
Comparison of Plasma Metabolome Response to Diets Enriched in Soybean and Partially-Hydrogenated Soybean Oil in Moderately Hypercholesterolemic Adults-A Pilot Study

Abstract
1. Introduction
2. Methods and Materials
2.1. Study Participants, Design and Diet Intervention
2.2. Untargeted Metabolomics
2.2.1. Primary Metabolites, Complex Lipids and Biogenic Amines Extraction, Data Acquisition and Processing
2.2.2. Pathway Analysis
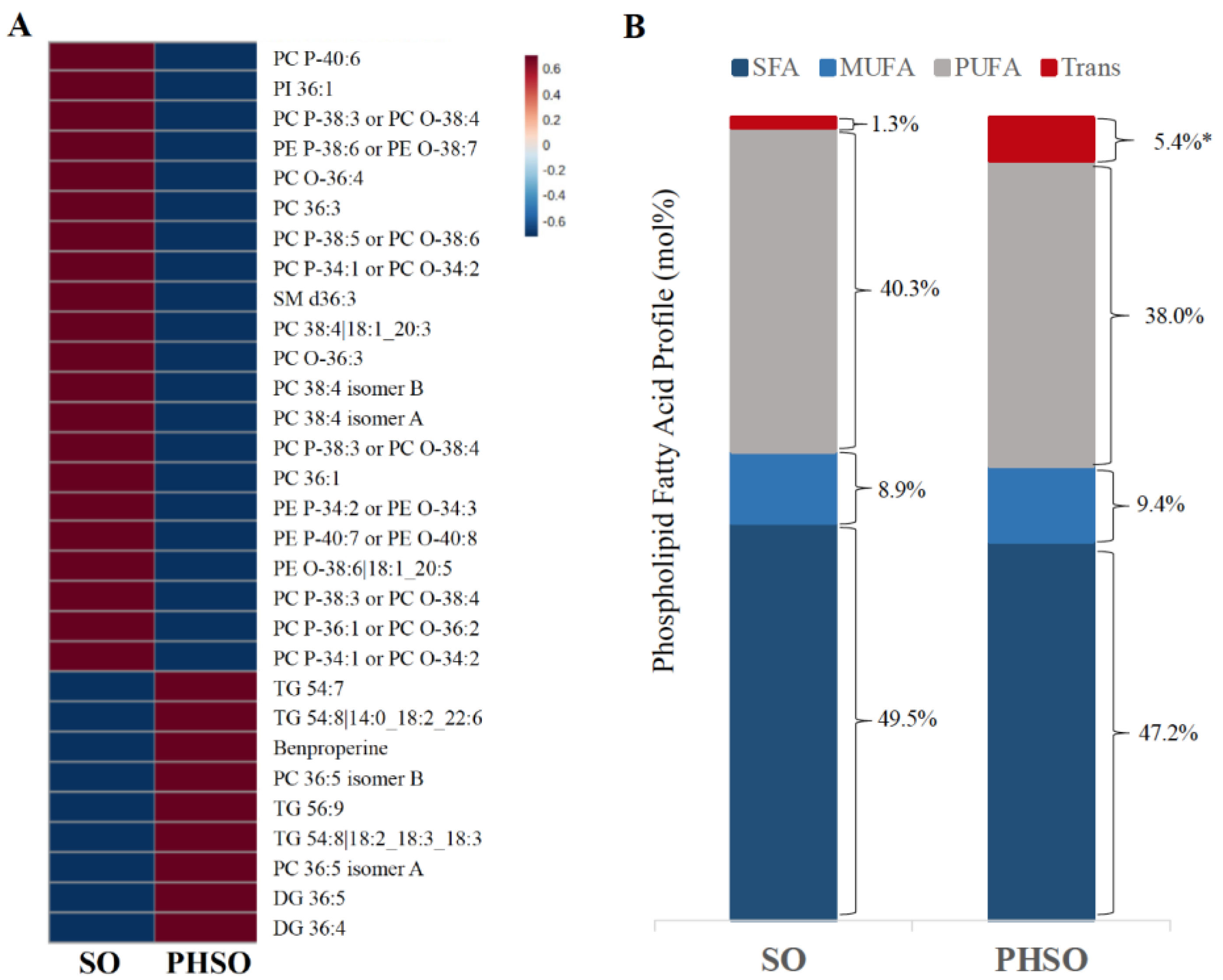
2.3. Plasma Lipid and Phospholipid Fatty Acid Profiles
2.4. Statistical Analysis
3. Results
3.1. Participant Characteristics and Serum Lipid and Phospholipid Fatty Acid Profiles after Consumption of SO and PHSO-Enriched Diets
3.2. Metabolite Profiling
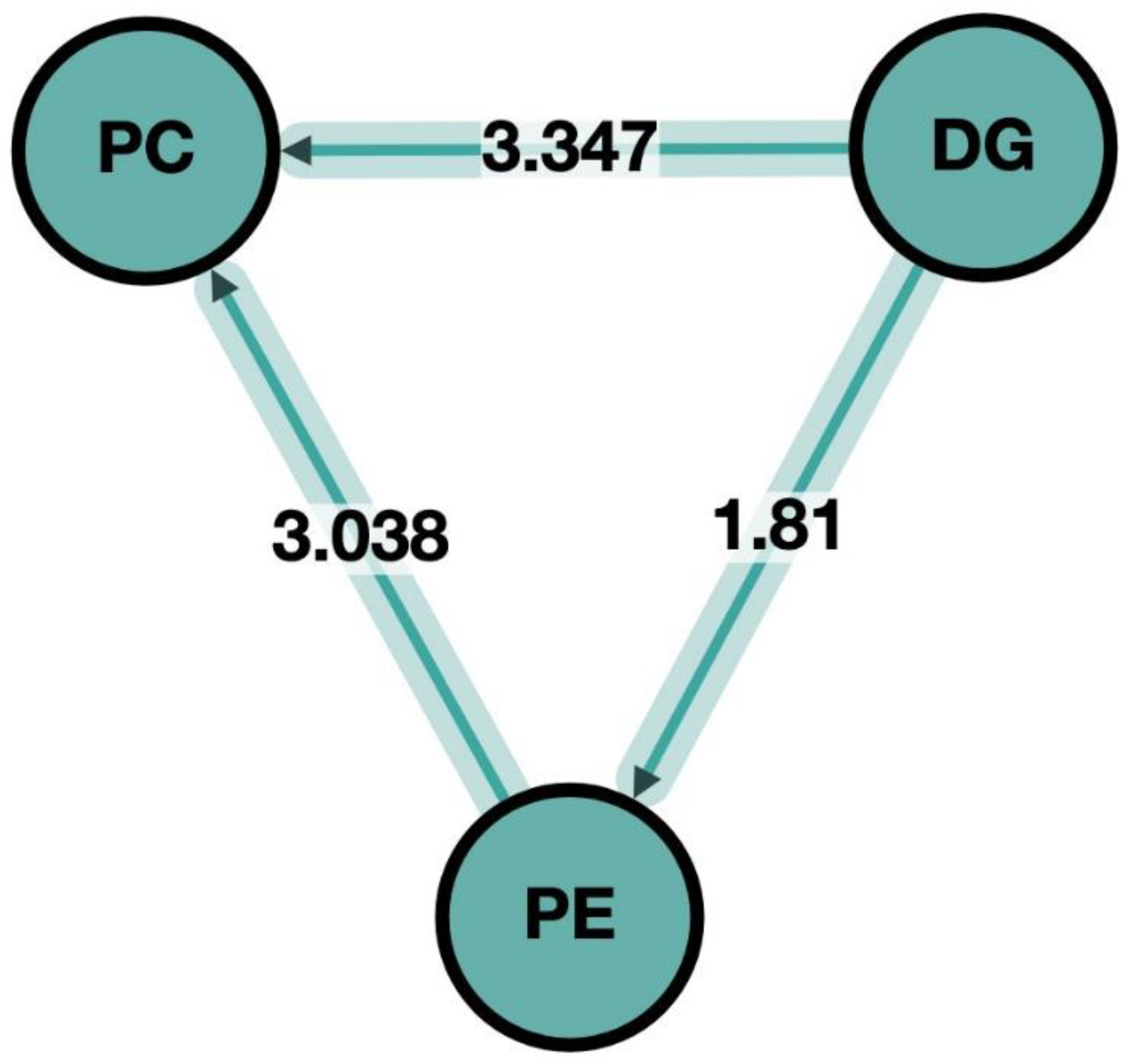
3.3. Pathway Analysis in Lipid Maps
3.4. Associations between Plasma Metabolites and Lipoprotein Fractions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Bingham, S.A. Biomarkers in nutritional epidemiology. Public Health Nutr. 2002, 5, 821–827. [Google Scholar] [CrossRef]
- Lichtenstein, A.H.; Appel, L.J.; Vadiveloo, M.; Hu, F.B.; Kris-Etherton, P.M.; Rebholz, C.M.; Sacks, F.M.; Thorndike, A.N.; Van Horn, L.; Wylie-Rosett, J.; et al. 2021 Dietary guidance to improve cardiovascular health: A scientific statement from the American Heart Association. Circulation 2021, 144, e472–e487. [Google Scholar] [CrossRef]
- Arnett, D.K.; Blumenthal, R.S.; Albert, M.A.; Buroker, A.B.; Goldberger, Z.D.; Hahn, E.J.; Himmelfarb, C.D.; Khera, A.; Lloyd-Jones, D.; McEvoy, J.W.; et al. 2019 ACC/AHA Guideline on the primary prevention of cardiovascular disease: A report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. Circulation 2019, 140, e596–e646. [Google Scholar] [CrossRef] [PubMed]
- Dayton, S.; Pearce, M.L.; Hashimoto, S.; Dixon, W.J.; Tomiyasu, U. A controlled clinical trial of a diet high in unsaturated fat in preventing complications of atherosclerosis. Circulation 1969, 40, II-1–II-63. [Google Scholar] [CrossRef] [PubMed]
- Leren, P. The Oslo Diet-Heart Study: Eleven-year report. Circulation 1970, 42, 935–942. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.N.; Ball, K.P.; Antonis, A.; Brigden, W.W.; Burns-Cox, C.J.; Fulton, M.; Haigh, M.; Hall, G.J.L.; Hanington, E.; Heady, J.A.; et al. Controlled trial of soya-bean oil in myocardial infarction. Lancet 1968, 2, 693–699. [Google Scholar] [CrossRef]
- Turpeinen, O.; Karvonen, M.J.; Pekkarinen, M.; Miettinen, M.; Elosuo, R.; Paavilainen, E. Dietary prevention of coronary heart disease: The Finnish Mental Hospital Study. Int. J. Epidemiol. 1979, 8, 99–118. [Google Scholar] [CrossRef]
- Miettinen, M.; Turpeinen, O.; Karvonen, M.J.; Pekkarinen, M.; Paavilainen, E.; Elosuo, R. Dietary prevention of coronary heart disease in women: The Finnish Mental Hospital Study. Int. J. Epidemiol. 1983, 12, 17–25. [Google Scholar] [CrossRef]
- Miettinen, M.; Turpeinen, O.; Karvonen, M.J.; Elosuo, R.; Paavilainen, E. Effect of cholesterol-lowering diet on mortality from coronary heart-disease and other causes: A twelve-year clinical trial in men and women. Lancet 1972, 2, 835–838. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Cobb, L.K.; Vesper, H.W.; Asma, S. Global surveillance of trans-fatty acids. Prev. Chronic Dis. 2019, 16, E147. [Google Scholar] [CrossRef] [PubMed]
- Ates, A.M.; Bukowski, M. Examining Record Soybean Oil Prices in 2021–22. Economic Research Service. Available online: https://www.ers.usda.gov/amber-waves/2022/december/examining-record-soybean-oil-prices-in-2021-22/ (accessed on 22 January 2023).
- Wanders, A.J.; Zock, P.L.; Brouwer, I.A. Trans fat intake and its dietary sources in general populations worldwide: A systematic review. Nutrients 2017, 9, 840. [Google Scholar] [CrossRef] [PubMed]
- Vega-López, S.; Ausman, L.M.; Jalbert, S.M.; Erkkilä, A.T.; Lichtenstein, A.H. Palm and partially hydrogenated soybean oils adversely alter lipoprotein profiles compared with soybean and canola oils in moderately hyperlipidemic subjects. Am. J. Clin. Nutr. 2006, 84, 54–62. [Google Scholar] [CrossRef] [PubMed]
- de Souza, R.J.; Mente, A.; Maroleanu, A.; Cozma, A.I.; Ha, V.; Kishibe, T.; Uleryk, E.; Budylowski, P.; Schünemann, H.; Beyene, J.; et al. Intake of saturated and trans unsaturated fatty acids and risk of all-cause mortality, cardiovascular disease, and type 2 diabetes: Systematic review and meta-analysis of observational studies. BMJ 2015, 351, h3978. [Google Scholar] [CrossRef] [PubMed]
- Rafiq, T.; Azab, S.M.; Teo, K.K.; Thabane, L.; Anand, S.S.; Morrison, K.M.; de Souza, R.J.; Britz-McKibbin, P. Nutritional metabolomics and the classification of dietary biomarker candidates: A critical review. Adv. Nutr. 2021, 12, 2333–2357. [Google Scholar] [CrossRef]
- O’Gorman, A.; Morris, C.; Ryan, M.; O’Grada, C.M.; Roche, H.M.; Gibney, E.R.; Gibney, M.J.; Brennan, L. Habitual dietary intake impacts on the lipidomic profile. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2014, 966, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Hustad, K.S.; Rundblad, A.; Ottestad, I.; Christensen, J.J.; Holven, K.B.; Ulven, S.M. Comprehensive lipid and metabolite profiling in healthy adults with low and high consumption of fatty fish: A cross-sectional study. Br. J. Nutr. 2021, 125, 1034–1042. [Google Scholar] [CrossRef] [PubMed]
- Azab, S.M.; de Souza, R.J.; Teo, K.K.; Anand, S.S.; Williams, N.C.; Holzschuher, J.; McGlory, C.; Philips, S.M.; Britz-McKibbin, P. Serum nonesterified fatty acids have utility as dietary biomarkers of fat intake from fish, fish oil, and dairy in women. J. Lipid Res. 2020, 61, 933–944. [Google Scholar] [CrossRef] [PubMed]
- McNairn, M.; Brito, A.; Dillard, K.; Heath, H.; Pantaleon, M.; Fanter, R.; Pilolla, K.; Amin, S.; La Frano, M.R. Postprandial dried blood spot-based nutritional metabolomic analysis discriminates a high-fat, high-protein meat-based diet from a high carbohydrate vegan diet: A randomized controlled crossover trial. J. Acad. Nutr. Diet. 2021, 121, 931–941 e932. [Google Scholar] [CrossRef]
- Brouwer-Brolsma, E.M.; Brennan, L.; Drevon, C.A.; van Kranen, H.; Manach, C.; Dragsted, L.O.; Roche, H.M.; Andres-Lacueva, C.; Bakker, S.J.L.; Bouwman, J. Combining traditional dietary assessment methods with novel metabolomics techniques: Present efforts by the Food Biomarker Alliance. Proc. Nutr. Soc. 2017, 76, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.S.; Kuo, C.H.; Yang, H.C.; Liang, Y.J.; Huang, C.J.; Sheen, L.Y.; Pan, W.H. Postprandial metabolomics response to various cooking oils in humans. J. Agric. Food Chem. 2018, 66, 4977–4984. [Google Scholar] [CrossRef]
- Gürdeniz, G.; Rago, D.; Bendsen, N.T.; Savorani, F.; Astrup, A.; Dragsted, L.O. Effect of trans fatty acid intake on LC-MS and NMR plasma profiles. PLoS ONE 2013, 8, e69589. [Google Scholar] [CrossRef] [PubMed]
- Guggisberg, D.; Burton-Pimentel, K.J.; Walther, B.; Badertscher, R.; Blaser, C.; Portmann, R.; Schmid, A.; Radtke, T.; Saner, H.; Fournier, N.; et al. Molecular effects of the consumption of margarine and butter varying in trans fat composition: A parallel human intervention study. Lipids Health Dis. 2022, 21, 74. [Google Scholar] [CrossRef]
- Lichtenstein, A.H.; Matthan, N.R.; Jalbert, S.M.; Resteghini, N.A.; Schaefer, E.J.; Ausman, L.M. Novel soybean oils with different fatty acid profiles alter cardiovascular disease risk factors in moderately hyperlipidemic subjects. Am. J. Clin. Nutr. 2006, 84, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Lichtenstein, A.H.; Ausman, L.M.; Carrasco, W.; Gualtieri, L.J.; Jenner, J.L.; Ordovas, J.M.; Nicolosi, R.J.; Goldin, B.R.; Schaefer, E.J. Rice bran oil consumption and plasma lipid levels in moderately hypercholesterolemic humans. Arter. Thromb. 1994, 14, 549–556. [Google Scholar] [CrossRef]
- Lichtenstein, A.H.; Ausman, L.M.; Jalbert, S.M.; Vilella-Bach, M.; Jauhiainen, M.; McGladdery, S.; Erkkilä, A.T.; Ehnholm, C.; Frohlich, J.; Schaefer, E.J. Efficacy of a Therapeutic Lifestyle Change/Step 2 diet in moderately hypercholesterolemic middle-aged and elderly female and male subjects. J. Lipid Res. 2002, 43, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Fiehn, O. Metabolomics by Gas Chromatography-Mass Spectrometry: Combined Targeted and Untargeted Profiling. Curr. Protoc. Mol. Biol. 2016, 114, 30.34.31–30.34.32. [Google Scholar] [CrossRef] [PubMed]
- Fiehn, O.; Kind, T. Metabolite profiling in blood plasma. Methods Mol. Biol. 2007, 358, 3–17. [Google Scholar] [CrossRef]
- Huang, N.K.; Matthan, N.R.; Matuszek, G.; Lichtenstein, A.H. Plasma Metabolite Profiles Following Consumption of Animal Protein and Soybean-Based Diet in Hypercholesterolemic Postmenopausal Women. Metabolites 2022, 12, 209. [Google Scholar] [CrossRef]
- Kind, T.; Liu, K.H.; Lee, D.Y.; DeFelice, B.; Meissen, J.K.; Fiehn, O. LipidBlast in silico tandem mass spectrometry database for lipid identification. Nat. Methods 2013, 10, 755–758. [Google Scholar] [CrossRef]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Oresic, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef]
- Fahy, E.; Sud, M.; Cotter, D.; Subramaniam, S. LIPID MAPS® online tools for lipid research. Nucleic Acids Res. 2007, 35, W606–W612. [Google Scholar] [CrossRef] [PubMed]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Matthan, N.R.; Ip, B.; Resteghini, N.; Ausman, L.M.; Lichtenstein, A.H. Long-term fatty acid stability in human serum cholesteryl ester, triglyceride, and phospholipid fractions. J. Lipid Res. 2010, 51, 2826–2832. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef] [PubMed]
- Saadatian-Elahi, M.; Slimani, N.; Chajes, V.; Jenab, M.; Goudable, J.; Biessy, C.; Ferrari, P.; Byrnes, G.; Autier, P.; Peeters, P.H.M.; et al. Plasma phospholipid fatty acid profles and their association with food intakes: Results from a cross -sectional study within the European prospective. Am. J. Clin. Nutr. 2009, 89, 331–346. [Google Scholar] [CrossRef] [PubMed]
- Meikle, P.J.; Barlow, C.K.; Mellett, N.A.; Mundra, P.A.; Bonham, M.P.; Larsen, A.; Cameron-Smith, D.; Sinclair, A.; Nestel, P.J.; Wong, G. Postprandial plasma phospholipids in men are influenced by the source of dietary fat. J. Nutr. 2015, 145, 2012–2018. [Google Scholar] [CrossRef]
- Hayes, K.C. Synthetic and modified glycerides: Effects on plasma lipids. Curr. Opin. Lipidol. 2001, 12, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Niu, S.L.; Mitchell, D.C.; Litman, B.J. Trans fatty acid derived phospholipids show increased membrane cholesterol and reduced receptor activation as compared to their cis analogs. Biochemistry 2005, 44, 4458–4465. [Google Scholar] [CrossRef] [PubMed]
- Matthan, N.R.; Welty, F.K.; Barrett, P.H.; Harausz, C.; Dolnikowski, G.G.; Parks, J.S.; Eckel, R.H.; Schaefer, E.J.; Lichtenstein, A.H. Dietary hydrogenated fat increases high-density lipoprotein apoA-I catabolism and decreases low-density lipoprotein apoB-100 catabolism in hypercholesterolemic women. Arter. Thromb. Vasc. Biol. 2004, 24, 1092–1097. [Google Scholar] [CrossRef]
- Yoon, Y.J.; Han, Y.M.; Choi, J.; Lee, Y.J.; Yun, J.; Lee, S.K.; Lee, C.W.; Kang, J.S.; Chi, S.W.; Moon, J.H.; et al. Benproperine, an ARPC2 inhibitor, suppresses cancer cell migration and tumor metastasis. Biochem. Pharm. 2019, 163, 46–59. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Variables | SO-Enriched Diet | PHSO-Enriched Diet |
|---|---|---|
| Total cholesterol, mmol/L | 5.3 ± 0.77 | 5.6 ± 0.76 * |
| LDL-C, mmol/L | 3.4 ± 0.53 | 3.6 ± 0.55 * |
| HDL-C, mmol/L | 1.3 ± 0.26 | 1.4 ± 0.29 |
| VLDL-C, mmol/L | 0.62 ± 0.34 | 0.63 ± 0.36 |
| TG, mmol/L | 1.7 ± 0.97 | 1.7 ± 0.86 |
| TC/HDL-C | 4.12 | 4.22 |
| LDL-C/HDL-C | 2.62 | 2.72 |
| Metabolites | Fold | VIP Score 2 | ROC Curve | ||
|---|---|---|---|---|---|
| Change 1 | FDR | AUC 3 | p-Value | ||
| TG 56:9 | 2.35 | 5.42 × 10−4 | 3.51 | 1.00 | 7.55 × 10−6 |
| TG 54:8 (TG 18:2_18:3_18:3) | 3.97 | 3.80 × 10−4 | 3.07 | 0.96 | 3.30 × 10−4 |
| Benproperine | 2.23 | 6.70 × 10−4 | 3.06 | 0.93 | 3.45 × 10−4 |
| DG 36:5 | 2.01 | 6.20 × 10−5 | 2.76 | 0.90 | 1.97 × 10−3 |
| TG 48:5 | 2.91 | 9.24 × 10−3 | 2.59 | 0.87 | 4.31 × 10−3 |
| TG 54:7 | 2.92 | 7.52 × 10−4 | 2.93 | 0.87 | 7.93 × 10−4 |
| TG 54:6 (TG 18:2_18:2_18:2) | 2.03 | 6.05 × 10−3 | 2.40 | 0.78 | 9.57 × 10−3 |
| Plasma Metabolite | Diet | VLDL | LDL | HDL |
|---|---|---|---|---|
| TG 56:9 | SO PHSO | 0.639 * 0.286 | 0.267 0.793 * | −0.358 −0.074 |
| TG 54:8 (TG 18:2_18:3_18:3) | SO PHSO | 0.375 0.464 | 0.063 0.603 | −0.087 −0.240 |
| TG 48:5 | SO PHSO | 0.192 0.005 | −0.444 −0.348 | −0.509 −0.143 |
| TG 54:7 | SO PHSO | 0.398 0.486 | 0.120 0.547 | −0.080 −0.245 |
| TG 54:6 (TG 18:2_18:2_18:2) | SO PHSO | 0.379 0.410 | −0.055 0.357 | −0.345 −0.277 |
| DG 36:5 | SO PHSO | 0.054 0.087 | −0.090 0.255 | −0.046 −0.049 |
| Benproperine | SO PHSO | 0.772 * 0.580 | 0.069 0.038 | −0.065 −0.180 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, N.K.; Lichtenstein, A.H.; Matuszek, G.; Matthan, N.R. Comparison of Plasma Metabolome Response to Diets Enriched in Soybean and Partially-Hydrogenated Soybean Oil in Moderately Hypercholesterolemic Adults-A Pilot Study. Metabolites 2023, 13, 474. https://doi.org/10.3390/metabo13040474
Huang NK, Lichtenstein AH, Matuszek G, Matthan NR. Comparison of Plasma Metabolome Response to Diets Enriched in Soybean and Partially-Hydrogenated Soybean Oil in Moderately Hypercholesterolemic Adults-A Pilot Study. Metabolites. 2023; 13(4):474. https://doi.org/10.3390/metabo13040474
Chicago/Turabian StyleHuang, Neil K., Alice H. Lichtenstein, Gregory Matuszek, and Nirupa R. Matthan. 2023. "Comparison of Plasma Metabolome Response to Diets Enriched in Soybean and Partially-Hydrogenated Soybean Oil in Moderately Hypercholesterolemic Adults-A Pilot Study" Metabolites 13, no. 4: 474. https://doi.org/10.3390/metabo13040474
APA StyleHuang, N. K., Lichtenstein, A. H., Matuszek, G., & Matthan, N. R. (2023). Comparison of Plasma Metabolome Response to Diets Enriched in Soybean and Partially-Hydrogenated Soybean Oil in Moderately Hypercholesterolemic Adults-A Pilot Study. Metabolites, 13(4), 474. https://doi.org/10.3390/metabo13040474