthalmology 2021, 128, 1580–1591. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Xu, Q.; Li, Z.; Ren, Y.; Jiao, Q.; Wang, L.; Wang, Y. Role of the angiopoietin-like protein family in the progression of NAFLD. Heliyon 2024, 10, e27739. [Google Scholar] [CrossRef]
- Sacks, D.; Baxter, B.; Campbell, B.C.V.; Carpenter, J.S.; Cognard, C.; Dippel, D.; Eesa, M.; Fischer, U.; Hausegger, K.; Hirsch, J.A.; et al. Multisociety Consensus Quality Improvement Revised Consensus Statement for Endovascular Therapy of Acute Ischemic Stroke. Int. J. Stroke Off. J. Int. Stroke Soc. 2018, 13, 612–632. [Google Scholar] [CrossRef]
- Liu, R.; Fu, M.; Chen, P.; Liu, Y.; Huang, W.; Sun, X.; Zhu, P.; Wen, Z.; Cheng, Y. Emerging roles of angiopoietin-like 4 in human tumors (Review). Int. J. Oncol. 2025, 66, 9. [Google Scholar] [CrossRef]
- Yu, C.G.; Yuan, S.S.; Yang, L.Y.; Ke, J.; Zhang, L.J.; Lang, J.N.; Zhang, D.W.; Zhao, S.Z.; Zhao, D.; Feng, Y.M. Angiopoietin-like 3 Is a Potential Biomarker for Retinopathy in Type 2 Diabetic Patients. Am. J. Ophthalmol. 2018, 191, 34–41. [Google Scholar] [CrossRef]
- Fang, C.; Huang, Y.; Guo, H.; Gao, Y.; Ji, X.; Hu, J. Lipasin, a biomarker of diabetic retinopathy. Diabetes Res. Clin. Pract. 2016, 115, 96–98. [Google Scholar] [CrossRef]
- Lu, Q.; Lu, L.; Chen, W.; Lu, P. Expression of angiopoietin-like protein 8 correlates with VEGF in patients with proliferative diabetic retinopathy. Graefe’s Arch. Clin. Exp. Ophthalmol. 2017, 255, 1515–1523. [Google Scholar] [CrossRef]
- Kuo, T.C.; Tan, C.T.; Chang, Y.W.; Hong, C.C.; Lee, W.J.; Chen, M.W.; Jeng, Y.M.; Chiou, J.; Yu, P.; Chen, P.S.; et al. Angiopoietin-like protein 1 suppresses SLUG to inhibit cancer cell motility. J. Clin. Investig. 2013, 123, 1082–1095. [Google Scholar] [CrossRef]
- Chatterjee, T.K.; Aronow, B.J.; Tong, W.S.; Manka, D.; Tang, Y.; Bogdanov, V.Y.; Unruh, D.; Blomkalns, A.L.; Piegore, M.G., Jr.; Weintraub, D.S.; et al. Human coronary artery perivascular adipocytes overexpress genes responsible for regulating vascular morphology, inflammation, and hemostasis. Physiol. Genom. 2013, 45, 697–709. [Google Scholar] [CrossRef]
- Chen, H.A.; Kuo, T.C.; Tseng, C.F.; Ma, J.T.; Yang, S.T.; Yen, C.J.; Yang, C.Y.; Sung, S.Y.; Su, J.L. Angiopoietin-like protein 1 antagonizes MET receptor activity to repress sorafenib resistance and cancer stemness in hepatocellular carcinoma. Hepatology 2016, 64, 1637–1651. [Google Scholar] [CrossRef]
- Wang, D.; Li, H.; Zeng, T.; Chen, Q.; Huang, W.; Huang, Y.; Liao, Y.; Jiang, Q. Exosome-transmitted ANGPTL1 suppresses angiogenesis in glioblastoma by inhibiting the VEGFA/VEGFR2/Akt/eNOS pathway. J. Neuroimmunol. 2024, 387, 578266. [Google Scholar] [CrossRef]
- Doi, Y.; Ninomiya, T.; Hirakawa, Y.; Takahashi, O.; Mukai, N.; Hata, J.; Iwase, M.; Kitazono, T.; Oike, Y.; Kiyohara, Y. Angiopoietin-like protein 2 and risk of type 2 diabetes in a general Japanese population: The Hisayama study. Diabetes Care 2013, 36, 98–100. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.; Lu, Z.; Zheng, J.; Wan, X.; Chen, X.; Hirayasu, K.; Sun, H.; Lam, Y.; Chen, L.; Wang, Q.; et al. A motif in LILRB2 critical for Angptl2 binding and activation. Blood 2014, 124, 924–935. [Google Scholar] [CrossRef] [PubMed]
- Richardson, M.R.; Robbins, E.P.; Vemula, S.; Critser, P.J.; Whittington, C.; Voytik-Harbin, S.L.; Yoder, M.C. Angiopoietin-like protein 2 regulates endothelial colony forming cell vasculogenesis. Angiogenesis 2014, 17, 675–683. [Google Scholar] [CrossRef]
- Farhat, N.; Thorin-Trescases, N.; Mamarbachi, M.; Villeneuve, L.; Yu, C.; Martel, C.; Duquette, N.; Gayda, M.; Nigam, A.; Juneau, M.; et al. Angiopoietin-like 2 promotes atherogenesis in mice. J. Am. Heart Assoc. 2013, 2, e000201. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.; Miyata, K.; Tazume, H.; Sakaguchi, H.; Kadomatsu, T.; Horio, E.; Takahashi, O.; Komohara, Y.; Araki, K.; Hirata, Y.; et al. Perivascular adipose tissue-secreted angiopoietin-like protein 2 (Angptl2) accelerates neointimal hyperplasia after endovascular injury. J. Mol. Cell. Cardiol. 2013, 57, 1–12. [Google Scholar] [CrossRef]
- Ide, S.; Toiyama, Y.; Shimura, T.; Kawamura, M.; Yasuda, H.; Saigusa, S.; Ohi, M.; Tanaka, K.; Mohri, Y.; Kusunoki, M. Angiopoietin-Like Protein 2 Acts as a Novel Biomarker for Diagnosis and Prognosis in Patients with Esophageal Cancer. Ann. Surg. Oncol. 2015, 22, 2585–2592. [Google Scholar] [CrossRef]
- Robciuc, M.R.; Maranghi, M.; Lahikainen, A.; Rader, D.; Bensadoun, A.; Öörni, K.; Metso, J.; Minicocci, I.; Ciociola, E.; Ceci, F.; et al. Angptl3 deficiency is associated with increased insulin sensitivity, lipoprotein lipase activity, and decreased serum free fatty acids. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 1706–1713. [Google Scholar] [CrossRef]
- Tarugi, P.; Bertolini, S.; Calandra, S. Angiopoietin-like protein 3 (ANGPTL3) deficiency and familial combined hypolipidemia. J. Biomed. Res. 2019, 33, 73–81. [Google Scholar] [CrossRef]
- Ortega-Senovilla, H.; Schaefer-Graf, U.; Meitzner, K.; Abou-Dakn, M.; Herrera, E. Decreased concentrations of the lipoprotein lipase inhibitor angiopoietin-like protein 4 and increased serum triacylglycerol are associated with increased neonatal fat mass in pregnant women with gestational diabetes mellitus. J. Clin. Endocrinol. Metab. 2013, 98, 3430–3437. [Google Scholar] [CrossRef] [PubMed]
- Georgiadi, A.; Wang, Y.; Stienstra, R.; Tjeerdema, N.; Janssen, A.; Stalenhoef, A.; van der Vliet, J.A.; de Roos, A.; Tamsma, J.T.; Smit, J.W.; et al. Overexpression of angiopoietin-like protein 4 protects against atherosclerosis development. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 1529–1537. [Google Scholar] [CrossRef] [PubMed]
- Robciuc, M.R.; Naukkarinen, J.; Ortega-Alonso, A.; Tyynismaa, H.; Raivio, T.; Rissanen, A.; Kaprio, J.; Ehnholm, C.; Jauhiainen, M.; Pietiläinen, K.H. Serum angiopoietin-like 4 protein levels and expression in adipose tissue are inversely correlated with obesity in monozygotic twins. J. Lipid Res. 2011, 52, 1575–1582. [Google Scholar] [CrossRef]
- Jonker, J.T.; Smit, J.W.; Hammer, S.; Snel, M.; van der Meer, R.W.; Lamb, H.J.; Mattijssen, F.; Mudde, K.; Jazet, I.M.; Dekkers, O.M.; et al. Dietary modulation of plasma angiopoietin-like protein 4 concentrations in healthy volunteers and in patients with type 2 diabetes. Am. J. Clin. Nutr. 2013, 97, 255–260. [Google Scholar] [CrossRef]
- Katano, H.; Yamada, K. Upregulation of ANGPTL4 messenger RNA and protein in severely calcified carotid plaques. J. Stroke Cerebrovasc. Dis. Off. J. Natl. Stroke Assoc. 2014, 23, 933–947. [Google Scholar] [CrossRef]
- Tan, Z.W.; Teo, Z.; Tan, C.; Choo, C.C.; Loo, W.S.; Song, Y.; Tam, Z.Y.; Ng, S.P.; Koh, H.Z.; Ng, Y.S.; et al. ANGPTL4 T266M variant is associated with reduced cancer invasiveness. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 1525–1536. [Google Scholar] [CrossRef]
- Tan, M.J.; Teo, Z.; Sng, M.K.; Zhu, P.; Tan, N.S. Emerging roles of angiopoietin-like 4 in human cancer. Mol. Cancer Res. MCR 2012, 10, 677–688. [Google Scholar] [CrossRef]
- Shibata, K.; Nakayama, T.; Hirakawa, H.; Hidaka, S.; Nagayasu, T. Clinicopathological significance of angiopoietin-like protein 4 expression in oesophageal squamous cell carcinoma. J. Clin. Pathol. 2010, 63, 1054–1058. [Google Scholar] [CrossRef]
- Goh, Y.Y.; Pal, M.; Chong, H.C.; Zhu, P.; Tan, M.J.; Punugu, L.; Lam, C.R.; Yau, Y.H.; Tan, C.K.; Huang, R.L.; et al. Angiopoietin-like 4 interacts with integrins beta1 and beta5 to modulate keratinocyte migration. Am. J. Pathol. 2010, 177, 2791–2803. [Google Scholar] [CrossRef]
- Zhu, P.; Tan, M.J.; Huang, R.L.; Tan, C.K.; Chong, H.C.; Pal, M.; Lam, C.R.; Boukamp, P.; Pan, J.Y.; Tan, S.H.; et al. Angiopoietin-like 4 protein elevates the prosurvival intracellular O2(-):H2O2 ratio and confers anoikis resistance to tumors. Cancer Cell 2011, 19, 401–415. [Google Scholar] [CrossRef]
- Costa, R.A.; Cardoso, J.C.; Power, D.M. Evolution of the angiopoietin-like gene family in teleosts and their role in skin regeneration. BMC Evol. Biol. 2017, 17, 14. [Google Scholar] [CrossRef] [PubMed]
- Namkung, J.; Koh, S.B.; Kong, I.D.; Choi, J.W.; Yeh, B.I. Serum levels of angiopoietin-related growth factor are increased in metabolic syndrome. Metab. Clin. Exp. 2011, 60, 564–568. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, K.; Hossein-Nezhad, A.; Chamari, M.; Shahbazi, S. Evidence of a role of ANGPTL6 in resting metabolic rate and its potential application in treatment of obesity. Minerva Endocrinol. 2011, 36, 13–21. [Google Scholar] [PubMed]
- Peek, R.; van Gelderen, B.E.; Bruinenberg, M.; Kijlstra, A. Molecular cloning of a new angiopoietinlike factor from the human cornea. Investig. Ophthalmol. Vis. Sci. 1998, 39, 1782–1788. [Google Scholar]
- Saccà, S.C.; Izzotti, A. Focus on molecular events in the anterior chamber leading to glaucoma. Cell. Mol. Life Sci. CMLS 2014, 71, 2197–2218. [Google Scholar] [CrossRef]
- Parri, M.; Pietrovito, L.; Grandi, A.; Campagnoli, S.; De Camilli, E.; Bianchini, F.; Marchiò, S.; Bussolino, F.; Jin, B.; Sarmientos, P.; et al. Angiopoietin-like 7, a novel pro-angiogenetic factor over-expressed in cancer. Angiogenesis 2014, 17, 881–896. [Google Scholar] [CrossRef]
- Fu, Z.; Yao, F.; Abou-Samra, A.B.; Zhang, R. Lipasin, thermoregulated in brown fat, is a novel but atypical member of the angiopoietin-like protein family. Biochem. Biophys. Res. Commun. 2013, 430, 1126–1131. [Google Scholar] [CrossRef]
- Crunkhorn, S. Metabolic disorders: Betatrophin boosts β-cells. Nat. Rev. Drug Discov. 2013, 12, 504. [Google Scholar] [CrossRef]
- Kugelberg, E. Diabetes: Betatrophin--inducing β-cell expansion to treat diabetes mellitus? Nat. Rev. Endocrinol. 2013, 9, 379. [Google Scholar] [CrossRef]
- Tseng, Y.H.; Yeh, Y.H.; Chen, W.J.; Lin, K.H. Emerging regulation and function of betatrophin. Int. J. Mol. Sci. 2014, 15, 23640–23657. [Google Scholar] [CrossRef]
- Dong, X.Y.; Pang, X.W.; Yu, S.T.; Su, Y.R.; Wang, H.C.; Yin, Y.H.; Wang, Y.D.; Chen, W.F. Identification of genes differentially expressed in human hepatocellular carcinoma by a modified suppression subtractive hybridization method. Int. J. Cancer 2004, 112, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Keles, A.; Sonmez, K.; Erol, Y.O.; Ayyıldız, S.N.; Ogus, E. Vitreous levels of vascular endothelial growth factor, stromal cell-derived factor-1α, and angiopoietin-like protein 2 in patients with active proliferative diabetic retinopathy. Graefe’s Arch. Clin. Exp. Ophthalmol. 2021, 259, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Zou, W.; Chen, B.; Zou, C.; Zhao, M.; Zheng, Z. ANGPTL-4 correlates with vascular endothelial growth factor in patients with proliferative diabetic retinopathy. Graefe’s Arch. Clin. Exp. Ophthalmol. 2016, 254, 1281–1288. [Google Scholar] [CrossRef]
- Blot, G.; Karadayi, R.; Przegralek, L.; Sartoris, T.M.; Charles-Messance, H.; Augustin, S.; Negrier, P.; Blond, F.; Muñiz-Ruvalcaba, F.P.; Rivera-de la Parra, D.; et al. Perilipin 2-positive mononuclear phagocytes accumulate in the diabetic retina and promote PPARγ-dependent vasodegeneration. J. Clin. Investig. 2023, 133, e161348. [Google Scholar] [CrossRef]
- Yan, J.; Li, W.J.; Qin, Y.Z.; Qiu, X.Y.; Qin, L.; Li, J.M. Aqueous angiopoietin-like levels correlate with optical coherence tomography angiography metrics in diabetic macular edema. Int. J. Ophthalmol. 2021, 14, 1888–1894. [Google Scholar] [CrossRef]
- Xu, Q.; Gong, C.; Qiao, L.; Feng, R.; Liu, H.; Liu, Y.; Ji, S.; Zhang, Y.; Wu, S.; Li, S. Aqueous Level of ANGPTL4 Correlates with the OCTA Metrics of Diabetic Macular Edema in NPDR. J. Diabetes Res. 2022, 2022, 8435603. [Google Scholar] [CrossRef]
- Perdiguero, E.G.; Galaup, A.; Durand, M.; Teillon, J.; Philippe, J.; Valenzuela, D.M.; Murphy, A.J.; Yancopoulos, G.D.; Thurston, G.; Germain, S. Alteration of developmental and pathological retinal angiogenesis in angptl4-deficient mice. J. Biol. Chem. 2011, 286, 36841–36851. [Google Scholar] [CrossRef]
- Wang, N.; Li, H.; Sun, Q.; Han, X.; Su, S. The Exploration of Disturbance of Capillary and Photoreceptor Communication Networks in Diabetic Retinopathy Through Single-Cell RNA-Seq. J. Cell. Mol. Med. 2025, 29, e70442. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Zhang, D.; Jiang, Z.Y.; Lu, X.Q.; Zheng, X.; Yu, Y.J.; Wang, Y.G.; Dong, J. Positive Association Between Betatrophin and Diabetic Retinopathy Risk in Type 2 Diabetes Patients. Horm. Metab. Res. 2016, 48, 169–173. [Google Scholar] [CrossRef]
- Pehlivan, V.; Gursu, M.F.; Citak, O.; Onalan, E.; Yakar, B.; Donder, E. Evaluation of Serum ANGPTL8/Betatrophin and Cartonectin/CTRP3 Levels in Diabetic and Non-Diabetic Retinopathy. J. Coll. Physicians Surg. Pak. JCPSP 2023, 33, 66–72. [Google Scholar] [CrossRef]
- Feng, S.; Yu, H.; Yu, Y.; Geng, Y.; Li, D.; Yang, C.; Lv, Q.; Lu, L.; Liu, T.; Li, G.; et al. Levels of Inflammatory Cytokines IL-1β, IL-6, IL-8, IL-17A, and TNF-α in Aqueous Humour of Patients with Diabetic Retinopathy. J. Diabetes Res. 2018, 2018, 8546423. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, S.; Seishima, R.; Yamasaki, J.; Hattori, K.; Ogiri, M.; Matsui, S.; Shigeta, K.; Okabayashi, K.; Nagano, O.; Li, L.; et al. Angiopoietin-like 4 promotes glucose metabolism by regulating glucose transporter expression in colorectal cancer. J. Cancer Res. Clin. Oncol. 2022, 148, 1351–1361. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Deng, X.; Jia, J.; Zhao, L.; Wang, C.; Cai, Z.; Guo, C.; Yang, L.; Wang, D.; Ma, S.; et al. Angiopoietin-like protein 8 (betatrophin) inhibits hepatic gluconeogenesis through PI3K/Akt signaling pathway in diabetic mice. Metab. Clin. Exp. 2022, 126, 154921. [Google Scholar] [CrossRef]
- Wu, Z.; Liu, J.; Chen, G.; Du, J.; Cai, H.; Chen, X.; Ye, G.; Luo, Y.; Luo, Y.; Zhang, L.; et al. CD146 is a Novel ANGPTL2 Receptor that Promotes Obesity by Manipulating Lipid Metabolism and Energy Expenditure. Adv. Sci. 2021, 8, 2004032. [Google Scholar] [CrossRef]
- Leander, K.; Chen, Y.Q.; Vikström, M.; Silveira, A.; Fisher, R.M.; Konrad, R.J.; van‘t Hooft, F.M. Circulating ANGPTL3/8 Concentrations Are Associated With an Atherogenic Lipoprotein Profile and Increased CHD Risk in Swedish Population-Based Studies. Arterioscler. Thromb. Vasc. Biol. 2025, 45, 443–451. [Google Scholar] [CrossRef]
- Leth-Espensen, K.Z.; Kristensen, K.K.; Kumari, A.; Winther, A.L.; Young, S.G.; Jørgensen, T.J.D.; Ploug, M. The intrinsic instability of the hydrolase domain of lipoprotein lipase facilitates its inactivation by ANGPTL4-catalyzed unfolding. Proc. Natl. Acad. Sci. USA 2021, 118, e2026650118. [Google Scholar] [CrossRef]
- Chaube, B.; Citrin, K.M.; Sahraei, M.; Singh, A.K.; de Urturi, D.S.; Ding, W.; Pierce, R.W.; Raaisa, R.; Cardone, R.; Kibbey, R.; et al. Suppression of angiopoietin-like 4 reprograms endothelial cell metabolism and inhibits angiogenesis. Nat. Commun. 2023, 14, 8251. [Google Scholar] [CrossRef]
- Zhang, R.; Zhang, K. An updated ANGPTL3-4-8 model as a mechanism of triglyceride partitioning between fat and oxidative tissues. Prog. Lipid Res. 2022, 85, 101140. [Google Scholar] [CrossRef]
- Siddiqa, A.; Ahmad, J.; Ali, A.; Paracha, R.Z.; Bibi, Z.; Aslam, B. Structural characterization of ANGPTL8 (betatrophin) with its interacting partner lipoprotein lipase. Comput. Biol. Chem. 2016, 61, 210–220. [Google Scholar] [CrossRef]
- Silbernagel, G.; Chen, Y.Q.; Li, H.; Lemen, D.; Wen, Y.; Zhen, E.Y.; Rief, M.; Kleber, M.E.; Delgado, G.E.; Sarzynski, M.A.; et al. Associations of Circulating ANGPTL3, C-Terminal Domain-Containing ANGPTL4, and ANGPTL3/8 and ANGPTL4/8 Complexes with LPL Activity, Diabetes, Inflammation, and Cardiovascular Mortality. Circulation 2025, 151, 218–234. [Google Scholar] [CrossRef]
- Xu, Z.; Jiang, G. ANGPTL4-A protein involved in glucose metabolism, lipid metabolism, and tumor development. J. Gene Med. 2024, 26, e3740. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, A.; Ma, T.; Menon, D.; Deshpande, M.; Jee, K.; Dinabandhu, A.; Vancel, J.; Lu, D.; Montaner, S. Angiopoietin-like 4 binds neuropilins and cooperates with VEGF to induce diabetic macular edema. J. Clin. Investig. 2019, 129, 4593–4608. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Lu, P.; Chen, W.; Lu, L.; Zheng, Z. ANGPTL-4 induces diabetic retinal inflammation by activating Profilin-1. Exp. Eye Res. 2018, 166, 140–150. [Google Scholar] [CrossRef]
- Gomez Perdiguero, E.; Liabotis-Fontugne, A.; Durand, M.; Faye, C.; Ricard-Blum, S.; Simonutti, M.; Augustin, S.; Robb, B.M.; Paques, M.; Valenzuela, D.M.; et al. ANGPTL4-αvβ3 interaction counteracts hypoxia-induced vascular permeability by modulating Src signalling downstream of vascular endothelial growth factor receptor 2. J. Pathol. 2016, 240, 461–471. [Google Scholar] [CrossRef]
- Hao, H.; Guo, Z.; Li, Z.; Li, J.; Jiang, S.; Fu, J.; Jiao, Y.; Deng, X.; Han, S.; Li, P. Modified Bu-Fei decoction inhibits lung metastasis via suppressing angiopoietin-like 4. Phytomedicine 2022, 106, 154409. [Google Scholar] [CrossRef]
- Du, X.; Yang, L.; Kong, L.; Sun, Y.; Shen, K.; Cai, Y.; Sun, H.; Zhang, B.; Guo, S.; Zhang, A.; et al. Metabolomics of various samples advancing biomarker discovery and pathogenesis elucidation for diabetic retinopathy. Front. Endocrinol. 2022, 13, 1037164. [Google Scholar] [CrossRef]
- Razavipour, S.F.; Yoon, H.; Jang, K.; Kim, M.; Nawara, H.M.; Bagheri, A.; Huang, W.C.; Shin, M.; Zhao, D.; Zhou, Z.; et al. C-terminally phosphorylated p27 activates self-renewal driver genes to program cancer stem cell expansion, mammary hyperplasia and cancer. Nat. Commun. 2024, 15, 5152. [Google Scholar] [CrossRef]
- Xu, Z.; Yang, J.; Zheng, H.; Xie, T.; Yang, Q.; Cai, J.; Sun, C.; Cao, Y.; Wu, M.; Liu, Y.; et al. Long Noncoding RNA PPT2-EGFL8 Regulates Pathological Retinal Neovascularization in PDR by Functioning as a Competing Endogenous RNA. Diabetes 2023, 72, 1012–1027. [Google Scholar] [CrossRef]
- Shakthiya, T.; Chand, L.; Annamalai, R.; Ka, A.S. Exploring the Association of VEGF-A and ANGPTL2 with the Prognosis of Non-proliferative and Proliferative Diabetic Retinopathy. Cureus 2024, 16, e68273. [Google Scholar] [CrossRef]
- Luo, F.; Wu, P.; Chen, J.; Guo, Y.; Wang, J.; Li, X.; Fang, Z. ANGPTL3 possibly promotes cardiac angiogenesis through improving proangiogenic ability of endothelial progenitor cells after myocardial infarction. Lipids Health Dis. 2018, 17, 184. [Google Scholar] [CrossRef]
- Xiong, Z.; Zhuang, R.L.; Yu, S.L.; Xie, Z.X.; Peng, S.R.; Li, Z.A.; Li, B.H.; Xie, J.J.; Li, Y.N.; Li, K.W.; et al. Cancer-associated fibroblasts regulate mitochondrial metabolism and inhibit chemosensitivity via ANGPTL4-IQGAP1 axis in prostate cancer. J. Adv. Res. 2024, 24, 00559. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhao, Y.; Zhu, P.; Jia, X.; Wang, C.; Zhang, Q.; Li, H.; Wang, J.; Hou, Y. Differential Proteomic Profiles of Coronary Serum Exosomes in Acute Myocardial Infarction Patients with or Without Diabetes Mellitus: ANGPTL6 Accelerates Regeneration of Endothelial Cells Treated with Rapamycin via MAPK Pathways. Cardiovasc. Drugs Ther. 2024, 38, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.; Yang, J.; Deng, G.; Li, D.; Zhang, S. Angiopoietin-like 4 promotes angiogenesis and neurogenesis in a mouse model of acute ischemic stroke. Brain Res. Bull. 2021, 168, 156–164. [Google Scholar] [CrossRef]
- Toyono, T.; Usui, T.; Yokoo, S.; Kimakura, M.; Nakagawa, S.; Yamagami, S.; Miyata, K.; Oike, Y.; Amano, S. Angiopoietin-like protein 2 is a potent hemangiogenic and lymphangiogenic factor in corneal inflammation. Investig. Ophthalmol. Vis. Sci. 2013, 54, 4278–4285. [Google Scholar] [CrossRef]
- Greatbatch, C.J.; Lu, Q.; Hung, S.; Barnett, A.J.; Wing, K.; Liang, H.; Han, X.; Zhou, T.; Siggs, O.M.; Mackey, D.A.; et al. High throughput functional profiling of genes at intraocular pressure loci reveals distinct networks for glaucoma. Hum. Mol. Genet. 2024, 33, 739–751. [Google Scholar] [CrossRef]
- Hirasawa, M.; Takubo, K.; Osada, H.; Miyake, S.; Toda, E.; Endo, M.; Umezawa, K.; Tsubota, K.; Oike, Y.; Ozawa, Y. Angiopoietin-like Protein 2 Is a Multistep Regulator of Inflammatory Neovascularization in a Murine Model of Age-related Macular Degeneration. J. Biol. Chem. 2016, 291, 7373–7385. [Google Scholar] [CrossRef]
- Wang, N.; Zou, C.; Zhao, S.; Wang, Y.; Han, C.; Zheng, Z. Fenofibrate Exerts Protective Effects in Diabetic Retinopathy via Inhibition of the ANGPTL3 Pathway. Investig. Ophthalmol. Vis. Sci. 2018, 59, 4210–4217. [Google Scholar] [CrossRef]
- Ahmad, Z.; Banerjee, P.; Hamon, S.; Chan, K.C.; Bouzelmat, A.; Sasiela, W.J.; Pordy, R.; Mellis, S.; Dansky, H.; Gipe, D.A.; et al. Inhibition of Angiopoietin-Like Protein 3 With a Monoclonal Antibody Reduces Triglycerides in Hypertriglyceridemia. Circulation 2019, 140, 470–486. [Google Scholar] [CrossRef]
- Gaudet, D.; Karwatowska-Prokopczuk, E.; Baum, S.J.; Hurh, E.; Kingsbury, J.; Bartlett, V.J.; Figueroa, A.L.; Piscitelli, P.; Singleton, W.; Witztum, J.L.; et al. Vupanorsen, an N-acetyl galactosamine-conjugated antisense drug to ANGPTL3 mRNA, lowers triglycerides and atherogenic lipoproteins in patients with diabetes, hepatic steatosis, and hypertriglyceridaemia. Eur. Heart J. 2020, 41, 3936–3945. [Google Scholar] [CrossRef]
- Raal, F.J.; Rosenson, R.S.; Reeskamp, L.F.; Hovingh, G.K.; Kastelein, J.J.P.; Rubba, P.; Ali, S.; Banerjee, P.; Chan, K.C.; Gipe, D.A.; et al. Evinacumab for Homozygous Familial Hypercholesterolemia. N. Engl. J. Med. 2020, 383, 711–720. [Google Scholar] [CrossRef]
- Watts, G.F.; Raal, F.J.; Chan, D.C. Transcriptomic therapy for dyslipidemias utilizing nucleic acids targeted at ANGPTL3. Future Cardiol. 2022, 18, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Watts, G.F.; Schwabe, C.; Scott, R.; Gladding, P.A.; Sullivan, D.; Baker, J.; Clifton, P.; Hamilton, J.; Given, B.; Melquist, S.; et al. RNA interference targeting ANGPTL3 for triglyceride and cholesterol lowering: Phase 1 basket trial cohorts. Nat. Med. 2023, 29, 2216–2223. [Google Scholar] [CrossRef]

{kind=link}
| ANGPTL | Main Tissue Expression | Receptor | Chromosome (Human) | Functional Description | Clinical Significance | Related Diseases | References |
|---|---|---|---|---|---|---|---|
| ANGPTL1 | Liver, muscle, thyroid gland, bladder, gallbladder, gastrointestinal tract (no esophagus), adipose tissue, skin | orphan nuclear receptor, site A apolipoprotein (AI) | 1 | Promotes angiogenesis, tissue repair, permeability, anti-apoptotic | Potential role in tissue regeneration | Cance, Cardiovascular diseases | [7,10,11,12,13] |
| ANGPTL2 | Heart, vessels, adipose tissue, kidney, lung, skeletal muscle | integrins α5β1 and Toll-like receptor 4 (TLR4), leukocyte immunoglobulin-like receptor B2 (LILRB 2) | 9 | Inflammation, angiogenesis, development of cancer, regulate lipid metabolism | Biomarker for metabolic disorders | Obesity, Diabetes, Atherosclerosis | [14,15,16,17,18,19] |
| ANGPTL3 | Liver, kidney | alpha-5/beta-3, LILRB2 (weak) | 1 | Angiogenesis, inhibits Lipoprotein lipase (LPL) activity, regulate lipid metabolism | Therapeutic target for lipid disorders | Hyperlipidemia, Atherosclerosis, Loss-of-function mutations | [20,21] |
| ANGPTL4 | Liver, adipose tissue, brain, intestine, thyroid, kidney, heart, muscle, ovary, testis, kidney urinary bladder, esophagus | fibronectin, vitronectin, integrin β1 and β5 | 19 | Inhibits LPL activity, regulates lipid metabolism, energy balance, angiogenesis, glucose metabolism, redox regulation, inflammation, development of cancer | Biomarker and therapeutic target | Obesity, Diabetes, Cancer | [22,23,24,25,26,27,28,29,30,31] |
| ANGPTL5 | Heart, adipose tissue, ovary, testis, skin | LILRB2 | 11 | Lipid, triglyceride metabolism | — | Cancer | [15,32] |
| ANGPTL6 | Liver, gallbladder, placenta | orphan of receptor | 19 | Angiogenesis, lipid metabolism, glucose metabolism | Potential role in vascular health | Cardiovascular diseases, Cancer | [16,33,34] |
| ANGPTL7 | Eye (trabecular meshwork) | LILRB2 (weak) | 1 | Regulates intraocular pressure | Potential therapeutic target for eye diseases | Glaucoma | [15,35,36,37] |
| ANGPTL8 | Liver, adipose tissue | orphan of receptor | 19 | Regulates insulin sensitivity, lipid metabolism | Therapeutic target for metabolic disorders | Obesity, Type 2 Diabetes | [38,39,40,41,42] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Liu, D.; Qiu, Y.; Hu, R.; Chen, S.; Xu, Y.; Qian, C.; Du, L.; Yuan, J.; Li, X. Recent Advancements in Understanding the Role and Mechanisms of Angiopoietin-like Proteins in Diabetic Retinopathy. Metabolites 2025, 15, 352. https://doi.org/10.3390/metabo15060352
Zhang X, Liu D, Qiu Y, Hu R, Chen S, Xu Y, Qian C, Du L, Yuan J, Li X. Recent Advancements in Understanding the Role and Mechanisms of Angiopoietin-like Proteins in Diabetic Retinopathy. Metabolites. 2025; 15(6):352. https://doi.org/10.3390/metabo15060352
Chicago/Turabian StyleZhang, Xinling, Dongang Liu, Yuting Qiu, Ruiyao Hu, Shiyu Chen, Yue Xu, Chenyan Qian, Lailing Du, Jinghua Yuan, and Xiaoping Li. 2025. "Recent Advancements in Understanding the Role and Mechanisms of Angiopoietin-like Proteins in Diabetic Retinopathy" Metabolites 15, no. 6: 352. https://doi.org/10.3390/metabo15060352
APA StyleZhang, X., Liu, D., Qiu, Y., Hu, R., Chen, S., Xu, Y., Qian, C., Du, L., Yuan, J., & Li, X. (2025). Recent Advancements in Understanding the Role and Mechanisms of Angiopoietin-like Proteins in Diabetic Retinopathy. Metabolites, 15(6), 352. https://doi.org/10.3390/metabo15060352