


A Pilot Study on Across-Generation Impacts of Maternal Heat Stress on Blood Metabolites of Female Holstein Dairy Calves



Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Housing System, Feeding and Management
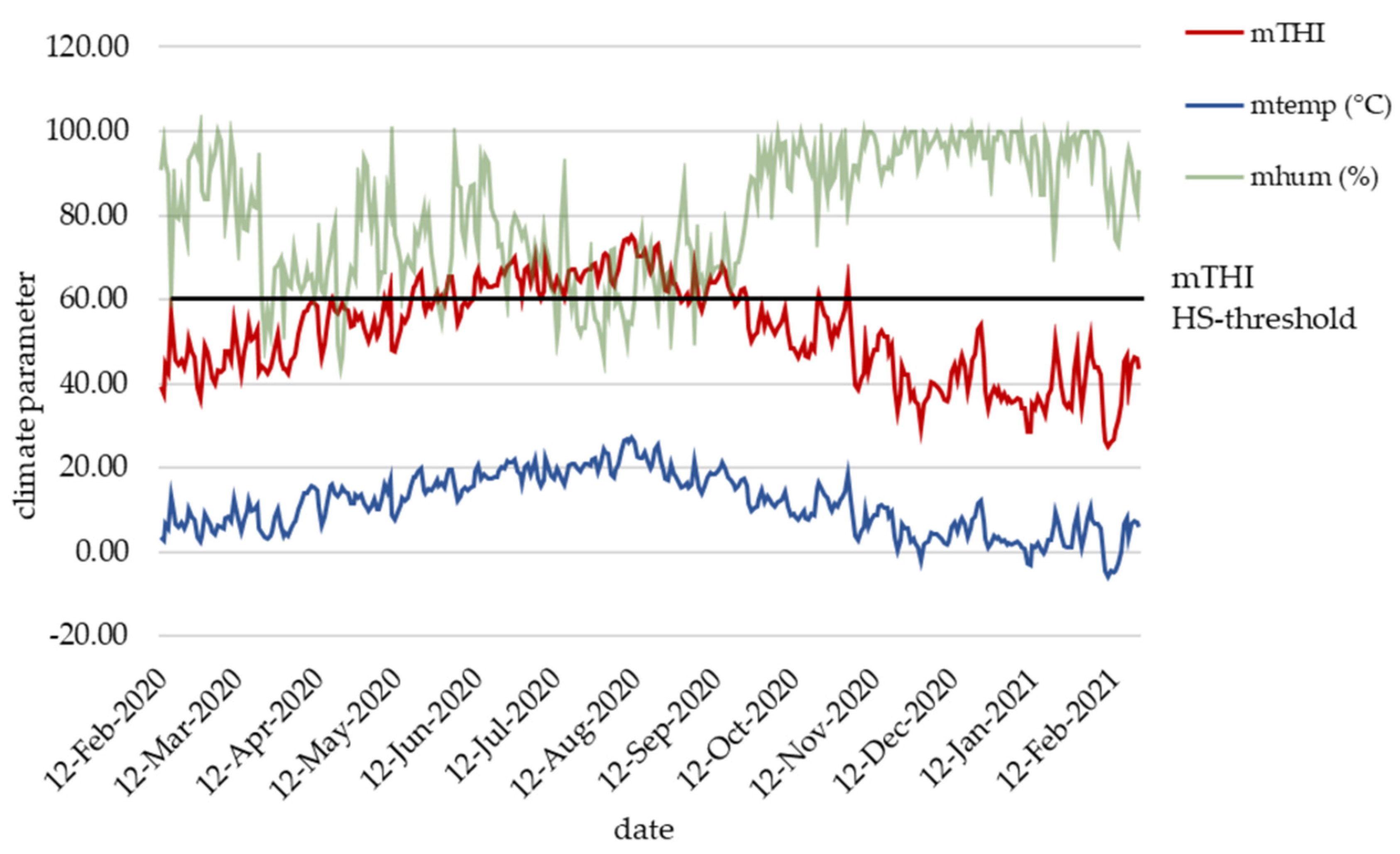
2.2. Climate Data Recording and HS Index
2.3. Blood Sampling and Blood Plasma Metabolome Profiling
2.4. Data Analysis and Visualization
- age of calf: blood sampling at 1st day of age = class 1; blood sampling at 2nd to 3rd day of age = class 2; blood sampling at 4th to 5th day of age = class 3; blood sampling at 6th to 7th day of age = class 4
- calving condition: no birth assistance = class 1; minor birth assistance or dystocia = class 2
- birth weight class for calves: birth weight from 34.8 kg to 39.0 kg = weight class 1; birth weight from 39.1 kg to 48.0 kg = weight class 2
- time of blood sampling: blood sampling from 10:00 a.m. to 12:00 p.m. (morning hours) = class 1; blood sampling from 12:01 p.m. to 15:15 p.m. (afternoon hours) = class 2
3. Results
3.1. Climate Data
3.2. Animal Performance
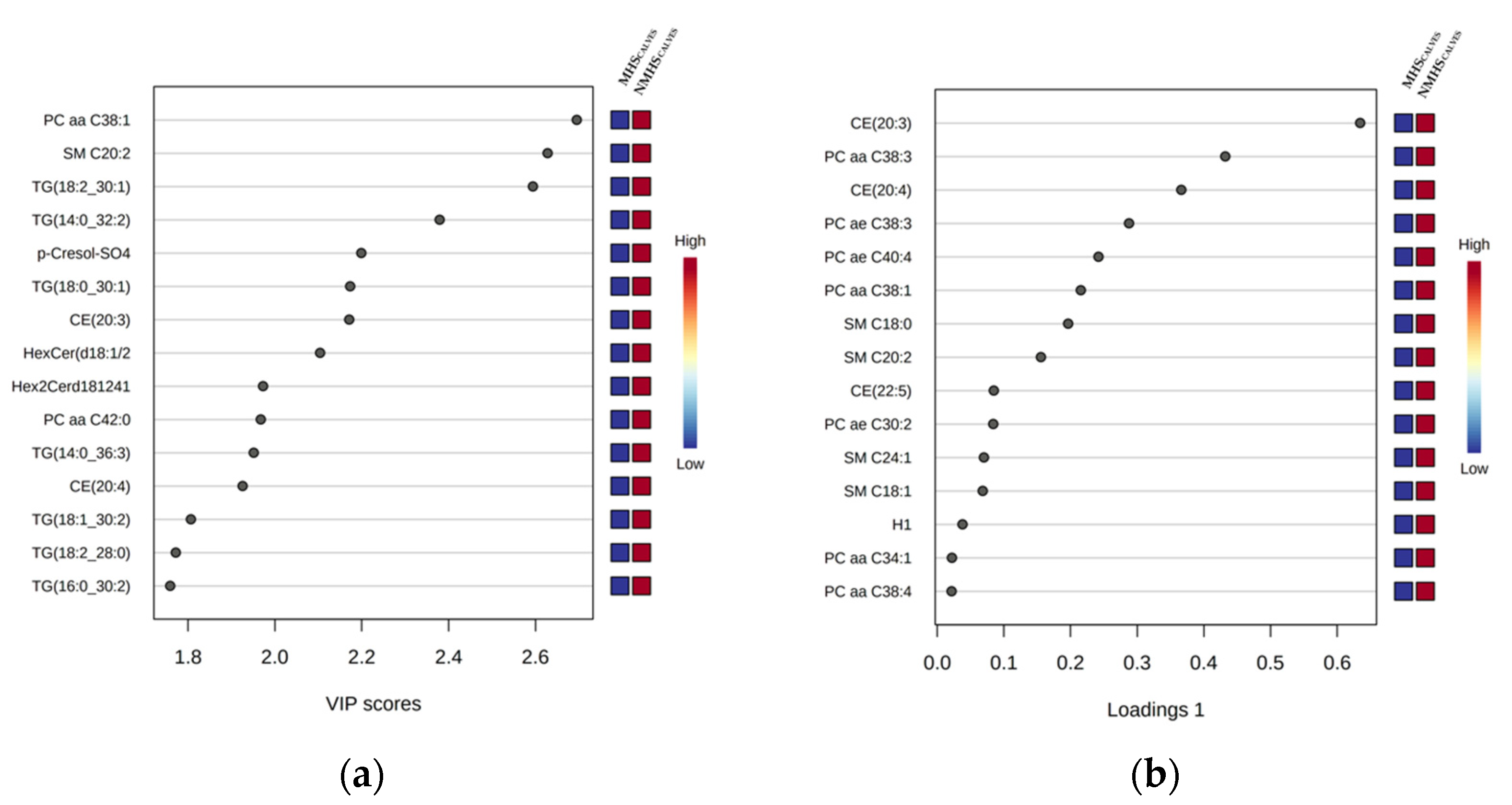
3.3. Phenotypic Correlations between Metabolite Concentrations and Birth Weight in Calves
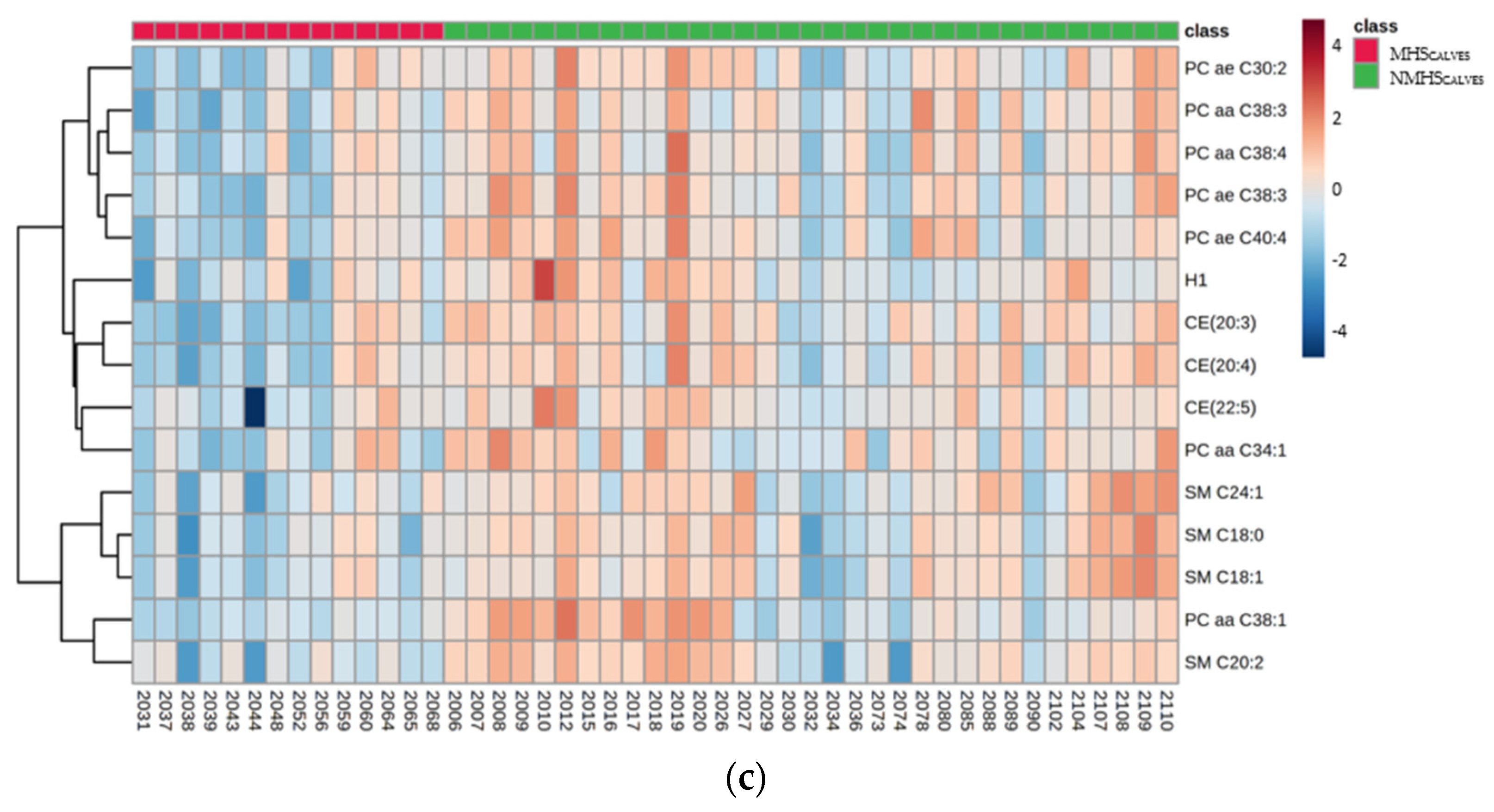
3.4. Effect of Maternal HS during the Last Week before Birth on Blood Metabolite Profiles of Calves
4. Discussion
4.1. Lipids
4.2. p-Cresol Sulfate
4.3. Hexoses (90 to 95% Glucose)
4.4. Epigenetics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tao, S.; Bubolz, J.W.; do Amaral, B.C.; Thompson, I.M.; Hayen, M.J.; Johnson, S.E.; Dahl, G.E. Effects of heat stress during the dry period on mammary gland development. J. Dairy Sci. 2011, 94, 5976–5986. [Google Scholar] [CrossRef] [PubMed]
- Dahl, G.E.; Tao, S.; Laporta, J. Heat stress impacts immune status in cow across the life cycle. Front. Vet. Sci. 2020, 7, 116. [Google Scholar] [CrossRef] [PubMed]
- Davidson, B.D.; Dado-Senn, B.; Rosa Padilla, N.; Fabris, T.F.; Casarotto, L.T.; Ouellet, V.; Toledo, I.M.; Dahl, G.E.; Laporta, J. Late-gestation heat stress abatement in dairy heifers promotes thermoregulation and improves productivity. J. Dairy Sci. 2021, 104, 2357–2368. [Google Scholar] [CrossRef]
- Collier, R.J.; Doelger, S.G.; Head, H.H.; Thatcher, W.W.; Wilcox, C.J. Effects of heat stress during pregnancy on maternal hormone concentrations, calf birth weight and postpartum milk yield of Holstein cows. J. Anim. Sci. 1982, 54, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, J.; Wang, F.; Xiao, J.; Wang, Y.; Yang, H.; Li, S.; Cao, Z. Heat stress on calves and heifers: A review. J. Anim. Sci. Biotechnol. 2020, 11, 79. [Google Scholar] [CrossRef]
- Laporta, J.; Ferreira, F.C.; Ouellet, V.; Dado-Senn, B.; Almeida, A.K.; De Vries, A.; Dahl, G.E. Late-gestation heat stress impairs daughter and granddaughter lifetime performance. J. Dairy Sci. 2020, 103, 7555–7568. [Google Scholar] [CrossRef]
- Davidson, B.D.; Dado-Senn, B.; Ouellet, V.; Dahl, G.E.; Laporta, J. Effect of late-gestation heat stress in nulliparous heifers on postnatal growth, passive transfer of immunoglobulin G, and thermoregulation of their calves. JDS Commun. 2021, 2, 165–169. [Google Scholar] [CrossRef]
- Ouellet, V.; Boucher, A.; Dahl, G.E.; Laporta, J. Consequences of maternal heat stress at different stages of embryonic and fetal development on dairy cow’s progeny. Anim. Front. 2021, 11, 48–56. [Google Scholar] [CrossRef]
- Skibiel, A.L.; Peñagaricano, F.; Amorín, R.; Ahmed, B.M.; Dahl, G.E.; Laporta, J. In utero heat stress alters the offspring epigenome. Sci. Rep. 2018, 8, 14609. [Google Scholar] [CrossRef]
- Skibiel, A.L.; Dado-Senn, B.; Fabris, T.F.; Dahl, G.E.; Laporta, J. In utero exposure to thermal stress has long-term effects on mammary gland microstructure and function in dairy cattle. PLoS ONE 2018, 13, e0206046. [Google Scholar] [CrossRef]
- Tao, S.; Dahl, G.E. Invited review: Heat stress effect during late gestation on dry cows and their calves. J. Dairy Sci. 2013, 96, 4079–4093. [Google Scholar] [CrossRef]
- Tao, S.; Monteiro, A.P.A.; Hayen, M.J.; Dahl, G.E. Short communication: Maternal heat stress during the dry period alters postnatal whole-body insulin response of calves. J. Dairy Sci. 2014, 97, 897–901. [Google Scholar] [CrossRef] [PubMed]
- Dado-Senn, B.; Vega Acosta, L.; Torres Rivera, M.; Field, S.L.; Marrero, M.G.; Davidson, B.D.; Tao, S.; Fabris, T.F.; Ortiz-Colón, G.; Dahl, G.E.; et al. Pre- and postnatal heat stress abatement affects dairy calf thermoregulation and performance. J. Dairy Sci. 2020, 103, 4822–4837. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Wang, W.; Zheng, N.; Cheng, J.; Li, S.; Zhang, Y.; Wang, J. Identification of diagnostic biomarkers and metabolic pathway shifts of heat-stressed lactating dairy cows. J. Proteom. 2015, 125, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Zheng, N.; Wang, W.; Cheng, J.; Li, S.; Zhang, Y.; Wang, J. Integrated metabolomics study of the milk of heat-stressed lactating dairy cows. Sci. Rep. 2016, 6, 24208. [Google Scholar] [CrossRef]
- Fan, C.; Su, D.; Tian, H.; Li, X.; Li, Y.; Ran, L.; Hu, R.; Cheng, J. Liver metabolic perturbations of heat-stressed lactating dairy cows. Asian-Australas. J. Anim. Sci. 2018, 31, 1244–1251. [Google Scholar] [CrossRef]
- Monteiro, A.P.A.; Guo, J.-R.; Weng, X.-S.; Ahmed, B.M.; Hayen, M.J.; Dahl, G.E.; Bernard, J.K.; Tao, S. Effect of maternal heat stress during the dry period on growth and metabolism of calves. J. Dairy Sci. 2016, 99, 3896–3907. [Google Scholar] [CrossRef]
- National Research Council (NRC) (U.S.). Committee on Physiological Effects of Environmental Factors on Animals. In A Guide to Environmental Research on Animals; National Academy of Sciences: Washington, DC, USA, 1971. [Google Scholar]
- Brügemann, K.; Gernand, E.; König von Borstel, U.; König, S. Defining and evaluating heat stress thresholds in different dairy cow production systems. Arch. Anim. Breed. 2012, 55, 13–24. [Google Scholar] [CrossRef]
- Halli, K.; Brügemann, K.; Bohlouli, M.; König, S. Time-lagged and acute impact of heat stress on production and fertility traits the local dual-purpose cattle breed “Rotes Höhenvieh” under pasture-based conditions. Transl. Anim. Sci. 2020, 4, txaa148. [Google Scholar] [CrossRef]
- Kipp, C.; Brügemann, K.; Zieger, P.; Mütze, K.; Möcklinghoff-Wicke, S.; König, S.; Halli, K. Across-generation effects of maternal heat stress during late gestation on production, female fertility and longevity traits in dairy cows. J. Dairy Res. 2021, 88, 147–153. [Google Scholar] [CrossRef]
- Yin, T.; Halli, K.; König, S. Direct genetic effects, maternal genetic effects, and maternal genetic sensitivity on prenatal heat stress for calf diseases and corresponding genomic loci in German Holsteins. J. Dairy Sci. 2022, 105, 6795–6808. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, S.L.; Guggenbichler, W.; Weinberger, K.M.; Graber, A.; Stöggl, W.M. Device for Quantitative Analysis of a Metabolite Profile. Patent No. EP 1875401 B1, 2014. [Google Scholar]
- Ramsay, S.L.; Stöggl, W.M.; Weinberger, K.M.; Graber, A.; Guggenbichler, W. Apparatus for Analyzing a Metabolic Profile. Patent No. EP 1897014 B1, 2014. [Google Scholar]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.-E.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucl. Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, L.; Johansson, E.; Kettaneh-Wold, N.; Wold, S. Introduction to Multi and Megavariate Data Analysis Using Projection Methods (PCA & PLS); Umetrics AB: Umeå, Sweden, 1999. [Google Scholar]
- Moore, K.L.; Johnston, D.J.; Graser, H.-U.; Herd, R. Genetic and phenotypic relationships between insulin-like growth factor-I (IGF-I) and net feed intake, fat, and growth traits in Angus beef cattle. Aust. J. Agric. Res. 2005, 56, 211–218. [Google Scholar] [CrossRef]
- Tosi, M.R.; Tugnoli, V. Cholesteryl esters in malignancy. Clin. Chim. Acta 2005, 359, 27–45. [Google Scholar] [CrossRef] [PubMed]
- Schären, M.; Riefke, B.; Slopianka, M.; Keck, M.; Grundemann, S.; Wichard, J.; Brunner, N.; Klein, S.; Snedec, T.; Theinert, K.B.; et al. Aspects of transition cow metabolomics—Part III: Alterations in the metabolome of liver and blood throughout the transition period in cows with different liver metabotypes. J. Dairy Sci. 2021, 104, 9245–9262. [Google Scholar] [CrossRef]
- Cole, L.K.; Vance, J.E.; Vance, D.E. Phosphatidylcholine biosynthesis and lipoprotein metabolism. BBA-Mol. Cell Biol. L 2012, 1821, 754–761. [Google Scholar] [CrossRef]
- Slotte, J.P. Biological functions of sphingomyelins. Prog. Lipid Res. 2013, 52, 424–437. [Google Scholar] [CrossRef]
- Nilsson, Å.; Duan, R.-D. Absorption and lipoprotein transport of sphingomyelin. J. Lipid Res. 2006, 47, 154–171. [Google Scholar] [CrossRef]
- Slotte, J.P. Molecular properties of various structurally defined sphingomyelins. Correlation of structure with function. Prog. Lipid Res. 2013, 52, 206–219. [Google Scholar] [CrossRef] [PubMed]
- Noble, R.C.; O’Kelly, J.C.; Moore, J.H. Observations on changes in lipid composition and lecithin-cholesterol-acyl transferase reaction of bovine plasma induced by heat exposure. Lipids 1973, 8, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Kenéz, Á.; Dänicke, S.; Rolle-Kampczyk, U.; von Bergen, M.; Huber, K.A. A metabolomics approach to characterize phenotypes of metabolic transition from late pregnancy to early lactation in dairy cows. Metabolomics 2016, 12, 165. [Google Scholar] [CrossRef]
- Blum, J.W.; Hadorn, U.; Sallmann, H.-P.; Schuep, W. Delaying colostrum intake by one day impairs plasma lipid, essential fatty acid, carotene, retinol and ɑ-Tocopherol status in neonatal calves. J. Nutr. 1997, 127, 2024–2029. [Google Scholar] [CrossRef] [PubMed]
- Carroll, K.K.; Hamilton, R.M.G.; Macleod, G.K. Plasma cholesterol levels in suckling and weanded calves, lambs, pigs and colts. Lipids 1973, 8, 635–640. [Google Scholar] [CrossRef]
- Kenéz, Á.; Koch, C.; Korst, M.; Kesser, J.; Eder, K.; Sauerwein, H.; Huber, K. Different milk feeding intensities during the first 4 weeks of rearing dairy calves: Part 3: Plasma metabolomics analysis reveals long-term metabolic imprinting in Holstein heifers. J. Dairy Sci. 2018, 101, 8446–8460. [Google Scholar] [CrossRef]
- Almoosavi, S.M.M.S.; Ghoorchi, T.; Naserian, A.A.; Khanaki, H.; Drackley, J.K.; Ghaffari, M.H. Effects of late gestation heat stress independent of reduced feed intake on colostrum, metabolism at calving, and milk yield in early lactation dairy cows. J. Dairy Sci. 2021, 104, 1744–1758. [Google Scholar] [CrossRef]
- Nardone, A.; Lacetera, N.; Bernabucci, U.; Ronchi. B. Composition of colostrum from dairy heifers exposed to high air temperatures during late pregnancy and the early postpartum period. J. Dairy Sci. 1997, 80, 838–844. [Google Scholar] [CrossRef]
- Roelfzema, H.; Broekhuyse, R.M.; Veerkamp, J.H. Lipids in tissues of the eye. XII. Phospholipid and sphingomyelin concentrations in bovine lenses in relation to differentiation and ageing. Exp. Eye Res. 1976, 23, 409–415. [Google Scholar] [CrossRef]
- Trabado, S.; Al-Salameh, A.; Croixmarie, V.; Masson, P.; Corruble, E.; Fève, B.; Colle, R.; Ripoll, L.; Walther, B.; Boursier-Neyret, C.; et al. The human plasma-metabolome: Reference values in 800 French healthy volunteers; impact of cholesterol, gender and age. PLoS ONE 2017, 12, e0173615. [Google Scholar] [CrossRef]
- Yu, Z.; Zhai, G.; Singmann, P.; He, Y.; Xu, T.; Prehn, C.; Römisch-Margl, W.; Lattka, E.; Gieger, C.; Soranzo, N.; et al. Human serum metabolic profiles are age dependent. Aging Cell 2012, 11, 960–967. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Sato, T.; Nomoto, K.; Tsuji, H. Identification of phenol- and p-cresol-producing intestinal bacteria by using media supplemented with tyrosine and its metabolites. FEMS Microbiol. Ecol. 2018, 94, fiy125. [Google Scholar] [CrossRef] [PubMed]
- Roediger, W.E.W.; Babidge, W. Human colonocyte detoxification. Gut 1997, 41, 731–734. [Google Scholar] [CrossRef]
- Sun, C.-Y.; Li, J.-R.; Wang, Y.-Y.; Lin, S.-Y.; Ou, Y.-C.; Lin, C.-J.; Wang, J.-D.; Liao, S.-L.; Chen, C.-J. P-Cresol sulfate caused behavior disorders and neurodegeneration in mice with unilateral nephrectomy involving oxidative stress and neuroinflammation. Int. J. Mol. Sci. 2020, 21, 6687. [Google Scholar] [CrossRef]
- Liu, W.-C.; Tomino, Y.; Lu, K.-C. Impacts of indoxyl sulfate and p-Cresol sulfate on chronic kidney disease and mitigating effects of AST-120. Toxins 2018, 10, 367. [Google Scholar] [CrossRef]
- Yokoyama, M.T.; Carlson, J.R. Production of Skatol and para-Cresol by a rumen Lactobacillus sp. Appl. Environ. Microb. 1981, 41, 71–76. [Google Scholar] [CrossRef]
- Smith, H.W. The development of the flora of the alimentary tract in young animals. J. Pathol. Bacteriol. 1965, 90, 459–513. [Google Scholar] [CrossRef]
- Ziarno, M.; Godlewska, A. Znaczenie i wykorzystani bakterii rodzaju Lactoccocus w mleczarstwie. Med. Wet 2008, 64, 35–39. [Google Scholar]
- Puppel, K.; Golebiewski, M.; Konopka, K.; Kunowska-Slósarz, M.; Slósarz, J.; Grodkowski, G.; Przysucha, T.; Balcerak, M.; Madras-Majewska, B.; Sakowski, T. Relationship between the quality of colostrum and the formation of microflora in the digestive tract of calves. Animals 2020, 10, 1293. [Google Scholar] [CrossRef]
- Tirone, T.A.; Brunicardi, F.C. Overview of glucose regulation. World J. Surg. 2001, 25, 461–467. [Google Scholar] [CrossRef]
- Wheelock, J.B.; Rhoads, R.P.; VanBaale, M.J.; Sanders, S.R.; Baumgard, L.H. Effects of heat stress on energetic metabolism in lactating Holstein cows. J. Dairy Sci. 2010, 93, 644–655. [Google Scholar] [CrossRef] [PubMed]
- Ronchi, B.; Bernabucci, U.; Lacetera, N.G.; Nardone, A.; Bertoni, G. Effects of heat stress on metabolic status of Friesian heifer calves. Zootenica Nutr. Anim. 1995, 21, 209–220. [Google Scholar]
- O’Brien, M.D.; Rhoads, R.P.; Sanders, S.R.; Duff, G.C.; Baumgard, L.H. Metabolic adaptations to heat stess in growing cattle. Domest. Anim. Endicrol. 2010, 38, 86–94. [Google Scholar] [CrossRef]
- Omran, F.I.; Ashour, G.; Youssef, M.M.; Shafie, M.M. Responses of hematology, blood metabolites, mineral ions and hormonal profile to heat stress for Egyptian buffalo-calves. Egypt J. Agric. Res. 2011, 89, 1129–1140. [Google Scholar] [CrossRef]
- Kaldes, M.Z.N. Heat-Induced Biological Changes as Heat Tolerance Indices Related to Growth Performance in Buffaloes. Ph.D. Thesis, Animal Production Departement, Ain Shams University, Cairo, Egypt, 2004. [Google Scholar]
- Chaiyabuter, N.; Buranakarl, C.; Muangcharoen, V.; Loypetjra, P.; Pichaicharnarong, A. Effects of acute heat stress on changes in the rate of liquid flow from the rumen turnover of body water of swamp buffalo (Bubalus Bubalis). J. Agric. Sci. 1987, 108, 549–553. [Google Scholar] [CrossRef]
- Guo, J.-R.; Monteiro, A.P.A.; Weng, X.-S.; Ahmed, B.M.; Laporta, J.; Hayen, M.J.; Dahl, G.E.; Bernard, J.K.; Tao, S. Short communication: Effect of maternal heat stress in late gestation on blood hormones and metabolites of newborn calves. J. Dairy Sci. 2016, 99, 6804–6807. [Google Scholar] [CrossRef]
- Humer, E.; Aditya, S.; Zebeli, Q. Innate immunity and metabolic responses in dairy cows challenged intramammarily with lipopolysaccharide after subacute ruminal acidosis. Animal 2018, 12, 2551–2560. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, G. Insulin and insulin resistance. Clin. Biochem. Rev. 2005, 26, 19–39. [Google Scholar]
- Singh, K.; Molenaar, A.J.; Swanson, K.M.; Gudex, B.; Arias, J.A.; Erdman, R.A.; Stelwagen, K. Epigenetics: A possible role in acute and transgenerational regulation of dairy cow milk production. Animal 2011, 6, 375–381. [Google Scholar] [CrossRef]
- Jaenisch, R.; Bird, A. Epigenetic regulation of gene expression: How the genome integrates intrinsic and environmental signals. Nat. Genet. 2003, 33, 245–254. [Google Scholar] [CrossRef]
- Strahl, B.D.; Allis, C.D. The language of covalent histone modifications. Nature 2000, 403, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Skinner, M.K.; Manikkam, M.; Guerrero-Bosagna, C. Epigenetic transgenerational actions of environmental factors in disease etiology. Trends Enocrinol. Metab. 2010, 21, 214–222. [Google Scholar] [CrossRef] [PubMed]
- D’Urso, A.; Brickner, J.H. Mechanisms of epigenetic memory. Trends Genet. 2014, 30, 230–236. [Google Scholar] [CrossRef]
- Ju, C.; Tacke, F. Hepatic macrophages in homeostasis and liver diseases: From pathogenesis to novel therapeutic strategies. Cell. Mol. Immunol. 2016, 13, 316–327. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Source | Metabolite | Model | Abbreviation |
|---|---|---|---|
| Plasma Samples of calves | PC aa C34:1 | yij = µ + ci + eij | (1) |
| PC aa C38:3 | |||
| PC aa C38:4 | |||
| PC ae C38:3 | |||
| PC ae C40:4 | |||
| CE(20:3) | |||
| CE(20:4) | |||
| CE(22:5) | |||
| Hexoses | |||
| PC aa C38:1 | yijk = µ + ci + ccj + eijk | (2) | |
| p-Cresol-SO4 | |||
| SM C18:0 | yijk = µ + ci + aj + eijk | (3) | |
| SM C18:1 | |||
| SM C24:1 | |||
| PC ae C30:2 |
| Calf | mTHI Class | Age (d) | Time of Blood Sampling | Birthweight (kg) | Calving Condition |
|---|---|---|---|---|---|
| 2031 | MHSCALF | 3 | 11:40 | 43.0 | no birth assistance |
| 2037 | MHSCALF | 6 | 10:35 | 39.5 | no birth assistance |
| 2038 | MHSCALF | 1 | 10:37 | 42.5 | minor birth assistance or dystocia |
| 2039 | MHSCALF | 4 | 13:50 | 35.5 | no birth assistance |
| 2043 | MHSCALF | 5 | 11:22 | 39.0 | no birth assistance |
| 2044 | MHSCALF | 3 | 11:25 | 39.0 | no birth assistance |
| 2048 | MHSCALF | 4 | 10:30 | 37.0 | no birth assistance |
| 2052 | MHSCALF | 5 | 12:10 | 39.0 | no birth assistance |
| 2056 | MHSCALF | 7 | 11:10 | 39.0 | no birth assistance |
| 2059 | MHSCALF | 5 | 13:15 | 39.4 | no birth assistance |
| 2060 | MHSCALF | 3 | 13:10 | 37.0 | minor birth assistance or dystocia |
| 2064 | MHSCALF | 2 | 13:14 | 37.0 | no birth assistance |
| 2065 | MHSCALF | 2 | 13:15 | 38.5 | no birth assistance |
| 2068 | MHSCALF | 6 | 11:40 | 39.0 | no birth assistance |
| 2006 | NMHSCALF | 1 | 15:00 | 41.0 | no birth assistance |
| 2007 | NMHSCALF | 1 | 15:05 | 41.5 | no birth assistance |
| 2008 | NMHSCALF | 4 | 11:23 | 40.0 | no birth assistance |
| 2009 | NMHSCALF | 3 | 11:20 | 39.0 | no birth assistance |
| 2010 | NMHSCALF | 1 | 11:27 | 39.5 | minor birth assistance or dystocia |
| 2012 | NMHSCALF | 5 | 12:56 | 40.0 | no birth assistance |
| 2015 | NMHSCALF | 4 | 11:42 | 42.0 | no birth assistance |
| 2016 | NMHSCALF | 2 | 11:52 | 45.5 | no birth assistance |
| 2017 | NMHSCALF | 7 | 12:02 | 39.0 | no birth assistance |
| 2018 | NMHSCALF | 2 | 12:00 | 46.0 | no birth assistance |
| 2019 | NMHSCALF | 6 | 10:50 | 48.0 | no birth assistance |
| 2020 | NMHSCALF | 3 | 10:55 | 39.4 | minor birth assistance or dystocia |
| 2026 | NMHSCALF | 4 | 15:15 | 43.4 | no birth assistance |
| 2027 | NMHSCALF | 6 | 13:17 | 41.7 | no birth assistance |
| 2029 | NMHSCALF | 1 | 10:40 | 43.0 | no birth assistance |
| 2030 | NMHSCALF | 4 | 13:35 | 36.5 | no birth assistance |
| 2032 | NMHSCALF | 2 | 10:01 | 42.2 | minor birth assistance or dystocia |
| 2034 | NMHSCALF | 1 | 10:00 | 44.7 | no birth assistance |
| 2036 | NMHSCALF | 3 | 10:10 | 34.8 | minor birth assistance or dystocia |
| 2073 | NMHSCALF | 5 | 12:25 | 38.0 | no birth assistance |
| 2074 | NMHSCALF | 2 | 12:26 | 35.5 | no birth assistance |
| 2078 | NMHSCALF | 7 | 13:07 | 39.0 | N/A |
| 2080 | NMHSCALF | 6 | 13:06 | 35.0 | no birth assistance |
| 2085 | NMHSCALF | 4 | 12:20 | NA | no birth assistance |
| 2088 | NMHSCALF | 7 | 13:45 | 38.0 | no birth assistance |
| 2089 | NMHSCALF | 2 | 13:47 | 37.0 | no birth assistance |
| 2090 | NMHSCALF | 1 | 14:35 | 37.5 | N/A |
| 2102 | NMHSCALF | 1 | 11:32 | 38.5 | no birth assistance |
| 2104 | NMHSCALF | 6 | 14:20 | 42.5 | no birth assistance |
| 2107 | NMHSCALF | 7 | 11:45 | 42.0 | N/A |
| 2108 | NMHSCALF | 7 | 11:42 | 38.5 | N/A |
| 2109 | NMHSCALF | 6 | 11:39 | 39.5 | N/A |
| 2110 | NMHSCALF | 4 | 11:35 | 41.0 | N/A |
| Dam Group | Parities | Mean Dry off Weight of mp Dams (kg) | Mean Dry off Milk Yield of mp Dams (kg) | Mean Weight of pp Dams before Birth (kg) |
|---|---|---|---|---|
| Dams of MHSCALVES | 1–4 | 754.8 ± 18.6 | 25.1 ± 4.7 | 708.2 ± 49.9 |
| Dams of NMHSCALVES | 1–5 | 754.5 ± 87.3 | 23.3 ± 6.3 | 705.1 ± 62.7 |
| Metabolite | Birth Weight | p-Value |
|---|---|---|
| PC aa C34:1 | 0.15 | 0.3332 |
| PC aa C38:3 | 0.14 | 0.3543 |
| PC aa C38:4 | 0.22 | 0.1352 |
| PC ae C38:3 | 0.20 | 0.1869 |
| PC ae C40:4 | 0.26 | 0.0865 |
| CE(20:3) | 0.27 | 0.0669 |
| CE(20:4) | 0.26 | 0.0768 |
| CE(22:5) | 0.15 | 0.3233 |
| Hexoses | 0.24 | 0.1121 |
| PC aa C38:1 | 0.26 | 0.0772 |
| p-Cresol-SO4 | −0.08 | 0.6019 |
| SM C18:0 | 0.17 | 0.2481 |
| SM C18:1 | 0.14 | 0.3643 |
| SM C24:1 | 0.05 | 0.7256 |
| PC ae C30:2 | 0.18 | 0.2288 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Halli, K.; Cohrs, I.; Brügemann, K.; Koch, C.; König, S. A Pilot Study on Across-Generation Impacts of Maternal Heat Stress on Blood Metabolites of Female Holstein Dairy Calves. Metabolites 2023, 13, 494. https://doi.org/10.3390/metabo13040494
Halli K, Cohrs I, Brügemann K, Koch C, König S. A Pilot Study on Across-Generation Impacts of Maternal Heat Stress on Blood Metabolites of Female Holstein Dairy Calves. Metabolites. 2023; 13(4):494. https://doi.org/10.3390/metabo13040494
Chicago/Turabian StyleHalli, Kathrin, Imke Cohrs, Kerstin Brügemann, Christian Koch, and Sven König. 2023. "A Pilot Study on Across-Generation Impacts of Maternal Heat Stress on Blood Metabolites of Female Holstein Dairy Calves" Metabolites 13, no. 4: 494. https://doi.org/10.3390/metabo13040494
APA StyleHalli, K., Cohrs, I., Brügemann, K., Koch, C., & König, S. (2023). A Pilot Study on Across-Generation Impacts of Maternal Heat Stress on Blood Metabolites of Female Holstein Dairy Calves. Metabolites, 13(4), 494. https://doi.org/10.3390/metabo13040494