
miR-1/AMPK-Mediated Glucose and Lipid Metabolism under Chronic Hypothermia in the Liver of Freshwater Drum, Aplodinotus grunniens


 ,
,
Abstract
1. Introduction
2. Results
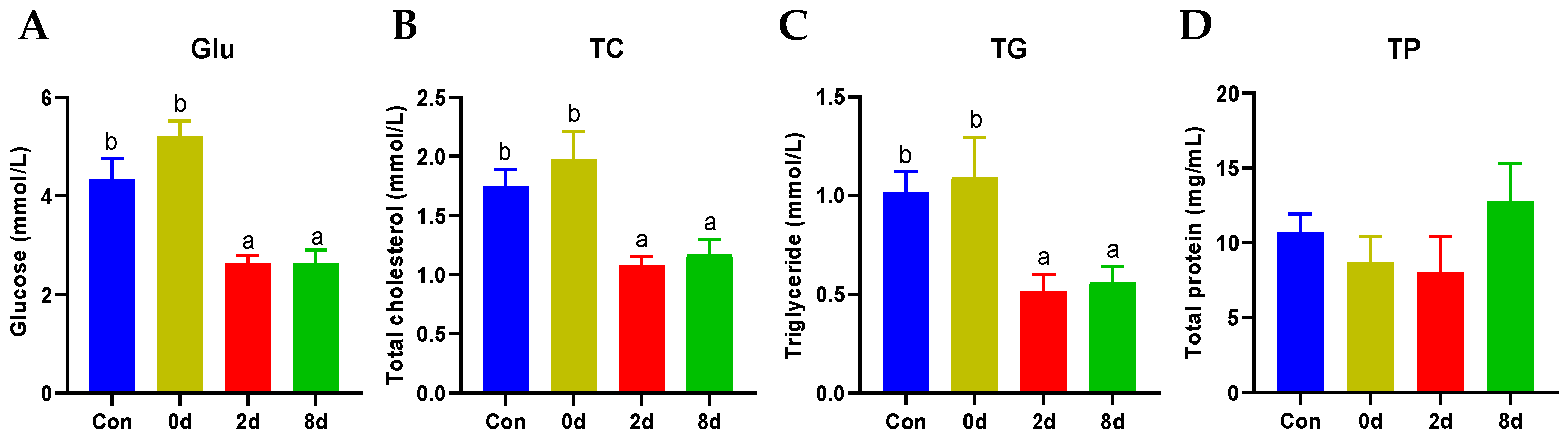
2.1. Effects of Chronic Hypothermia on Plasma Biochemical Parameters of Freshwater Drum
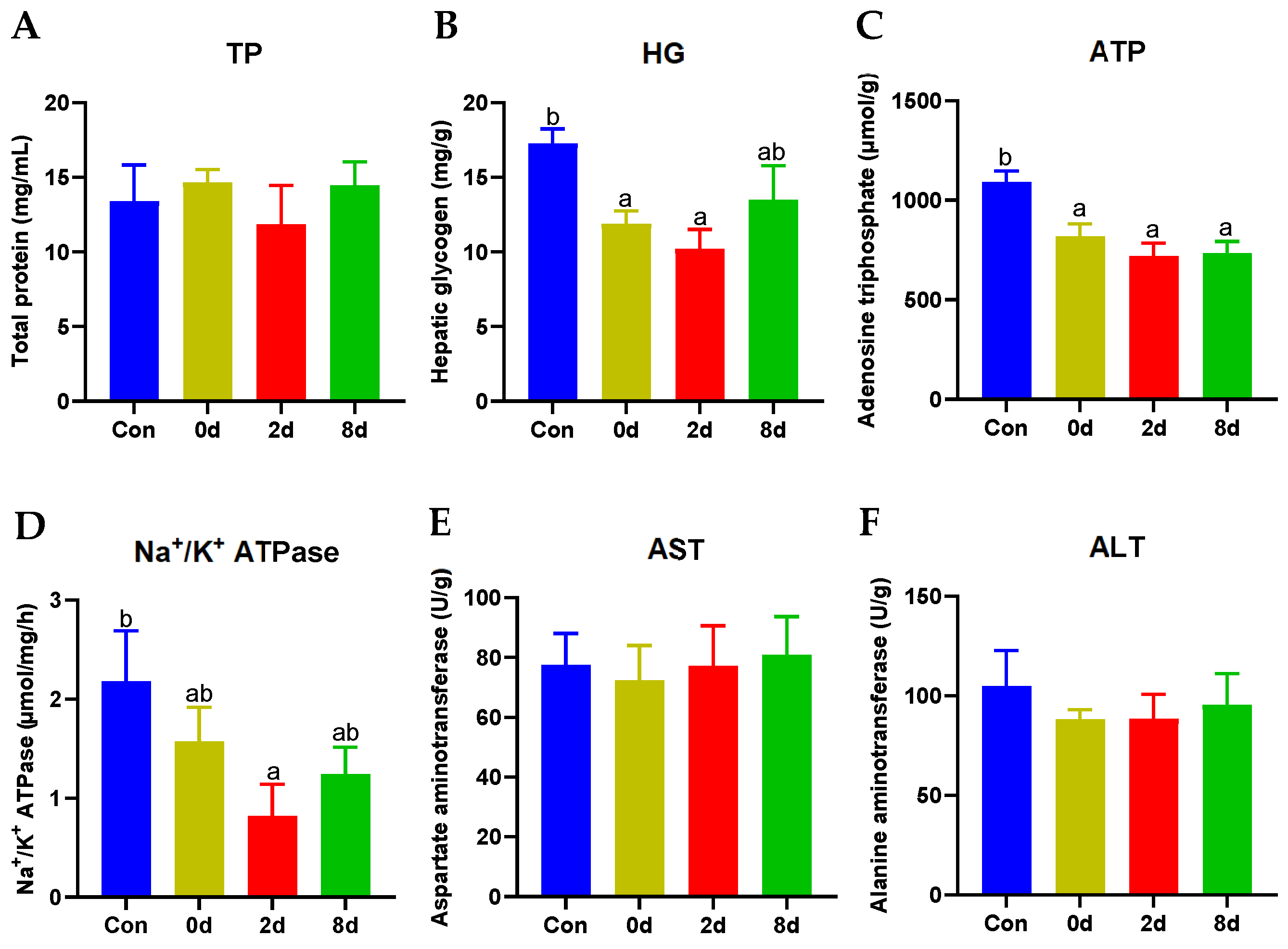
2.2. Effects of Chronic Hypothermia on Hepatic Biochemical Parameters of Freshwater Drum
2.3. Effects of Chronic Hypothermia on Glucometabolism of Freshwater Drum
2.4. Effects of Chronic Hypothermia on Lipid Metabolism of Freshwater Drum
2.5. miRNA Analysis Reveals AMPK Signaling Was Active to Glucose and Lipid Metabolism under Hypothermia Exposure
2.6. mRNA-miRNA Integrate Analysis Reveals miR-1 and AMPK Were Involved in Hypothermia Exposure
2.7. AMPK Signaling Was Dysregulated under Hypothermia Based on Transcriptomic Analysis
2.8. miR-1/AMPK Signaling Was Involved in Glucose and Fatty Acid Metabolism under Chronic Hypothermia
2.9. Epigenetic Schematic of AMPK Signaling under Hypothermia Induction in Freshwater Drum
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Experimental Animals and Rearing Conditions
4.3. Sample Collection
4.4. Plasma and Liver Biochemical Indicators Analysis
4.5. RNA Extraction and De Novo High-Throughput Sequencing
4.5.1. RNA Extraction, cDNA Library Construction, and RNA-Seq
4.5.2. De Novo Assembly, Functional Annotation and the Differentially Expressed Genes (DEGs) Analysis
4.6. miRNA Sequencing, Identification, and Target Gene Prediction
4.6.1. miRNA Sequencing
4.6.2. Differentially Expressed miRNAs (DEMs) Identification and Target Gene Prediction
4.7. Validation of Differentially Expressed Genes Obtained from RNA-Seq
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Song, C.; Cui, Y.; Liu, B.; Xie, J.; Ge, X.; Xu, P.; Lin, Y. HSP60 and HSP90β from blunt snout bream, Megalobrama amblycephala: Molecular cloning, characterization, and comparative response to intermittent thermal stress and Aeromonas hydrophila infection. Fish Shellfish Immunol. 2017, 74, 119–132. [Google Scholar] [CrossRef]
- Zhou, C.; Zhang, Z.; Zhang, L.; Liu, Y.; Liu, P.F. Effects of temperature on growth performance and metabolism of juvenile sea bass (Dicentrarchus labrax). Aquaculture 2021, 537, 736458. [Google Scholar] [CrossRef]
- Abram, Q.H.; Dixon, B.; Katzenback, B.A. Impacts of low temperature on the teleost immune system. Biology 2017, 6, 39. [Google Scholar] [CrossRef]
- Snyder, R.J.; Hennessey, T.M. Cold tolerance and homeoviscous adaptation in freshwater alewives (Alosa pseudoharengus). Fish Physiol. Biochem. 2003, 29, 117–126. [Google Scholar] [CrossRef]
- Donaldson, M.R.; Cooke, S.J.; Patterson, D.A.; Macdonald, J.S. Cold shock and fish. J. Fish Biol. 2008, 73, 1491–1530. [Google Scholar] [CrossRef]
- Kuo, C.M.; Hsieh, S.L. Comparisons of physiological and biochemical responses between milkfish (Chanos chanos) and grass carp (Ctenopharyngodon idella) to cold shock. Aquaculture 2006, 251, 525–536. [Google Scholar] [CrossRef]
- Qiang, J.; Yang, H.; Wang, H.; Kpundeh, M.D.; He, J.; Xu, P. Physiological responses and HSP70 mRNA expression in GIFT tilapia juveniles, Oreochromis niloticus under short-term crowding. Aquac. Res. 2015, 46, 335–345. [Google Scholar] [CrossRef]
- Farkas, T.; Csengeri, I.; Majoros, F.; Oláh, J. Metabolism of fatty acids in fish: III. Combined effect of environmental temperature and diet on formation and deposition of fatty acids in the carp, Cyprinus carpio Linnaeus 1758. Aquaculture 2015, 20, 29–40. [Google Scholar] [CrossRef]
- Sellner, P.A.; Hazel, J.R. Incorporation of polyunsaturated fatty acids into lipids of rainbow trout hepatocytes. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1982, 243, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Dzamko, N.L.; Steinberg, G.R. AMPK-dependent hormonal regulation of whole-body energy metabolism. Acta Physiol. 2009, 196, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Merrill, G.F.; Kurth, E.J.; Hardie, D.G.; Winder, W.W. AICA riboside increases AMP-activated protein kinase, fatty acid oxidation, and glucose uptake in rat muscle. Am. J. Physiol. 1997, 273, 1107–1112. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.T.; Kola, B.; Korbonits, M. AMPK as a mediator of hormonal signalling. J. Mol. Endocrinol. 2010, 44, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Healy, J.E.; Gearhart, C.N.; Bateman, J.L.; Handa, R.J.; Florant, G.L. AMPK and ACC change with fasting and physiological condition in euthermic and hibernating golden-mantled ground squirrels (Callospermophilus lateralis). Comp. Biochem. Physiol. Part A 2011, 159, 322–331. [Google Scholar] [CrossRef]
- McGarry, J.D.; Mills, S.E.; Long, C.S.; Foster, D.W. Observations on the affinity for carnitine, and malonyl-CoA sensitivity, of carnitine palmitoyltransferase I in animal and human tissues. Demonstration of the presence of malonyl-CoA in non-hepatic tissues of the rat. Biochem. J. 1983, 214, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Abu-Elheiga, L.; Matzuk, M.M.; Abo-hashema, K.A.H.; Wakil, S.J. Continuous Fatty Acid Oxidation and Reduced Fat Storage in Mice Lacking Acetyl-CoA Carboxylase 2. Science 2001, 291, 2613–2616. [Google Scholar] [CrossRef] [PubMed]
- Carlson, C.L.; Winder, W.W. Liver AMP-activated protein kinase and acetyl-CoA carboxylase during and after exercise. J. Appl. Physiol. 1999, 86, 669–674. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Horgan, R.P.; Kenny, L.C. ‘Omic’ technologies: Genomics, transcriptomics, proteomics and metabolomics. Obstet. Gynaecol. 2011, 13, 189–195. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef]
- Song, C.; Liu, B.; Ge, X.; Li, H.; Liu, B.; Xu, P. miR-34a/Notch1b mediated autophagy and apoptosis contributes to oxidative stress amelioration by emodin in the intestine of teleost Megalobrama amblycephala. Aquaculture 2021, 547, 737441. [Google Scholar] [CrossRef]
- Song, C.; Liu, B.; Xu, P.; Ge, X.; Tang, Y.; Li, H.; Su, S. mir-144 is the epigenetic target for emodin to ameliorate oxidative stress induced by dietary oxidized fish oil via Nrf2 signaling in Wuchang bream, Megalobrama amblycephala. Aquaculture 2021, 534, 736357. [Google Scholar] [CrossRef]
- Hernández-Gómez, R.E.; Contreras-Sánchez, W.M.; Hernández-Franyutti, A.; Perera-García, M.A.; Torres-Martínez, A. Testicular structure and development of the male germinal epithelium in the freshwater drum Aplodinotus grunniens (Perciformes: Sciaenidae) from the Usumacinta River, Southern Mexico. Acta Zool. 2021, 1–19. [Google Scholar] [CrossRef]
- Song, C.; Wen, H.; Liu, G.; Ma, X.; Lv, G.; Wu, N.; Chen, J.; Xue, M.; Li, H.; Xu, P. Gut Microbes Reveal Pseudomonas Medicates Ingestion Preference via Protein Utilization and Cellular Homeostasis Under Feed Domestication in Freshwater Drum, Aplodinotus grunniens. Front. Microbiol. 2022, 861705. [Google Scholar] [CrossRef] [PubMed]
- William, B.W. Life history aspects of smallmouth buffalo and freshwater drum in wheeler reservoir, Alabama. Proc. Annu. Conf. Southeast. Assoc. Game Fish Comm. 1968, 22, 479–495. [Google Scholar]
- Bodensteiner, L.S.; Lewis, W.M. Role of Temperature, Dissolved Oxygen, and Backwaters in the Winter Survival of Freshwater Drum (Aplodinotus grunniens) in the Mississippi River. Can. J. Fish. Aquat. Sci. 1992, 49, 173–184. [Google Scholar] [CrossRef]
- Ou, Y.J. Impact on fishery production by frost disaster in southern China. Chin. Fish. Econ. 2008, 26, 89–93. [Google Scholar]
- Lahnsteiner, F.; Caberlotto, S. Motility of gilthead seabream sparus aurata spermatozoa and its relation to temperature, energy metabolism and oxidative stress. Aquaculture 2012, 370–371, 76–83. [Google Scholar] [CrossRef]
- Pilkis, S.J.; Granner, D.K. Molecular Physiology of the Regulation of Hepatic Gluconeogenesis and Glycolysis. Annu. Rev. Physiol. 1992, 54, 885–909. [Google Scholar] [CrossRef]
- Metón, I.; Fernández, F.; Baanante, I.V. Short- and long-term effects of refeeding on key enzyme activities in glycolysis-gluconeogenesis in the liver of gilthead seabream (Sparus aurata). Aquaculture 2003, 225, 99–107. [Google Scholar] [CrossRef]
- Shaklee, J.B.; Christiansen, J.A.; Sidell, B.D.; Prosser, C.L.; Whitt, G.S. Molecular aspects of temperature acclimation in fish: Contributions of changes in enzyme activities and isozyme patterns to metabolic reorganization in the green sunfish. J. Exp. Zool. Part A Ecol. Genet. Physiol. 1977, 201, 1–20. [Google Scholar] [CrossRef]
- Mark, A.S. Lipid dynamics in fish: Aspects of absorption, transportation, deposition and mobilization. Comp. Biochem. Physiol. Part B Comp. Biochem. 1988, 90, 679–690. [Google Scholar] [CrossRef]
- Martins, D.A.; Rocha, F.; Castanheira, F.; Mendes, A.; Pousão-Ferreira, P.; Bandarra, N.; Coutinho, J.; Morais, S.; Yúfera, M.; Conceição, L.E.C.; et al. Effects of dietary arachidonic acid on cortisol production and gene expression in stress response in Senegalese sole (Solea senegalensis) post-larvae. Fish Physiol. Biochem. 2013, 39, 1223–1238. [Google Scholar] [CrossRef]
- Choi, S.S.; Diehl, A.M. Hepatic triglyceride synthesis and nonalcoholic fatty liver disease. Curr. Opin. Lipidol. 2008, 19, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Temkin, S.M.; Hawkridge, A.M.; Guo, C.; Wang, W.; Wang, X.Y.; Fang, X.J. Fatty acid oxidation: An emerging facet of metabolic transformation in cancer. Cancer Lett. 2018, 435, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Munday, M.R. Regulation of mammalian acetyl-coa carboxylase. Biochem. Soc. Trans. 2002, 30, 1059–1064. [Google Scholar] [CrossRef] [PubMed]
- Ke, R.; Xu, Q.; Li, C.; Luo, L.; Huang, D. Mechanisms of AMPK in the maintenance of ATP balance during energy metabolism. Cell Biol. Int. 2017, 42, 384–392. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Ye, C.; Guo, Z.; Wang, A. Immune and physiological responses of pufferfish (Takifugu obscurus) under cold stress. Fish Shellfish Immunol. 2017, 64, 137–145. [Google Scholar] [CrossRef]
- Ibarz, A.; Blasco, J.; Beltrán, M.; Gallardo, M.A.; Sánchez, J.; Sala, R.; Fernández-Borràs, J. Cold-induced alterations on proximate composition and fatty acid profiles of several tissues in gilthead sea bream (Sparus aurata). Aquaculture 2005, 249, 477–486. [Google Scholar] [CrossRef]
- Ibarz, A.; Beltrán, M.; Fernández-Borràs, J.; Gallardo, M.A.; Sánchez, J.; Blasco, J. Alterations in lipid metabolism and use of energy depots of gilthead sea bream (Sparus aurata) at low temperatures. Aquaculture 2007, 262, 470–480. [Google Scholar] [CrossRef]
- Galt, N.J.; Froehlich, J.M.; McCormick, S.D.; Biga, P.R. A comparative evaluation of crowding stress on muscle HSP90 and myostatin expression in salmonids. Aquaculture 2018, 483, 141–148. [Google Scholar] [CrossRef]
- Marqueze, A.; Garbino, C.F.; Trapp, M.; Kucharski, L.C.; Fagundes, M.; Ferreira, D.; Koakoski, G.; Rosa, J.G.S. Protein and lipid metabolism adjustments in silver catfish (Rhamdia quelen) during different periods of fasting and refeeding. Braz. J. Biol. 2018, 78, 464–471. [Google Scholar] [CrossRef]
- Fynn-Aikins, K.; Hughes, S.G.; Vandenberg, G.W. Protein retention and liver aminotransferase activities in Atlantic salmon fed diets containing different energy sources. Comp. Biochem. Physiol. Part A Physiol. 1995, 111, 163–170. [Google Scholar] [CrossRef]
- Li, S.; Ji, H.; Zhang, B.; Zhou, J.; Yu, H. Defatted black soldier fly (Hermetia illucens) larvae meal in diets for juvenile jian carp (cyprinus carpio var. jian): Growth performance, antioxidant enzyme activities, digestive enzyme activities, intestine and hepatopancreas histological structure. Aquaculture 2017, 477, 62–70. [Google Scholar] [CrossRef]
- Ambros, V. The functions of animal micrornas. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef]
- Sun, J.; Zhao, L.; Wu, H.; Lian, W.; Cui, C.; Du, Z.; Luo, W.; Li, M.; Yang, S. Analysis of miRNA-seq in the liver of common carp (Cyprinus carpio L.) in response to different environmental temperatures. Funct. Integr. Genom. 2019, 19, 265–280. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Dai, Z.; Chen, S.; Chen, L. MicroRNA-mediated gene regulation plays a minor role in the transcriptomic plasticity of cold-acclimated zebrafish brain tissue. BMC Genom. 2011, 12, 605. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Liu, Q.; Zhao, L.; Cui, C.; Wu, H.; Liao, L.; Tang, G.; Yang, S. Potential regulation by miRNAs on glucose metabolism in liver of common carp (Cyprinus carpio) at different temperatures. Comp. Biochem. Physiol. Part D Genom. Proteom. 2019, 32, 100628. [Google Scholar] [CrossRef] [PubMed]
- Jia, Q.; An, W.; Xu, Y.; Qiao, S.; Chen, W. Role of microrna-1-mediated amp-activated protein kinase pathway in cardiac fibroblasts induced by high glucose in rats. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue 2018, 30, 145–150. [Google Scholar] [CrossRef]
- Xu, W.; Zhang, Z.; Zou, K.; Cheng, Y.; Yang, M.; Chen, H.; Wang, H.; Zhao, J.; Chen, P.; He, L. MiR-1 suppresses tumor cell proliferation in colorectal cancer by inhibition of Smad3-mediated tumor glycolysis. Cell Death Dis. 2017, 8, e2761. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Chen, W.; Han, D.; Zhu, X.; Yang, Y.; Jin, J.; Liu, H.; Xie, S. Effects of food restriction on growth, body composition and gene expression related in regulation of lipid metabolism and food intake in grass carp. Aquaculture 2017, 469, 28–35. [Google Scholar] [CrossRef]
- Li, H.; Qiang, J.; Song, C.; Xu, P. Transcriptome profiling reveal Acanthopanax senticosus improves growth performance, immunity and antioxidant capacity by regulating lipid metabolism in GIFT (Oreochromis niloticus). Comp. Biochem. Physiol. Part D Genom. Proteom. 2020, 37, 100784. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Liu, B.; Xu, P.; Xie, J.; Ge, X.; Zhou, Q.; Sun, C.; Zhang, H.; Shan, F.; Yang, Z. Oxidized Fish Oil Injury Stress in Megalobrama Amblycephala: Evaluated by Growth, Intestinal Physiology, and Transcriptome-Based PI3K-Akt/NF-κB/TCR Inflammatory Signaling. Fish Shellfish Immunol. 2018, 81, 446–455. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence (5′-3′) |
|---|---|---|
| miRNA primer | ||
| miR-1 | F | CGGGAATGTAAAGAAGTATG |
| U6 | F | CTCGCTTCGGCAGCACA |
| R | AACGCTTCACGAATTTGCGT | |
| mRNA primer | ||
| AMPK | F | TCCCTCCTACAGCAACAAC |
| R | GACGCCAGGTAGAAATCC | |
| PEPCK | F | TCGTCTCATTCGGTAGTGGT |
| R | CGTAGGTTGCCTTGGTTGT | |
| G6Pase | F | GCGGACCTGAGGAACACCTT |
| R | AAACACCTGCGGCTCCCATA | |
| TORC2 | F | AAGTTTAGCGAGAAGATAGCG |
| R | TCAAGAGTGGATGGGAAGG | |
| PFK-2 | F | CCAGATAATGAGGAGGGTTT |
| R | TCTTGTTGTATTTGTGGCATC | |
| GS | F | GAGCCTCCGCACATCGTA |
| R | CGCCTGCTTCCTTATCCA | |
| SREBP1 | F | TTCCTCTCCCTCAACCCTCT |
| R | TTACGGGCTCTCCATACACC | |
| FAS | F | TGGCATCGAGTACAACAAGC |
| R | TTGGCACGAAGTAGCATCAC | |
| SCD1 | F | CGGGGCTTCTTCTTCTCTCA |
| R | GAAGCACATGACTAGCACGG | |
| ACC1 | F | CTGGAGGAGACGGTGAAAAG |
| R | TGCGTATCTGCTTGAGGATG | |
| CPT1 | F | GCACCAGAACCTTTACCGA |
| R | TCCGCCCAGTGATATGAAC | |
| eEF-2 | F | TCAGCCCTGTGGTGAGAAT |
| R | TGTGCTTGTTGGGTGACTTT | |
| β-actin | F | AGGCTGTGCTGTCCCTGTAT |
| R | GCTGTGGTGGTGAAGGAGTAG | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.; Song, C.; Wen, H.; Liu, G.; Wu, N.; Li, H.; Xue, M.; Xu, P. miR-1/AMPK-Mediated Glucose and Lipid Metabolism under Chronic Hypothermia in the Liver of Freshwater Drum, Aplodinotus grunniens. Metabolites 2022, 12, 697. https://doi.org/10.3390/metabo12080697
Chen J, Song C, Wen H, Liu G, Wu N, Li H, Xue M, Xu P. miR-1/AMPK-Mediated Glucose and Lipid Metabolism under Chronic Hypothermia in the Liver of Freshwater Drum, Aplodinotus grunniens. Metabolites. 2022; 12(8):697. https://doi.org/10.3390/metabo12080697
Chicago/Turabian StyleChen, Jianxiang, Changyou Song, Haibo Wen, Guangxiang Liu, Ningyuan Wu, Hongxia Li, Miaomiao Xue, and Pao Xu. 2022. "miR-1/AMPK-Mediated Glucose and Lipid Metabolism under Chronic Hypothermia in the Liver of Freshwater Drum, Aplodinotus grunniens" Metabolites 12, no. 8: 697. https://doi.org/10.3390/metabo12080697
APA StyleChen, J., Song, C., Wen, H., Liu, G., Wu, N., Li, H., Xue, M., & Xu, P. (2022). miR-1/AMPK-Mediated Glucose and Lipid Metabolism under Chronic Hypothermia in the Liver of Freshwater Drum, Aplodinotus grunniens. Metabolites, 12(8), 697. https://doi.org/10.3390/metabo12080697