


Effects of Carbohydrase Supplementation on Growth Performance, Intestinal Digestive Enzymes and Flora, Glucose Metabolism Enzymes, and glut2 Gene Expression of Hybrid Grouper (Epinephelus fuscoguttatus♀ × E. lanceolatus♂) Fed Different CHO/L Ratio Diets

Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diets and Animals
2.2. Sample Collection and Chemical Analysis
2.3. Proximate Composition Analysis
2.4. Plasma Biochemical Indexes and Glycogen Content Assay
2.5. Digestive Enzyme and Glucose Metabolic Enzyme Activity Assay
2.6. Real-Time Quantitative RT-PCR Analysis of Gene Expression
2.7. Intestine Microbiological Analysis
2.8. Statistical Analysis
3. Results
3.1. Growth Performance, Survival and Feed Utilization
3.2. Whole-Body Composition Analysis
3.3. Plasma Biochemical Composition Measurement
3.4. Intestinal Digestive Enzyme Activities Analysis
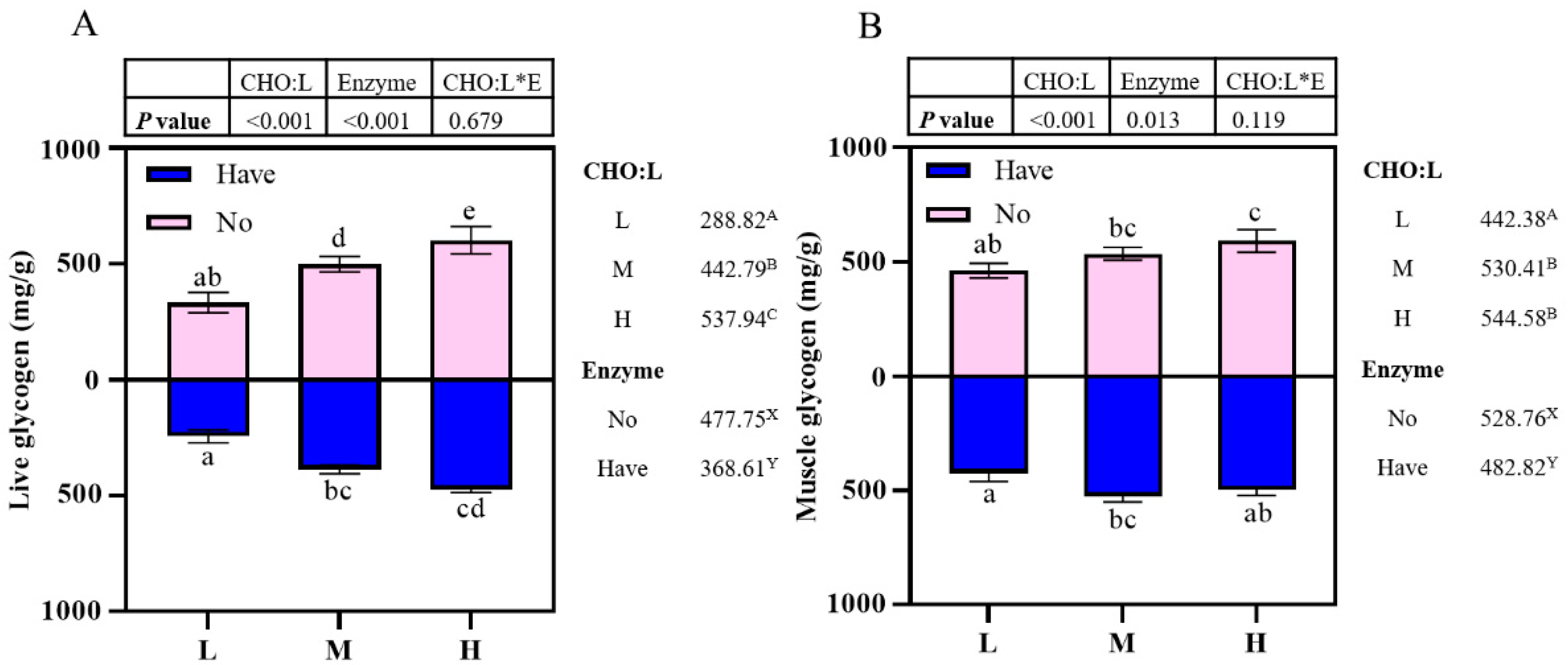
3.5. Glycogen Content Assay
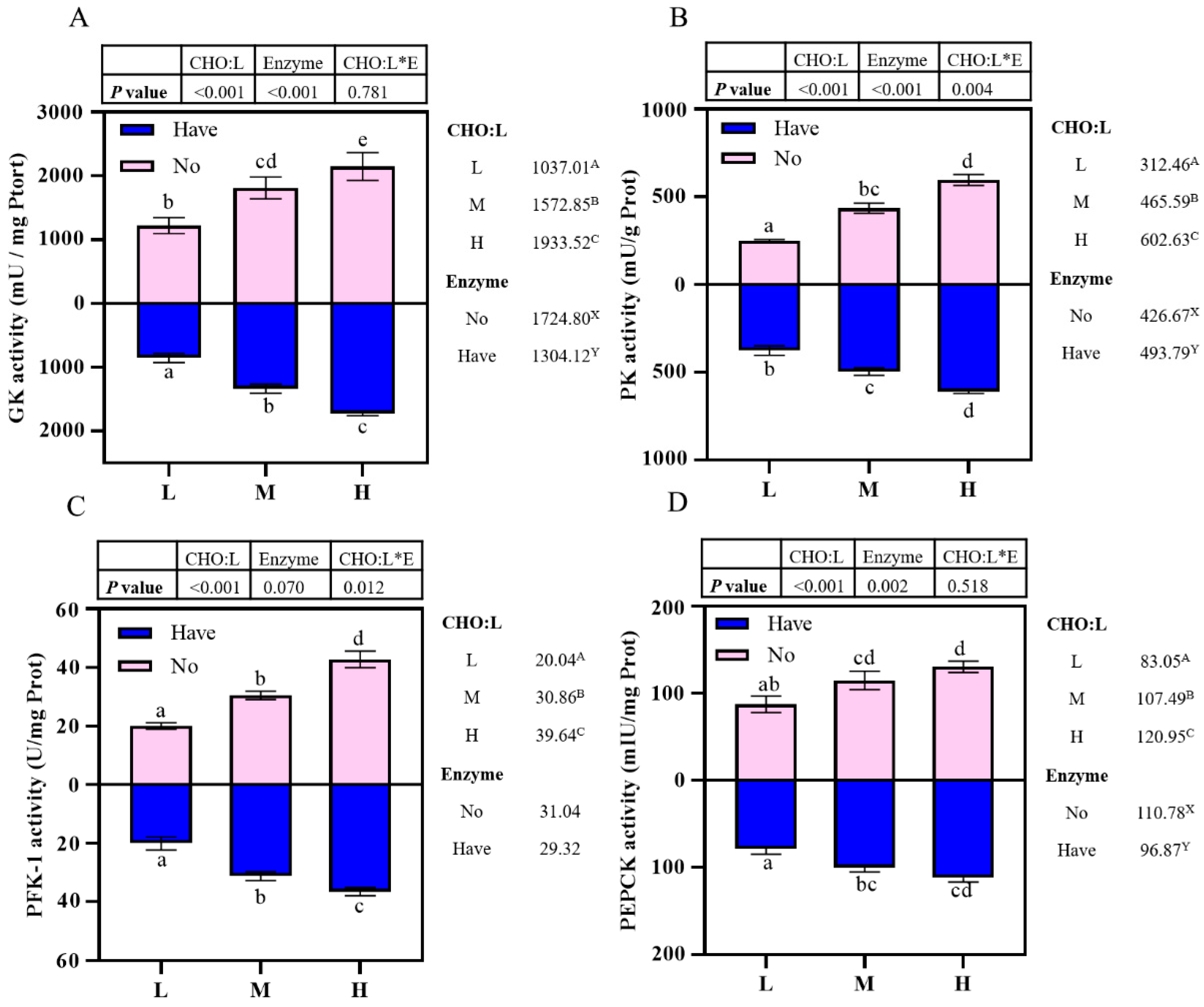
3.6. Enzyme Activity Related to Carbohydrate Metabolism in the Liver
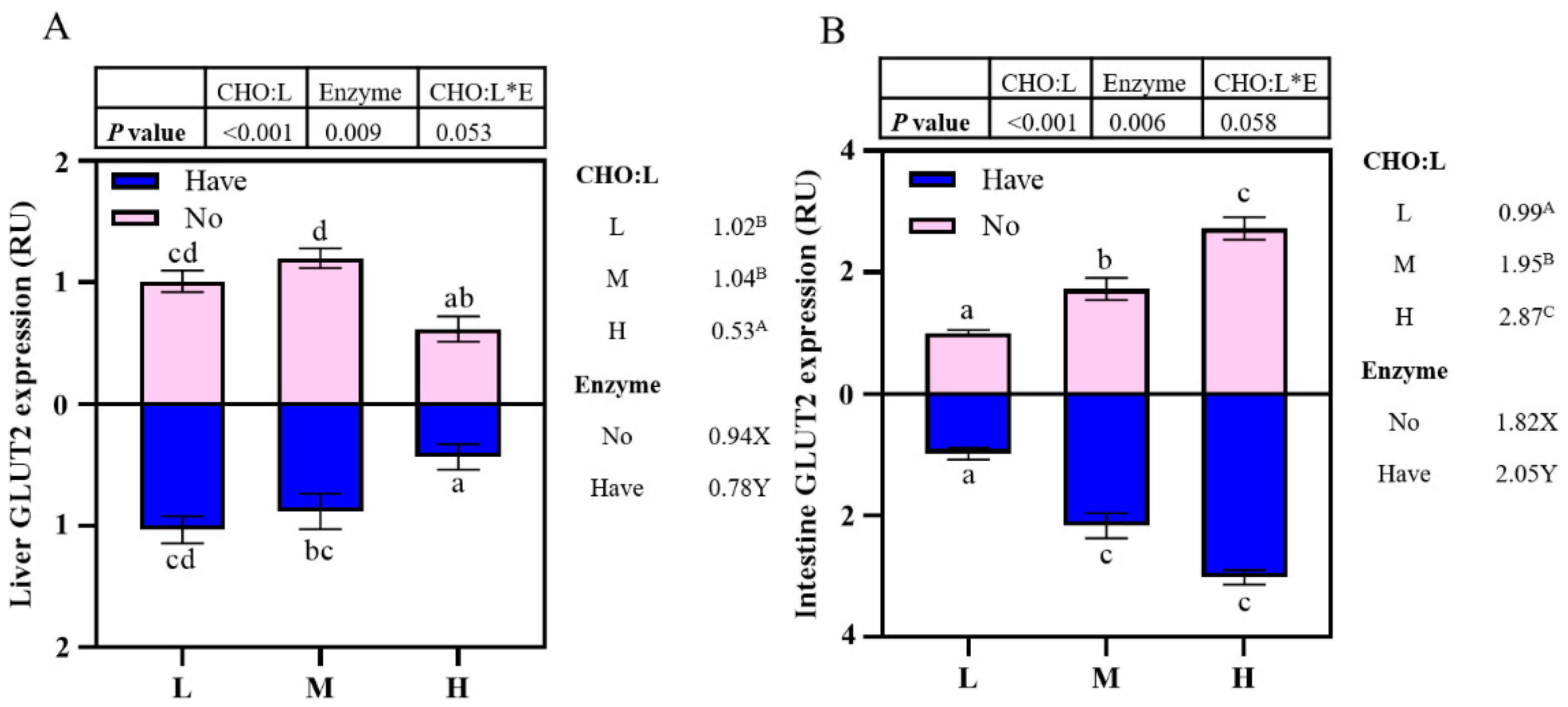
3.7. The Impact of Different Diets on glut2 mRNA Gene Expression
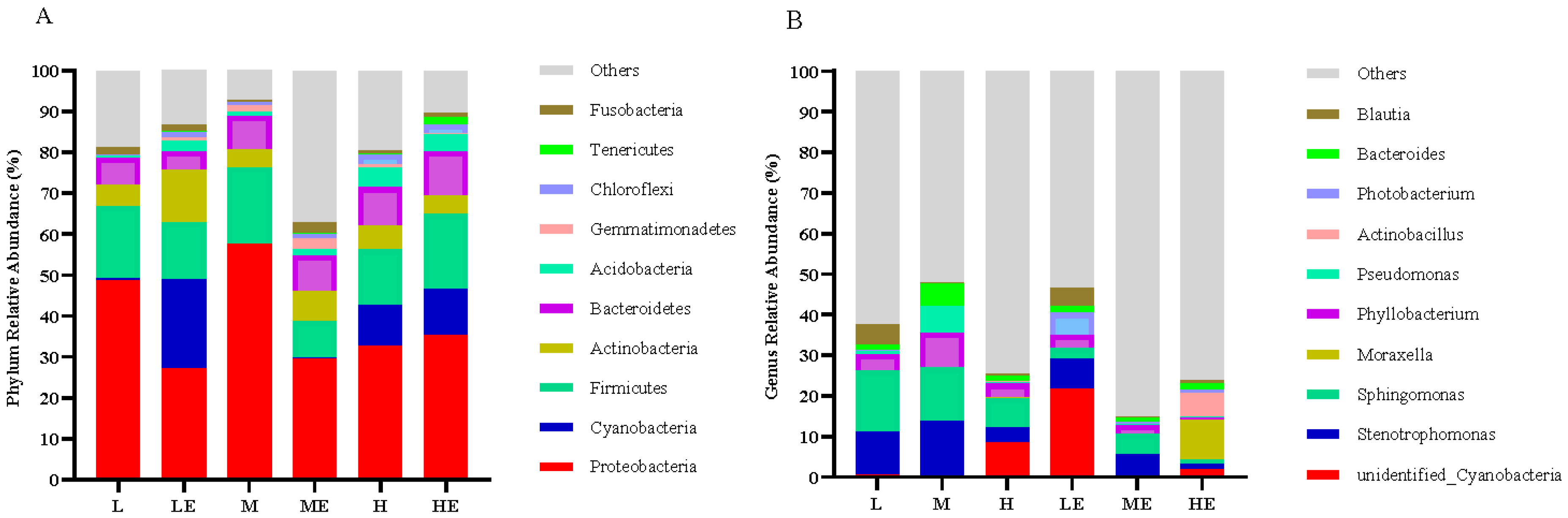
3.8. The Impact of Different Diets on Intestinal Microflora
3.8.1. Analysis at the Phylum and Genus Level
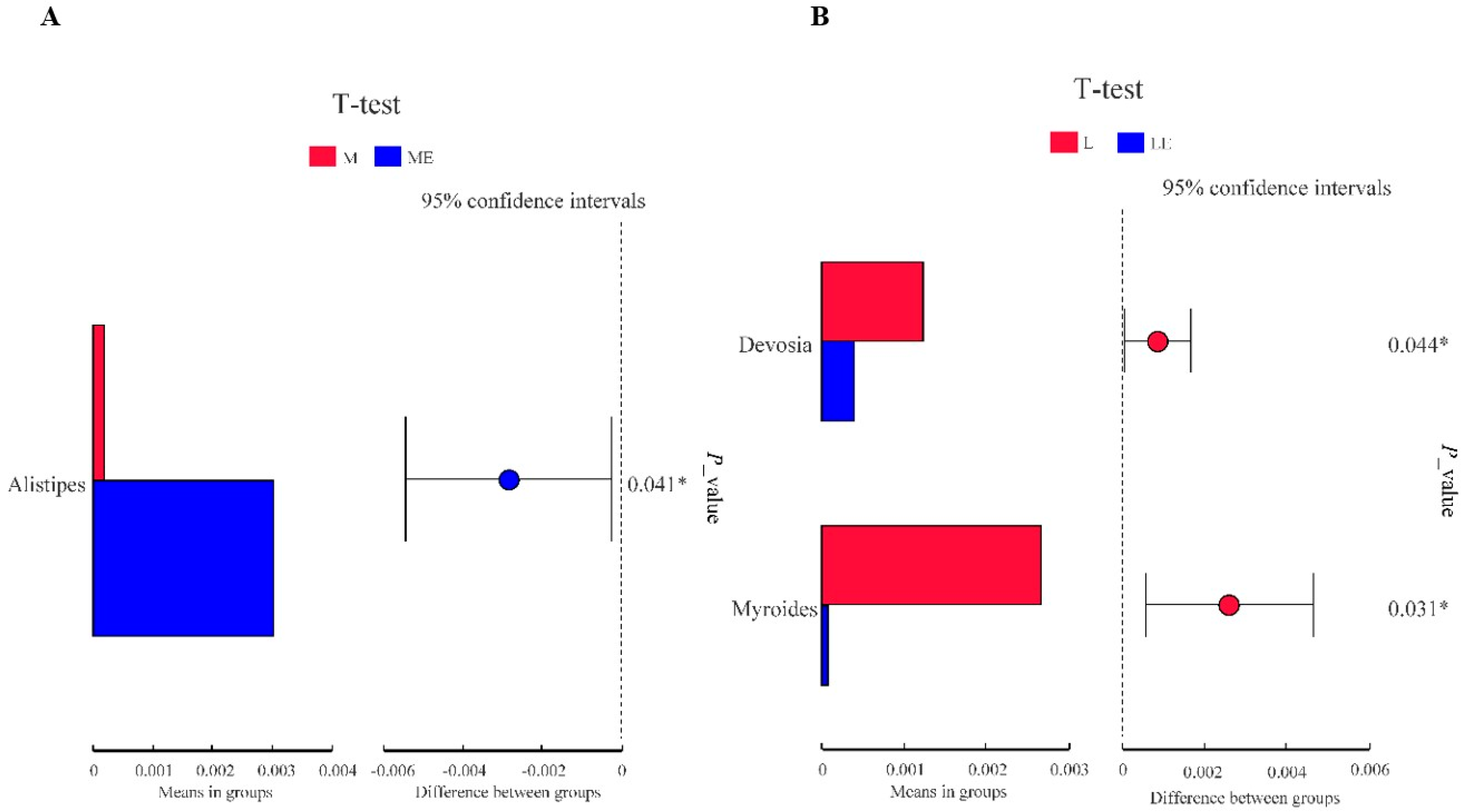
3.8.2. Multi Sample Comparative Analysis
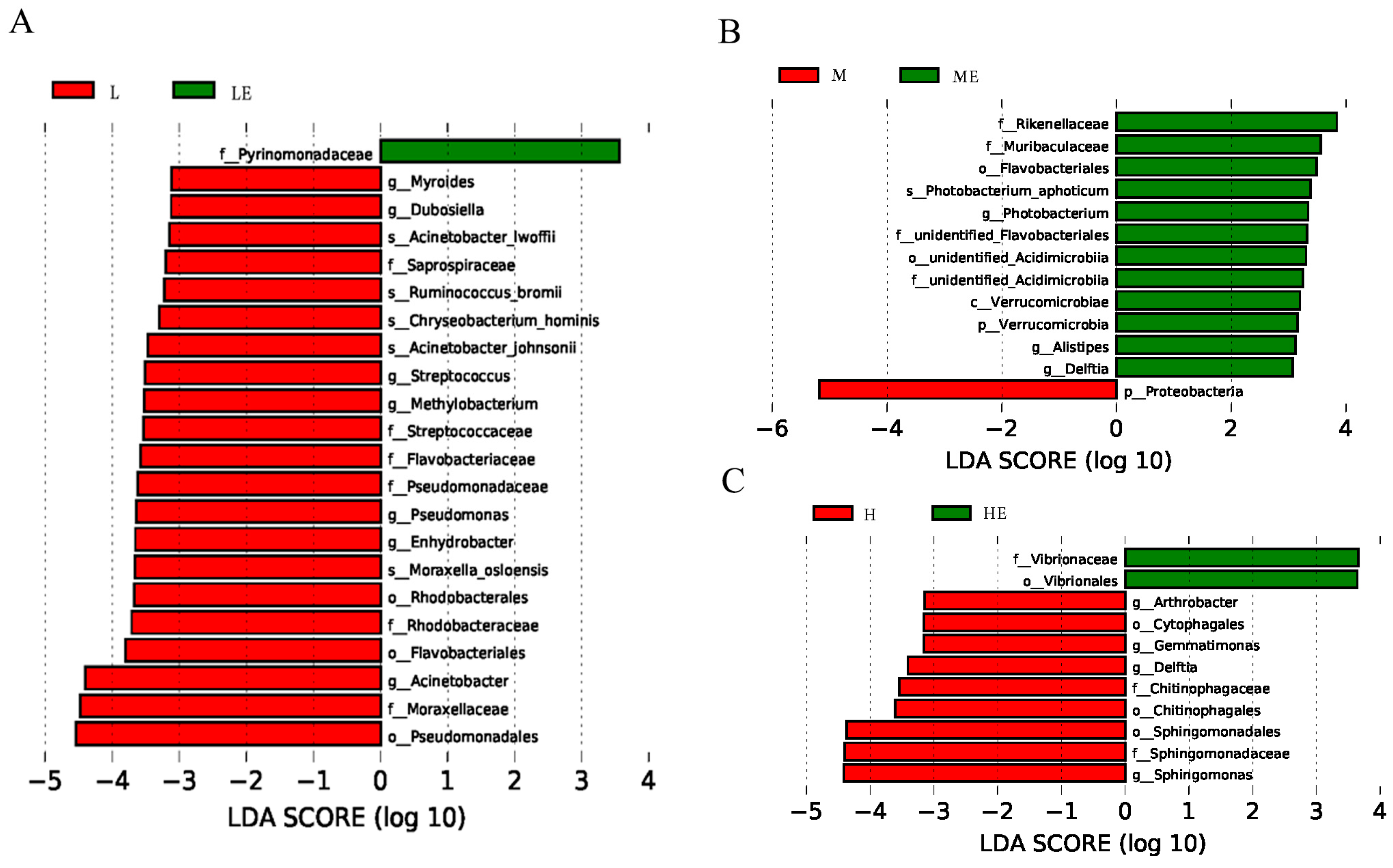
3.8.3. Linear Discriminant Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dong, L.F.; Tong, T.; Zhang, Q.; Wang, Q.C.; Xu, M.Z.; Yu, H.R.; Wang, J. Effects of dietary carbohydrate to lipid ratio on growth, feed utilization, body composition and digestive enzyme activities of golden pompano (Trachinotus ovatus). Aquac. Nutr. 2018, 24, 341–347. [Google Scholar] [CrossRef]
- Wang, L.N.; Liu, W.B.; Lu, K.L.; Xu, W.N.; Cai, D.S.; Zhang, C.N.; Qian, Y. Effects of dietary carbohydrate/lipid ratios on non-specific immune responses, oxidative status and liver histology of juvenile yellow catfish Pelteobagrus fulvidraco. Aquaculture 2014, 426, 41–48. [Google Scholar] [CrossRef]
- Council, N.R. Nutrient Requirements of Fish; National Academies Press: Washington, DC, USA, 1993. [Google Scholar]
- Dias, J.; Rueda-Jasso, R.; Panserat, S.; Conceicao, L.; Gomes, E.F.; Dinis, M.T. Effect of dietary carbohydrate-to-lipid ratios on growth, lipid deposition and metabolic hepatic enzymes in juvenile Senegalese sole (Solea senegalensis, Kaup). Aquac. Res. 2004, 35, 1122–1130. [Google Scholar] [CrossRef]
- Gao, W.; Liu, Y.J.; Tian, L.X.; Mai, K.S.; Liang, G.Y.; Yang, H.J.; Huai, M.Y.; Luo, W.J. Effect of dietary carbohydrate-to-lipid ratios on growth performance, body composition, nutrient utilization and hepatic enzymes activities of herbivorous grass carp (Ctenopharyngodon idella). Aquac. Nutr. 2009, 16, 327–333. [Google Scholar] [CrossRef]
- Zhou, Y.L.; Guo, J.L.; Tang, R.J.; Ma, H.J.; Chen, Y.J.; Lin, S.M. High dietary lipid level alters the growth, hepatic metabolism enzyme, and anti-oxidative capacity in juvenile largemouth bass Micropterus salmoides. Fish Physiol Biochem. 2020, 46, 125–134. [Google Scholar] [CrossRef]
- Ellis, S.; Reigh, R. Effects of dietary lipid and carbohy drate levels on growth and body composition of juvenile red drum, Sciaenops ocellatus. Aquaculture 1988, 97, 383–394. [Google Scholar] [CrossRef]
- Garling, D.L.; Wilson, R.P. Effects of Dietary Carbohydrate-to-Lipid Ratios on Growth and Body Composition of Fingerling Channel Catfish. Progress. Fish-Culturist. 1977, 39, 43–47. [Google Scholar] [CrossRef]
- Hjm, A.; Mmm, A.; Dcp, B.; Sml, A.; Yjc, A.; Li, L.A. Effect of dietary starch level on growth, metabolism enzyme and oxidative status of juvenile largemouth bass, Micropterus salmoides. ScienceDirect. Aquaculture 2019, 498, 482–487. [Google Scholar]
- Tan, X.Y.; Liu, Y.J.; Tian, L.X.; Yang, H.J.; Liang, G.Y.; Luo, Z.; Huang, J.W.; Guo, R. The Effects of Dietary Carbohydrate Levels on the Growth, Nutrient Composition of Juvenile Largemouth Bass Micropterus salmoides. Acta Sci. Nat. Univ. Sunyatseni. 2005, 44, 258–263. [Google Scholar]
- Kaushik, S.J.; Seiliez, I. Protein and amino acid nutrition and metabolism in fish: Current knowledge and future needs. Aquac. Res. 2010, 41, 322–332. [Google Scholar] [CrossRef]
- Buchanan, J.; Sarac, H.Z.; Poppi, D.; Cowan, R.T. Effects of enzyme addition to canola meal in prawn diets. Aquaculture 1997, 151, 29–35. [Google Scholar] [CrossRef]
- Wood, I.S.; Trayhurn, P. Glucose transporters (GLUT and SGLT): Expanded families of sugar transport proteins. Br. J. Nutr. 2003, 89, 3–9. [Google Scholar] [CrossRef]
- Hall, J.R.; Short, C.E.; Driedzic, W.R. Sequence of Atlantic cod (Gadus morhua) GLUT4, GLUT2 and GPDH: Developmental stage expression, tissue expression and relationship to starvation-induced changes in blood glucose. J. Exp. Biol. 2006, 209, 4490–4502. [Google Scholar] [CrossRef]
- Mueckler, M.; Thorens, B. The SLC2 (GLUT) family of membrane transporters. Mol. Asp. Med. 2013, 34, 121–138. [Google Scholar] [CrossRef]
- Pan, L.; Shen, J.; Liu, H.; Tan, B.; Dong, X.; Yang, Q.; Chi, S.; Zhang, S. Molecular cloning and characterization of GLUT2 from pompano (Trachinotus ovatus), and its gene expression in response to exogenous enzymes supplementation in high carbohydrate diet. Aquac. Rep. 2022, 22, 100999. [Google Scholar] [CrossRef]
- Dennis, L.P.; Ashford, G.; Thai, T.Q.; In, V.V.; Ninh, N.H.; Elizur, A. Hybrid grouper in Vietnamese aquaculture: Production approaches and profitability of a promising new crop. Aquaculture 2020, 522, 735108. [Google Scholar] [CrossRef]
- Chen, G.; Qian, J.; Liu, H.; Tan, B.; Dong, X.; Yang, Q.; Chi, S.; Zhang, S. Dietary carbohydrate-to-lipid ratios modulate juvenile hybrid grouper (♀ Epinephelus fuscoguttatus × ♂ E. lanceolatus): Effects on growth, serum biochemistry, intestinal digestion and hepatic metabolism of glucose and lipid. Aquac. Nutr. 2020, 27, 1370–1382. [Google Scholar] [CrossRef]
- Gao, Y.; Luo, Y.; Li, X.; Dong, Y.; Liao, Y.; Yao, W.; Jin, Z.; Wu, X. Effects of Dietary Carbohydrate/Lipid Ratios on Growth, Feed Utilization, Hematology Parameters, and Intestinal Digestive Enzyme Activities of Juvenile Hybrid Grouper (Brown-Marbled Grouper Epinephelus fuscoguttatus♀ × Giant Grouper E. lanceolatus♂). N. Am. J. Aquac. 2018, 80, 418–426. [Google Scholar] [CrossRef]
- Liu, H.; Yang, J.J.; Dong, X.H.; Tan, B.P.; Zhang, S.; Chi, S.Y.; Yang, Q.H.; Liu, H.Y.; Yang, Y.Z. Effects of different dietary carbohydrate-to-lipid ratios on growth, plasma biochemical indexes, digestive, and immune enzymes activities of sub-adult orange-spotted grouper Epinephelus coioides. Fish Physiol Biochem. 2020, 46, 1409–1420. [Google Scholar] [CrossRef]
- Chi, S.Y.; Wang, X.W.; Tan, B.P.; Yang, Q.H.; Dong, X.H.; Liu, H.Y.; Zhang, S. Effects of dietary methionine on the growth performance, anti-oxidation and activities of gluconeogensis-related enzyme in juvenile groupers coioides. Acta Hydrobiol. Sin. 2015, 39, 645–652. [Google Scholar]
- Li, R.X.; Liu, H.Y.; Dong, X.H.; Chi, S.Y.; Yang, Q.H.; Zhang, S.; Tan, B.P. Molecular characterization and expression analysis of glucose transporter 1 and hepatic glycolytic enzymes activities from herbivorous fish Ctenopharyngodon idellus in respond to a glucose load after the adaptation to dietary carbohydrate levels. Aquaculture 2018, 492, 290–299. [Google Scholar] [CrossRef]
- Rider, S.A.; Davies, S.J.; Jha, A.N.; Fisher, A.A.; Knight, J.; Sweetman, J.W. Supra-nutritional dietary intake of selenite and selenium yeast in normal and stressed rainbow trout (Oncorhynchus mykiss): Implications on selenium status and health responses. Aquaculture 2009, 295, 282–291. [Google Scholar] [CrossRef]
- Wu, M.; Lu, S.; Wu, X.; Jiang, S.; Luo, Y.; Yao, W.; Jin, Z. Effects of dietary amino acid patterns on growth, feed utilization and hepatic IGF-I, TOR gene expression levels of hybrid grouper (Epinephelus fuscoguttatus ♀×Epinephelus lanceolatus ♂) juveniles. Aquac.-Amst. 2017, 468, 508–514. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data using Real-Time Quantitative PCR. Methods 2002, 25, 402–408. [Google Scholar] [CrossRef]
- Coutinho, J.J.d.; Maria, N.L.; Sandre, L.C.G.; Costa, J.I.; GeraldoMartins, M.I.E.; CéliaPortella, M.; JoséCarneiro, D. Carbohydrate-to-lipid ratio in extruded diets for NILE tilapia farmed in net cages. Aquaculture 2018, 497, 520–525. [Google Scholar] [CrossRef]
- Yang, M.; Deng, K.; Pan, M.; Gu, Z.; Liu, D.; Zhang, Y.; Zhang, W.; Mai, K. Glucose and lipid metabolic adaptations during postprandial starvation of Japanese flounder Paralichthys olivaceus previously fed different levels of dietary carbohydrates. Aquac.-Amst. 2019, 501, 416–429. [Google Scholar] [CrossRef]
- Zhou, P.P.; Wang, M.Q.; Xie, F.J.; Deng, D.F.; Zhou, Q.J. Effects of dietary carbohydrate to lipid ratios on growth performance, digestive enzyme and hepatic carbohydrate metabolic enzyme activities of large yellow croaker (Larmichthys crocea). Aquaculture 2016, 452, 45–51. [Google Scholar] [CrossRef]
- Li, S.; Yin, J.; Zhang, H.; Liu, Z.; Chen, N. Effects of dietary carbohydrate and lipid levels on growth performance, feed utilization, body composition and non-specific immunity of large yellow croaker (Larimichthys crocea). Aquac. Nutr. 2019, 25, 995–1005. [Google Scholar] [CrossRef]
- Storebakken, T.; Kvien, I.S.; Shearer, K.D.; Grisdale-Helland, B.; Helland, S.J.; Berge, G.M. The apparent digestibility of diets containing fish meal, soybean meal or bacterial meal fed to Atlantic salmon (Salmo salar): Evaluation of different faecal collection methods. Aquac. Amst. 1998, 169, 195–210. [Google Scholar] [CrossRef]
- Chou, B.S.; Shiau, S.Y. Optimal dietary lipid level for growth of juvenile hybrid tilapia, Oreochromis niloticus X Oreochromis aureus. Aquaculture 1996, 143, 185–195. [Google Scholar] [CrossRef]
- Luo, Y.; Wu, X.; Li, W.; Jiang, S.; Lu, S.; Wu, M. Effects of Different Corn Starch Levels on Growth, Protein Input, and Feed Utilization of Juvenile Hybrid Grouper (male × female ). North Am. J. Aquac. 2016, 78, 168–173. [Google Scholar] [CrossRef]
- Kumar, S.; Sahu, N.P.; Pal, A.K.; Choudhury, D.; Mukherjee, S.C. Studies on digestibility and digestive enzyme activities in Labeo rohita (Hamilton) juveniles: Effect of microbial α-amylase supplementation in non-gelatinized or gelatinized corn-based diet at two protein levels. Fish Physiol. Biochem. 2006, 32, 209–220. [Google Scholar] [CrossRef]
- Liu, N.N.; Zhe, C.; Liu, G.L.; Chen, T.J.; Hong, J.; Zhong, H.; Chi, Z.M. α-Amylase, glucoamylase and isopullulanase determine molecular weight of pullulan produced by Aureobasidium melanogenum P16. Int. J. Biol. Macromol. 2018, 117, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Castillo, S.; Gatlin, D.M. Dietary supplementation of exogenous carbohydrase enzymes in fish nutrition: A review. Aquaculture 2015, 435, 286–292. [Google Scholar] [CrossRef]
- Jantrarotai, W.; Sitasit, P.; Rajchapakdee, S. The optimum carbohydrate to lipid ratio in hybrid Clarias catfish (Clarias macrocephalus× C. gariepinus) diets containing raw broken rice. Aquaculture 1994, 127, 61–68. [Google Scholar] [CrossRef]
- Cowey, C.B.; Cho, C.Y. Nutritional requirements of fish. Proc. Nutr. Soc. 1993, 52, 417–426. [Google Scholar] [CrossRef]
- López, L.M.; OlmosSoto, J.; TrejoEscamilla, I.; FloresIbarra, M.; Ochoa, L.; Drawbridge, M.; Peres, H. Evaluation of carbohydrate-to-lipid ratio in diets supplemented with Bacillus subtilis probiotic strain on growth performance, body composition and digestibility in juvenile white seabass (Atractoscion nobilis, Ayres 1860). Aquac. Res. 2016, 47, 1864–1873. [Google Scholar] [CrossRef]
- Tan, Q.; Xie, S.; Zhu, X.; Lei, W.; Yang, Y. Effect of dietary carbohydrate-to-lipid ratios on growth and feed utilization in Chinese longsnout catfish (Leiocassis longirostris Günther). J. Appl. Ichthyol. 2007, 23, 605–610. [Google Scholar] [CrossRef]
- CouncilResources, N. Nutrient Requirements of Fish and Shrimp; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Christelle, R.; Jacqueline, A.; Mireille, C.; Jean, R.; Sadasivam, K. Dietary lipid level, hepatic lipogenesis and flesh quality in turbot (Psetta maxima). Aquaculture 2001, 193, 291–309. [Google Scholar]
- Enes, P.; Peres, H.; Sanchez-Gurmaches, J.; Navarro, I.; Gutierrez, J.; Oliva-Teles, A. Insulin and IGF-I response to a glucose load in European sea bass (Dicentrarchus labrax) juveniles. Aquaculture 2011, 315, 321–326. [Google Scholar] [CrossRef]
- Olefsky, J.M. Insulin resistance and the pathogenesis of non-insulin dependent diabetes mellitus: Cellular and molecular mechanisms. Adv. Exp. Med. Biol. 1993, 334, 129–150. [Google Scholar] [PubMed]
- Czech, M.P. Molecular Actions of Insulin on Glucose Transport. Annu. Rev. Nutr. 1995, 15, 441–471. [Google Scholar] [CrossRef] [PubMed]
- Polakof, S.; Panserat, S. How Tom Moon’s research highlighted the question of glucose tolerance in carnivorous fish. Comp. Biochem. Physiol. Part B 2016, 199, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Enes, P.; Panserat, S.; Kaushik, S.; Oliva-Teles, A. Hepatic glucokinase and glucose-6-phosphatase responses to dietary glucose and starch in gilthead sea bream (Sparus aurata) juveniles reared at two temperatures. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2008, 149, 80–86. [Google Scholar] [CrossRef]
- Capilla, E.; Médale, F.; Navarro, I.; Panserat, S.; Vachot, C.; Kaushik, S.; Gutiérrez, J. Muscle insulin binding and plasma levels in relation to liver glucokinase activity, glucose metabolism and dietary carbohydrates in rainbow trout. Regul. Pept. 2003, 110, 123–132. [Google Scholar] [CrossRef]
- Caseras, A.; Metón, I.; Fernández, F.; Baanante, I.V. Glucokinase gene expression is nutritionally regulated in liver of gilthead sea bream (Sparus aurata). Biochim. Biophys. Acta 2000, 1493, 135–141. [Google Scholar] [CrossRef]
- Metón, I.; Fernández, F.; Baanante, I.V. Short- and long-term effects of refeeding on key enzyme activities in glycolysis-gluconeogenesis in the liver of gilthead seabream (Sparus aurata). Aquaculture 2003, 225, 99–107. [Google Scholar] [CrossRef]
- Enes, P.; Panserat, S.; Kaushik, S.; Oliva-Teles, A. Effect of normal and waxy maize starch on growth, food utilization and hepatic glucose metabolism in European sea bass (Dicentrarchus labrax) juveniles. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2006, 143, 89–96. [Google Scholar] [CrossRef]
- Enes, P.; Panserat, S.; Kaushik, S.; Oliva-Teles, A. Growth performance and metabolic utilization of diets with native and waxy maize starch by gilthead sea bream (Sparus aurata) juveniles. Aquac.-Amst. 2008, 274, 101–108. [Google Scholar] [CrossRef]
- Moreira, I.S.; Peres, H.; Couto, A.; Enes, P.; Oliva-Teles, A. Temperature and dietary carbohydrate level effects on performance and metabolic utilisation of diets in European sea bass (Dicentrarchus labrax) juveniles. Aquaculture 2008, 274, 153–160. [Google Scholar] [CrossRef]
- Wang, G.Y.; Liu, B.; Xie, J.; Xian, G.E. Research progress of several carbohydrate metabolic key enzymes in fish. J. Shanghai Fish. Univ. 2008, 17, 377–383. [Google Scholar]
- Baanante, I.V.; Frutos, P.D.; Bonamusa, L. Regulation of fish glycolysis—Gluconeogenesis: Role of fructose 2,6 P2 and PFK-2. Comp. Biochem. Physiol. Part B Comp. Biochem. 1991, 100, 11–17. [Google Scholar] [CrossRef]
- Metón, I.; Caseras, A.; Fernández, F.; Baanante, I.V. 6-Phosphofructo-2-kinase/fructose-2,6-bisphosphatase gene expression is regulated by diet composition and ration size in liver of gilthead sea bream, Sparus aurata. BBA-Gene Struct. Expr. 2000, 1491, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.; Xie, S.; Zhu, X.; Lei, W.; Yang, Y.; Tan, Q.; Xie, S.; Zhu, X.; Lei, W.; Yang, Y. Effect of dietary carbohydrate sources on growth performance and utilization for gibel carp (Carassius auratus gibelio) and Chinese longsnout catfish (Leiocassis longirostris Gunther). Aquac. Nutr. 2010, 12, 61–70. [Google Scholar] [CrossRef]
- Panserat, S.; Plagnes-Juan, E.; Kaushik, S. Gluconeogenic enzyme gene expression is decreased by dietary carbohydrates in common carp (Cyprinus carpio) and gilthead seabream (Sparus aurata). BBA-Gene Struct. Expr. 2002, 1579, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Sabat, P.; Río, C. Lack of significant long-term effect of dietary carbohydrates on hepatic glucose-6-phosphatase expression in rainbow trout (Oncorhynchus mykiss). J. Nutr. Biochem. 2000, 11, 22–29. [Google Scholar]
- TranulisMichael, A.; Dregni, O.; Christophersen, B.; Krogdahi, A.; Borrebaek, B. A glucokinase-like enzyme in the liver of atlantic salmon (Salmo salar). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1996, 114, 35–39. [Google Scholar]
- Pilkis, S.J. Molecular Physiology Of The Regulation Of Hepatic Gluconeogenesis And Glycolysis. Annu. Rev. Physiol. 1992, 54, 885–909. [Google Scholar] [CrossRef]
- Krasnov, A.; Teerijoki, H.; Molsa, H. Rainbow trout (Onchorhynchus mykiss) hepatic glucose transporter. Biochim. Biophys. Acta 2001, 1520, 174–178. [Google Scholar] [CrossRef]
- Uldry, M.; Ibberson, M.; Hosokawa, M.; Thorensã, B. GLUT2 is a high a⁄nity glucosamine transporter. FEBS Lett. 2002, 524, 199–203. [Google Scholar] [CrossRef]
- Terova, G.; Rimoldi, S.; Brambilla, F.; Gornati, R.; Saroglia, M. In vivo regulation of GLUT2 mRNA in sea bass (Dicentrarchus labrax) in response to acute and chronic hypoxia. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2009, 152, 306–316. [Google Scholar] [CrossRef]
- Panserat, S.; Plagnes-Juan, E.; Kaushik, S.J. Nutritional regulation and tissue specifity of gene expression for proteins involved in hepatic glucose metabolism in rainbow trout (Oncorhynchus mykiss). J. Exp. Biol. 2001, 204, 2351–2360. [Google Scholar] [CrossRef] [PubMed]
- Hakim, Y.; Uni, Z.; Hulata, G.; Harpaz, S. Relationship between intestinal brush border enzymatic activity and growth rate in tilapias fed diets containing 30% or 48% protein. Aquaculture 2006, 257, 420–428. [Google Scholar] [CrossRef]
- Couto, A.; Enes, P.; Peres, H.; Oliva-Teles, A. Effect of water temperature and dietary starch on growth and metabolic utilization of diets in gilthead sea bream (Sparus aurata) juveniles. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2008, 151, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Ren, M.; Ai, Q.; Mai, K.; Ma, H.; Wang, X. Effect of dietary carbohydrate level on growth performance, body composition, apparent digestibility coefficient and digestive enzyme activities of juvenile cobia, Rachycentron canadum L. Aquac. Res. 2011, 42, 1467–1475. [Google Scholar] [CrossRef]
- Zhou, Y.; Yuan, X.; Liang, X.F.; Fang, L.; Li, J.; Guo, X.; Bai, X.; He, S. Enhancement of growth and intestinal flora in grass carp: The effect of exogenous cellulase. Aquaculture 2013, 416–417, 1–7. [Google Scholar] [CrossRef]
- Jiang, T.T.; Feng, L.; Liu, Y.; Jiang, W.D.; Jiang, J.; Li, S.H.; Tang, L.; Kuang, S.Y.; Zhou, X.Q. Effects of exogenous xylanase supplementation in plant protein-enriched diets on growth performance, intestinal enzyme activities and microflora of juvenile Jian carp (Cyprinus carpio var. Jian). Aquac. Nutr. 2014, 20, 632–645. [Google Scholar] [CrossRef]
- Lin, S.; Mai, K.; Tan, B. Effects of exogenous enzyme supplementation in diets on growth and feed utilization in tilapia, Oreochromis niloticus x O. aureus. Aquac. Res. 2007, 38, 1645–1653. [Google Scholar] [CrossRef]
- Song, Z.; Wu, T. Progress in the study of normal intestinal flora in fish. Aquat. Sci. 2007, 26, 471–474. [Google Scholar]
- Hugo, C.J.; Bruun, B.; Jooste, P.J. The genera empedobacter and myroides. Prokaryotes 2006, 7, 630–637. [Google Scholar]
- Oliphant, K.; Allen-Vercoe, E. Macronutrient metabolism by the human gut microbiome: Major fermentation by-products and their impact on host health. Microbiome 2019, 7, 91. [Google Scholar] [CrossRef]
- Su, X.L.; Tian, Q.; Zhang, J.; Yuan, X.Z.; Shi, X.S.; Guo, R.B.; Qiu, Y.L. Acetobacteroides hydrogenigenes gen. nov., sp. nov., an anaerobic hydrogen-producing bacterium in the family Rikenellaceae isolated from a reed swamp. Int. J. Syst. Evol. Microbiol. 2014, 64, 2986–2991. [Google Scholar] [CrossRef] [PubMed]
- Du, S.; Sun, X.; Zhang, J.; Lin, D.; Chen, R.; Cui, Y.; Xiang, S.; Wu, Z.; Ding, T. Metagenome-Assembled Genomes Reveal Mechanisms of Carbohydrate and Nitrogen Metabolism of Schistosomiasis-Transmitting Vector Biomphalaria Glabrata. Microbiol. Spectr. 2022, 10, e01843-21. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (%) | Groups | |||||
|---|---|---|---|---|---|---|
| L | M | H | LE | ME | HE | |
| Fishmeal a | 26 | 26 | 26 | 26 | 26 | 26 |
| Casein a | 25 | 25 | 25 | 25 | 25 | 25 |
| Wheat gluten a | 6 | 6 | 6 | 6 | 6 | 6 |
| Corn starch a | 15 | 24 | 33 | 15 | 24 | 33 |
| Fish oil a | 12.63 | 8.63 | 4.37 | 12.63 | 8.63 | 4.37 |
| Soybean lecithin a | 1 | 1 | 1 | 1 | 1 | 1 |
| Vitamin premix b | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 |
| Mineral premix c | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 |
| Enzyme | 0 | 0 | 0 | 0.05 | 0.05 | 0.05 |
| Calcium monophosphate d | 1 | 1 | 1 | 1 | 1 | 1 |
| Choline chloride d | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 |
| Antioxidants a | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 | 0.03 |
| Attractant a | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 |
| Vitamin C a | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 |
| Carboxymethyl cellulose e | 2 | 2 | 2 | 2 | 2 | 2 |
| Microcrystalline cellulose e | 9.99 | 4.99 | 0.25 | 9.94 | 4.94 | 0.25 |
| Total | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 |
| Proximate composition | ||||||
| Moisture | 10.85 | 10.06 | 11.11 | 9.57 | 10.62 | 10.67 |
| Crude protein | 43.20 | 43.14 | 44.12 | 43.42 | 43.49 | 43.57 |
| Crude lipid | 15.96 | 12.27 | 8.03 | 16.06 | 12.05 | 8.73 |
| Ash | 6.46 | 6.3 | 6.25 | 6.18 | 6.24 | 6.34 |
| Carbohydrate | 15 | 24 | 33 | 15 | 24 | 33 |
| Carbohydrate/lipid | 0.91 | 1.92 | 3.91 | 0.91 | 1.92 | 3.91 |
| Gross energy (MJ Kg−1) | 19.62 | 19.58 | 19.54 | 19.62 | 19.58 | 19.54 |
| Primer name | Forward (5′-3′) | Reverse (5′-3′) | Accession No. |
|---|---|---|---|
| Glut2 | CTCCGATTCCAGAACGACTACTC | TGGAATAGGACCAGGACCGAT | KY656467.1 |
| β-Actin | TACGAGCTGCCTGACGGACA | GGCTGTGATCTCCTTCTGC | AY510710.2 |
| Groups | CHO:L | Enzyme | IBW(g) | FBW(g) | WGR(%) | SGR(% day−1) | SR(%) | FCR |
|---|---|---|---|---|---|---|---|---|
| L | 0.91 | 9.78 ±0.01 a | 44.48 ± 4.39 a | 333.40 ± 26.71 a | 2.82 ± 0.12 a | 95.56 ± 3.85 a | 1.20 ± 0.08 b | |
| M | 1.92 | 9.78 ± 0.00 a | 54.27 ± 2.38 b | 448.60 ± 22.61 cd | 3.27 ± 0.08 cd | 98.89 ± 1.92 a | 0.96 ± 0.10 a | |
| H | 3.91 | 9.78 ± 0.01 a | 50.66 ± 2.55 ab | 412.05 ± 21.21 bc | 3.14 ± 0.08 bc | 98.89 ± 1.92 a | 1.07 ± 0.07 ab | |
| LE | 0.91 | E | 9.78 ± 0.01 a | 47.12 ± 1.35 a | 371.00 ± 14.35 ab | 2.98 ± 0.06 ab | 97.78 ± 1.92 a | 1.16 ± 0.04 b |
| ME | 1.92 | E | 9.78 ± 0.01 a | 56.39 ± 2.21 b | 463.61 ± 19.12 cd | 3.32 ± 0.07 cd | 97.78 ± 1.92 a | 0.94 ± 0.08 a |
| HE | 3.91 | E | 9.78 ± 0.00 a | 57.02 ± 1.26 b | 476.41 ± 7.97 d | 3.36 ± 0.03 d | 98.89 ± 1.92 a | 0.93 ± 0.06 a |
| Means of main effect | ||||||||
| L | 0.91 | 9.78 | 45.80 A | 352.20 A | 2.90 A | 96.67 | 1.19 B | |
| M | 1.92 | 9.78 | 55.33 B | 456.11 B | 3.30 B | 98.33 | 0.95 A | |
| H | 3.91 | 9.78 | 53.84 B | 444.23 B | 3.25 B | 98.89 | 1.00 A | |
| No | 9.78 | 49.80 X | 398.02 X | 3.08 X | 97.78 | 1.082 | ||
| Have | 9.78 | 53.51 Y | 437.01 Y | 3.22 Y | 98.15 | 1.012 | ||
| Two-way ANOVA: p-values | ||||||||
| CHO:L | 1.00 | 0.010 | <0.001 | <0.001 | 0.194 | <0.001 | ||
| Enzyme | 0.78 | <0.001 | 0.001 | 0.002 | 0.090 | 0.06 | ||
| CHO:L*Enzyme | 0.49 | 0.333 | 0.135 | 0.178 | 0.115 | 0.30 | ||
| Groups | CHO:L | Enzyme | Moisture | Crude Protein | Crude Lipid | Ash |
|---|---|---|---|---|---|---|
| L | 0.91 | 66.70 ± 4.56 | 58.42 ± 2.09 | 28.50 ± 1.74 c | 14.53 ± 0.41 | |
| M | 1.92 | 69.93 ± 1.43 | 58.36 ± 1.40 | 26.26 ± 2.19 bc | 13.90 ± 0.57 | |
| H | 3.91 | 70.92 ± 9.17 | 59.06 ± 1.60 | 22.29 ± 2.28 ab | 13.85 ± 0.74 | |
| LE | 0.91 | E | 74.02 ± 6.92 | 57.33 ± 2.54 | 28.27 ± 1.66 c | 13.68 ± 0.13 |
| ME | 1.92 | E | 67.83 ± 5.52 | 58.42 ± 1.52 | 27.37 ± 1.82 bc | 13.80 ± 0.36 |
| HE | 3.91 | E | 71.62 ± 3.47 | 59.59 ± 2.36 | 19.60 ± 1.79 a | 14.56 ± 0.52 |
| Means of main effect | ||||||
| L | 0.91 | 70.36 | 57.87 | 28.39 B | 14.11 | |
| M | 1.92 | 68.88 | 58.39 | 26.82 B | 13.85 | |
| H | 3.91 | 71.27 | 59.33 | 20.94 A | 14.21 | |
| No | 69.18 | 58.611 | 25.69 | 14.09 | ||
| Have | 71.16 | 58.447 | 25.08 | 14.01 | ||
| Two-way ANOVA: p-values | ||||||
| CHO:L | 0.770 | 0.456 | <0.001 | 0.453 | ||
| Enzyme | 0.479 | 0.862 | 0.521 | 0.740 | ||
| CHO:L*Enzyme | 0.373 | 0.767 | 0.262 | 0.055 | ||
| Groups | CHO:L | Enzyme | TG (mmol/L) | TC (mmol/L) | GLU (mmol/mL) | INS (m U/L) |
|---|---|---|---|---|---|---|
| L | 0.91 | 0.61 ± 0.04 d | 2.87 ± 0.08 d | 4.79 ± 0.09 b | 29.59 ± 0.70 a | |
| M | 1.92 | 0.49 ± 0.04 c | 1.79 ± 0.21 c | 6.10 ± 0.38 c | 38.86 ± 0.49 b | |
| H | 3.91 | 0.41 ± 0.06 abc | 1.14 ± 0.10 ab | 7.46 ± 0.17 d | 47.64 ± 2.25 cd | |
| LE | 0.91 | E | 0.49 ± 0.02 bc | 1.90 ± 0.03 c | 4.24 ± 0.25 b | 44.37 ± 1.41 c |
| ME | 1.92 | E | 0.39 ± 0.02 ab | 1.40 ± 0.16 b | 4.18 ± 0.58 b | 50.96 ± 1.71 d |
| HE | 3.91 | E | 0.31 ± 0.04 a | 0.89 ± 0.20 a | 3.10 ± 0.25 a | 58.34 ± 1.29 e |
| Means of main effect | ||||||
| L | 0.91 | 0.55 C | 2.39 C | 4.52 A | 36.98 A | |
| M | 1.92 | 0.44 B | 1.60 B | 5.14 B | 44.91 B | |
| H | 3.91 | 0.36 A | 1.02 A | 5.28 B | 52.99 C | |
| No | 0.50 X | 1.936 X | 6.11 X | 38.69 X | ||
| Have | 0.40 Y | 1.396 Y | 3.84 Y | 51.22 Y | ||
| Two-way ANOVA: p-values | ||||||
| CHO:L | <0.001 | <0.001 | 0.004 | <0.001 | ||
| Enzyme | <0.001 | <0.001 | <0.001 | <0.001 | ||
| CHO:L*Enzyme | 0.791 | 0.002 | <0.001 | 0.081 | ||
| Groups | CHO:L | Enzyme | Trypsin(U/mg Prot) | Amylase(IU/mg Prot) | Lipase(mU/mg Prot) |
|---|---|---|---|---|---|
| L | 0.91 | 2868.34 ± 117.13 a | 252.22 ± 14.46 a | 1115.49 ± 23.44 b | |
| M | 1.92 | 3997.42 ± 156.84 bc | 553.33 ± 12.79 b | 960.36 ± 26.88 a | |
| H | 3.91 | 3933.64 ± 160.74 bc | 673.75 ± 8.98 c | 920.68 ± 36.56 a | |
| LE | 0.91 | E | 3585.75 ± 166.99 b | 261.73 ± 27.35 a | 1131.23 ± 52.55 b |
| ME | 1.92 | E | 4457.23 ± 155.06 d | 618.85 ± 33.17 c | 1006.00 ± 19.95 a |
| HE | 3.91 | E | 4281.28 ± 178.43 cd | 1000.99 ± 22.07 d | 933.43 ± 27.09 a |
| Means of main effect | |||||
| L | 0.91 | 3227.04 A | 256.98 A | 1123.36 C | |
| M | 1.92 | 4227.32 B | 586.09 B | 983.18 B | |
| H | 3.91 | 4107.46 B | 837.36 C | 927.06 A | |
| No | 3599.80 X | 493.10 X | 998.84 | ||
| Have | 4108.09 Y | 627.19 Y | 1023.55 | ||
| Two-way ANOVA: p-values | |||||
| CHO:L | <0.001 | <0.001 | <0.001 | ||
| Enzyme | <0.001 | <0.001 | 0.137 | ||
| CHO:L*Enzyme | 0.155 | <0.001 | 0.645 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Pan, L.; Shen, J.; Tan, B.; Dong, X.; Yang, Q.; Chi, S.; Zhang, S. Effects of Carbohydrase Supplementation on Growth Performance, Intestinal Digestive Enzymes and Flora, Glucose Metabolism Enzymes, and glut2 Gene Expression of Hybrid Grouper (Epinephelus fuscoguttatus♀ × E. lanceolatus♂) Fed Different CHO/L Ratio Diets. Metabolites 2023, 13, 98. https://doi.org/10.3390/metabo13010098
Liu H, Pan L, Shen J, Tan B, Dong X, Yang Q, Chi S, Zhang S. Effects of Carbohydrase Supplementation on Growth Performance, Intestinal Digestive Enzymes and Flora, Glucose Metabolism Enzymes, and glut2 Gene Expression of Hybrid Grouper (Epinephelus fuscoguttatus♀ × E. lanceolatus♂) Fed Different CHO/L Ratio Diets. Metabolites. 2023; 13(1):98. https://doi.org/10.3390/metabo13010098
Chicago/Turabian StyleLiu, Hongyu, Ling Pan, Jianfei Shen, Beiping Tan, Xiaohui Dong, Qihui Yang, Shuyan Chi, and Shuang Zhang. 2023. "Effects of Carbohydrase Supplementation on Growth Performance, Intestinal Digestive Enzymes and Flora, Glucose Metabolism Enzymes, and glut2 Gene Expression of Hybrid Grouper (Epinephelus fuscoguttatus♀ × E. lanceolatus♂) Fed Different CHO/L Ratio Diets" Metabolites 13, no. 1: 98. https://doi.org/10.3390/metabo13010098
APA StyleLiu, H., Pan, L., Shen, J., Tan, B., Dong, X., Yang, Q., Chi, S., & Zhang, S. (2023). Effects of Carbohydrase Supplementation on Growth Performance, Intestinal Digestive Enzymes and Flora, Glucose Metabolism Enzymes, and glut2 Gene Expression of Hybrid Grouper (Epinephelus fuscoguttatus♀ × E. lanceolatus♂) Fed Different CHO/L Ratio Diets. Metabolites, 13(1), 98. https://doi.org/10.3390/metabo13010098