Abstract
Calcium (Ca) represents about 40% of the total mineral mass, mainly in the bone, providing mechanical strength to the skeleton and teeth. An adequate Ca intake is necessary for bone growth and development in children and adolescents and for maintaining bone mineral loss in elderly age. Ca deficiency predisposes to osteopenia and osteoporosis. Healthy nutrition, including an adequate intake of Ca-rich food, is paramount to prevent and cure osteoporosis. Recently, several clinical studies have demonstrated that, in conditions of Ca dysmetabolism, Ca-rich mineral water is beneficial as a valuable source of Ca to be used as an alternative to caloric Ca-rich dairy products. Although promising, these data have been collected from small groups of participants. Moreover, they mainly regard the effect of Ca-rich mineral water on bone metabolism. In contrast, an investigation of the effect of Ca supplementation on systemic metabolism is needed to address the spreading of systemic metabolic dysfunction often associated with Ca dysmetabolism. In the present study, we analyzed urine and blood sera of 120 women in perimenopausal condition who were subjected for six months to 2l daily consumption of bicarbonate–calcium mineral water marketed under ®Lete. Remarkably, this water, in addition to being rich in calcium and bicarbonate, is also low in sodium. A complete set of laboratory tests was carried out to investigate whether the specific water composition was such to confirm the known therapeutic effects on bone metabolism. Second, but not least, urine and blood sera were analyzed using NMR-based metabolomic procedures to investigate, other than the action on Ca metabolism, potential system-wide metabolic effects. Our data show that Lete water is a valid supplement for compensating for Ca dysmetabolism and preserving bone health and integrity.
1. Introduction
Bone mass and density are determined by genetics, hormones, physical activity, and nutrition. While genetic factors are important in growth and peak bone development, an adequate intake of bone nutrients is the main factor for optimal genetic potential expression and bone maintenance during adulthood [1]. An optimal Calcium (Ca) intake is necessary for bone health at all stages of life: for bone growth and development in children and adolescents and for maintaining bone mineral loss in elderly age [2]. Accordingly, dietary requirements for Ca are determined by the need for bone development and maintenance: higher during childhood and adolescence, during pregnancy and lactation, and for the elderly. The recommended dietary allowance for Ca varies between 700 and 1200 mg/day throughout life, as stated both at the international level by the United States Department of Agriculture (USDA), and at the Italian level by the Reference Levels of Nutrients and Energy Intake for the Italian population (LARN) [3].
Ca represents about 40% of the total mineral mass, mainly in the bone where, combined with phosphorus in hydroxyapatite crystals Ca10(PO4)6(OH)2, it provides mechanical strength to the skeleton and teeth [4]. Ca is unique among nutrients in that the body’s reserve is also functional: increasing bone mass is linearly related to reducing fracture risk. The remaining roles of body Ca in metabolism serve as a signal for vital physiological processes, including vascular contraction, blood clotting, muscle contraction, and nerve transmission [5].
Most of the Ca, not stored in the bones, is present in the blood serum as an ion. The concentration of serum ionized Ca is regulated in healthy subjects by the action of calciotropic hormones: parathyroid hormone (PTH), 1,25-dihydroxyvitamin D [1,25(OH)2D], Fibroblast Growth Factor 23 (FGF23), and calcitonin A, the decrease of which in serum Ca concentration increases the secretion of PTH, favoring the release of Ca from the bone and the Ca tubular reabsorption, thus reducing urinary Ca excretion. At the same time, PTH stimulates the secretion of 1,25(OH)2D in the kidneys, which favors active Ca absorption in the gut. Increases in serum Ca are reversed by calcitonin, whose action is controlled by 1,25(OH)2D and PTH secretion. FGF23 controls the phosphate serum levels and, therefore, indirectly also calcemia [6,7].
Ca deficiency predisposes to pathologies such as osteopenia, osteoporosis, kidney stones, cancer, hypertension, obesity, and insulin resistance [8,9,10]. Alteration in the Ca metabolism is frequent in women after menopause, reflecting systemic metabolic changes which occur in response to a deficit in hormonal estrogen regulation. As a result, approximately 30% of all postmenopausal women suffer from osteopenia or osteoporosis, a pathological condition characterized by decreased bone tissue density [11]. As a manifestation of a systemic metabolic condition, osteoporosis is frequently associated with other pathological metabolic conditions such as hyperglycemia, high blood pressure, and high BMI. These symptoms, typical of the metabolic syndrome and results of a systemic inflammatory condition, may worsen osteoporosis prognosis and increase the risk of bone fracture [12].
The control of Ca metabolism for the diagnosis of osteoporosis is based on the measurement of the bone mineral density; however, blood and urine laboratory tests are necessary to have integrative information on the health status of the bone and the bioavailability of Ca in response to dietary or pharmacological treatments. Based on the players involved in Ca metabolism, the concentrations of the following serum and urinary metabolites are considered: calcium, phosphate, magnesium, total proteins, PTH, 1,25(OH)2D (vitamin D), bone alkaline phosphatase, and beta cross laps; moreover, a complete urine test of fasting morning and urinary Ca as well as phosphate excretion is conducted and evaluated by 24 h urine collection [13].
The first intervention to prevent and cure osteoporosis is based on a healthy lifestyle, including correct nutrition with an adequate Ca intake (1000–1500 mg/day), and regular physical activity [14]. Despite that, food consumption surveys indicate that the diet in the West is generally poor in products rich in Ca since milk and its derivatives are considered fat and hypercaloric foods. To reduce the trend of low Ca intake, leading health authorities, such as the World Health Organization (WHO), recommend mineral water rich in Ca as a good source of Ca, easily assimilated and an alternative to caloric dietary products [3,15,16,17].
Water is the main constituent of the human body and is involved in many bodily functions, including being the carrier of nutrients to reach biological fluids and the primary vehicle to eliminate waste and toxins [18]. According to the National Recommended Energy and Nutrient Intake Levels (LARN) for water, a 1.2 to 2.5 L daily intake is essential to maintain body water equilibrium. However, needs may vary among people, depending on age, physical activity, personal circumstances, and weather conditions. The USDA reports that food, beverages, and drinking water intake should represent 2.7 L per day in women aged 19–50 [19]. Natural mineral waters are classified with respect to the mineral content. In particular, the 2009/54/EC directive established that “Water with Calcium” is that which includes Ca content > 150 mg/L [20]. Several clinical studies have been carried out to evaluate the possibility of using Ca from mineral water as a dietary supplement for Ca deficiency [6,21,22,23]. By measuring Ca and Ca-related metabolite concentrations in blood and urine, Ca from mineral water is demonstrated to be comparable or higher than that of Ca taken with dairy products [23,24,25,26].
These studies, in support of Ca-rich mineral waters as a valid integration for Ca deficiency, are conducted on small groups of participants. Moreover, they are exclusively focused on the effect of Ca-rich mineral water on Ca absorption and bone metabolism [27,28,29]. On the other hand, several metabolic changes are demonstrated to be associated with Ca deficit and bone dysmetabolism; therefore, data regarding a potential effect of Ca-rich mineral water on systemic metabolism are missing. To bridge this gap, in the present study, we analyzed urine and blood sera of 120 women in perimenopausal condition who were subjected for six months to 2l daily consumption of Ca-rich mineral water marketed under ®Lete. Remarkably, this water, in addition to being rich in Ca and bicarbonate, is also poor in sodium.
A complete set of laboratory tests was carried out to investigate whether the specific composition of ®Lete was such to confirm the therapeutic effects on bone metabolism previously observed for other bicarbonate–calcium waters. Second, in this study, we aimed to investigate possible system-wide metabolic effects occurring due to taking a bicarbonate–calcium water poor in sodium, other than the effect on Ca metabolism. In addition to the laboratory tests, the urine and blood of the women recruited in this study were analyzed using NMR-based metabolomic procedures.
Metabolomics is the large-scale study of metabolites within biological fluids, describing a fingerprint of the metabolic conditions of an organism. NMR spectroscopy combined with high-resolution mass spectrometry (HRMS) is a robust technique for metabolomic analysis [30,31]. Our NMR-based metabolomic analysis allowed for the observation of the metabolic changes associated with Ca-rich water treatment, which include, but are not limited to, Ca metabolism. Our data show that water rich in Ca and poor in sodium is a valid supplement to compensate for Ca dysmetabolism and preserve bone health and integrity. The improvement of Ca homeostasis is associated with the compensation of biochemical parameters related to systemic dysmetabolism, thus suggesting a beneficial effect in controlling eventual systemic inflammation.
2. Materials and Methods
2.1. Participants
A total of 120 perimenopausal women were recruited at the Avellino (Italy) Azienda Ospedaliera “San Giuseppe Moscati” between June 2021 and April 2022. Their demographics are reported in Table 1.

Table 1.
Demographics and clinical information of the participants.
2.2. Inclusion and Exclusion Criteria
The inclusion criteria for participants were female sex, aged 40–65, and 19–30 Kg/m2 body mass index (BMI). The exclusion criteria were the inclusion in other study protocols, pharmacological therapies in progress (estrogen-replacement hormone therapy, therapy for osteoporosis, corticosteroid therapy, insulin therapy), Ca supplements, presence of medium–severe renal and hepatic diseases, and documented severe osteoporosis (T-score MOC < −2.5).
2.3. Study Design
The women were randomly and in a blinded manner divided into the intervention group (79 participants) and the control group (41 participants). The institutional Ethics Committee CAMPANIA NORD for clinical trials and bio-medical research located at Azienda Ospedaliera “San Giuseppe Moscati,” Avellino, Italy, approved the study protocol (Prot. N.: NKCL2101; reg. N. CECN/1522), which followed the Declaration of Helsinki, according to the International Guidelines of Good Clinical Practice and the regulations of clinical trials. Informed written consent was obtained from participants after providing information about the nature, purpose, and procedures of the study (ClinicalTrials.gov Identifier: NCT05854342, University of Salerno Protocol Record CECN/1522).
2.4. Clinical Assessment and Intervention
All the participants were subjected to chemical and biochemical laboratory tests. Blood samples were analyzed in the clinical laboratory using automated analyzers and available commercial kits. Quantitative evaluation of the following clinical parameters was conducted for all participants at T0 and T6: glycemia (GLIC); creatinine (CREA); sodium (Na); calcium (Ca); alkaline phosphatase (ALP); cholesterol (COL); high-density lipoprotein (HDL); low-density lipoprotein (LDL); phosphorus (P); magnesium (Mg); urine creatinine (CREA U); urinary sodium (U Na); urinary calcium (U Ca); urinary phosphorus (U P); urinary magnesium (U Mg); creatinine clearance (CL CR); albumin (ALB); parathormone (PTH); osteocalcin (OSTEO); vitamin D (1,25(OH)2D); insulinemia (INSUL); pH; specific weight (SPEC W); bicarbonate (HCO3−); ionized calcium (Ca++). In addition, the BMI at t0 was calculated as indicated in the inclusion and exclusion criteria of this study.
2.5. Dietary Assessment and Intervention
The 121 participants were tested before (T0) and after six months (T6) of treatment. Firstly, each patient was given a food frequency questionnaire (FFQ) to assess Ca consumption [32]. The Ca intake for each food was calculated by multiplying the intake of mg of Ca/100 g of the product with the number of times it was taken per week. According to LARN recommendations, the daily Ca intake of a woman between 40 and 65 years is between 800 and 1200 mg/day [8]. The participants’ average Ca intake before treatment was 901.41 mg/day for the intervention group and 920.37 mg/day for the control group. After that, they were subjected to clinical nutritional evaluation. Nutrition experts evaluated the patient’s nutritional habits at T0. From the nutritional analysis of 120 participants, a Mediterranean diet emerged (Supplementary Table S1). After the initial assessment, experts provided dietary recommendations, suggesting the consumption of low-index glycemic carbohydrates, 1.0 g/pro kilo per day proteins, and a high content of unsaturated lipids. Deprivation of acidic foods and controlled Ca intake <700 mg/day was recommended. Then, they were subjected to clinical nutritional evaluation and provided with dietary recommendations, suggesting the consumption of low-index glycemic carbohydrates, 1.0 g/pro kilo per day proteins, and a high content of unsaturated lipids. Deprivation of acidic foods and controlled Ca intake <700 mg/day was recommended.
The intervention-group women were recommended to drink 2 L of bicarbonate–calcic mineral water daily, while the control-group women were recommended to drink 2 L of CO2-added oligomineral water. The water was suggested to be consumed as 500 mL with meals and the remaining 1500 mL during the day. The experimental water selected for this study is the natural bicarbonate–calcic sparkling mineral water Lete (Acqua Lete®; Società Generale delle Acque Minerali, Pratella, CE, Italy), shipped directly to the testing laboratory from its bottling facility. The chemical composition of the water is reported in Table 2.
Table 2.
Bicarbonate–calcic composition.
2.6. Sample Pre-Treatment for NMR Analysis
Serum and urinary samples for metabolomic analysis were collected following the standard operating procedure (SOP) for metabolomic-grade serum samples [33]. First, 5 mm heavy-walled NMR tubes were prepared using 200 µL of serum sample added to 300 µL of buffer phosphate (0.075 M Na2HPO4∙7H2O, 4% NaN3 in H2O used as preservative and 3-(trimethylsilyl)-2,2,3,3-tetradeuteropropionic acid, sodium salt (TSP-d4) used as an internal reference for the alignment and quantification of NMR spectra signals) at pH 7.4. A total of 1.5 mL of 24 h urine was previously centrifuged at 15.000× g for 10 min at 4 °C to pellet any particulates in the sample, and then the urine was filtered using a 0.2 µm filter. A total of 500 μL of urine was transferred into a new tube and mixed with 50 μL of 50 mM phosphate buffer in 99.8% D2O [33,34].
2.7. NMR Data Acquisition and Processing
NMR experiments were conducted at 298 K on a Bruker Avance 600 MHz spectrometer equipped with a 5 mm triple-resonance z-gradient. NMR spectra were processed and visualized using Topspin 3.2 (Bruker Biospin Bruker Biospin, Fällanden, Switzerland). 1D-NOESY experiments were conducted using the excitation sculpting pulse sequence for water suppression. The experiments’ results were collected using a 14 ppm sweep width, 192 transients of 16 complex points, and an acquisition time of 4 s transient and 60 ms mixing time. For identifying and quantifying metabolites, we used the online software Bayesil, (http://bayesil.ca, accessed on 1 September 2023) [35], which automatically processes and analyzes monodimensional 1H-NMR spectra of ultra-filtered biological samples. The glucose duplet signal at 5.4 ppm was used as a chemical shift reference, whereas the quantification was based on the peak intensity of TSP-d4, used as an internal reference compound.
2.8. Statistical Analysis
Biochemical parameters were analyzed using R-package 4.2.2 applying t-test and considering the parameters with p-value < 0.05 as significant [36]. The samples’ metabolomics data were normalized using sum, log-transformed, and Pareto-scaled and analyzed using MixOmics R-package (mixOmics-package) by the multivariate unsupervised method: principal component analysis (PCA) and multivariate supervised partial least-squares discriminant analysis (PLS-DA) [37]. A sample plot was used to observe the clustering of metabolomic profiles of the samples. In the graph, samples are represented as points positioned based on their projection on the selected latent components of the data. The correlations between the variables were plotted using a circular correlation plot obtained with the R Mix Omics package [37]. Data clustering was confirmed by the approach of distance metrics calculated by the centroid method, maximum distance, and Mahalanobis distance (Supplementary Figures S1–S4). The contribution of each variable for each component is represented in the loading plot, where the bar length corresponds to the importance of the variable in the construction of the given component, which can be positive or negative [37]. The analysis of the pathways was carried out by analysis with the Enrichment tool. The output of Metaboanalyst 5.0 was examined, and the KEGG paths [38] were chosen according to the lower false discoveries (FDR), p-value < 0.05, and the hits value related to the number of metabolites belonging to the pathway, >1. For a more precise visualization of common metabolites between the two biofluids, a Venn diagram was constructed using Interact Venn www.interactivenn.net (accessed on 20 August 2023) [39]. OmicsNet https://www.omicsnet.ca, accessed on 1 September 2023, was used for the creation of biological network and pathway prediction. The matrices of serum and urinary metabolites’ concentration, expressed by KEGG code, and the relative fold change value were analyzed by Omics-Net. The prediction of the pathways involved in the metabolomic profile was made by setting the Function Explorer analysis using the KEGG database [40].
3. Results
3.1. Clinical Analysis
The participants’ average Ca intake before treatment was 901.41 mg/day for the intervention group and 920.37 mg/day for the control group.
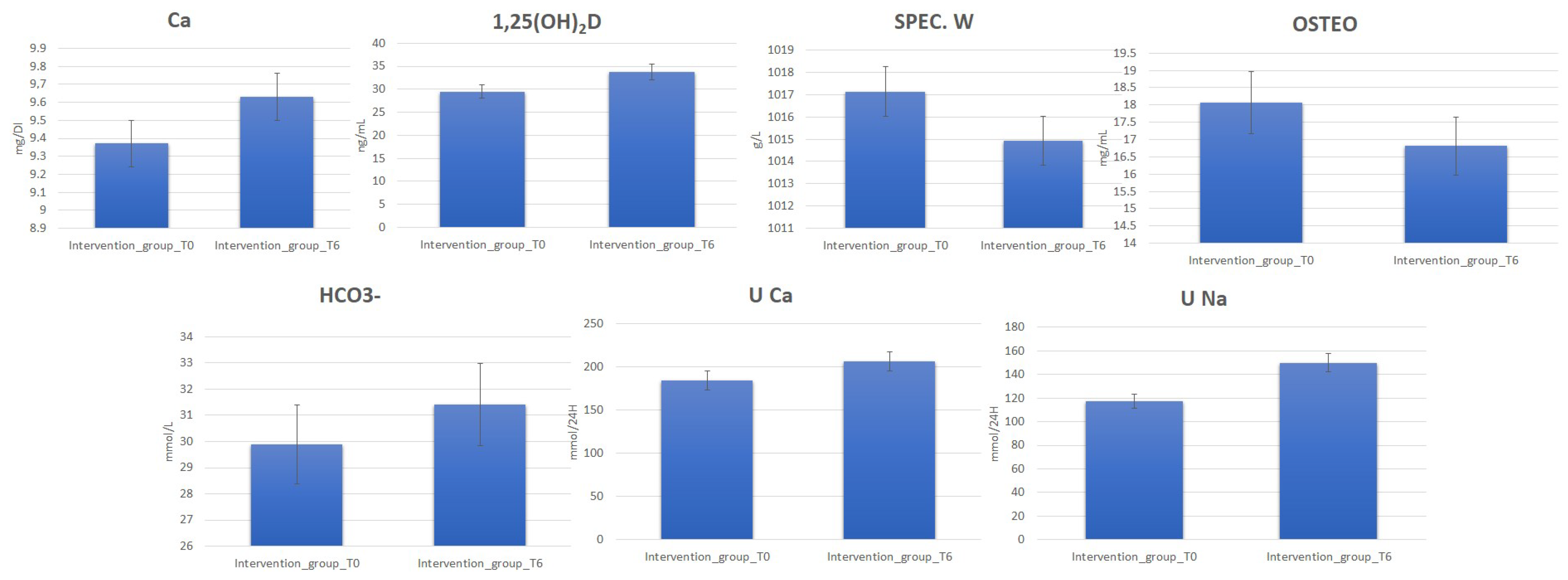
Clinical data deriving from blood and urine laboratory tests for 120 perimenopause women belonging to the intervention and control groups before (T0) (Supplementary Table S2) and after 6 months (T6) of water diet–therapy treatment were statistically analyzed using t-test to assess their significance (p-value). The analysis reporting clinical parameters indicates an increase in the urinary excretion of Na and Ca. Moreover, increased 1,25(OH)2D, Ca, and HCO3− blood values were detected for most subjects in the intervention group at T6. On the contrary, a decrease in OSTEO and urinary-specific weight (SPEC. W) was observed (Figure 1, Supplementary Table S3).
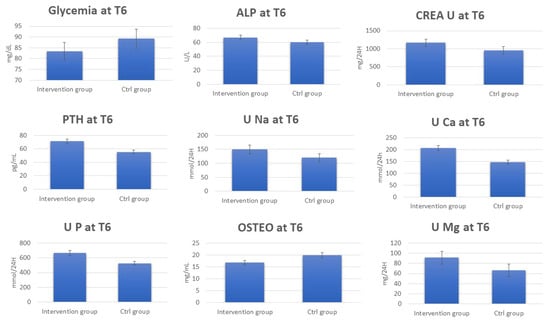
Figure 1.
Change in blood and urine concentrations of several important biomarkers of women subjected to 6 months of Ca–bicarbonate water diet–therapy treatment and women subjected to CO2-supplemented oligomineral water consumption for 6 months.
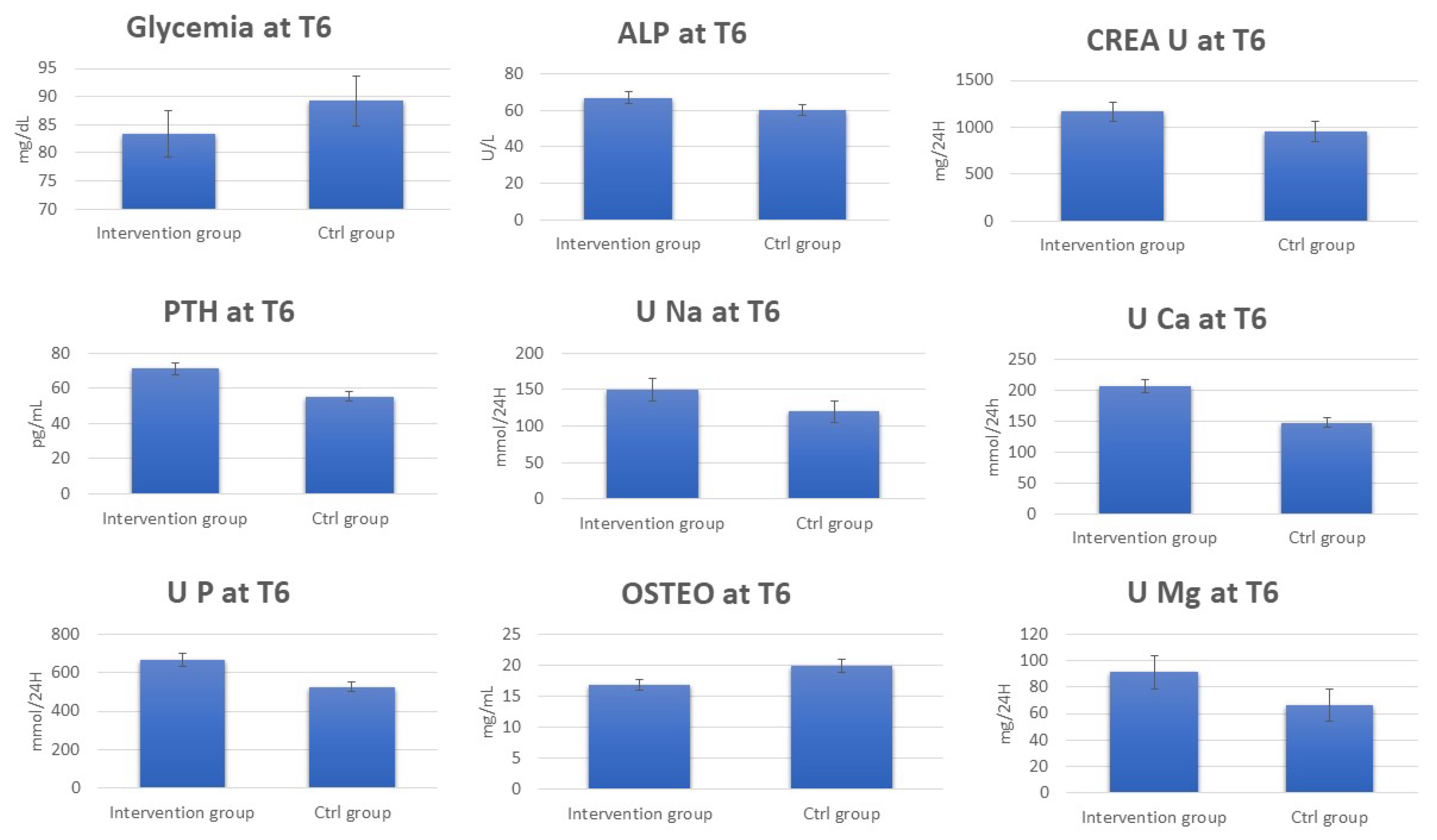
The data regarding the laboratory tests for the patients in the control group show the absence of any significant change in the urinary and blood parameters at T6 (Supplementary Tables S3 and S4). Interestingly, by comparing serum and urinary laboratory test data for the intervention and control group at T6, significant increases in the concentrations were observed for PTH, ALP, CREA U, U Ca, U Na, U P, U Mg, and Ca2+ in the intervention group at T6. On the other hand, OSTEO and glycemia decreased (Figure 2, Supplementary Table S5).
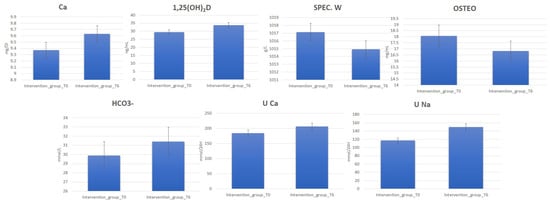
Figure 2.
Change in blood and urine concentrations of several important biomarkers of women subjected to 6 months of Ca–bicarbonate-rich water diet–therapy treatment. Clinical parameters are collected at T0 and T6 (p-value < 0.05).
3.2. Multivariate Statistical and Pathway Analysis
Data matrices, including the urine and serum metabolites’ concentrations derived from NMR analysis, were analyzed using a multivariate PCA unsupervised approach and PLS-DA supervised approach (R package: MixOmics [37]) [41]. The matrices analyzed consisted of 73 metabolites detected in the urinary extract and 43 in the blood sera.
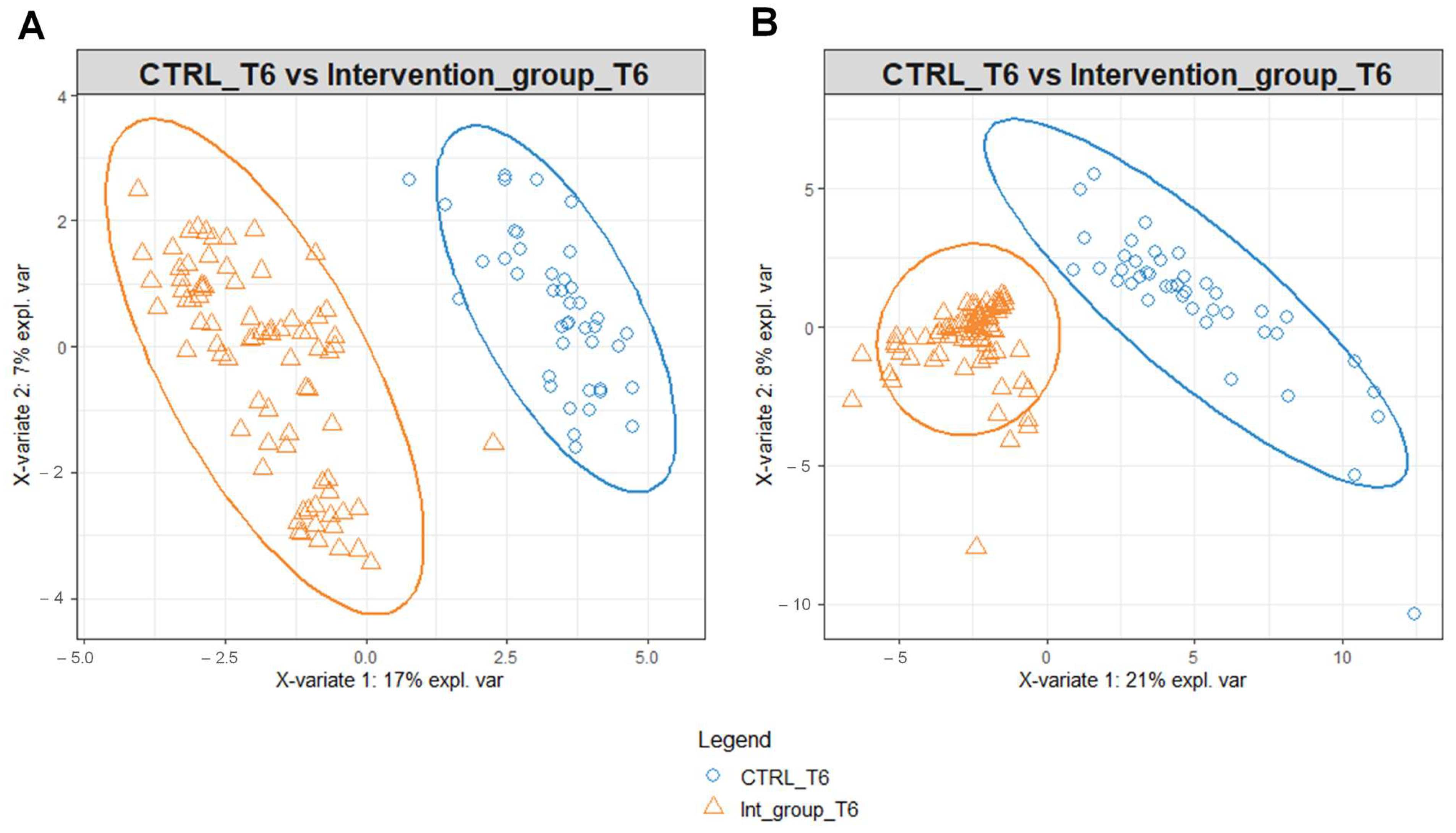
To identify possible modifications in the metabolomic profiles in response to the different water intakes for the intervention and the control group, we performed PLS-DA on the data matrices using serum and urine metabolites detected at T6 as variables (Figure 3). PLS-DA score plot identified a separation of intervention and control group serum (Figure 3A) and urine (Figure 3B) metabolomic profile at T6.
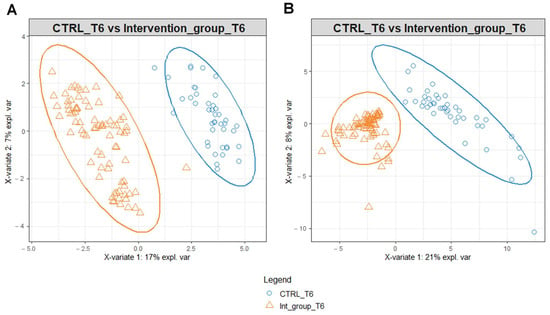
Figure 3.
PLS-DA score plot for 1H-NMR collected in 1D-NOESY spectra related to the serum (A) and urine (B) of the intervention and control group at T6.
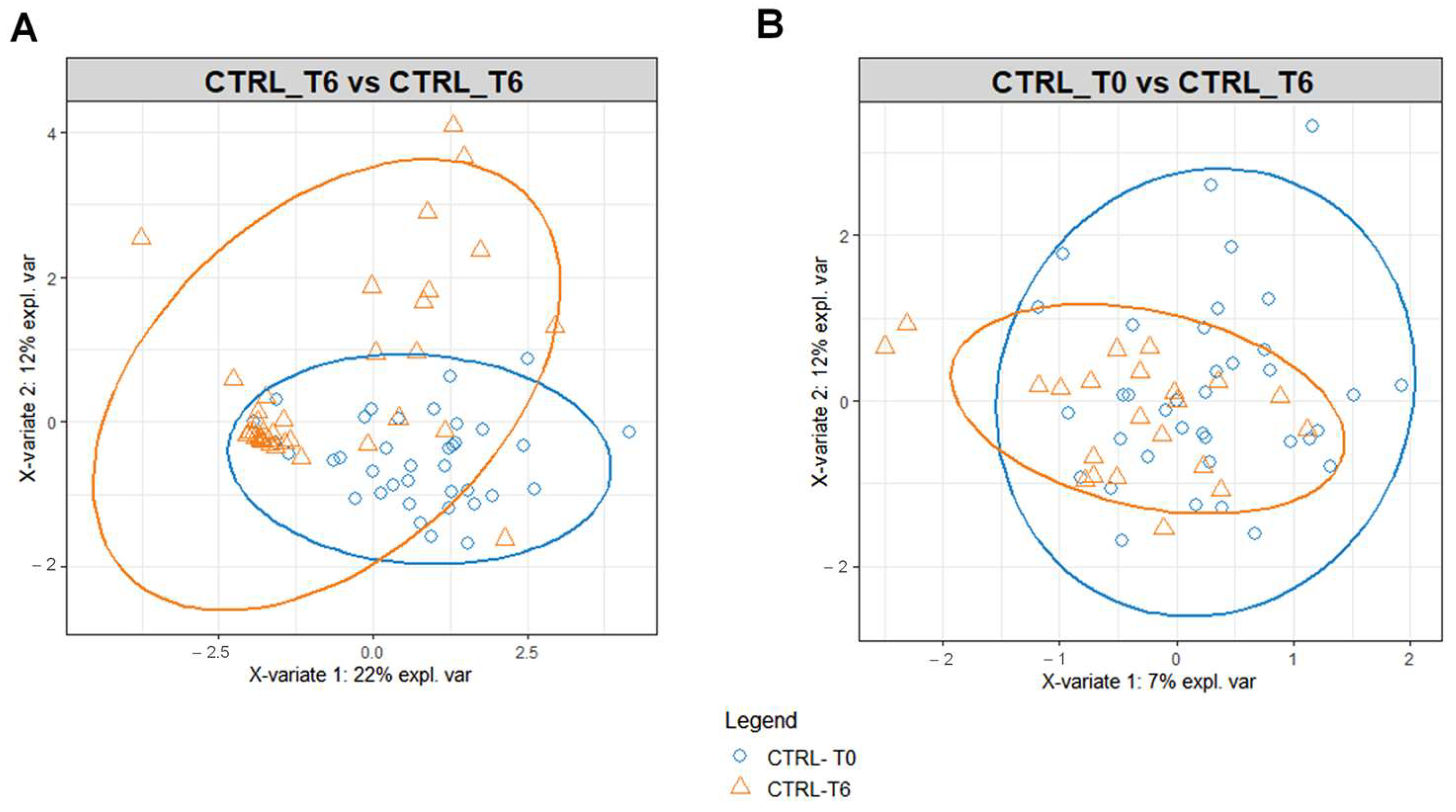
The benefit of Ca–bicarbonate water compared to CO2-supplemented oligomineral water is confirmed by the absence of clustering in the control group’s metabolomic profiles (Figure 4), in contrast to what was observed in the intervention group (Figure 5A,B).
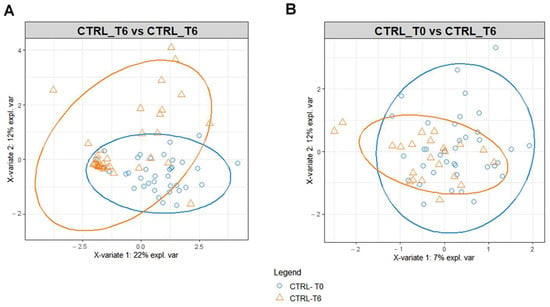
Figure 4.
PLS-DA score plot for 1H-NMR collected in 1D-NOESY spectra related to the serum (A) and urine (B) of the control at T0 (blue circles) and control group at T6 (orange triangles).
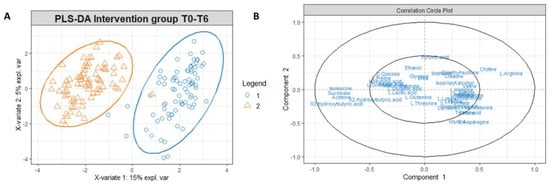
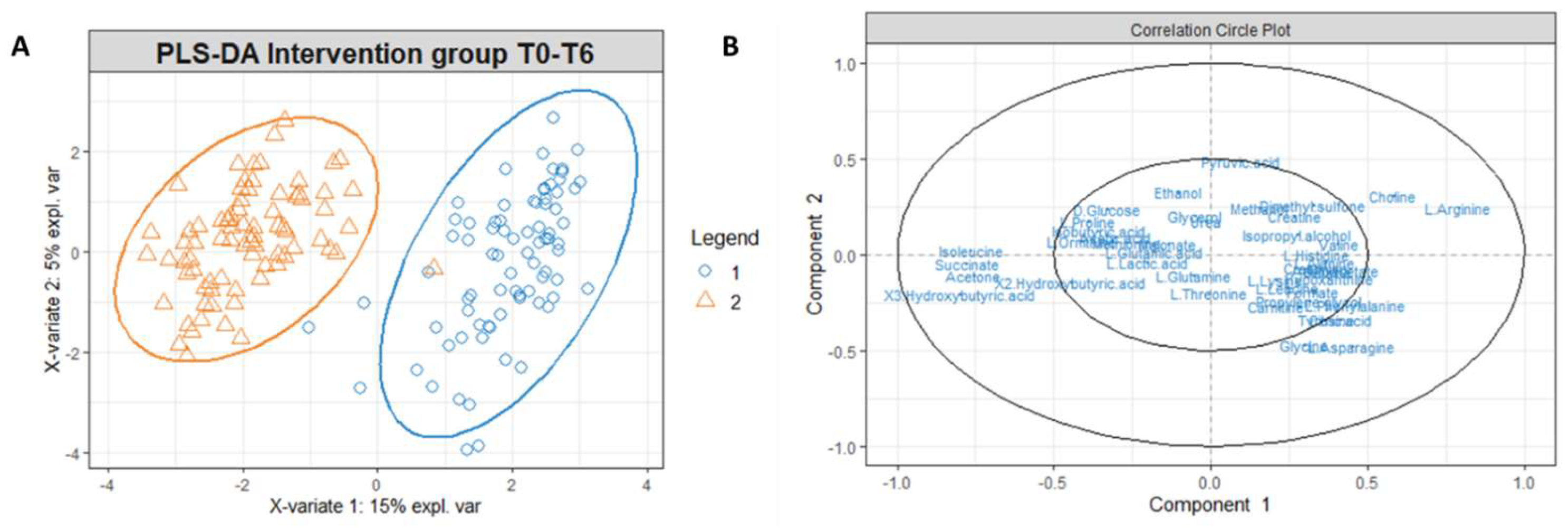
Figure 5.
(A) PLS-DA score plot for 1H-NMR collected in 1D-NOESY spectra related to the serum of the intervention group at T0 (blue circles) and intervention group at T6 (orange triangles). (B) Correlation circle plot applied to Component 1 and Component 2. Metabolites that have a greater distance from the origin of the axes represent the variables that most affect clustering. The proximity of the metabolites shows the correlation between the metabolites.
To investigate which metabolites are primarily responsible for the differences in the intervention group’s metabolomic profiles at T0 and T6, we used t-tests with a p-value threshold < 0.05, and fold change with a threshold equal to or superior to 75% (Supplementary Tables S6 and S7). In Figure 5B, the correlation circle plot applied to Components 1 and 2 indicates the variables that most affect the clustering (in the graph, the metabolites distant from the axes’ origin) and the correlation between the metabolites (evident in the proximity between the metabolites). Accordingly, an inspection of Figure 6B indicates that the metabolites significantly responsible for clusterization are isoleucine, succinate, and ketone bodies; on the other hand, a correlation exists between ketone bodies (3-hydroxybutyrate, 2-hydroxybutyrate and acetone) and succinate, as well as between isoleucine arginine and choline, as shown from the proximity of position of these metabolites on the plot.
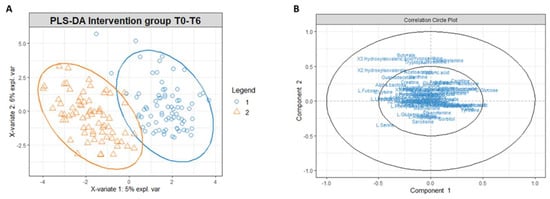
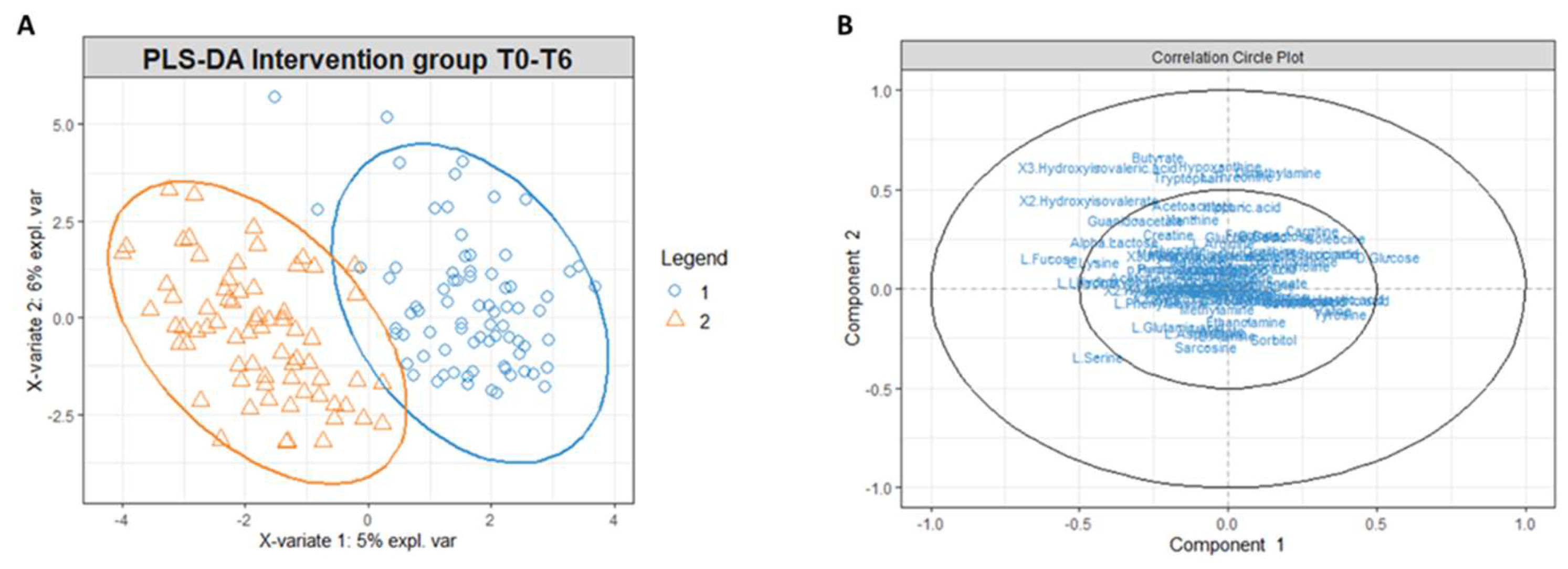
Figure 6.
(A) PLS-DA score plot for 1H NMR collected in 1D 1H NOESY spectra related to the urine of the intervention group at t0 (blue circles) and intervention group at t6 (orange triangles). (B) Correlation circle plot applied to Component 1 and Component 2. Metabolites that have a greater distance from the origin of the axes represent the variables that most affect clustering. The proximity of the metabolites shows the correlation between the metabolites.
Similarly to the serum extract, PLS-DA conducted on the urinary metabolites’ concentrations matrix revealed a different metabolomic profile of the intervention group at T0 and T6 (Figure 6A). The correlation circle plot highlighted that fatty acids such as 2-hydroxy isovalerate and 3-hydroxy isovalerate are responsible for the metabolomic modification between T0 and T6, and the changes of these metabolites correlate with changes in the concentrations of butyrate, tryptophan, threonine, dimethylamine, and hypoxanthine (Figure 6B).
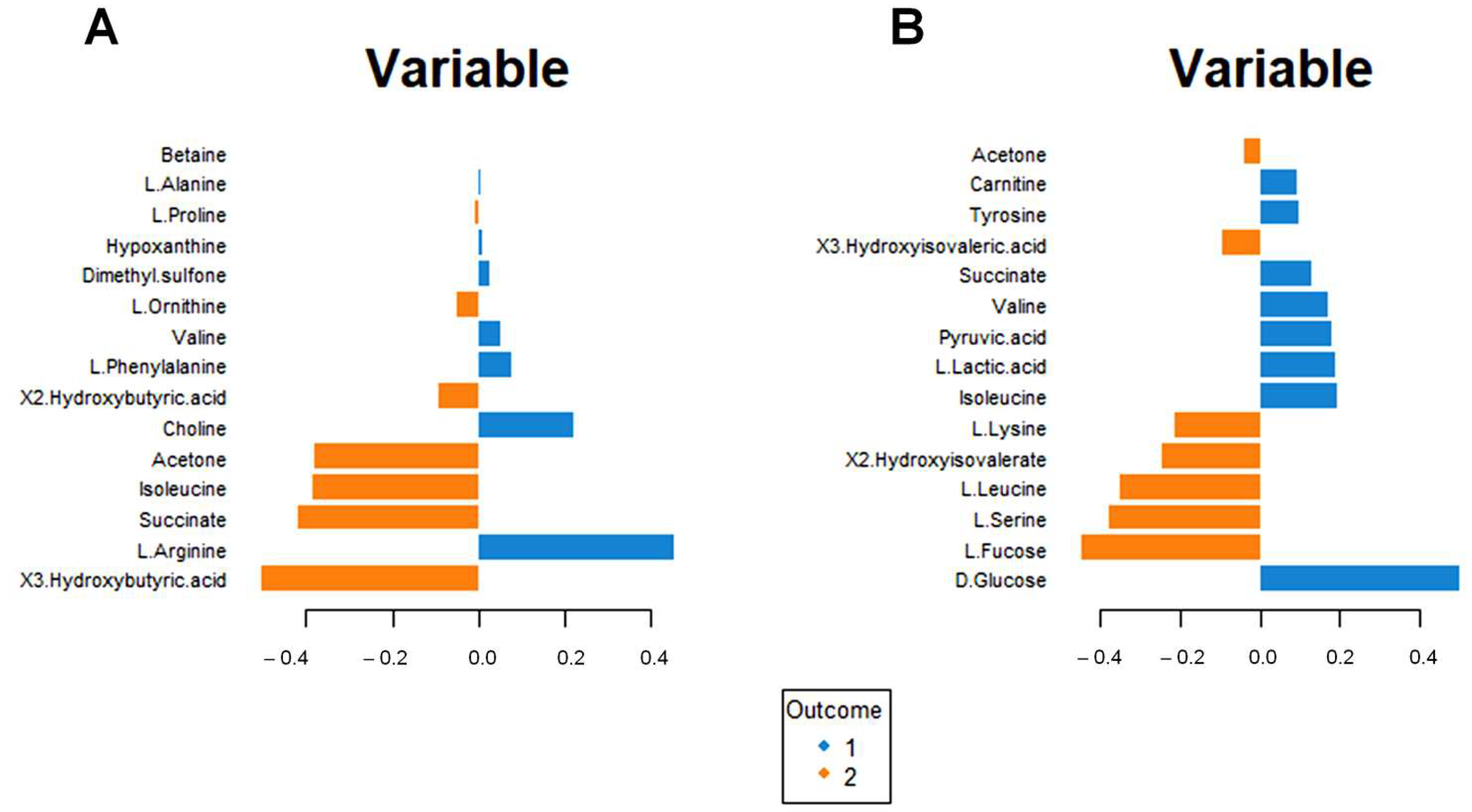
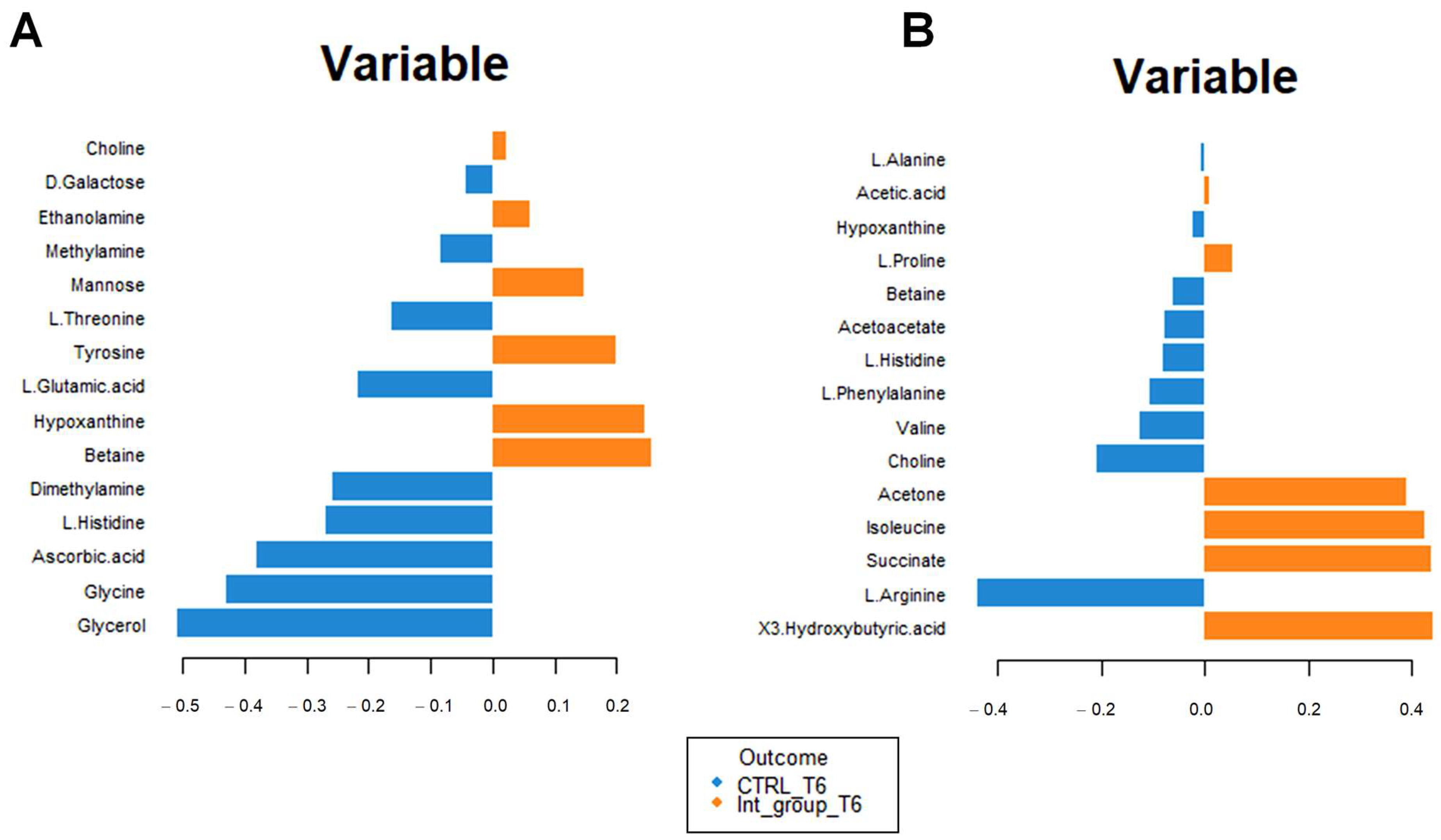
Figure 7A shows the incidence of each metabolite for separating the blood sera metabolomic profiles of the intervention group at T0 and T6. Higher concentrations of ketone bodies (3-hydroxybutyrate, 2-hydroxybutyrate, and acetone) are evident for the intervention group at T6. Moreover, significant modifications in the amino acid concentrations result in increased Isoleucine and Ornithine and lower Arginine, Valine, and Phenylalanine concentrations.
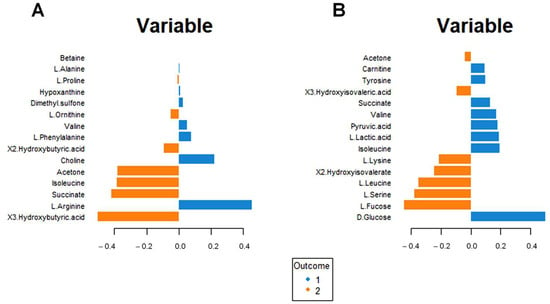
Figure 7.
Loading plot related to the matrix of serum (A) and urinary (B) metabolites. The variables responsible for metabolomic profile differences are ordered according to values of increasing importance from bottom to top. Colors indicate the cluster where the median is maximum for each metabolite (blue: Intervention group T0; orange: Intervention group T6). The metabolite corresponding to an orange band has high concentrations in the intervention group at T6, while the metabolite corresponding to a blue band has higher concentrations in the intervention group at T0.
A similar analysis for the urine metabolomic profile (Figure 7B) indicates a reduction in glucose, lactic acid, and pyruvate concentrations (suggestive of altered energy metabolism) and an increase in fucose, 3-hydroxyisovalerate, and 2-hydroxyisovalerate (Figure 7B); in addition, lower valine and tyrosine concentration, as well as higher lysine, leucine, and serine concentrations are found.
Interestingly, increased urinary succinate and isoleucine concentrations are associated with decreased serum succinate and isoleucine concentrations.
In order to understand which variables were mostly affected by Ca–bicarbonate water intake, we carried out the loading plot analysis using the serum and urinary matrices of the control and the intervention group at T6.
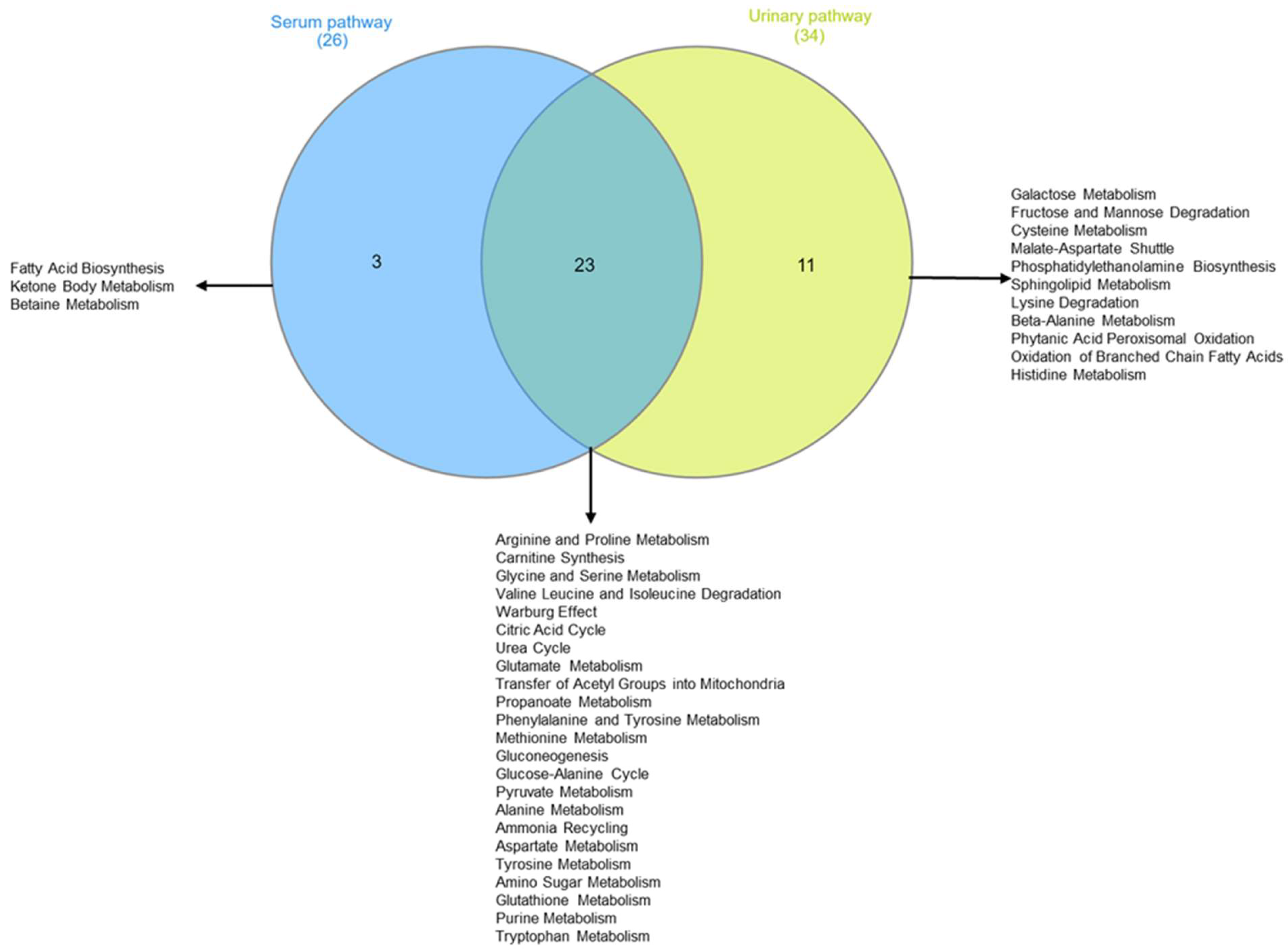
Figure 8A confirms the high concentrations of 3-hydroxybutyrate, succinate, and acetone in the intervention compared to the control group at T6. The impact of bicarbonate–Ca water intake on amino acid metabolism is confirmed by increased serum concentrations of isoleucine and proline and decreased urinary excretion of glycine, histidine, and glutamate in the intervention group at T6 (Figure 8B). To interpret the change in metabolite concentrations in terms of the modifications of biochemical pathways, we carried out pathway analysis using MetaboAnalyst 5.0 (Table 3). Furthermore, we identified the common dysregulated pathways between the two biofluids using the Venn diagram (Figure 9) [42].
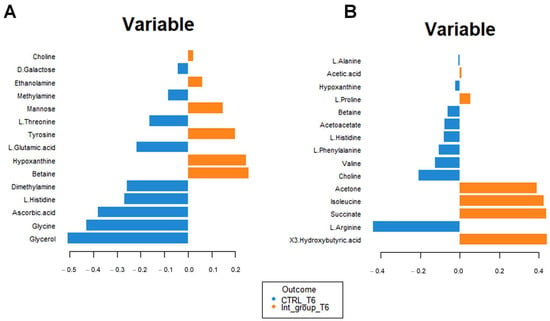
Figure 8.
Loading plot related to the matrix of serum (A) and urinary (B) metabolites. The variables are ordered according to values of increasing importance from bottom to top. Colors indicate the cluster where the median is maximum for each metabolite (blue: control group T6; orange: intervention group T6). The metabolite corresponding to an orange band has high concentrations in the intervention group at T6, while the metabolite corresponding to a blue band has higher.
Table 3.
Metabolic pathways corresponding to serum and urinary metabolomic profile of intervention group. The pathways are classified according to p-values, Holm adjustment, and FDR values. The pathways are selected based on Hits > 2, Raw p < 0.05, and Holm adjust, FDR < 1.
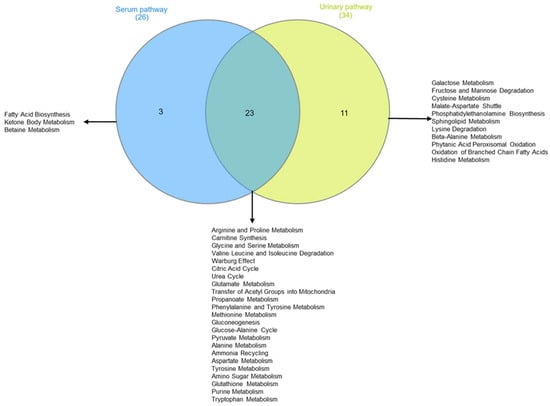
Figure 9.
Two-sets Venn diagram intersecting metabolomic pathway identified by pathway enrichment analysis of intervention group serum and urine.
Table 3 shows that, according to the change in metabolite concentrations, biochemical pathways related to fatty acids’ biosynthesis and ketone body metabolism are modified (pathways’ selection criteria are hits > 2 and p-value < 0.05). Moreover, those biochemical pathways related to amino acid metabolism appear significantly altered: arginine and proline; glycine and serine; glutamate, alanine, and methionine; valine, leucine, and isoleucine degradation. Finally, metabolite-altered concentrations in urine are consistent with the alteration in biochemical pathways related to galactose metabolism, fructose, and mannose degradation.
4. Discussion
Osteoporosis is a chronic disease characterized by a reduction in bone density. It is prevalent in postmenopausal women [43], causing a significant risk of bone fractures [44]. The primary intervention to control osteoporosis is a healthy lifestyle based on correct nutrition and regular physical activity. A diet with adequate Ca intake is a fundamental recommendation [45].
Milk and its derivatives are considered a primary Ca source [23,24,25,26]. However, their consumption has progressively been reduced as they are regarded as hypercaloric food. To favor the consumption of Ca in the diet, the leading health authority, WHO, has recently recommended diet supplementation with Ca-rich water. Indeed, in vitro and in vivo experiments have proved the efficacy of alkaline Ca-rich water to favor bone matrix affixing [13]. At the same time, several clinical studies have demonstrated that bicarbonate–calcic water is an efficacious nutritional supplement to preserve an adequate Ca content in pathological conditions characterized by bone matrix loss [46,47]. While all these data demonstrate the efficacy of bicarbonate–calcic water on Ca metabolism and bone status [6,13,48], data regarding the impact of bicarbonate–calcium water supplements on the systemic metabolic balance are missing.
Here, we reported an analysis of the blood and urine of 121 perimenopausal women using standard laboratory tests (Supplementary Tables S2–S5, Figure 1 and Figure 2) and NMR-based metabolomics analysis. The women were divided into an intervention group advised to drink 2l of bicarbonate–calcic water daily for 6 months, and a control group advised to drink 2l of CO2-supplemented oligomineral water daily.
Ca metabolism and bone health status may be appreciated by measuring blood Ca concentration and the markers responsible for its regulation: PTH, 25(OH)vitamin D, and calcitonin, as well as several ions and enzymes whose concentrations are linked to Ca, such as phosphate, magnesium, total proteins, and bone alkaline phosphatase [13].
Accordingly, all these parameters were monitored in the blood and by 24 h urine collection. After six months, the subjects drinking bicarbonate–calcium water showed increased blood and urine Ca concentration values, indicating a correct Ca absorption in response to the Ca-rich water treatment. In line with this metabolic change, increases in PTH, 1,25(OH)2D, and urinary excretion of P and Mg were evident; moreover, there was a decrease in OSTEO and urinary-specific weight. Interestingly, the latter are products of bone catabolism, suggesting that their decrease is a limitation of bone catabolism. In response to the Lete® water treatment, the intervention group also exhibited an increase in urinary Na, typical of the diuretic water effect. Remarkably, this is specific, as it was not observed at T6 in the control group, although subjected to the 2l water treatment.
Clinical laboratory tests indicate that several parameters not strictly related to the Ca metabolism changed in the intervention group at T6 (Figure 2). For example, a decrease in glycemia related to energetic metabolism is evident, hinting at a more general effect on metabolism associated with the improved metabolism of Ca.
To explore this further, we performed an NMR-based metabolomic analysis of the urine and sera of the women enrolled in this study at T0 and T6. While the metabolomic profile of the women in the intervention and the control group at T0 is very similar, separation is observable at T6 (Figure 3A,B). This is due to the different effect of the Lete® diet–therapy treatment, which induces significant metabolomic changes in the control group at T6, while the same is not observable for CO2-enriched mineral water (Figure 4A,B).
In search of the metabolites responsible for the modification of the metabolome in the intervention group, we performed loading plot analysis. Accordingly, we observed a statistically significant decrease in glucose (Figure 7B, Supplementary Table S7) after the intake of Lete. Moreover, the hypoglycemic effect is associated with modification of the succinate concentrations (increased in the serum and decreased in the urine at T6) (Figure 7A,B). By looking at the biochemical pathway to comprehend the significance of these metabolic changes, we observed that the hypoglycemic effect associated with the modification of the succinate concentrations may reflect an enhancement of the mitochondrial activity (Table 3, Figure 9). A significant amount of scientific data shows that altered mitochondrial activity induces systemic metabolic dysfunction and inflammation.
These data show that bicarbonate–calcium water treatment, in addition to the effects on Ca and bone metabolism, may improve systemic inflammatory conditions. Interestingly, severe osteoporosis, with a high risk of bone fracture, is frequently associated with several symptoms of metabolic syndrome and hyperglycemia, suggesting that a general condition of energy dysmetabolism and systemic inflammation also worsens this pathology [49,50].
The pathway analysis confirms these data, indicating that the changes in the metabolite concentrations are associated with the modulation of metabolic biochemical networks related to the improvements in the mitochondrial activity’s (i) citric acid cycle, (ii) gluconeogenesis, (iii) acyl group transfer, and (iv) pyruvate metabolism (Table 3, Figure 9).
The metabolomic profile of women in the intervention group is characterized by increased concentrations of several metabolites associated with the stimulation of tissue repair, bone fixing, and collagen synthesis. Specifically, we observed high concentrations of ornithine, an amino acid involved in collagen synthesis (Figure 8A) [51]. Stimulation of collagen synthesis was also confirmed by the upregulation of arginine and proline metabolism, evident from the blood and urine pathway analysis (Figure 9 Table 3). Then, we observed high concentrations of succinate, 3-hydroxybutyrate, and acetone, metabolites involved in the in vitro differentiation of murine osteoblast and in vivo bone mineralization (Figure 8A,B) [52,53]. In addition, previous scientific studies have shown that the variation in the ketone bodies, in particular of 3-hydroxybutyrate and the energy pathways shown in Table 3, could predict increased osteoblast activity against osteoclast remodeling [50,53,54]
According to sera and urine metabolomic data, mineral water treatment induces modifications in the biochemical pathways related to the metabolisms of the branched-chain amino acids valine, leucine, and isoleucine, and aromatic amino acid phenylalanine and arginine (Table 3). This confirms previously published data according to which high valine, arginine, and phenylalanine concentrations may be osteoporosis biomarkers [55].
Taken together, our data show that bicarbonate–calcic water intake for six months, in addition to inducing an improvement in the biochemical parameters related to Ca metabolism and bone health, can (i) restore mitochondrial energy pathways, (ii) promote the application of bone mass and collagen, and (iii) reduce the biomarkers of osteoporosis. Therefore, the alkaline environment improves bone metabolism toward a lower risk of osteoporosis and bone fractures.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/metabo13111109/s1, Supplementary Table S1: Eating habits at t0. Number of patients taking at least once a week a portion of the tabulated food. Supplementary Table S2: Intervention group and control group’s clinical average ± standard deviation parameters at t0. p-value was calculated using t-test. Supplementary Table S3: Intervention group’s clinical average ± standard deviation parameters at t0 and t6. p-value was calculated using t-test. Supplementary Table S4: Control group’s clinical average ± standard deviation parameters at t0 and t6. p-value was calculated using t-test. Supplementary Table S5: Intervention group and control group’s clinical average ± standard deviation parameters at t6. p-value was calculated using t-test. Supplementary Table S6: Important serum and urine metabolites identified by Fold Change and logarithmic Fold Change (log2(FC)) parameters calculated. Supplementary Table S7: Important serum and urinary metabolites identified by t-tests values, p-values (threshold < 0.05), logarithmic p-values, and false discovery rate (FDR) parameters calculated for the most statistically significative compounds. Supplementary Figure S1: Sample prediction area plot carried out using Centroid (a), Maximum distance (b), and Mahalanobis (c) showing the distribution of serum polar extract of intervention group at T0 (1) vs. intervention group at T6 (2) in validation area. Supplementary Figure S2: Sample prediction area plot carried out using Centroid (a), Maximum distance (b), and Mahalanobis (c) showing the distribution of urine polar extract intervention group at T0 (1) vs. intervention group at T6 (2) in validation area. Supplementary Figure S3: Sample prediction area plot carried out using Centroid (a), Maximum distance (b), and Mahalanobis (c) showing the distribution of serum polar extract intervention group at T6 vs. CTRL group at T6 in validation area. Supplementary Figure S4: Sample prediction area plot carried out using Centroid (a), Maximum distance (b), and Mahalanobis (c) showing the distribution of urine polar extract intervention group at T6 vs. CTRL group at T6 in validation area.
Author Contributions
Conceptualization, L.R., A.M.D. and G.C.; methodology, I.P., M.D., A.C., P.M. and C.P.; software, C.M.; validation, C.M., M.G. and I.P.; formal analysis, C.M.; investigation, I.P.; resources, L.R.; data curation, I.P.; writing—review and editing, A.S., M.B., A.M.D. and C.M.; visualization, C.M.; supervision, A.M.D.; project administration, L.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The institutional Ethics Committee CAMPANIA NORD for clinical trials and bio-medical research located at Azienda Ospedaliera “San Giuseppe Moscati,” Avellino, Italy, approved the study protocol (Prot. N.: NKCL2101; reg. N. CECN/1522), which followed the Declaration of Helsinki, according to the International Guidelines of Good Clinical Prac-tice and the regulations of clinical trials. Informed written consent was obtained from participants after providing information about the nature, purpose, and procedures of the study (ClinicalTrials.gov Identifier: NCT05854342, University of Salerno Protocol Record CECN/1522).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data is available within the article.
Acknowledgments
We thanks Amos Paternopoli, Amos Montoro, and Amdos Forino for helping to recruit patients for the study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Weaver, C.M. The role of nutrition on optimizing peak bone mass. Asia Pac. J. Clin. Nutr. 2008, 17 (Suppl. 1), 135–137. [Google Scholar] [PubMed]
- Flynn, A. The role of dietary calcium in bone health. Proc. Nutr. Soc. 2003, 62, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Cormick, G.; Belizán, J.M. Calcium Intake and Health. Nutrients 2019, 11, 1606. [Google Scholar] [CrossRef] [PubMed]
- Winkler, A.; Knoche, M. Calcium and the physiology of sweet cherries: A review. Sci. Hortic. 2019, 245, 107–115. [Google Scholar] [CrossRef]
- Power, M.L.; Heaney, R.P.; Kalkwarf, H.J.; Pitkin, R.M.; Repke, J.T.; Tsang, R.C.; Schulkin, J. The role of calcium in health and disease. Am. J. Obstet. Gynecol. 1999, 181, 1560–1569. [Google Scholar] [CrossRef] [PubMed]
- Heaney, R.P.; Dowell, M.S. Absorbability of the calcium in a high-calcium mineral water. Osteoporos. Int. 1994, 4, 323–324. [Google Scholar] [CrossRef] [PubMed]
- DeGrazia, J.A.; Ivanovich, P.; Fellows, H.; Rich, C. A double isotope method for measurement of intestinal absorption of calcium in man. J. Lab. Clin. Med. 1965, 66, 822–829. [Google Scholar]
- Thor, K. Calcium—Nutrient and messenger. Front. Plant Sci. 2019, 10, 440. [Google Scholar] [CrossRef]
- Mente, A. High Urinary Calcium Excretion and Familial Aggregation of Hypertension, Kidney Stone Disease, Obesity, Excessive Weight Gain and Type 2 Diabetes in Patients with Calcareous Stones; University of Toronto: Toronto, ON, Canada, 2007. [Google Scholar]
- Shadman, A.; Bastani, B. Kidney calculi: Pathophysiology and as a systemic disorder. Iran. J. Kidney Dis. 2017, 11, 180. [Google Scholar]
- Fitzpatrick, L.A. Secondary causes of osteoporosis. Mayo Clin. Proc. 2002, 77, 453–468. [Google Scholar] [CrossRef]
- Heaney, R.P. Pathophysiology of osteoporosis. Endocrinol. Metab. Clin. N. Am. 1998, 27, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Bacciottini, L.; Tanini, A.; Falchetti, A.; Masi, L.; Franceschelli, F.; Pampaloni, B.; Giorgi, G.; Brandi, M.L. Calcium bioavailability from a calcium-rich mineral water, with some observations on method. J. Clin. Gastroenterol. 2004, 38, 761–766. [Google Scholar] [CrossRef] [PubMed]
- Carpintero, P.; Gil-Garay, E.; Hernández-Vaquero, D.; Ferrer, H.; Munuera, L. Interventions to improve inpatient osteoporosis management following first osteoporotic fracture: The PREVENT project. Arch. Orthop. Trauma Surg. 2009, 129, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Rylander, R.; Arnaud, M.J. Mineral water intake reduces blood pressure among subjects with low urinary magnesium and calcium levels. BMC Public Health 2004, 4, 56. [Google Scholar] [CrossRef] [PubMed]
- Vannucci, L.; Fossi, C.; Quattrini, S.; Guasti, L.; Pampaloni, B.; Gronchi, G.; Giusti, F.; Romagnoli, C.; Cianferotti, L.; Marcucci, G.J.N. Calcium intake in bone health: A focus on calcium-rich mineral waters. Nutrients 2018, 10, 1930. [Google Scholar] [CrossRef] [PubMed]
- WHO. WHO Handbook on Indoor Radon: A Public Health Perspective; World Health Organization: Geneva, Switzerland, 2009. [Google Scholar]
- Quattrini, S.; Pampaloni, B.; Brandi, M.L. Natural mineral waters: Chemical characteristics and health effects. Clin. Cases Miner. Bone Metab. 2016, 13, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Cotruvo, J.A.; Bartram, J. Calcium and Magnesium in Drinking-Water: Public Health Significance; World Health Organization: Geneva, Switzerland, 2009. [Google Scholar]
- European Union. Directive 54/EC of the European Parliament and of the Council of 18 June 2009 on the exploitation and marketing of natural mineral waters. Off. J. Eur. Union 2009, 164. [Google Scholar]
- Halpern, G.M.; Van de Water, J.; Delabroise, A.-M.; Keen, C.L.; Gershwin, M.E. Comparative uptake of calcium from milk and a calcium-rich mineral water in lactose intolerant adults: Implications for treatment of osteoporosis. Am. J. Prev. Med. 1991, 7, 379–383. [Google Scholar] [CrossRef]
- Couzy, F.; Kastenmayer, P.; Vigo, M.; Clough, J.; Munoz-Box, R.; Barclay, D.V. Calcium bioavailability from a calcium-and sulfate-rich mineral water, compared with milk, in young adult women. Am. J. Clin. Nutr. 1995, 62, 1239–1244. [Google Scholar] [CrossRef]
- Greupner, T.; Schneider, I.; Hahn, A. Calcium bioavailability from mineral waters with different mineralization in comparison to milk and a supplement. J. Am. Coll. Nutr. 2017, 36, 386–390. [Google Scholar] [CrossRef]
- Guillemant, J.; Le, H.; Guillemant, S.; Delabroise, A.; Arnaud, M.J. Acute effects induced by a calcium-rich mineral water on calcium metabolism and on parathyroid function. Osteoporos. Int. 1997, 7, 85–86. [Google Scholar] [CrossRef]
- Wynn, E.; Raetz, E.; Burckhardt, P.J. The composition of mineral waters sourced from Europe and North America in respect to bone health: Composition of mineral water optimal for bone. Br. J. Nutr. 2008, 101, 1195–1199. [Google Scholar] [CrossRef] [PubMed]
- Van Dokkum, W.; De La Gueronniere, V.; Schaafsma, G.; Bouley, C.; Luten, J.; Latge, C. Bioavailability of calcium of fresh cheeses, enteral food and mineral water. A study with stable calcium isotopes in young adult women. Br. J. Nutr. 1996, 75, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Aptel, I.; Cance-Rouzaud, A.; Grandjean, H.; Epidos Study Group. Association between calcium ingested from drinking water and femoral bone density in elderly women: Evidence from the EPIDOS cohort. J. Bone Miner. Res. 1999, 14, 829–833. [Google Scholar] [CrossRef] [PubMed]
- Guillemant, J.; Le, H.-T.; Accarie, C.; du Montcel, S.T.; Delabroise, A.-M.; Arnaud, M.J.; Guillemant, S. Mineral water as a source of dietary calcium: Acute effects on parathyroid function and bone resorption in young men. Am. J. Clin. Nutr. 2000, 71, 999–1002. [Google Scholar] [CrossRef] [PubMed]
- Massey, L.K.; Whiting, S.J. Dietary salt, urinary calcium, and bone loss. J. Bone Miner. Res. 1996, 11, 731–736. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Sun, H.; Wang, P.; Han, Y.; Wang, X. Modern analytical techniques in metabolomics analysis. Analyst 2012, 137, 293–300. [Google Scholar] [CrossRef]
- Utpott, M.; Rodrigues, E.; de Oliveira Rios, A.; Mercali, G.D.; Flôres, S.H. Metabolomics: An analytical technique for food processing evaluation. Food Chem. 2022, 366, 130685. [Google Scholar] [CrossRef]
- Battistini, N.; Caselli, D.; Bedogni, G.; Gatti, G. Food intake in university students and its impact on nutritional status. Nutr. Res. 1992, 12, 223–233. [Google Scholar] [CrossRef]
- Bernini, P.; Bertini, I.; Luchinat, C.; Nincheri, P.; Staderini, S.; Turano, P. Standard operating procedures for pre-analytical handling of blood and urine for metabolomic studies and biobanks. J. Biomol. NMR 2011, 49, 231–243. [Google Scholar] [CrossRef]
- Emwas, A.-H.; Roy, R.; McKay, R.T.; Ryan, D.; Brennan, L.; Tenori, L.; Luchinat, C.; Gao, X.; Zeri, A.C.; Gowda, G.N.; et al. Recommendations and standardization of biomarker quantification using NMR-based metabolomics with particular focus on urinary analysis. J. Proteome Res. 2016, 15, 360–373. [Google Scholar] [CrossRef] [PubMed]
- Ravanbakhsh, S.; Liu, P.; Bjordahl, T.C.; Mandal, R.; Grant, J.R.; Wilson, M.; Eisner, R.; Sinelnikov, I.; Hu, X.; Luchinat, C. Accurate, fully-automated NMR spectral profiling for metabolomics. PLoS ONE 2015, 10, e0124219. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.; Singh, U.; Pandey, C.M.; Mishra, P.; Pandey, G. Application of student’s t-test, analysis of variance, and covariance. Ann. Card. Anaesth. 2019, 22, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Rohart, F.; Gautier, B.; Singh, A.; Lê Cao, K.-A. mixOmics: An R package for ‘omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Nakaya, A. The KEGG databases at GenomeNet. Nucleic Acids Res. 2002, 30, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Heberle, H.; Meirelles, G.V.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Pang, Z.; Lu, Y.; Ewald, J.; Xia, J. OmicsNet 2.0: A web-based platform for multi-omics integration and network visual analytics. Nucleic Acids Res. 2022, 50, W527–W533. [Google Scholar] [CrossRef] [PubMed]
- Lipovetsky, S. Book Review: Multivariate Data Integration Using R: Methods and Applications with the mixOmics Package, by Kim-Ahn Lê Cao and Zoe Welham; CRC/CRC Press: Boca Raton, FL, USA; Chapman and Hall: Boca Raton, FL, USA; Taylor & Francis Group: Abingdon, UK, 2022; pp. xxi+308. ISBN 9780367460945. [Google Scholar]
- Xia, J.; Wishart, D.S. Web-based inference of biological patterns, functions and pathways from metabolomic data using MetaboAnalyst. Nat. Protoc. 2011, 6, 743–760. [Google Scholar] [CrossRef]
- Parsons, L.C. Osteoporosis: Incidence, prevention, and treatment of the silent killer. Nurs. Clin. 2005, 40, 119–133. [Google Scholar] [CrossRef]
- Rachner, T.D.; Khosla, S.; Hofbauer, L.C. Osteoporosis: Now and the future. Lancet 2011, 377, 1276–1287. [Google Scholar] [CrossRef]
- Lau, E.M.C.; Woo, J. Nutrition and osteoporosis. Curr. Opin. Rheumatol. 1998, 10, 368–372. [Google Scholar] [CrossRef] [PubMed]
- Fasihi, S.; Fazelian, S.; Farahbod, F.; Moradi, F.; Dehghan, M. Effect of Alkaline Drinking Water on Bone Density of Postmenopausal Women with Osteoporosis. J. Menopausal Med. 2021, 27, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Wynn, E.; Krieg, M.A.; Aeschlimann, J.M.; Burckhardt, P. Alkaline mineral water lowers bone resorption even in calcium sufficiency: Alkaline mineral water and bone metabolism. Bone 2009, 44, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Pagano, I.; Rastrelli, L. Calcium from a calcium-rich mineral water: Supplementation and bioavailability. Pharmacologyonline 2017, 1, 132–138. [Google Scholar]
- Da, W.; Tao, L.; Zhu, Y. The Role of Osteoclast Energy Metabolism in the Occurrence and Development of Osteoporosis. Front. Endocrinol. 2021, 12, 675385. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.C.; Guntur, A.R.; Long, F.; Rosen, C.J. Energy Metabolism of the Osteoblast: Implications for Osteoporosis. Endocr. Rev. 2017, 38, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Harada, D.; Nagamachi, S.; Aso, K.; Ikeda, K.; Takahashi, Y.; Furuse, M. Oral administration of l-ornithine increases the content of both collagen constituting amino acids and polyamines in mouse skin. Biochem. Biophys. Res. Commun. 2019, 512, 712–715. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Hu, G.; Karner, C.M. Bioenergetic Metabolism In Osteoblast Differentiation. Curr. Osteoporos. Rep. 2022, 20, 53–64. [Google Scholar] [CrossRef]
- Cao, Q.; Zhang, J.; Liu, H.; Wu, Q.; Chen, J.; Chen, G.Q. The mechanism of anti-osteoporosis effects of 3-hydroxybutyrate and derivatives under simulated microgravity. Biomaterials 2014, 35, 8273–8283. [Google Scholar] [CrossRef]
- Zhao, Y.; Zou, B.; Shi, Z.; Wu, Q.; Chen, G.-Q. The effect of 3-hydroxybutyrate on the in vitro differentiation of murine osteoblast MC3T3-E1 and in vivo bone formation in ovariectomized rats. Biomaterials 2007, 28, 3063–3073. [Google Scholar] [CrossRef]
- Qiu, C.; Yu, F.; Su, K.; Zhao, Q.; Zhang, L.; Xu, C.; Hu, W.; Wang, Z.; Zhao, L.; Tian, Q.; et al. Multi-omics Data Integration for Identifying Osteoporosis Biomarkers and Their Biological Interaction and Causal Mechanisms. iScience 2020, 23, 100847. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).