
Metabolomics and Multi-Omics Determination of Potential Plasma Biomarkers in PRV-1-Infected Atlantic Salmon

 ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Trial and Sampling
2.2. Metabolomic Analyses
2.2.1. Targeted Metabolomics Using Standardized Lipid and Metabolic Profiling
2.2.2. Untargeted Metabolomics
2.3. Data Processing
2.3.1. Targeted Metabolomics
2.3.2. Untargeted Metabolomics
2.3.3. Statistical Analyses
2.4. Multi-Omics Analysis
2.4.1. Datasets Included in the Multi-Omics Analysis
2.4.2. Multi-Block Sparse Partial Least-Squares Discriminant Analysis (sPLS-DA)
3. Results
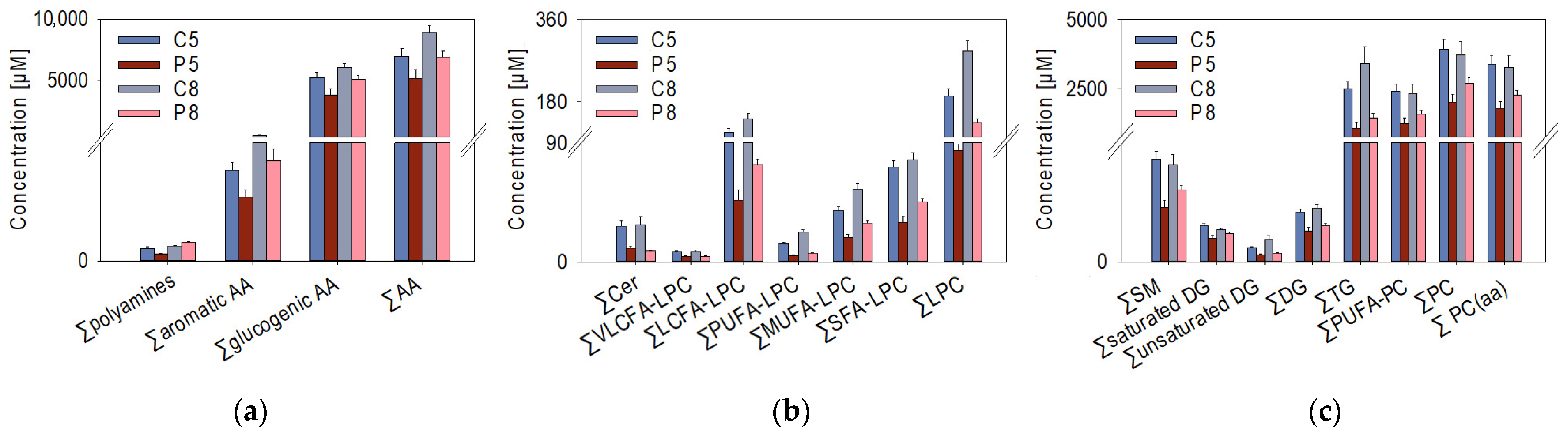
3.1. Targeted Analysis Using the AbsoluteIDQ® p400 HR Kit
3.1.1. Univariate Analysis of the Targeted Metabolomics Data
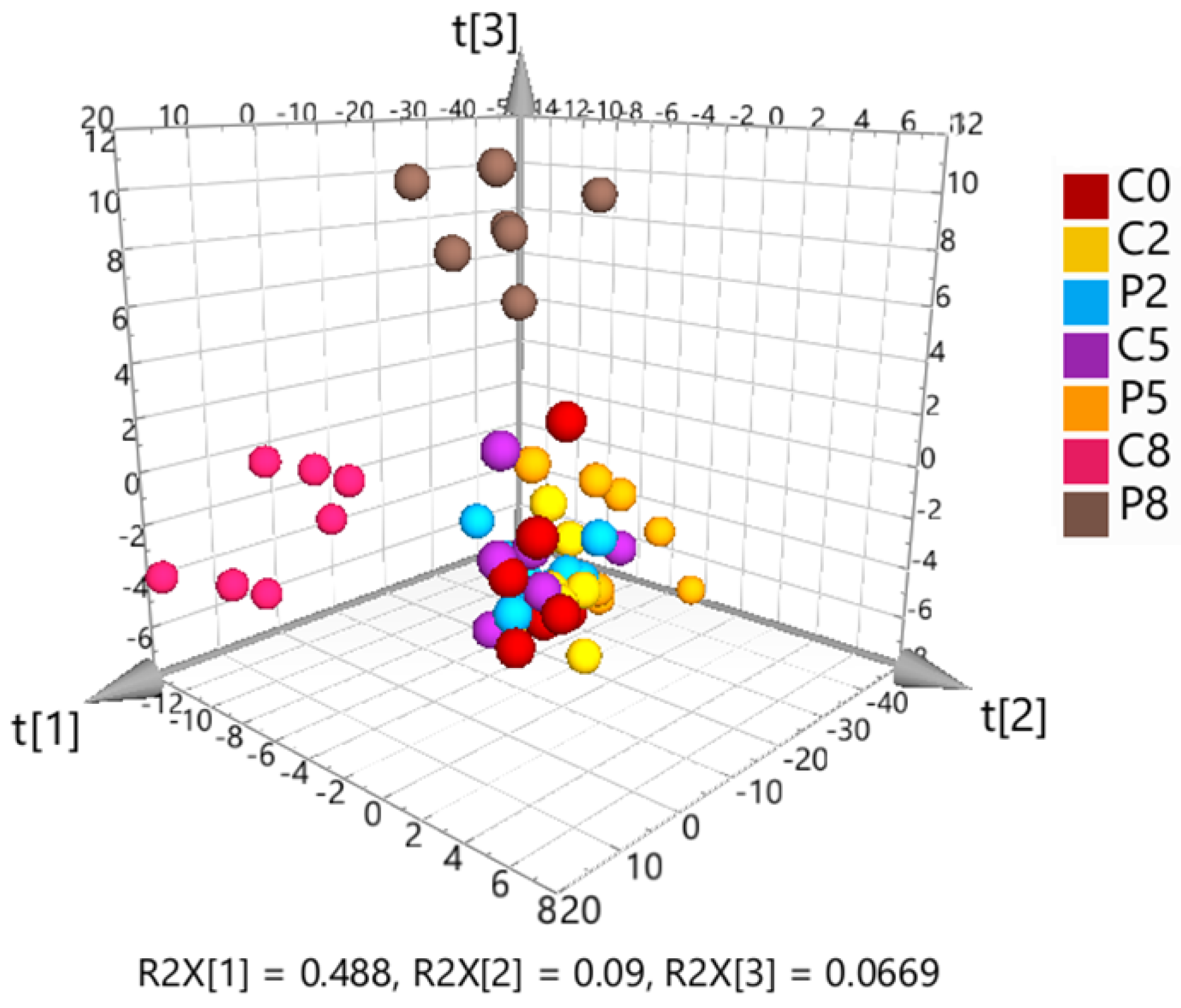
3.1.2. Multivariate Analysis of the Targeted Metabolomics Data
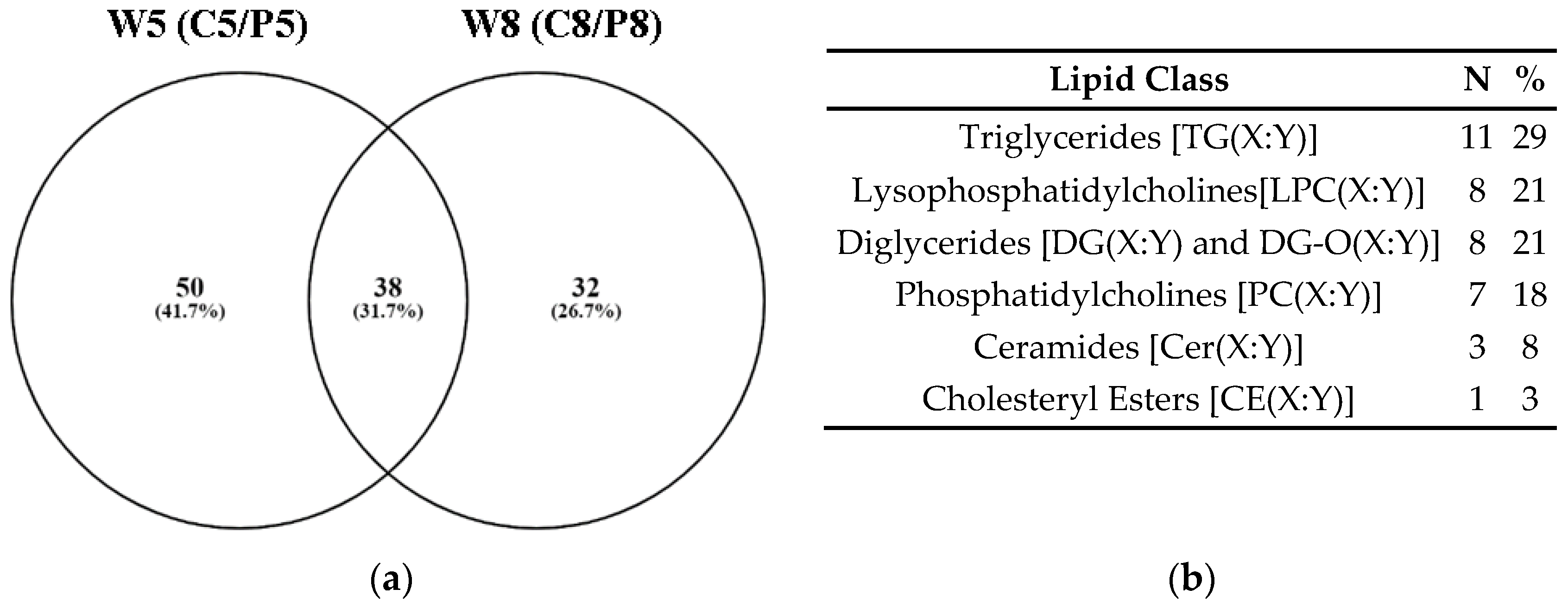
3.2. Untargeted Metabolomics
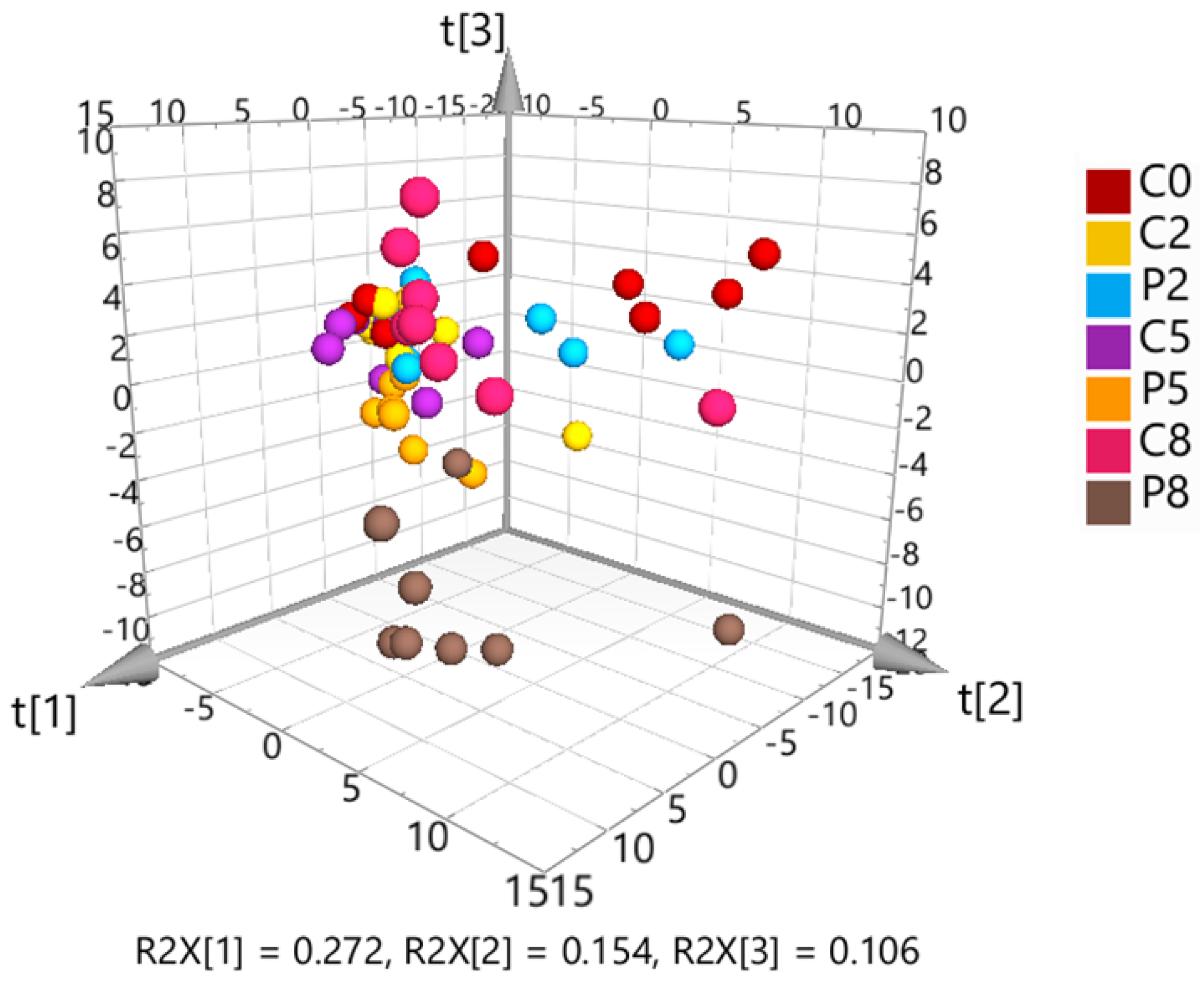
3.2.1. Multivariate Analysis of the Untargeted Metabolomics Data
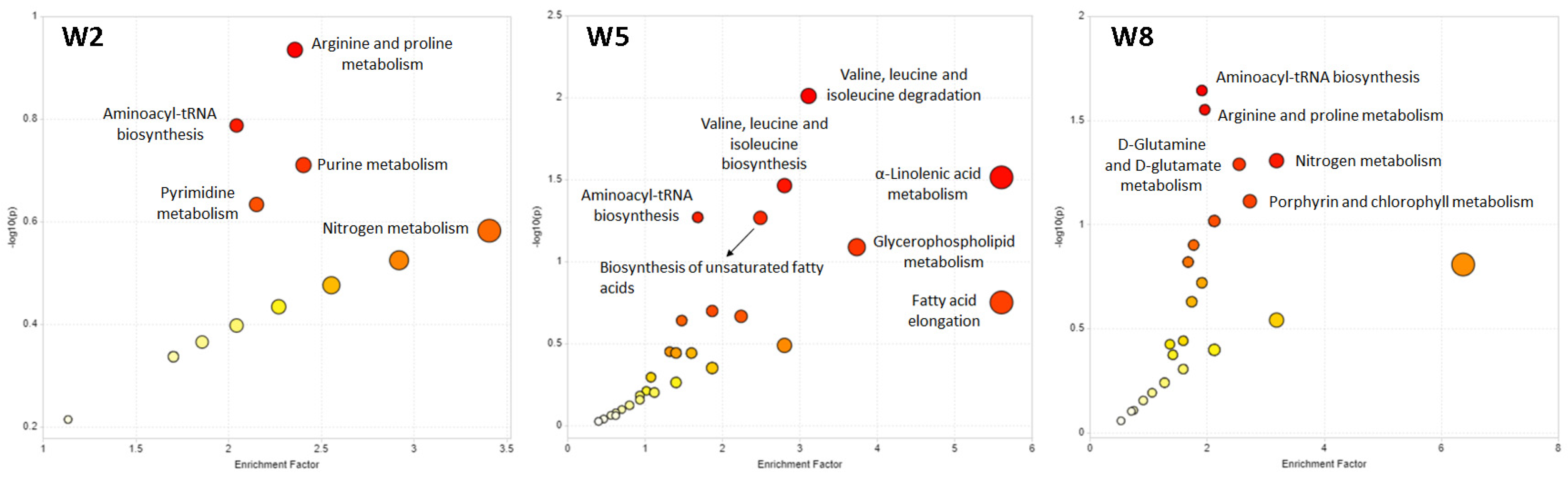
3.2.2. Biological Pathway Analysis of the Untargeted Metabolomics Data
3.3. Multi-Omics for the Integration of Targeted and Untargeted Metabolomics Data with Proteomics Data Available for the Same Salmon Plasma Samples
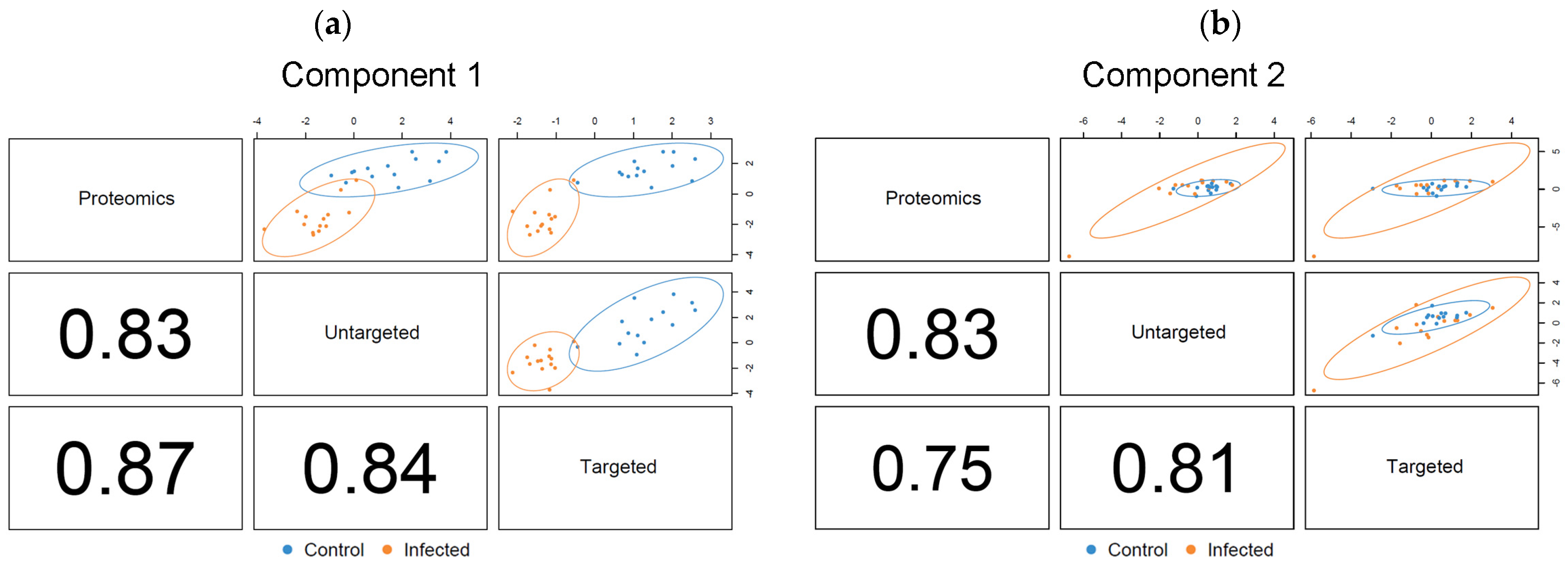
3.3.1. Correlation Analysis between the Data Blocks Using PLS2
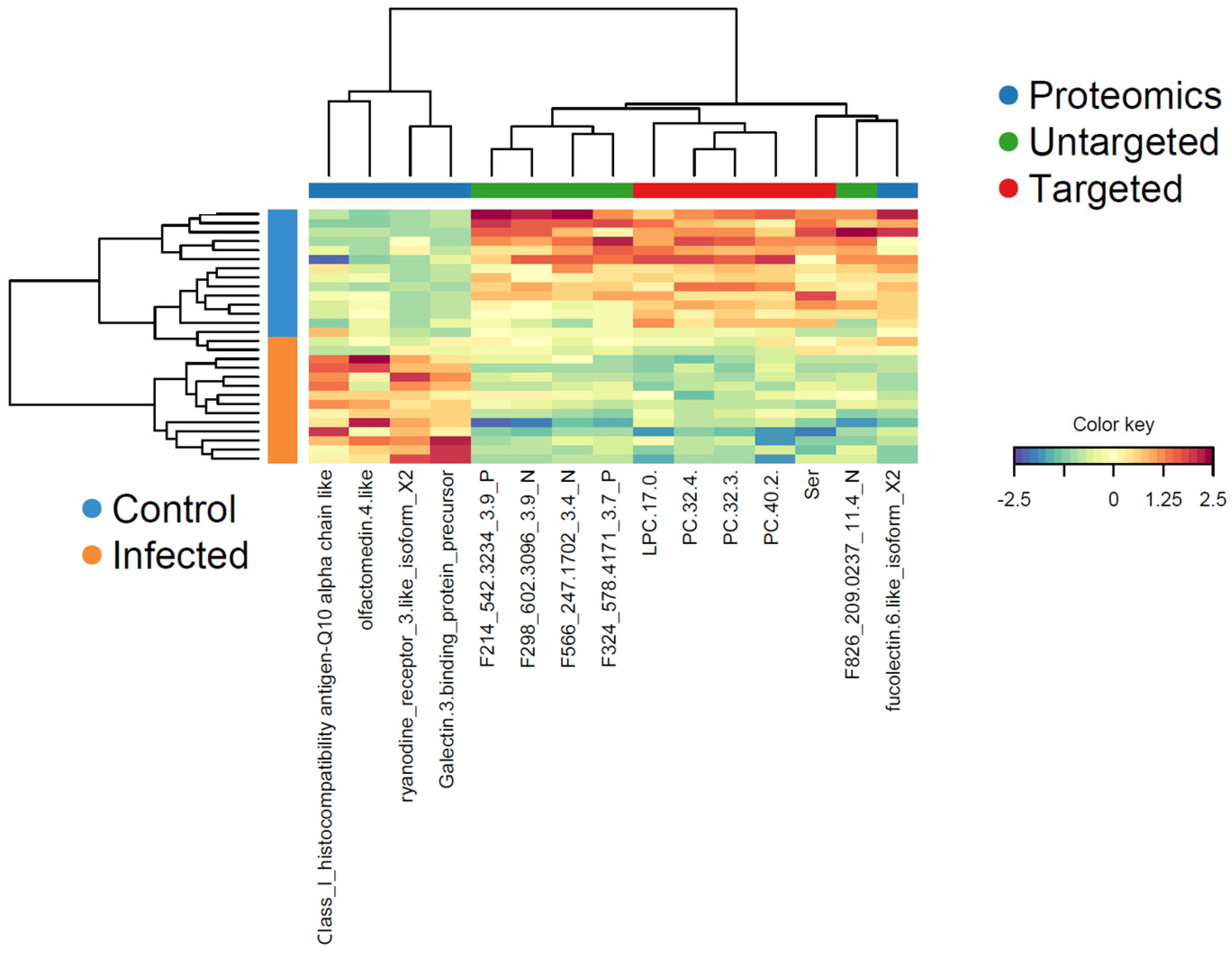
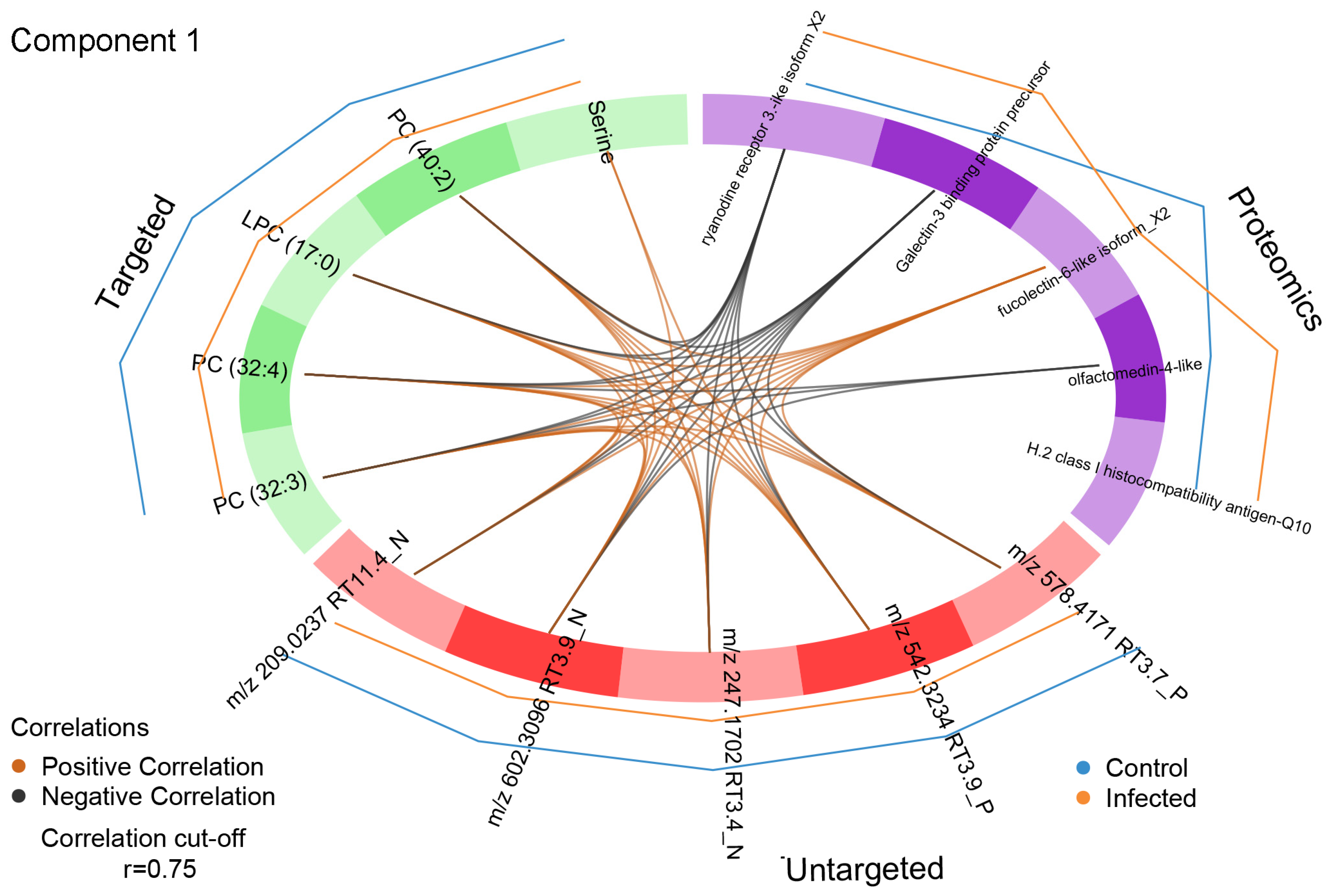
3.3.2. Determination of Relevant Components for Describing the Optimal DIABLO Model
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raposo de Magalhães, C.S.F.; Cerqueira, M.A.C.; Schrama, D.; Moreira, M.J.V.; Boonanuntanasarn, S.; Rodrigues, P.M.L. A Proteomics and other Omics approach in the context of farmed fish welfare and biomarker discovery. Rev. Aquac. 2020, 12, 122–144. [Google Scholar] [CrossRef]
- Mohanty, B.P.; Mohanty, S.; Mitra, T.; Mahanty, A.; Ganguly, S.; Singh, S. Omics technology in fisheries and aquaculture. In Advances in Fish Research; Mohanty, B.P., Ed.; Narendra Publishing House: Delhi, India, 2019; Volume 7, pp. 1–30. [Google Scholar]
- Natnan, M.E.; Mayalvanan, Y.; Jazamuddin, F.M.; Aizat, W.M.; Low, C.F.; Goh, H.H.; Azizan, K.A.; Bunawan, H.; Baharum, S.N. Omics strategies in current advancements of infectious fish disease management. Biology 2021, 10, 1086. [Google Scholar] [CrossRef]
- Alfaro, A.C.; Young, T. Showcasing metabolomic applications in aquaculture: A review. Rev. Aquac. 2018, 10, 135–152. [Google Scholar] [CrossRef]
- Nissa, M.U.; Banerjee, A.; Pinto, N.; Srivastava, S. Proteo-metabolomic technologies in context of fishery/aquacultural research and applications. In Frontiers in Aquaculture Biotechnology; Lakra, W.S., Goswami, M., Trudeau, V.L., Eds.; Academic Press: Cambridge, MS, USA, 2023; pp. 259–274. [Google Scholar] [CrossRef]
- Patti, G.J.; Yanes, O.; Siuzdak, G. Innovation: Metabolomics: The apogee of the omics trilogy. Nat. Rev. Mol. Cell Biol. 2012, 13, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Chubukov, V.; Uhr, M.; Le Chat, L.; Kleijn, R.J.; Jules, M.; Link, H.; Aymerich, S.; Stelling, J.; Sauer, U. Transcriptional regulation is insufficient to explain substrate-induced flux changes in Bacillus subtilis. Mol. Syst. Biol. 2013, 9, 709. [Google Scholar] [CrossRef] [PubMed]
- Rise, M.L.; Martyniuk, C.J.; Chen, M. Comparative physiology and aquaculture: Toward Omics-enabled improvement of aquatic animal health and sustainable production. Comp. Biochem. Physiol. D 2019, 31, 100603. [Google Scholar] [CrossRef] [PubMed]
- Kullgren, A.; Jutfelt, F.; Fontanillas, R.; Sundell, K.; Samuelsson, L.; Wiklander, K.; Kling, P.; Koppe, W.; Larsson, D.G.J.; Björnsson, B.T.; et al. The impact of temperature on the metabolome and endocrine metabolic signals in Atlantic salmon (Salmo salar). Comp. Biochem. Physiol. A 2013, 164, 44–53. [Google Scholar] [CrossRef]
- Zhou, H.; Liu, Y.; Fan, J.; Huang, H.; Deng, J.; Tan, B. Feeding rainbow trout with different types of non-starch polysaccharides: Impacts on serum metabolome and gut microbiota. Metabolites 2022, 12, 1167. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Li, X.; Xu, Z.; Yang, H.; Lin, X.; Leng, X. Replacing fishmeal with cottonseed protein concentrate in practical diet of largemouth bass (Micropterus salmoides): Growth, flesh quality and metabolomics. Aquaculture 2024, 579, 740164. [Google Scholar] [CrossRef]
- Lulijwa, R.; Alfaro, A.C.; Young, T. Metabolomics in salmonid aquaculture research: Applications and future perspectives. Rev. Aquac. 2022, 14, 547–577. [Google Scholar] [CrossRef]
- Su, Y.; Gu, J.Y.; Zhou, Y.G.; Dong, Y.W. Metabolomic responses of Atlantic salmon (Salmo salar) cultured during the pre-smolt, smolt and post-smolt stages. Aquaculture 2024, 582, 740552. [Google Scholar] [CrossRef]
- Lazado, C.C.; Breiland, M.W.; Furtado, F.; Burgerhout, E.; Strand, D. The circulating plasma metabolome of Neoparamoeba perurans-infected Atlantic salmon (Salmo salar). Microb. Pathogenesis 2022, 166, 105553. [Google Scholar] [CrossRef] [PubMed]
- Lokman, P.M.; Symonds, J.E. Molecular and biochemical tricks of the research trade:-omics approaches in finfish aquaculture. N. Z. J. Mar. Freshw. Res. 2014, 48, 492–505. [Google Scholar] [CrossRef]
- Rangel-Huerta, O.D.; Ivanova, L.; Hytterød, S.; Moen, A.; Uhlig, S.; Hagen, A.G.; Olstad, K.; Fæste, C.K. Integrated Omics in salmon skin mucus for determining consequences of chloramine treatment against parasite infestation. J. Integr. OMICS 2020, 10, 5–17. [Google Scholar] [CrossRef]
- Segner, H.; Sundh, H.; Buchmann, K.; Douxfils, J.; Sundell, K.S.; Mathieu, C.; Ruane, N.; Jutfelt, F.; Toften, H.; Vaughan, L. Health of farmed fish: Its relation to fish welfare and its utility as welfare indicator. Fish Physiol. Biochem. 2012, 38, 85–105. [Google Scholar] [CrossRef] [PubMed]
- Yıldız, H.Y.; Seçer, F.S. Stress and fish health: Towards an understanding of allostatic load. In Trends in Fisheries and Aquatic Animal Health; Berillis, P., Ed.; Bentham Science Publishers: Sharjah, United Arab Emirates, 2017; pp. 133–154. [Google Scholar] [CrossRef]
- Sommerset, I.; Wiik-Nielsen, J.; Moldal, T.; Oliveira, V.H.S.; Svendsen, J.C.; Haukaas, A.; Brun, E. Fiskehelserapporten; Norwegian Veterinary Institute: Ås, Norway, 2023; ISSN nr. 1893-1480. [Google Scholar]
- Finstad, Ø.W.; Dahle, M.K.; Lindholm, T.H.; Nyman, I.B.; Løvoll, M.; Wallace, C.; Olsen, C.M.; Rimstad, E. Piscine orthoreovirus (PRV) infects Atlantic salmon erythrocytes. Vet. Res. 2014, 45, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Dhamotharan, K.; Bjørgen, H.; Malik, M.S.; Nyman, I.B.; Markussen, T.; Dahle, M.K.; Koppang, E.O.; Wessel, Ø.; Rimstad, E. Dissemination of piscine orthoreovirus-1 (PRV-1) in Atlantic salmon (Salmo salar) during the early and regenerating phases of infection. Pathogens 2020, 9, 143. [Google Scholar] [CrossRef] [PubMed]
- Mikalsen, A.B.; Haugland, O.; Rode, M.; Solbakk, I.T.; Evensen, O. Atlantic salmon reovirus infection causes a CD8 T cell myocarditis in Atlantic salmon (Salmo salar L.). PLoS ONE 2012, 7, e37269. [Google Scholar] [CrossRef]
- Wessel, Ø.; Hansen, E.F.; Dahle, M.K.; Alarcon, M.; Vatne, N.A.; Nyman, I.B.; Soleim, K.B.; Dhamotharan, K.; Timmerhaus, G.; Markussen, T.; et al. Piscine orthoreovirus-1 isolates differ in their ability to induce heart and skeletal muscle inflammation in Atlantic salmon (Salmo salar). Pathogens 2020, 9, 1050. [Google Scholar] [CrossRef]
- Wessel, O.; Braaen, n.; Alarcon, M.; Haatveit, H.; Roos, N.; Markussen, T.; Tengs, T.; Dahle, M.K.; Rimstad, E. Infection with purified piscine orthoreovirus demonstrates a causal relationship with heart and skeletal muscle inflammation in Atlantic salmon. PLoS ONE 2017, 12, e0183781. [Google Scholar] [CrossRef]
- Kannimuthu, D.; Roh, H.; Morton, H.C.; Peñaranda, M.M.D.; Vossgård, A.; Hansen, T.; Fjelldal, P.G.; Karlsbakk, E.; Fiksdal, I.; Dahle, M.K.; et al. Experimental transmission of piscine orthoreovirus-1 (PRV-1) in different life stages of Atlantic salmon (Salmo salar) and brown trout (Salmo trutta). Front. Mar. Sci. 2023, 10, 1151577. [Google Scholar] [CrossRef]
- Brimsholm, M.; Fjelldal, P.G.; Hansen, T.; Fraser, T.K.; Solberg, M.; Glover, K.; Koppang, E.O.; Bjørgen, H. Red and melanized focal changes in white skeletal muscle in Atlantic salmon (Salmo salar): Comparative analysis of farmed, wild and hybrid fish reared under identical conditions. J. Fish Dis. 2023, 46, 1377–1389. [Google Scholar] [CrossRef] [PubMed]
- Malik, M.S.; Teige, L.H.; Braaen, S.; Olsen, A.B.; Nordberg, M.; Amundsen, M.M.; Dhamotharan, K.; Svenning, S.; Edholm, E.S.; Takano, T.; et al. Piscine orthoreovirus (PRV)-3, but not PRV-2, cross-protects against PRV-1 and heart and skeletal muscle inflammation in Atlantic salmon. Vaccines 2021, 9, 230. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.W.; Adams, K.J.; Adamski, J.; Asad, Y.; Borts, D.; Bowden, J.A.; Byram, G.; Dang, V.; Dunn, W.B.; Fernandez, F.; et al. International ring trial of a high resolution targeted metabolomics and lipidomics platform for serum and plasma analysis. Anal. Chem. 2019, 91, 14407–14416. [Google Scholar] [CrossRef] [PubMed]
- Do, K.T.; Wahl, S.; Raffler, J.; Molnos, S.; Laimighofer, M.; Adamski, J.; Suhre, K.; Strauch, K.; Peters, A.; Gieger, C.; et al. Characterization of missing values in untargeted MS-based metabolomics data and evaluation of missing data handling strategies. Metabolomics 2018, 14, 128–145. [Google Scholar] [CrossRef]
- Huang, Y.; Mu, R.; Wen, D.; Grimsby, J.S.; Liang, M.; Rosenbaum, A.I. Differences in levels of phosphatidylinositols in healthy and stable coronary artery disease subjects revealed by HILIC-MRM method with SERRF normalization. PLoS ONE 2021, 16, e0252426. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.-E.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucl. Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]
- Bevilacqua, M.; Bro, R. Can we trust score plots? Metabolites 2020, 10, 278. [Google Scholar] [CrossRef]
- Dührkop, K.; Fleischauer, M.; Ludwig, M.; Aksenov, A.A.; Melnik, A.V.; Meusel, M.; Dorrestein, P.C.; Rousu, J.; Böcker, S. SIRIUS 4: A rapid tool for turning tandem mass spectra into metabolite structureinformation. Nat. Meth. 2019, 16, 299–302. [Google Scholar] [CrossRef]
- Dührkop, K.; Nothias, L.F.; Fleischauer, M.; Reher, R.; Ludwig, M.; Hoffmann, M.A.; Petras, D.; Gerwick, W.H.; Rousu, J.; Dorrestein, P.C.; et al. Systematic classification of unknown metabolites using high-resolution fragmentation mass spectra. Nat. Biotechnol. 2021, 39, 462–471. [Google Scholar] [CrossRef]
- Djoumbou Feunang, Y.; Eisner, R.; Knox, C.; Chepelev, L.; Hastings, J.; Owen, G.; Fahy, E.; Steinbeck, C.; Subramanian, S.; Bolton, E.; et al. ClassyFire: Automated chemical classification with a comprehensive, computable taxonomy. J. Cheminform. 2016, 8, 61. [Google Scholar] [CrossRef] [PubMed]
- Fæste, C.K.; Rangel Huerta, O.D.; Anonsen, J.H.; Tator, H.; Kuiper, R.V.; Dahle, M.K. Potential plasma biomarkers for the onset of heart and skeletal muscle inflammation from Piscine orthoreovirus-1 infection in Atlantic salmon (Salmo salar). Aquaculture 2024, 590, 741045. [Google Scholar] [CrossRef]
- Singh, A.; Shannon, C.P.; Gautier, B.; Rohart, F.; Vacher, M.; Tebbutt, S.J.; Cao, K.A.L. DIABLO: An integrative approach for identifying key molecular drivers from multi-omics assays. Bioinformatics 2019, 35, 3055–3062. [Google Scholar] [CrossRef] [PubMed]
- Rohart, F.; Gautier, B.; Singh, A.; Cao, K.A.L. mixOmics: An R package for ‘omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [PubMed]
- Downing, T.; Angelopoulos, N. A primer on correlation-based dimension reduction methods for multi-omics analysis. J. R. Soc. Interface 2023, 20, 20230344. [Google Scholar] [CrossRef] [PubMed]
- González, I.; Cao, K.A.L.; Davis, M.J.; Déjean, S. Visualising associations between paired ‘omics’ data sets. BioData Min. 2012, 5, 19. Available online: http://www.biodatamining.org/content/5/1/19 (accessed on 8 April 2024). [CrossRef] [PubMed]
- Ivanova, L.; Rangel-Huerta, O.D.; Tartor, H.; Gjessing, M.C.; Dahle, M.K.; Uhlig, S. Fish skin and gill mucus: A source of metabolites for non-invasive health monitoring and research. Metabolites 2021, 12, 28. [Google Scholar] [CrossRef] [PubMed]
- Forsythe, I.J.; Wishart, D.S. Exploring human metabolites using the human metabolome database. Curr. Protoc. Bioinform. 2009, 25, 14.8.1–14.8.45. [Google Scholar] [CrossRef]
- The Human Metabolome Database, Version 5.0. Available online: https://hmdb.ca/ (accessed on 8 April 2024).
- Molversmyr, E.; Devle, H.M.; Naess-Andresen, C.F.; Ekeberg, D. Identification and quantification of lipids in wild and farmed Atlantic salmon (Salmo salar), and salmon feed by GC-MS. Food Sci. Nutr. 2022, 10, 3117–3127. [Google Scholar] [CrossRef]
- Heaton, N.S.; Randall, G. Multifaceted roles for lipids in viral infection. Trends Microbiol. 2011, 19, 368–375. [Google Scholar] [CrossRef]
- Pezzotti, G. Raman spectroscopy in cell biology and microbiology. J. Raman Spectrosc. 2021, 52, 2348–2443. [Google Scholar] [CrossRef]
- Sanchez, E.L.; Lagunoff, M. Viral activation of cellular metabolism. Virology 2015, 479, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Casari, I.; Manfredi, M.; Metharom, P.; Falasca, M. Dissecting lipid metabolism alterations in SARS-CoV-2. Prog. Lipid Res. 2021, 82, 101092. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Chen, D.; Wu, L.; He, G.; Ye, W. Low serum cholesterol level among patients with COVID-19 Infection in Wenzhou, China (21 February 2020). [CrossRef]
- Beckmann, N.; Becker, K.A. Ceramide and related molecules in viral infections. Int. J. Mol. Sci. 2021, 22, 5676. [Google Scholar] [CrossRef] [PubMed]
- Havranek, K.E.; Reyes Ballista, J.M.; Hines, K.M.; Brindley, M.A. Untargeted lipidomics of vesicular stomatitis virus-infected cells and viral particles. Viruses 2021, 14, 3. [Google Scholar] [CrossRef] [PubMed]
- Belury, M.A.; Bowman, E.; Gabriel, J.; Snyder, B.; Kulkarni, M.; Palettas, M.; Mo, X.; Lake, J.E.; Zidar, D.; Sieg, S.F.; et al. Prospective analysis of lipid composition changes with antiretroviral therapy and immune activation in persons living with HIV. Pathog. Immun. 2017, 2, 376. [Google Scholar] [CrossRef] [PubMed]
- Perlemuter, G.; Sabile, A.; Letteron, P.; Vona, G.; Topilco, A.; Chrétien, Y.; Koike, K.; Pessayre, D.; Chapman, J.; Barba, G.; et al. Hepatitis C virus core protein inhibits microsomal triglyceride transfer protein activity and very low density lipoprotein secretion: A model of viral-related steatosis. FASEB J. 2002, 16, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.-Y.; Chuang, W.-L.; Ho, C.-K.; Hsieh, M.-Y.; Huang, J.-F.; Lee, L.-P.; Hou, N.-J.; Lin, Z.-Y.; Chen, S.-C.; Hsieh, M.-Y.; et al. Associations between hepatitis C viremia and low serum triglyceride and cholesterol levels: A community-based study. J. Hepatol. 2008, 49, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Johansen, L.H.; Dahle, M.K.; Wessel, Ø.; Timmerhaus, G.; Løvoll, M.; Røsæg, M.; Jørgensen, S.M.; Rimstad, E.; Krasnov, A. Differences in gene expression in Atlantic salmon parr and smolt after challenge with Piscine orthoreovirus (PRV). Mol. Immunol. 2016, 73, 138–150. [Google Scholar] [CrossRef]
- Park, E.-S.; Lee, J.H.; Hong, J.H.; Park, Y.K.; Lee, J.W.; Lee, W.-J.; Lee, J.W.; Kim, K.-H. Phosphatidylcholine alteration identified using MALDI imaging MS in HBV-infected mouse livers and virus-mediated regeneration defects. PLoS ONE 2014, 9, e103955. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-F.; Wu, W.-Y.; Qiu, G.-K.; Wang, H.; Li, W.-S.; Wang, Y.-L.; Jiang, Q.-Q.; Han, M.-F.; Ning, Q. Plasma lipidomics identifies novel biomarkers in patients with hepatitis B virus-related acute-on-chronic liver failure. Metabolomics 2017, 13, 76. [Google Scholar] [CrossRef]
- Xu, S.; Zheng, Z.; Pathak, J.L.; Cheng, H.; Zhou, Z.; Chen, Y.; Wu, Q.; Wang, L.; Zeng, M.; Wu, L. The emerging role of the serine incorporator protein family in regulating viral infection. Front. Cell Dev. Biol. 2022, 10, 856468. [Google Scholar] [CrossRef]
- Delova, A.; Pasc, A.; Monari, A. Interaction of the immune system TIM-3 protein with a model cellular membrane containing phosphatidyl-serine lipids. Chemistry 2024, 30, e202304318. [Google Scholar] [CrossRef] [PubMed]
- DeRoo, E.P.; Wrobleski, S.K.; Shea, E.M.; Al-Khalil, R.K.; Hawley, A.E.; Henke, P.K.; Myers, D.D., Jr.; Wakefield, T.W.; Diaz, J.A. The role of galectin-3 and galectin-3–binding protein in venous thrombosis. Blood 2015, 125, 1813–1821. [Google Scholar] [CrossRef] [PubMed]
- Zhen, S.; Cai, R.; Yang, X.; Ma, Y.; Wen, D. Association of serum galectin-3-binding protein and metabolic syndrome in a Chinese adult population. Front. Endocrin. 2021, 12, 726154. [Google Scholar] [CrossRef] [PubMed]
- Dara, M.; Giulianini, P.G.; Manfrin, C.; Parisi, M.G.; Parrinello, D.; La Corte, C.; Vasta, G.R.; Cammarata, M. F-type lectin from serum of the Antarctic teleost fish Trematomus bernacchii (Boulenger, 1902): Purification, structural characterization, and bacterial agglutinating activity. Comp. Biochem. Physiol. B 2021, 256, 110633. [Google Scholar] [CrossRef]
- Dulhunty, A.F.; Casarotto, M.G.; Beard, N.A. The ryanodine receptor: A pivotal Ca2+ regulatory protein and potential therapeutic drug target. Curr. Drug Targets 2011, 12, 709–723. [Google Scholar] [CrossRef]
- Liu, W.; Rodgers, G.P. Olfactomedin 4 is a biomarker for the severity of infectious diseases. Open Forum Infect. Dis. 2022, 9, ofac061. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Class 1 | Targeted by Kit 2 | Detected in Salmon Plasma 3 | Major Metabolites (Top Three) in Compound Class 4 |
|---|---|---|---|
| Acylcarnitines [AC(X:Y)] | 55 | 35 | AC(0:0), AC(2:0), AC(18:1) |
| Amino acids | 21 | 21 | glycine, glutamine, alanine |
| Biogenic amines | 21 | 10 | taurine, trans-4-OH-proline, putrescine |
| Cholesteryl Esters [CE(X:Y)] | 14 | 11 | CE(22:6), CE(20:5), CE(18:2) |
| Diglycerides [DG(X:Y) and DG-O(X:Y)] | 18 | 17 | DG(39:0), DG(36:2), DG-O(34:1) |
| Triglycerides [TG(X:Y)] | 42 | 30 | TG(54:3), TG(52:2), TG(54:4) |
| Lysophosphatidylcholines [LPC(X:Y)] | 24 | 17 | LPC(22:6), LPC(16:0), LPC(18:1) |
| Phosphatidylcholines [PC(X:Y) and PC-O(X:Y)] | 172 | 95 | PC(34:1), PC(34:2), PC(36:4) |
| Ceramides [Cer(X:Y)] | 9 | 4 | Cer(42:2), Cer(42:1), Cer(34:1) |
| Sphingomyelins [SM(X:Y)] | 31 | 22 | SM(42:2), SM(38:2), SM(40:2) |
| Sum hexoses [including glucose] | 1 | 1 | H1 |
| Models | R2X | R2Y | Q2 | Δ%R2Y−Q2 | CV-ANOVA | Permutation |
|---|---|---|---|---|---|---|
| C2 vs. P2 | 0.348 | 0.930 | 0.555 | 40% | 0.047 | valid |
| C5 vs. P5 | 0.722 | 0.606 | 0.522 | 14% | 0.017 | valid |
| C8 vs. P8 | 0.634 | 0.986 | 0.954 | 3.2% | 0.0000049 | valid |
| Models | R2X | R2Y | Q2 | Δ%R2Y−Q2 | CV-ANOVA | Permutation |
|---|---|---|---|---|---|---|
| C2 vs. P2 | 0.233 | 0.701 | 0.440 | 37% | 0.0413 | valid |
| C5 vs. P5 | 0.227 | 0.809 | 0.604 | 25% | 0.0061 | valid |
| C8 vs. P8 | 0.342 | 0.897 | 0.838 | 6.6% | 0.00001 | valid |
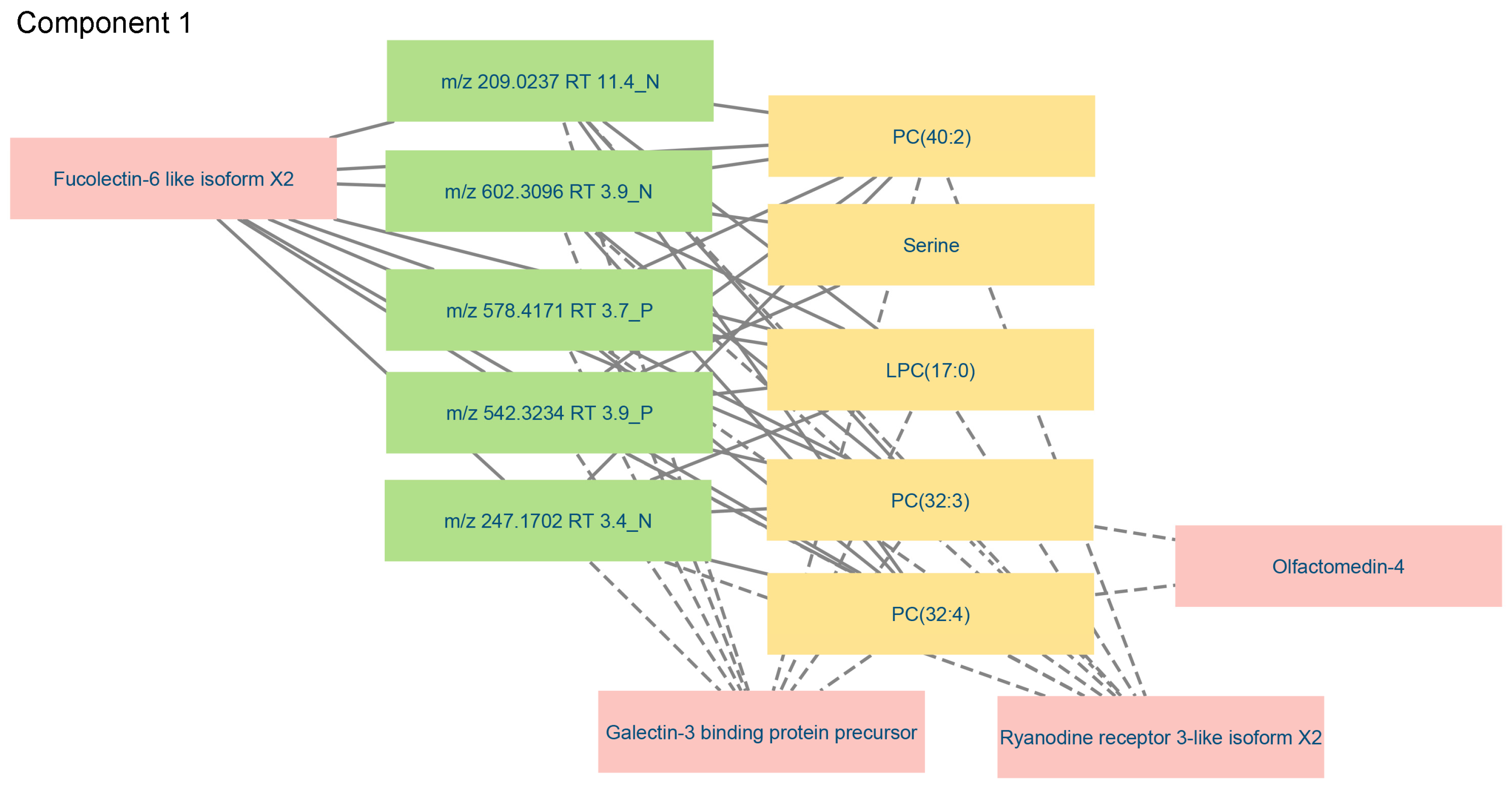
| Targeted Metabolomics Block 1 | Untargeted Metabolomics Block 2 | Proteomics Block |
|---|---|---|
| Component 1 | ||
| LPC (17:0) | m/z 578.4171 RT 3.7 P (LPC(22:1)) | Ryanodine receptor 3-like isoform X2 |
| PC (32:3) | m/z 542.3234 RT 3.9 P (PC(20:5)) | Fucolectin-6-like isoform X2 |
| PC (32:4) | m/z 209.0237 RT 11.4 N | Olfactomedin 4-like |
| PC (40:2) | m/z 247.1702 RT 3.4 N (FA(16:4)) | Galectin-3-binding protein precursor |
| Serine | m/z 602.3096 RT 3.9 N (phosphatidylserine) | H-2 class I histocompatibility antigen-Q10 alpha chain-like |
| Component 2 | ||
| Alanine | m/z 212.1387 RT 9.8 P (amino acid and derivatives) | Histone H3-like partial |
| AC (6:0) | m/z 149.0454 RT 12.6 N (D-xylose) | ATP dependent 6-phospho-fructokinase-muscle type like |
| AC (12:0-DC) | m/z 168.1492 RT 9.8 P | Glycine-rich RNA binding protein-like isoform X1 |
| ADMA | m/z 203.1499 RT 22.2 P (ADMA) | Histone H3-3 |
| SDMA | m/z 166.0144 RT 16.4 N | Barrier to autointegration factor |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ivanova, L.; Rangel-Huerta, O.D.; Tartor, H.; Dahle, M.K.; Uhlig, S.; Fæste, C.K. Metabolomics and Multi-Omics Determination of Potential Plasma Biomarkers in PRV-1-Infected Atlantic Salmon. Metabolites 2024, 14, 375. https://doi.org/10.3390/metabo14070375
Ivanova L, Rangel-Huerta OD, Tartor H, Dahle MK, Uhlig S, Fæste CK. Metabolomics and Multi-Omics Determination of Potential Plasma Biomarkers in PRV-1-Infected Atlantic Salmon. Metabolites. 2024; 14(7):375. https://doi.org/10.3390/metabo14070375
Chicago/Turabian StyleIvanova, Lada, Oscar D. Rangel-Huerta, Haitham Tartor, Maria K. Dahle, Silvio Uhlig, and Christiane Kruse Fæste. 2024. "Metabolomics and Multi-Omics Determination of Potential Plasma Biomarkers in PRV-1-Infected Atlantic Salmon" Metabolites 14, no. 7: 375. https://doi.org/10.3390/metabo14070375
APA StyleIvanova, L., Rangel-Huerta, O. D., Tartor, H., Dahle, M. K., Uhlig, S., & Fæste, C. K. (2024). Metabolomics and Multi-Omics Determination of Potential Plasma Biomarkers in PRV-1-Infected Atlantic Salmon. Metabolites, 14(7), 375. https://doi.org/10.3390/metabo14070375