
The Potential Role of Cyclopeptides from Pseudostellaria heterophylla, Linum usitatissimum and Drymaria diandra, and Peptides Derived from Heterophyllin B as Dipeptidyl Peptidase IV Inhibitors for the Treatment of Type 2 Diabetes: An In Silico Study


Abstract
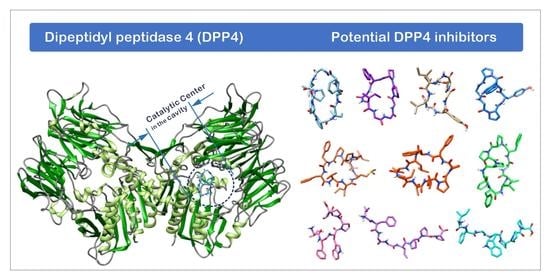
:
1. Introduction
1.1. About Diabetes
1.2. Mechanisms of Incretins (GLP-1 and GIP) in Glucose Homeostasis and Diabetes Treatment
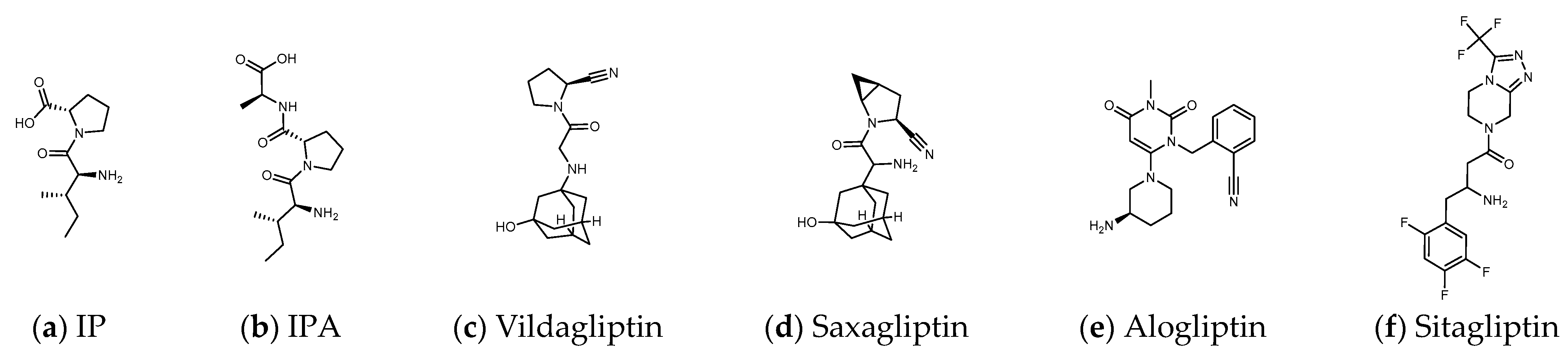
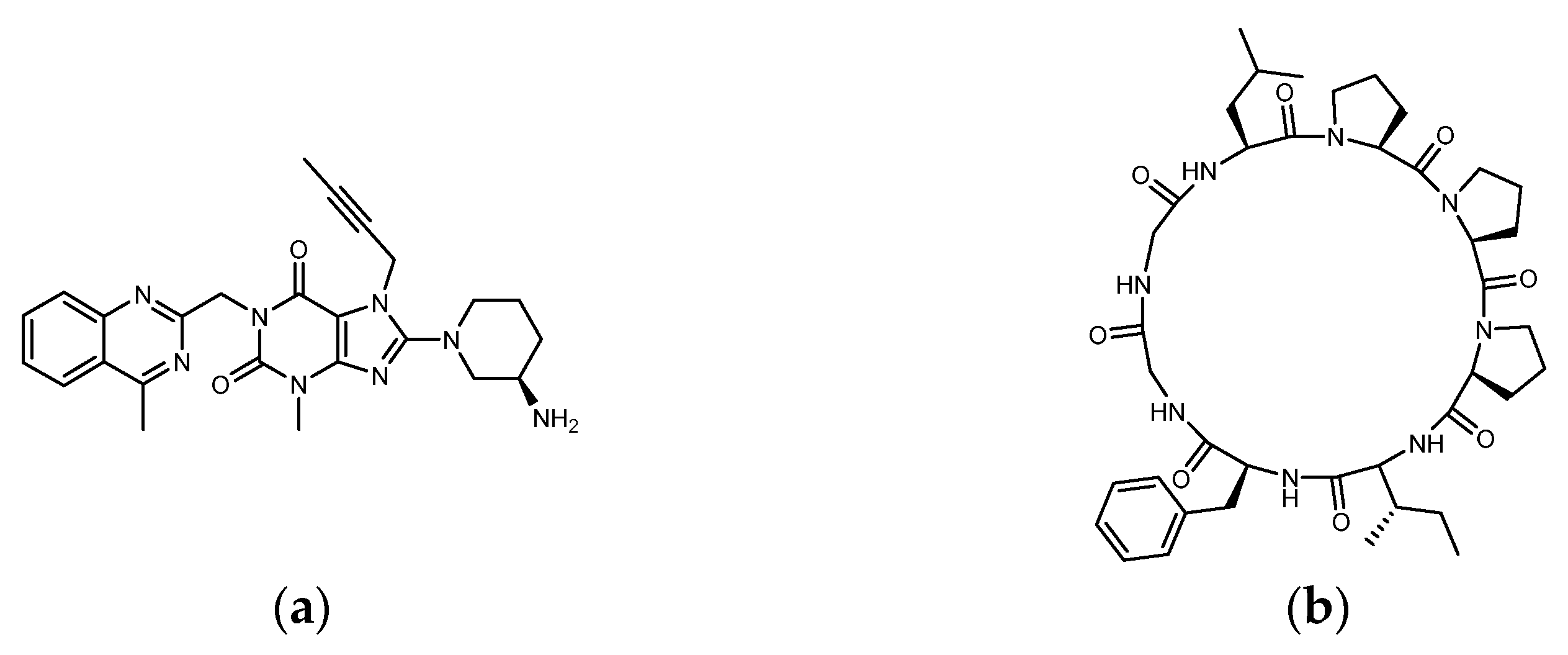
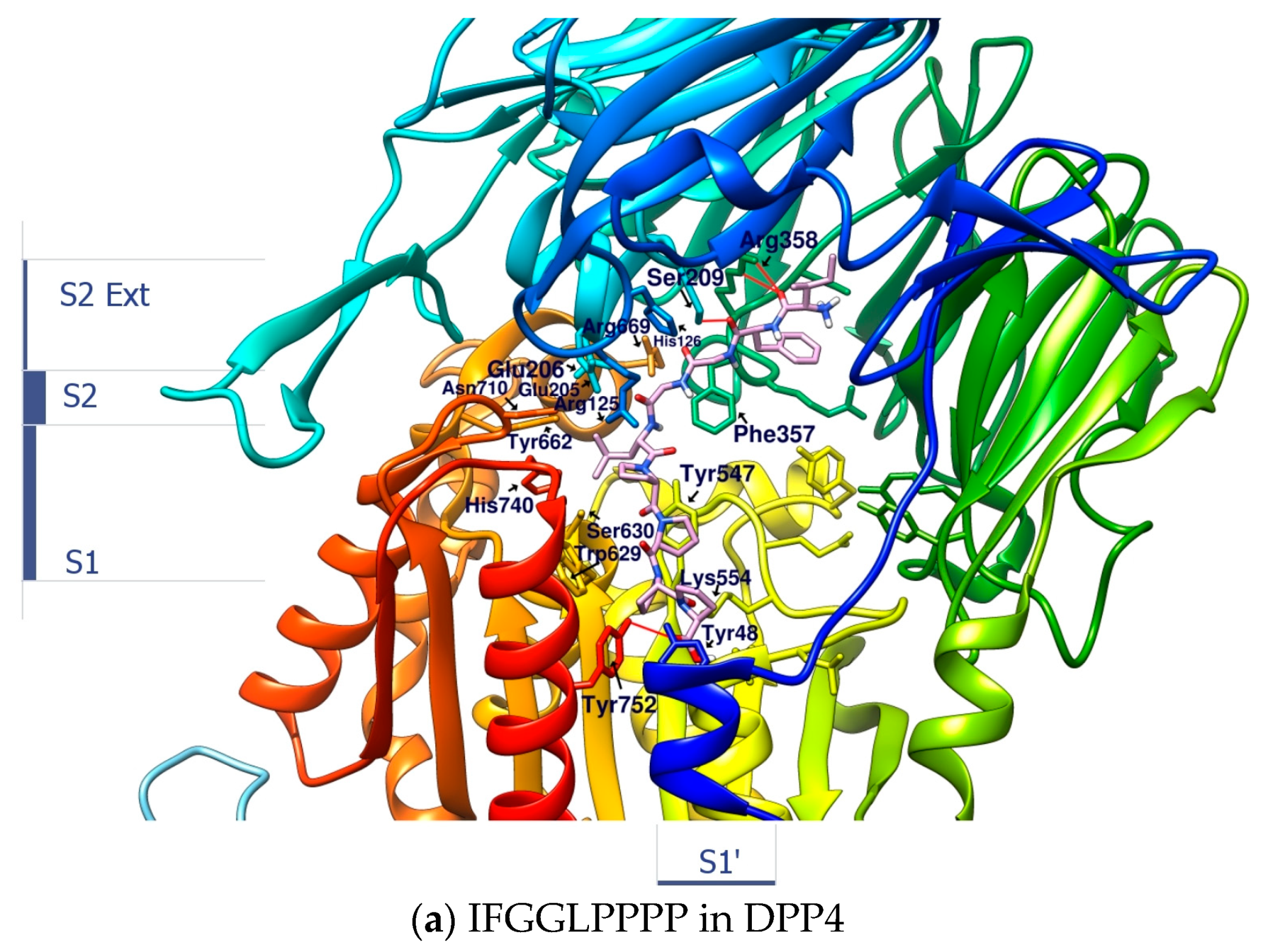
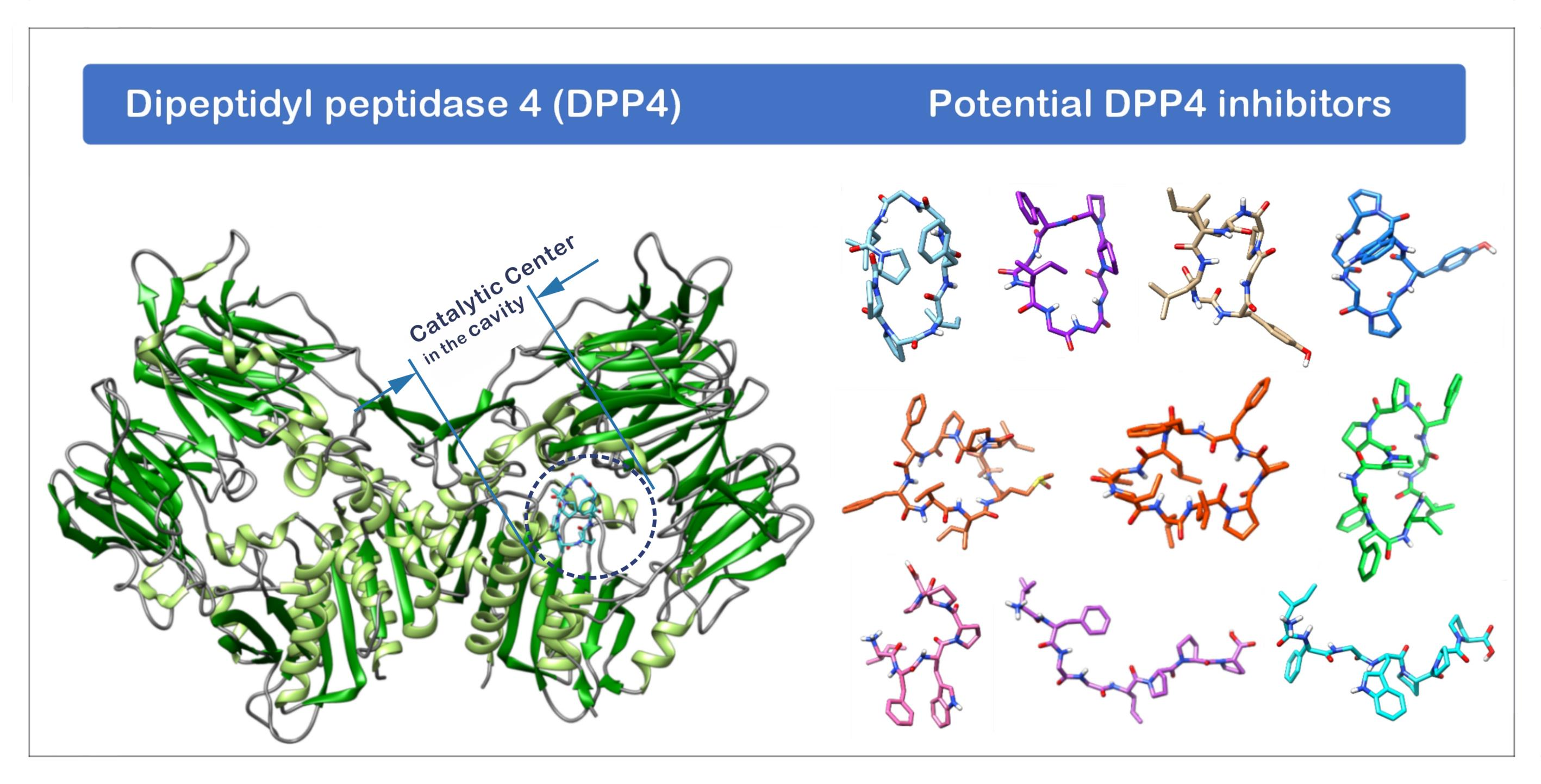
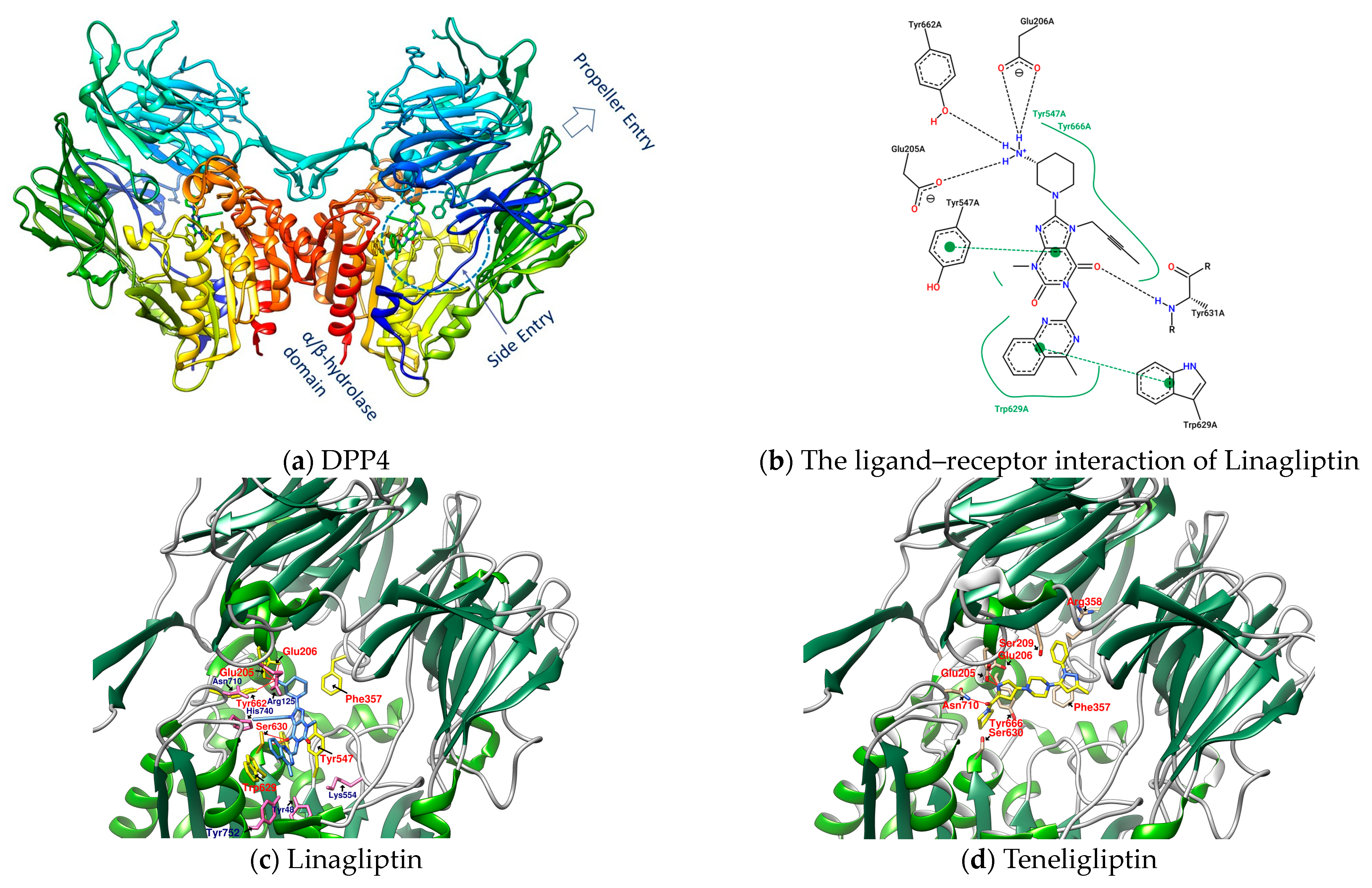
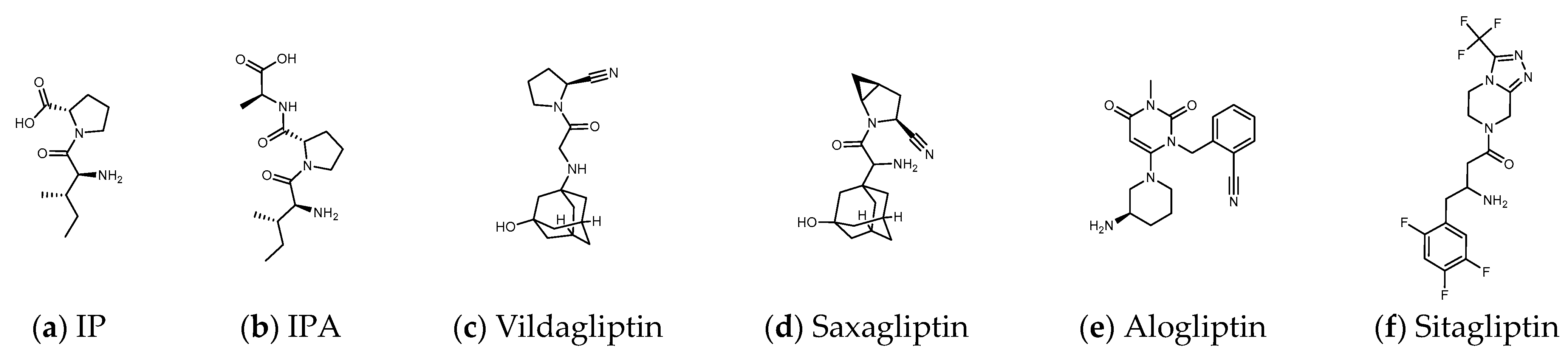
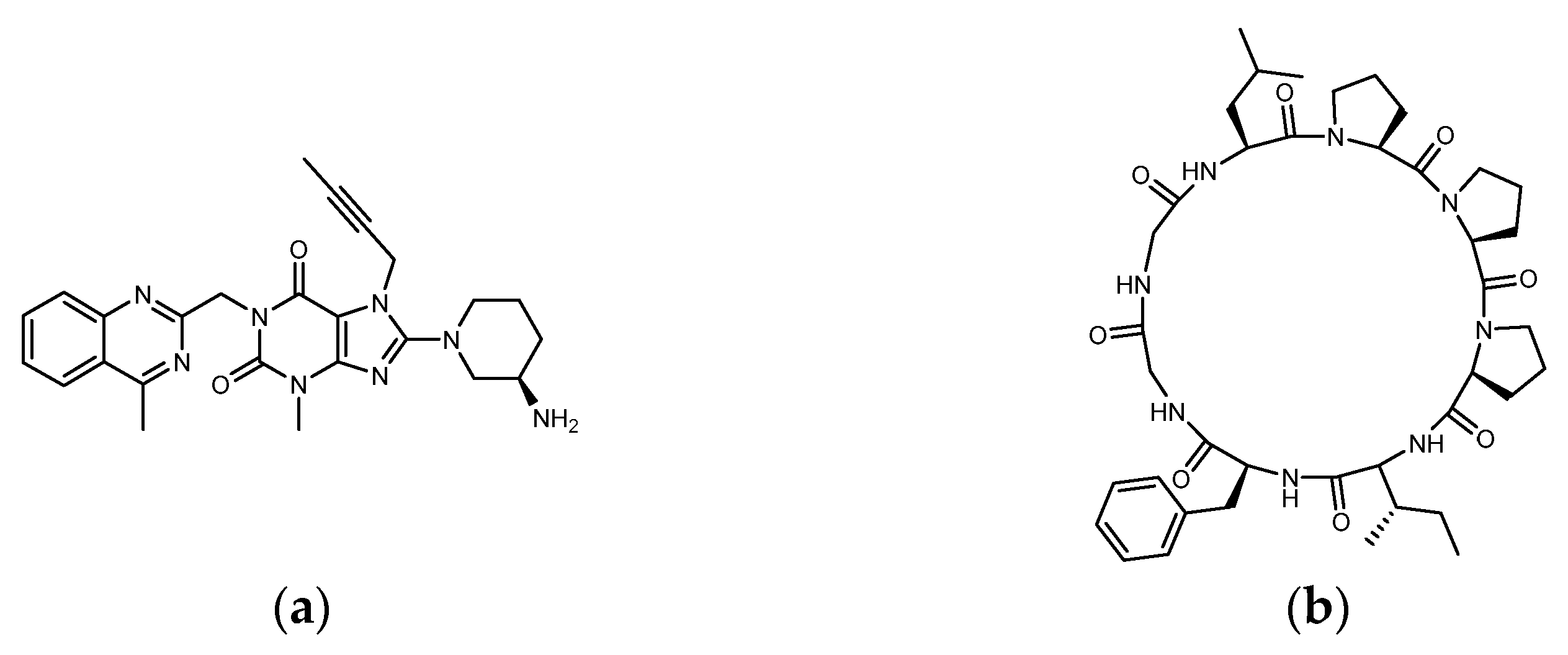
1.3. The Structure of DPP4 and the Interaction of DPP4 Inhibitors with DPP4
1.4. Natural Products with Relevant Reports on Lowering Blood Sugar and Their Mechanisms
1.5. Pseudostellaria Heterophylla, a Reported Natural Product with Hypoglycemic Effect
1.6. Linum Usitatissimum, Which Is Rich in Cyclic Peptides and Has Hypoglycemic Effect Reported
1.7. Drymaria Diandra, Which Is Rich in Cyclic Peptides and Has Hypoglycemic Effect Reported
1.8. Can Linear Precursors of Heterophyllin B “IFGGLPPP” Participate in DPP4 Inhibition?
2. Results and Discussion
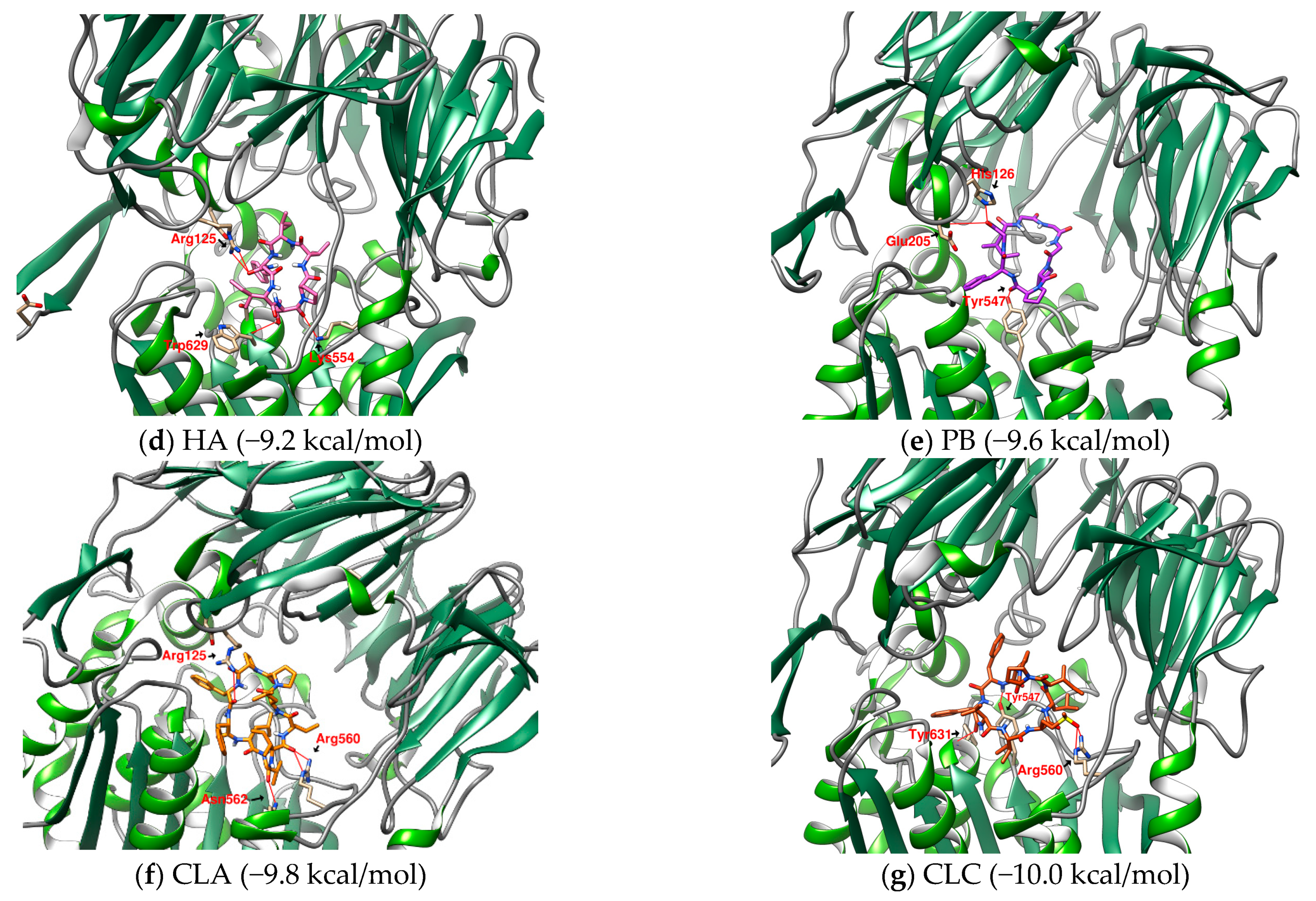
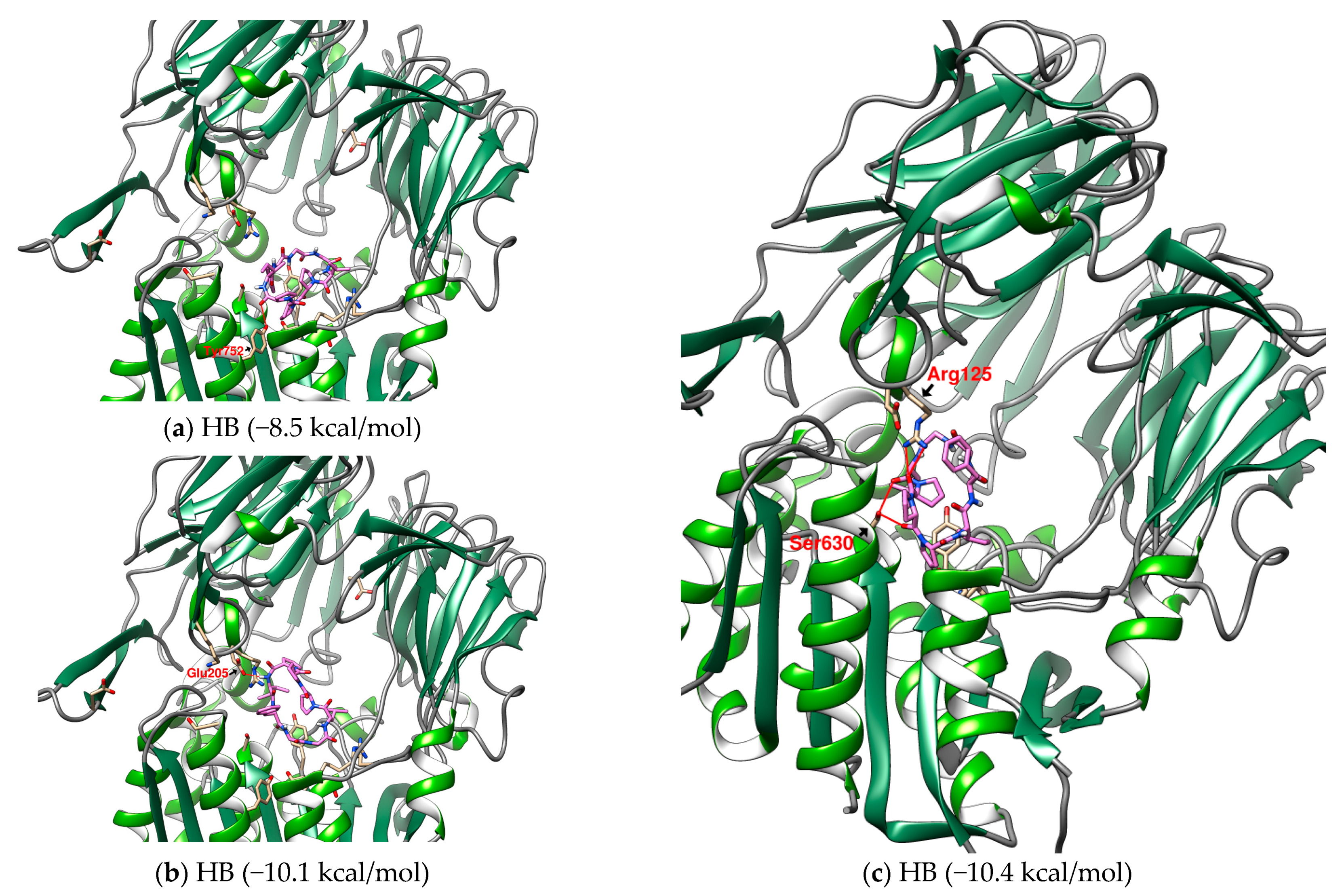
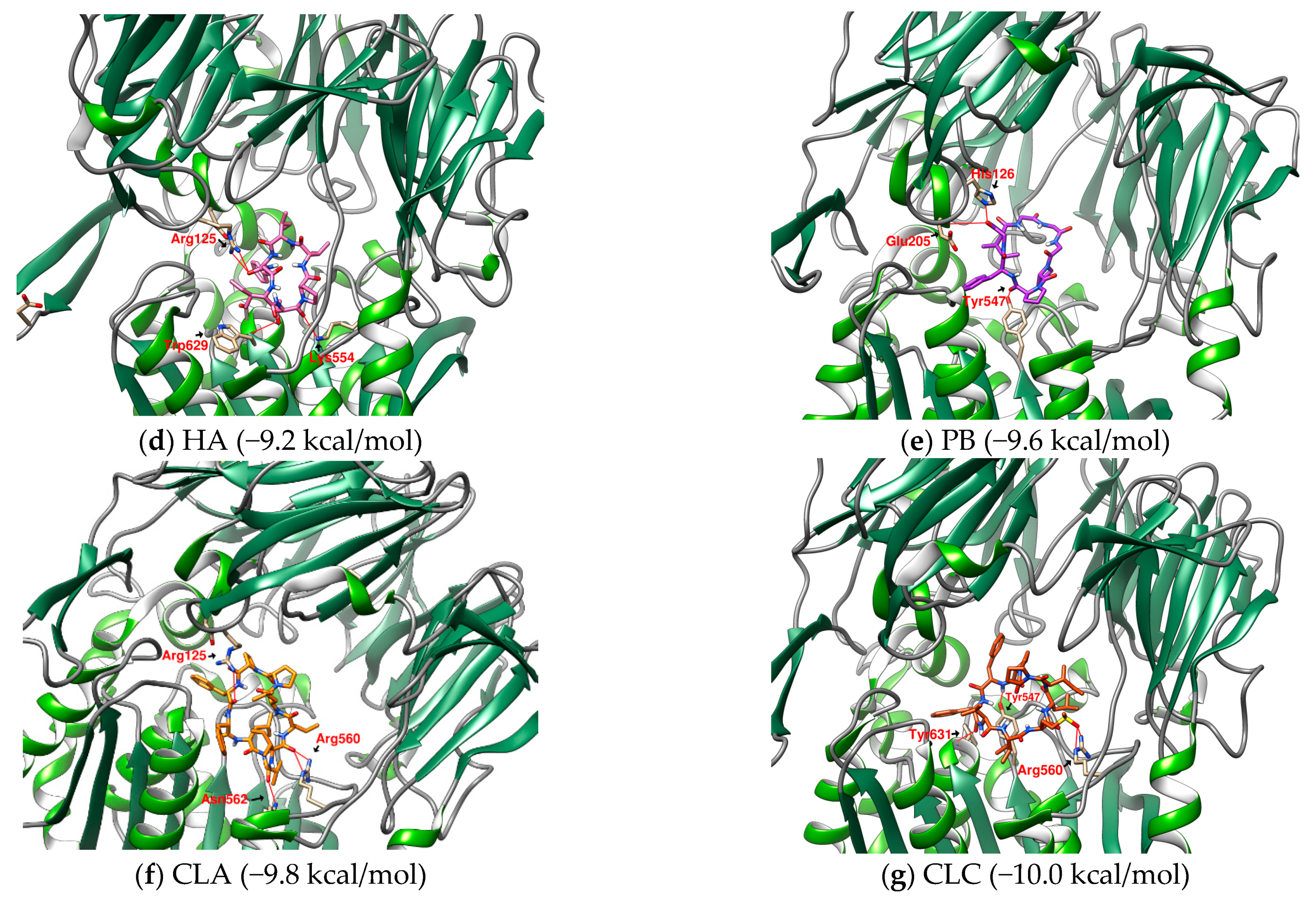
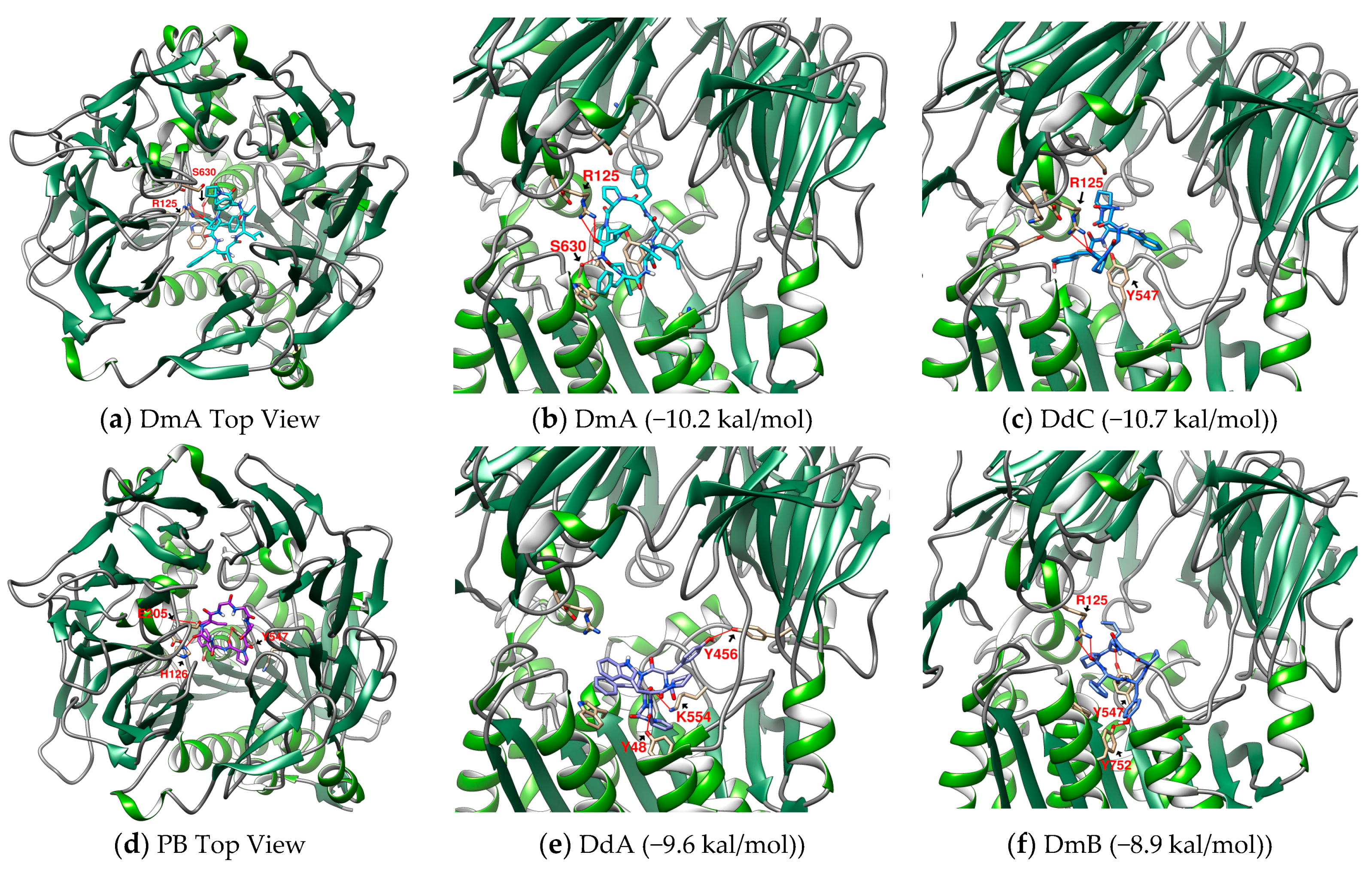
2.1. The Binding Affinity of Three Plant-Derived Cyclopeptides to DPP4 and Their Research Potential
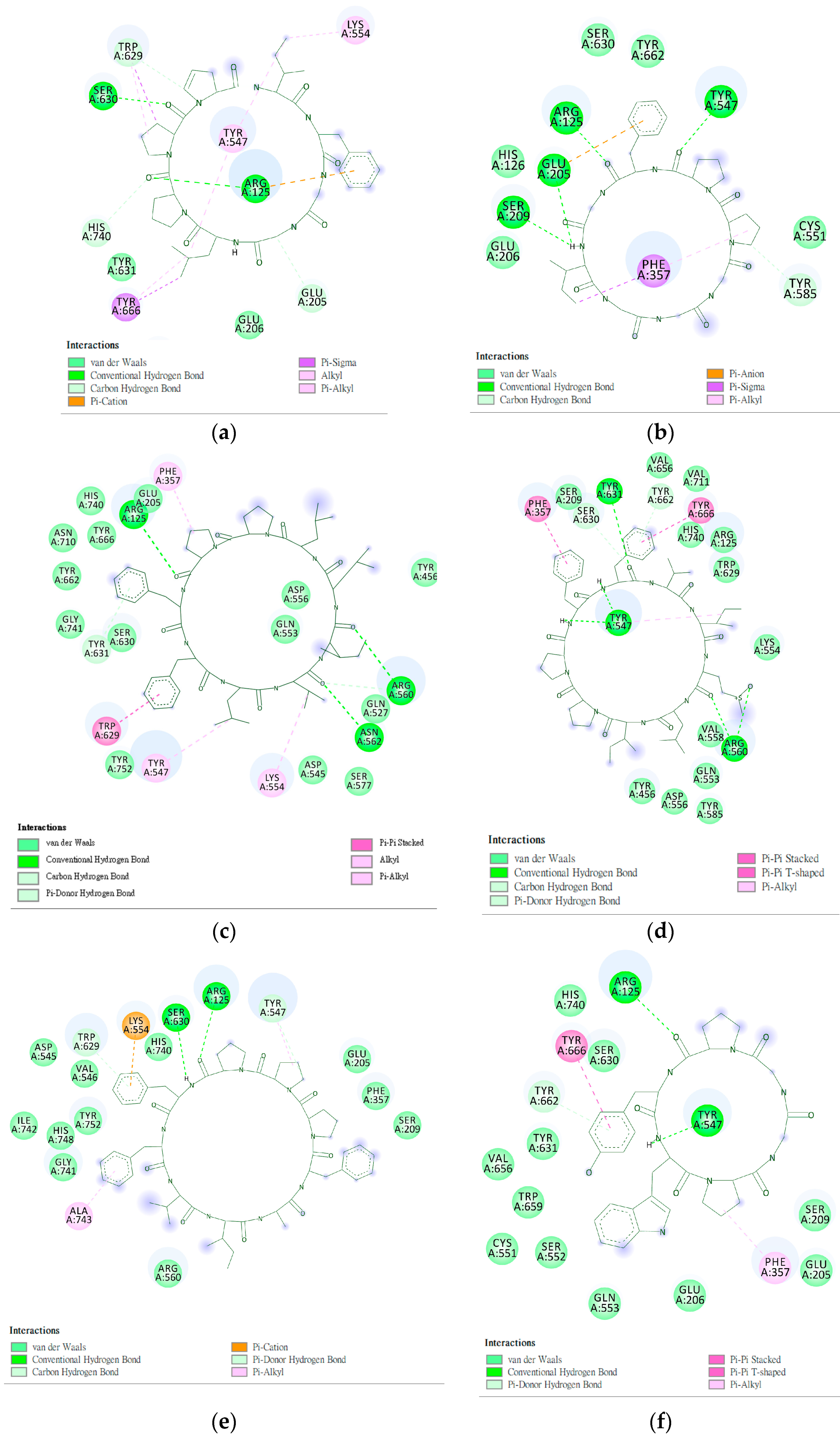
2.2. Analysis of the Configuration and Conformation of Plant Cyclopeptides Docking with DPP4
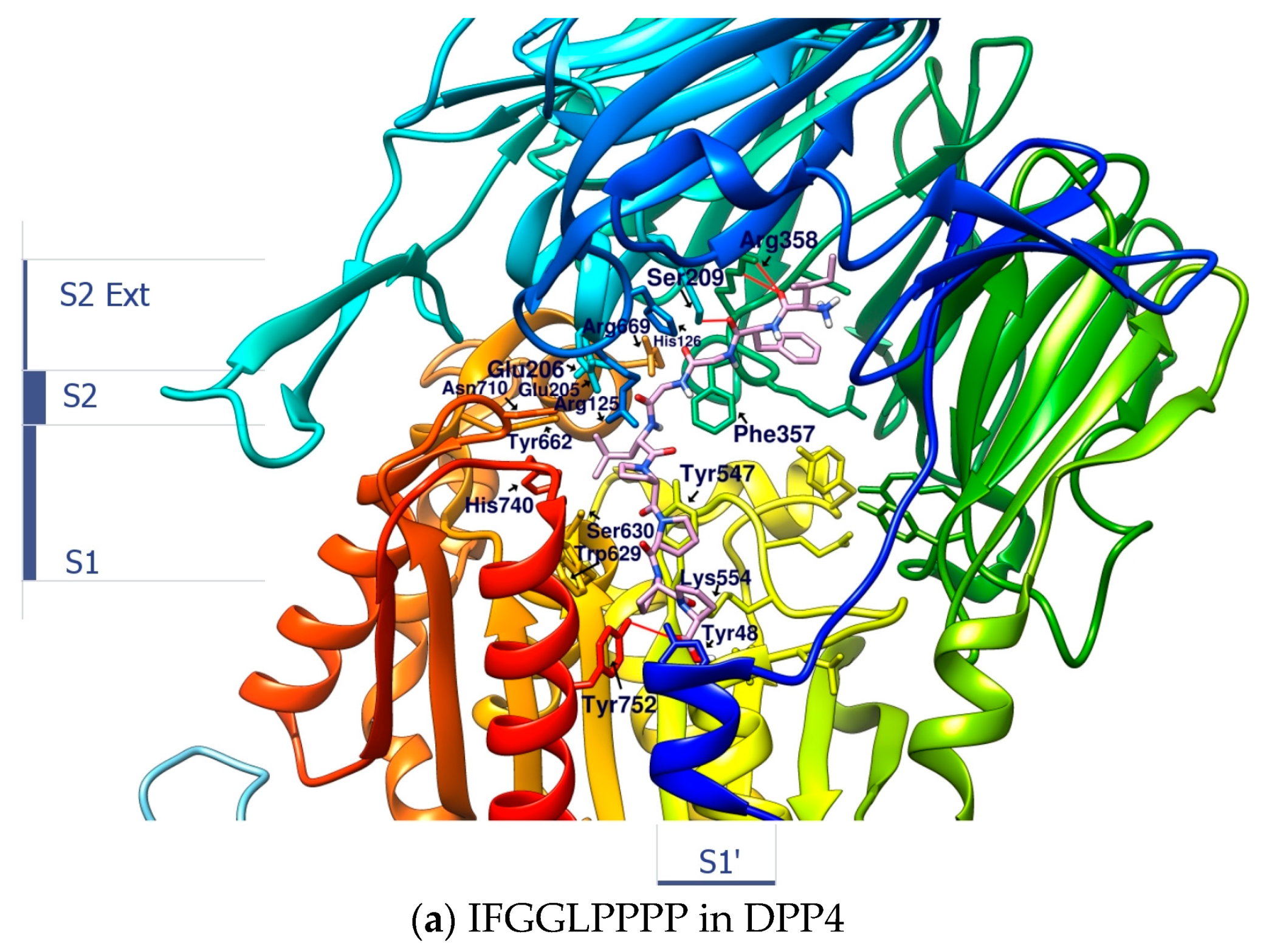
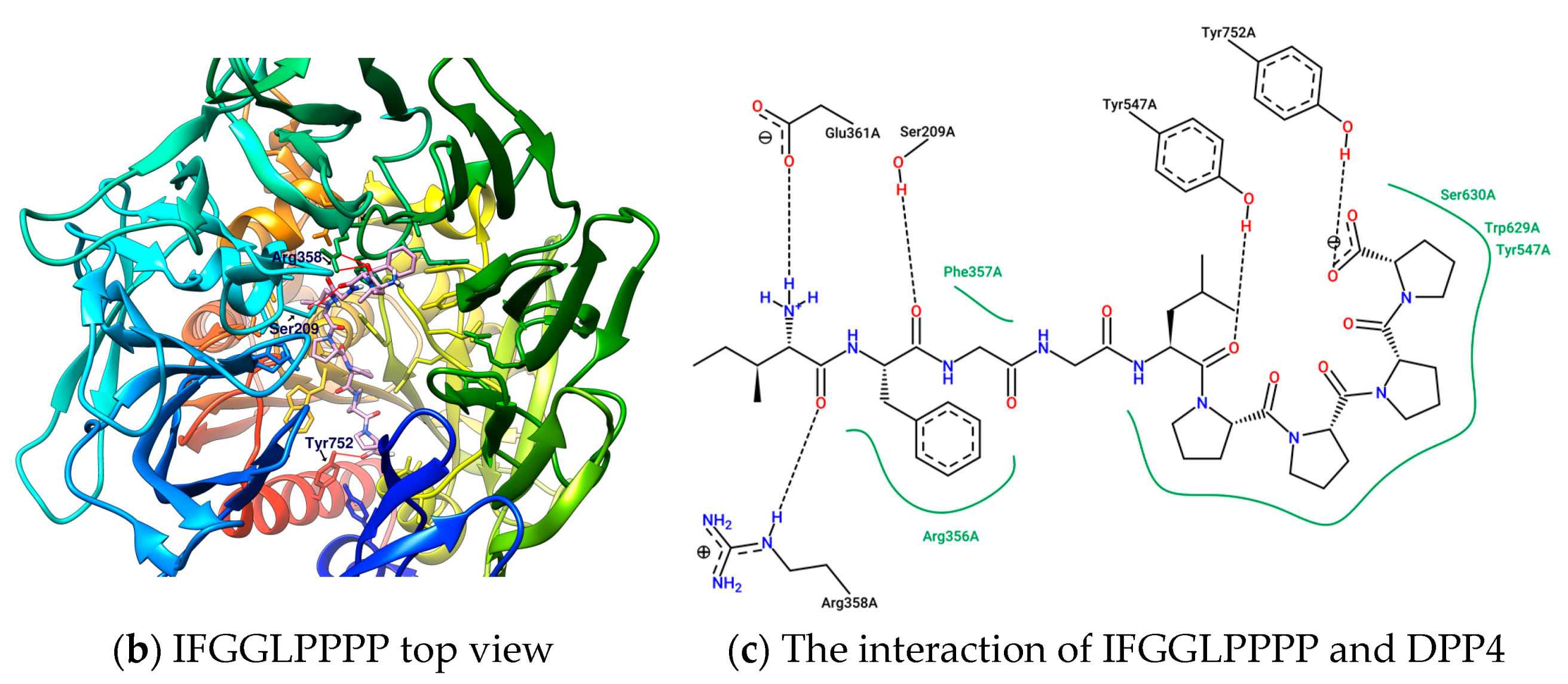
2.3. Linear Peptide ”IFGGLPPPP” as the Reference Coordinate of “IFGGLPPP” (HB Linear Precursor) Derivative
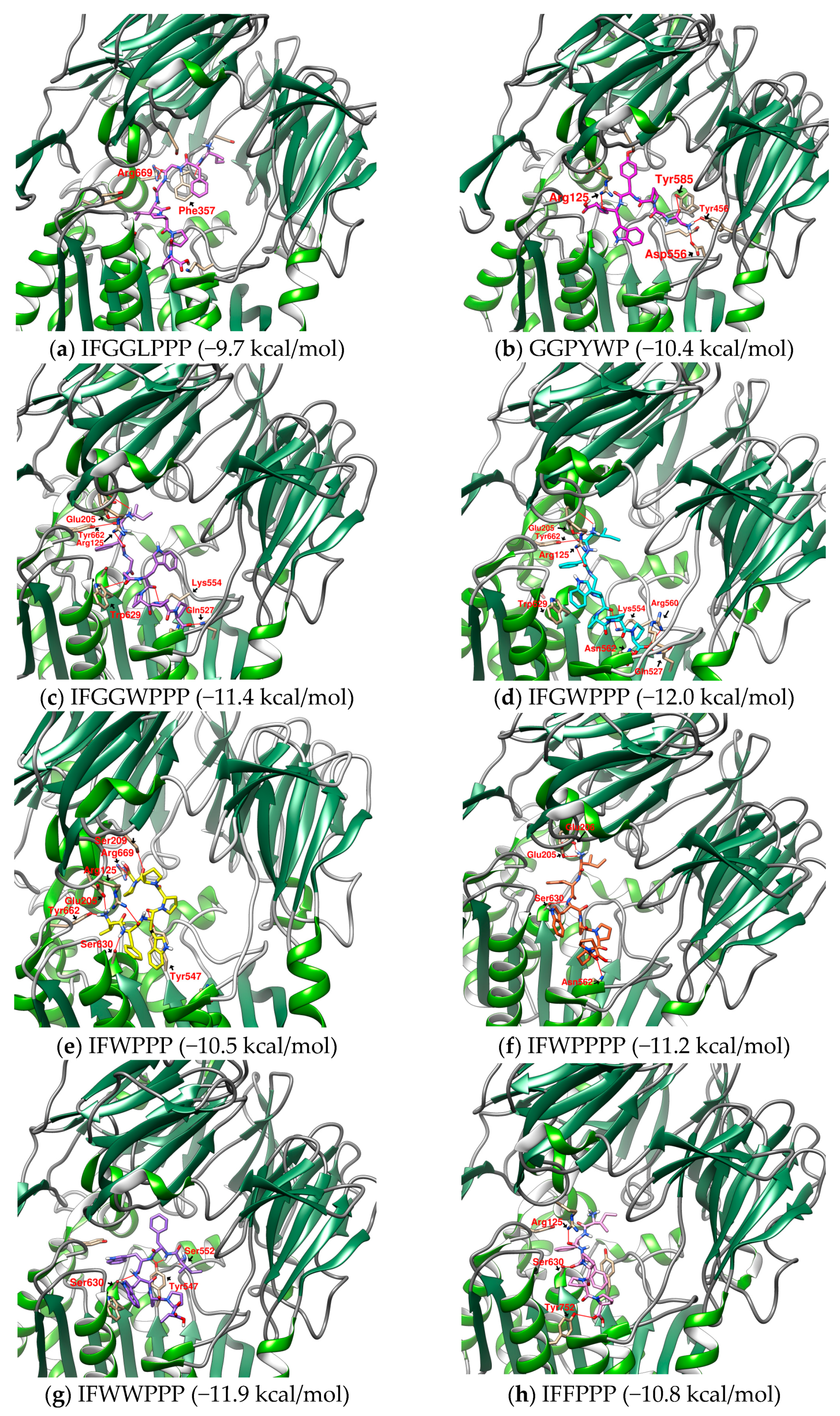
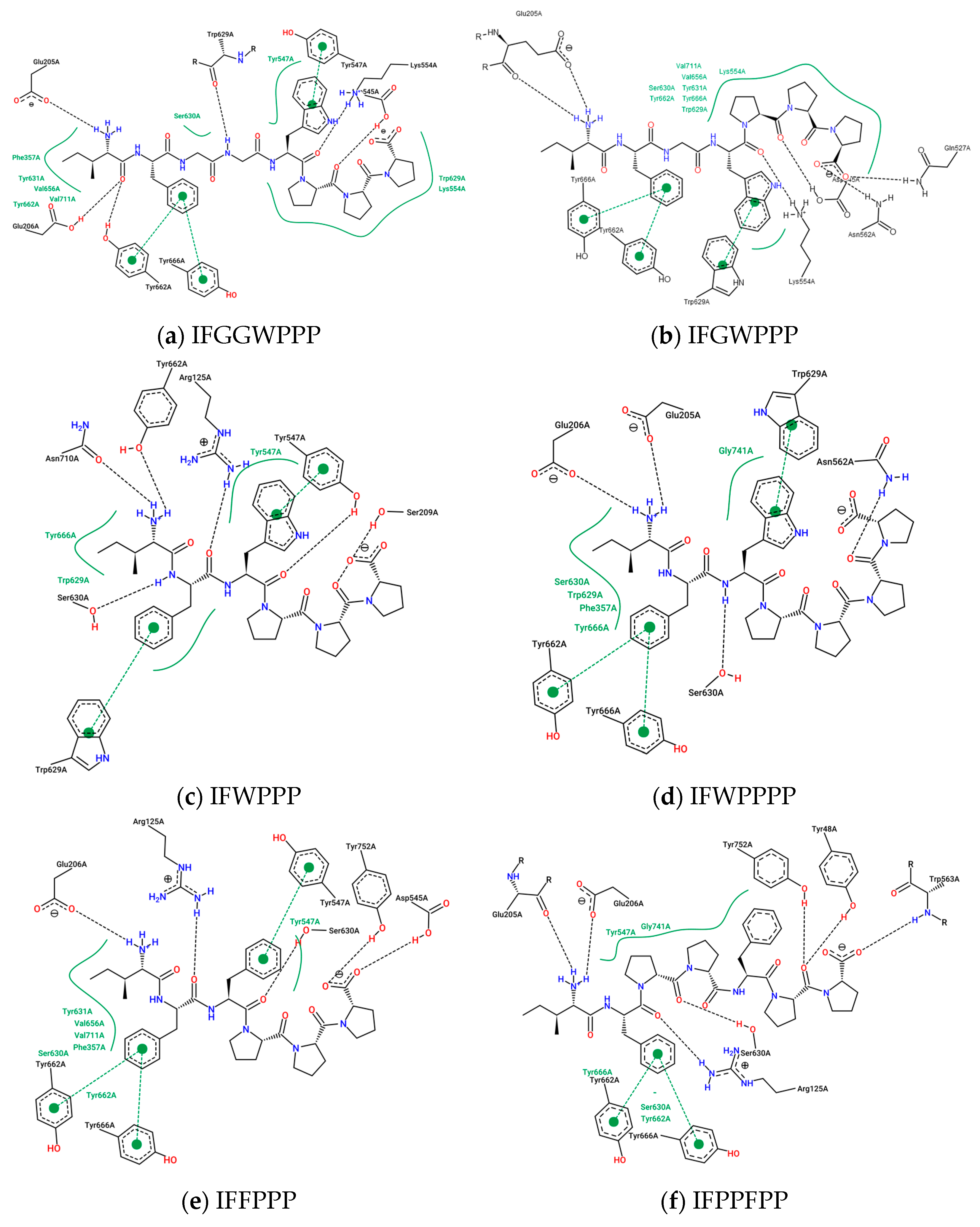
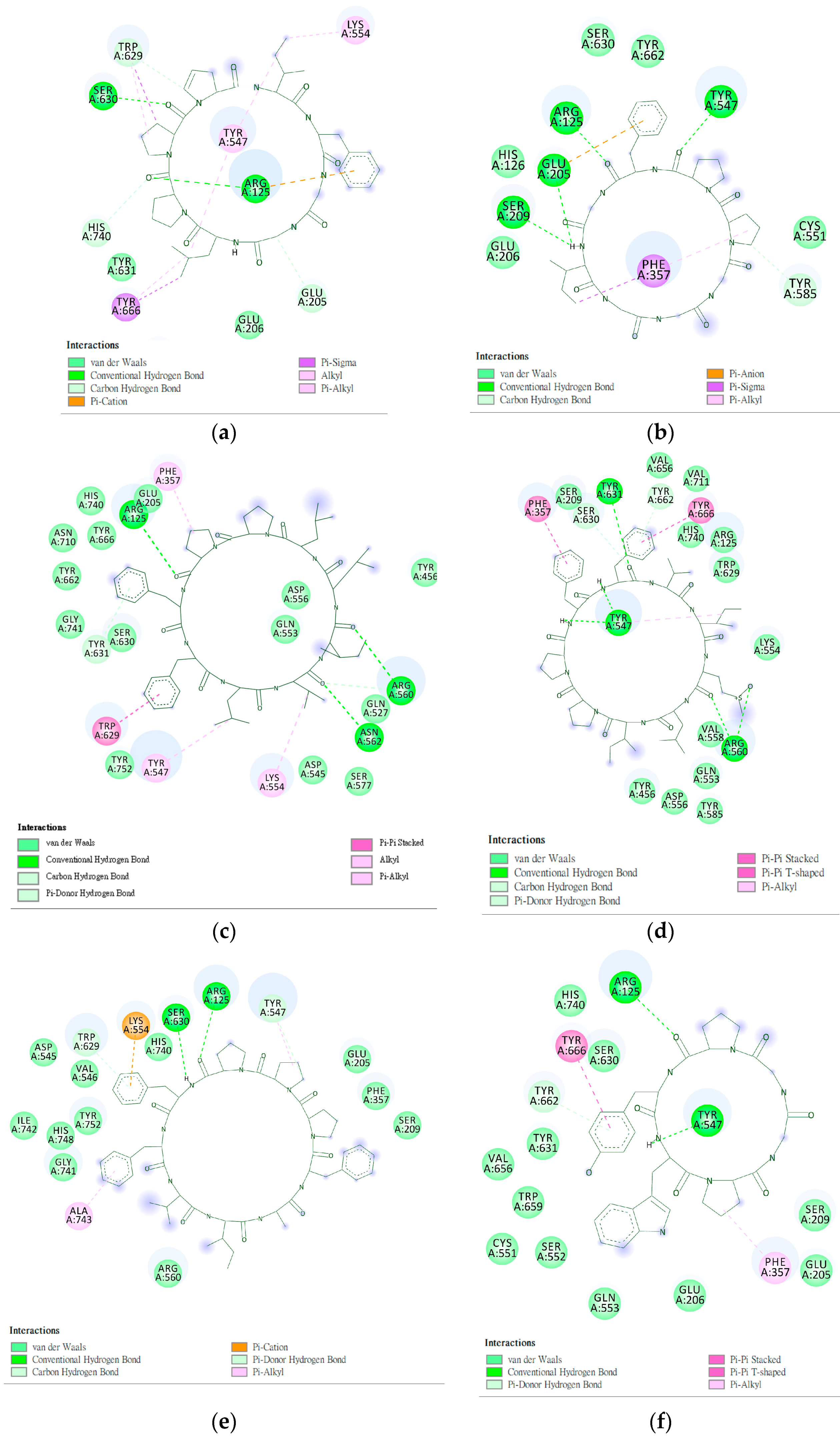
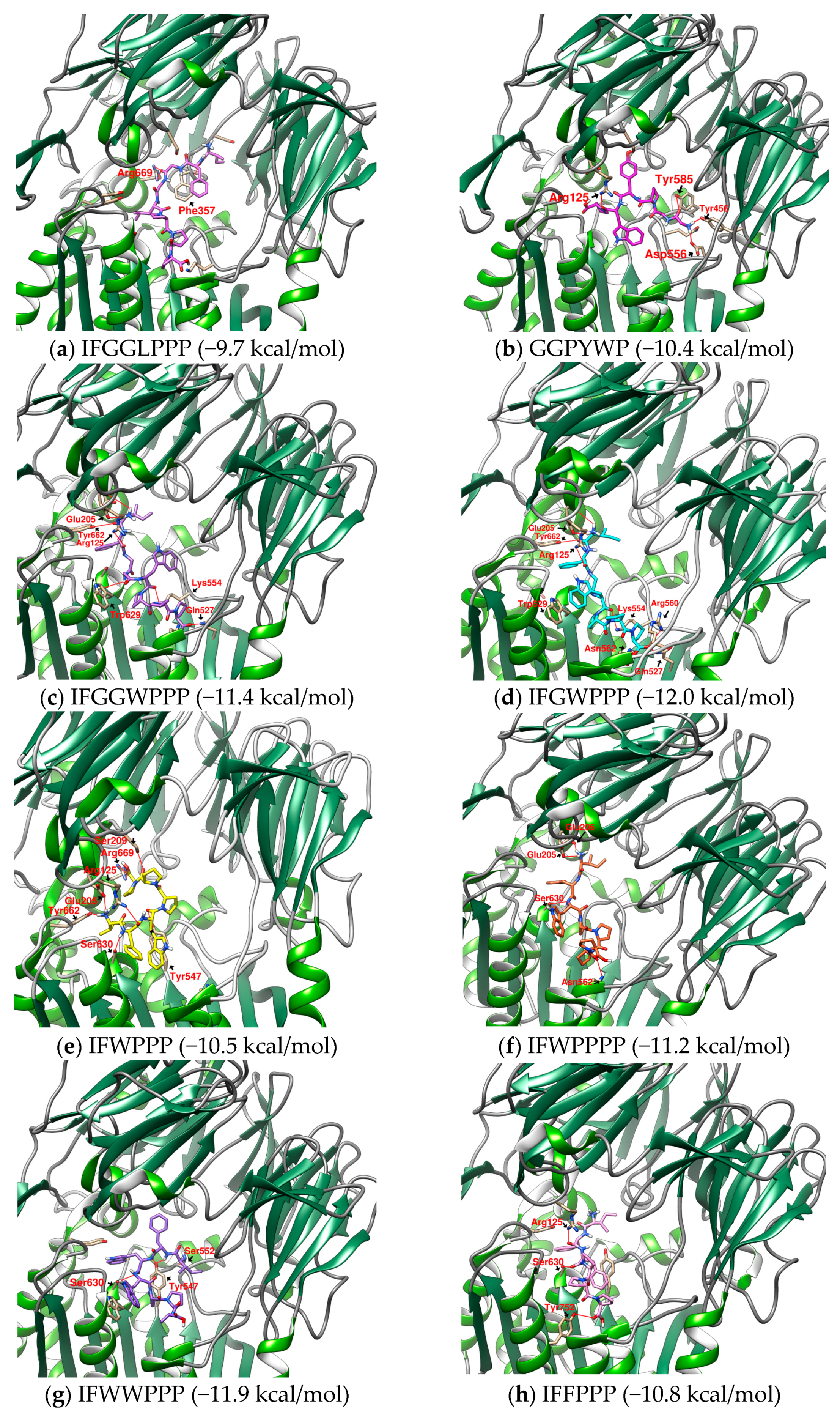
2.4. Design and Analysis of “IFGGLPPP” Derivatives as Potential DPP4 Inhibitors
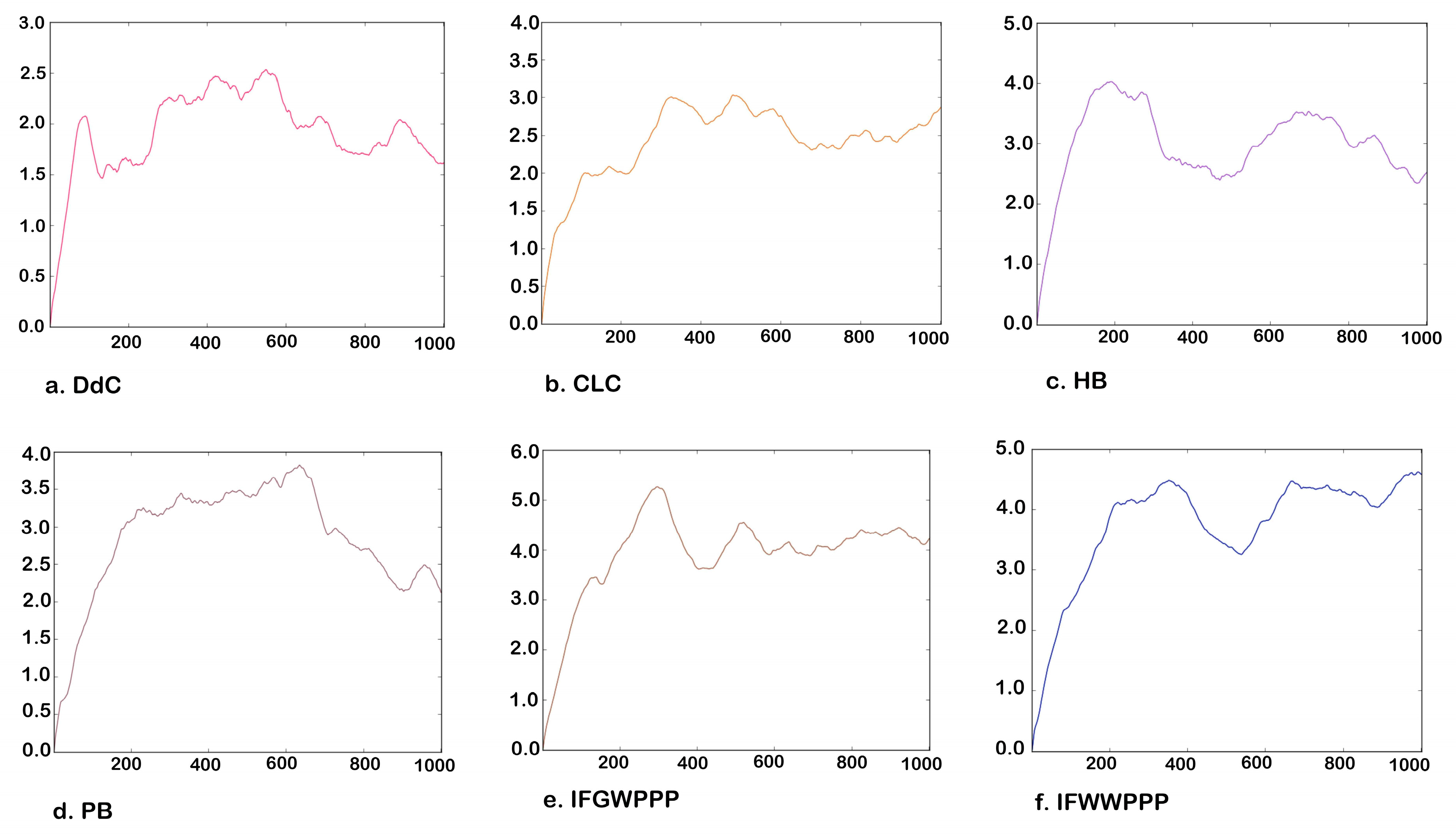
2.5. Molecular Dynamics Simulation of Potential Cyclic and Linear Peptides
3. Materials and Methods
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9(th) edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsalamandris, S.; Antonopoulos, A.S.; Oikonomou, E.; Papamikroulis, G.A.; Vogiatzi, G.; Papaioannou, S.; Deftereos, S.; Tousoulis, D. The Role of Inflammation in Diabetes: Current Concepts and Future Perspectives. Eur. Cardiol. 2019, 14, 50–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, R.M.M.; Chua, Z.J.Y.; Tan, J.C.; Yang, Y.; Liao, Z.; Zhao, Y. From Pre-Diabetes to Diabetes: Diagnosis, Treatments and Translational Research. Medicina 2019, 55, 546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baggio, L.L.; Drucker, D.J. Biology of incretins: GLP-1 and GIP. Gastroenterology 2007, 132, 2131–2157. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Egan, J.M. The role of incretins in glucose homeostasis and diabetes treatment. Pharm. Rev. 2008, 60, 470–512. [Google Scholar] [CrossRef] [Green Version]
- Godinho, R.; Mega, C.; Teixeira-de-Lemos, E.; Carvalho, E.; Teixeira, F.; Fernandes, R.; Reis, F. The Place of Dipeptidyl Peptidase-4 Inhibitors in Type 2 Diabetes Therapeutics: A “Me Too” or “the Special One” Antidiabetic Class? J. Diabetes Res. 2015, 2015, 806979. [Google Scholar] [CrossRef]
- Müller, T.D.; Finan, B.; Bloom, S.; D’Alessio, D.; Drucker, D.J.; Flatt, P.; Fritsche, A.; Gribble, F.; Grill, H.; Habener, J. Glucagon-like peptide 1 (GLP-1). Mol. Metab. 2019, 30, 72–130. [Google Scholar] [CrossRef]
- Deacon, C.F. Physiology and Pharmacology of DPP-4 in Glucose Homeostasis and the Treatment of Type 2 Diabetes. Front. Endocrinol. 2019, 10, 80. [Google Scholar] [CrossRef] [Green Version]
- Nauck, M.A.; Quast, D.R.; Wefers, J.; Meier, J.J. GLP-1 receptor agonists in the treatment of type 2 diabetes–state-of-the-art. Mol. Metab. 2021, 46, 101102. [Google Scholar] [CrossRef]
- Gallwitz, B. Clinical use of DPP-4 inhibitors. Front. Endocrinol. 2019, 10, 389. [Google Scholar] [CrossRef]
- Klemann, C.; Wagner, L.; Stephan, M.; von Hörsten, S. Cut to the chase: A review of CD26/dipeptidyl peptidase-4′s (DPP4) entanglement in the immune system. Clin. Exp. Immunol. 2016, 185, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, L.J. Sitagliptin: A Review in Type 2 Diabetes. Drugs 2017, 77, 209–224. [Google Scholar] [CrossRef] [PubMed]
- Engel, M.; Hoffmann, T.; Wagner, L.; Wermann, M.; Heiser, U.; Kiefersauer, R.; Huber, R.; Bode, W.; Demuth, H.U.; Brandstetter, H. The crystal structure of dipeptidyl peptidase IV (CD26) reveals its functional regulation and enzymatic mechanism. Proc. Natl. Acad. Sci. USA 2003, 100, 5063–5068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Röhrborn, D.; Wronkowitz, N.; Eckel, J. DPP4 in diabetes. Front. Immunol. 2015, 6, 386. [Google Scholar] [CrossRef] [Green Version]
- Mulvihill, E.E.; Drucker, D.J. Pharmacology, physiology, and mechanisms of action of dipeptidyl peptidase-4 inhibitors. Endocr. Rev. 2014, 35, 992–1019. [Google Scholar] [CrossRef] [Green Version]
- Eckhardt, M.; Langkopf, E.; Mark, M.; Tadayyon, M.; Thomas, L.; Nar, H.; Pfrengle, W.; Guth, B.; Lotz, R.; Sieger, P.; et al. 8-(3-(R)-aminopiperidin-1-yl)-7-but-2-ynyl-3-methyl-1-(4-methyl-quinazolin-2-ylmethyl)-3,7-dihydropurine-2,6-dione (BI 1356), a highly potent, selective, long-acting, and orally bioavailable DPP-4 inhibitor for the treatment of type 2 diabetes. J. Med. Chem. 2007, 50, 6450–6453. [Google Scholar] [CrossRef]
- Maezaki, H.; Tawada, M.; Yamashita, T.; Banno, Y.; Miyamoto, Y.; Yamamoto, Y.; Ikedo, K.; Kosaka, T.; Tsubotani, S.; Tani, A.; et al. Design of potent dipeptidyl peptidase IV (DPP-4) inhibitors by employing a strategy to form a salt bridge with Lys554. Bioorg. Med. Chem. Lett. 2017, 27, 3565–3571. [Google Scholar] [CrossRef]
- Yoshida, T.; Akahoshi, F.; Sakashita, H.; Kitajima, H.; Nakamura, M.; Sonda, S.; Takeuchi, M.; Tanaka, Y.; Ueda, N.; Sekiguchi, S. Discovery and preclinical profile of teneligliptin (3-[(2S, 4S)-4-[4-(3-methyl-1-phenyl-1H-pyrazol-5-yl) piperazin-1-yl] pyrrolidin-2-ylcarbonyl] thiazolidine): A highly potent, selective, long-lasting and orally active dipeptidyl peptidase IV inhibitor for the treatment of type 2 diabetes. Bioorganic Med. Chem. 2012, 20, 5705–5719. [Google Scholar] [CrossRef]
- Berger, J.P.; SinhaRoy, R.; Pocai, A.; Kelly, T.M.; Scapin, G.; Gao, Y.D.; Pryor, K.A.D.; Wu, J.K.; Eiermann, G.J.; Xu, S.S.; et al. A comparative study of the binding properties, dipeptidyl peptidase-4 (DPP-4) inhibitory activity and glucose-lowering efficacy of the DPP-4 inhibitors alogliptin, linagliptin, saxagliptin, sitagliptin and vildagliptin in mice. Endocrinol. Diabetes Metab. 2018, 1, e00002. [Google Scholar] [CrossRef] [Green Version]
- Ríos, J.L.; Francini, F.; Schinella, G.R. Natural products for the treatment of type 2 diabetes mellitus. Planta Med. 2015, 81, 975–994. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.S.; Liang, L.F.; Guo, Y.W. Natural products possessing protein tyrosine phosphatase 1B (PTP1B) inhibitory activity found in the last decades. Acta Pharm. Sin. 2012, 33, 1217–1245. [Google Scholar] [CrossRef] [PubMed]
- Hossain, U.; Das, A.K.; Ghosh, S.; Sil, P.C. An overview on the role of bioactive α-glucosidase inhibitors in ameliorating diabetic complications. Food Chem. Toxicol. 2020, 145, 111738. [Google Scholar] [CrossRef] [PubMed]
- Saleem, S.; Jafri, L.; Haq, I.U.; Chang, L.C.; Calderwood, D.; Green, B.D.; Mirza, B. Plants Fagonia cretica L. and Hedera nepalensis K. Koch contain natural compounds with potent dipeptidyl peptidase-4 (DPP-4) inhibitory activity. J. Ethnopharmacol. 2014, 156, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Zhang, Y.; Zhu, J.; Li, B.; Li, Z.; Zhu, W.; Shi, J.; Jia, Q.; Li, Y. Recent progress in natural products as DPP-4 inhibitors. Future Med. Chem. 2015, 7, 1079–1089. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Chen, X.; Chin, Y.; Zheng, J.; Lim, P.E.; Xue, C.; Tang, Q. Identification of curcumin as a potential α-glucosidase and dipeptidyl-peptidase 4 inhibitor: Molecular docking study, in vitro and in vivo biological evaluation. J. Food Biochem. 2021, 46, e13686. [Google Scholar] [CrossRef]
- Lacroix, I.M.; Li-Chan, E.C. Evaluation of the potential of dietary proteins as precursors of dipeptidyl peptidase (DPP)-IV inhibitors by an in silico approach. J. Funct. Foods 2012, 4, 403–422. [Google Scholar] [CrossRef]
- Oseguera-Toledo, M.E.; de Mejía, E.G.; Reynoso-Camacho, R.; Cardador-Martínez, A.; Amaya-Llano, S.L. Proteins and bioactive peptides. Nutrafoods 2014, 13, 147–157. [Google Scholar] [CrossRef]
- Lacroix, I.M.; Li-Chan, E.C. Isolation and characterization of peptides with dipeptidyl peptidase-IV inhibitory activity from pepsin-treated bovine whey proteins. Peptides 2014, 54, 39–48. [Google Scholar] [CrossRef]
- Uenishi, H.; Kabuki, T.; Seto, Y.; Serizawa, A.; Nakajima, H. Isolation and identification of casein-derived dipeptidyl-peptidase 4 (DPP-4)-inhibitory peptide LPQNIPPL from gouda.a-type cheese and its effect on plasma glucose in rats. Int. Dairy J. 2012, 22, 24–30. [Google Scholar] [CrossRef]
- Velarde-Salcedo, A.J.; Barrera-Pacheco, A.; Lara-González, S.; Montero-Morán, G.M.; Díaz-Gois, A.; González de Mejia, E.; Barba de la Rosa, A.P. In vitro inhibition of dipeptidyl peptidase IV by peptides derived from the hydrolysis of amaranth (Amaranthus hypochondriacus L.) proteins. Food Chem. 2013, 136, 758–764. [Google Scholar] [CrossRef]
- Hatanaka, T.; Inoue, Y.; Arima, J.; Kumagai, Y.; Usuki, H.; Kawakami, K.; Kimura, M.; Mukaihara, T. Production of dipeptidyl peptidase IV inhibitory peptides from defatted rice bran. Food Chem. 2012, 134, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.J.; Shakerian, F.; Zhao, J.; Li, S.P. Chemistry, pharmacology and analysis of Pseudostellaria heterophylla: A mini-review. Chin. Med. 2019, 14, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, W.; Zhou, T.; Li, J.; Jiang, W.; Zhang, J.; Xiao, C.; Wei, D.; Yang, C.; Xu, R.; Gong, A.; et al. The Biosynthesis of Heterophyllin B in Pseudostellaria heterophylla From prePhHB-Encoded Precursor. Front. Plant. Sci. 2019, 10, 1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, K.; Zhu, F.; Guo, L.; Zhu, Y.; Li, W.; Xiong, X. Retrospective study on Professor Zhongying Zhou’s experience in Traditional Chinese Medicine treatment on diabetic nephropathy. J. Tradit. Chin. Med. 2013, 33, 262–267. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Pang, W.; Chen, J.; Bai, S.; Zheng, Z.; Wu, X. Hypoglycemic effect of polysaccharides with different molecular weight of Pseudostellaria heterophylla. BMC Complement. Altern. Med. 2013, 13, 267. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Li, Y.W.; Jia, N.; Song, X.M.; Duan, J.L.; Weng, Y.; Guan, Y.; Zhou, D.; Wang, L.; Xi, M.M.; et al. Insulin-secretagogue activity of eleven plant extracts and twelve pure compounds isolated from Aralia taibaiensis. Life Sci. 2013, 92, 131–136. [Google Scholar] [CrossRef]
- Zhou, J.; Tan, N. Application of a new TLC chemical method for detection of cyclopeptides in plants. Chin. Sci. Bull. 2000, 45, 1825–1831. [Google Scholar] [CrossRef]
- Tan, N.-H.; Zhou, J. Plant cyclopeptides. Chem. Rev. 2006, 106, 840–895. [Google Scholar] [CrossRef]
- Arnison, P.G.; Bibb, M.J.; Bierbaum, G.; Bowers, A.A.; Bugni, T.S.; Bulaj, G.; Camarero, J.A.; Campopiano, D.J.; Challis, G.L.; Clardy, J.; et al. Ribosomally synthesized and post-translationally modified peptide natural products: Overview and recommendations for a universal nomenclature. Nat. Prod. Rep. 2013, 30, 108–160. [Google Scholar] [CrossRef]
- Lu, F.; Yang, H.; Lin, S.D.; Zhao, L.; Jiang, C.; Chen, Z.B.; Liu, Y.Y.; Kan, Y.J.; Hu, J.; Pang, W.S. Cyclic Peptide Extracts Derived From Pseudostellaria heterophylla Ameliorates COPD via Regulation of the TLR4/MyD88 Pathway Proteins. Front. Pharm. 2020, 11, 850. [Google Scholar] [CrossRef]
- Zou, H.; Zhu, N.; Li, S. The emerging role of dipeptidyl-peptidase-4 as a therapeutic target in lung disease. Expert Opin. Ther. Targets 2020, 24, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, T.; Chen, W.; Htwe, Y.M.; Tatsumi, K.; Dudek, S.M. DPP4 inhibition by sitagliptin attenuates LPS-induced lung injury in mice. Am. J. Physiol. Lung Cell Mol. Physiol. 2018, 315, L834–L845. [Google Scholar] [CrossRef]
- Fang, W.Y.; Dahiya, R.; Qin, H.L.; Mourya, R.; Maharaj, S. Natural Proline-Rich Cyclopolypeptides from Marine Organisms: Chemistry, Synthetic Methodologies and Biological Status. Mar. Drugs 2016, 14, 194. [Google Scholar] [CrossRef] [Green Version]
- Meli, A.; Tedesco, C.; Della Sala, G.; Schettini, R.; Albericio, F.; De Riccardis, F.; Izzo, I. Phakellistatins: An Underwater Unsolved Puzzle. Mar. Drugs 2017, 15, 78. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Wu, J.; Bao, Y.; Jiang, S.; Wang, Z.; Jin, Y.; Qu, F. The conventional turns rather than irregular γ-/β-turn secondary structures accounting for the antitumor activities of cyclic peptide Phakellistatin 6 analogs. Tetrahedron 2020, 76, 130881. [Google Scholar] [CrossRef]
- Janusz, M.; Zabłocka, A. Colostral proline-rich polypeptides--immunoregulatory properties and prospects of therapeutic use in Alzheimer’s disease. Curr. Alzheimer Res. 2010, 7, 323–333. [Google Scholar] [CrossRef]
- Zorzi, A.; Deyle, K.; Heinis, C. Cyclic peptide therapeutics: Past, present and future. Curr. Opin. Chem. Biol. 2017, 38, 24–29. [Google Scholar] [CrossRef] [Green Version]
- Henninot, A.; Collins, J.C.; Nuss, J.M. The Current State of Peptide Drug Discovery: Back to the Future? J. Med. Chem. 2018, 61, 1382–1414. [Google Scholar] [CrossRef]
- Shim, Y.Y.; Song, Z.; Jadhav, P.D.; Reaney, M.J. Orbitides from flaxseed (Linum usitatissimum L.): A comprehensive review. Trends Food Sci. Technol. 2019, 93, 197–211. [Google Scholar] [CrossRef]
- Mani, U.V.; Mani, I.; Biswas, M.; Kumar, S.N. An open-label study on the effect of flax seed powder (Linum usitatissimum) supplementation in the management of diabetes mellitus. J. Diet. Suppl. 2011, 8, 257–265. [Google Scholar] [CrossRef]
- Pan, A.; Sun, J.; Chen, Y.; Ye, X.; Li, H.; Yu, Z.; Wang, Y.; Gu, W.; Zhang, X.; Chen, X.; et al. Effects of a flaxseed-derived lignan supplement in type 2 diabetic patients: A randomized, double-blind, cross-over trial. PLoS ONE 2007, 2, e1148. [Google Scholar] [CrossRef]
- Behsaz, B.; Mohimani, H.; Gurevich, A.; Prjibelski, A.; Fisher, M.; Vargas, F.; Smarr, L.; Dorrestein, P.C.; Mylne, J.S.; Pevzner, P.A. De Novo Peptide Sequencing Reveals Many Cyclopeptides in the Human Gut and Other Environments. Cell Syst. 2020, 10, 99–108.e105. [Google Scholar] [CrossRef] [Green Version]
- Nono, N.R.; Nzowa, K.L.; Barboni, L.; Tapondjou, A.L. Drymaria cordata (Linn.) Willd (Caryophyllaceae): Ethnobotany, Pharmacology and Phytochemistry. Adv. Biol. Chem. 2014, 2014. [Google Scholar] [CrossRef] [Green Version]
- Barua, C.C.; Roy, J.D.; Buragohain, B.; Barua, A.G.; Borah, P.; Lahkar, M. Analgesic and anti-nociceptive activity of hydroethanolic extract of Drymaria cordata Willd. Indian J. Pharm. 2011, 43, 121–125. [Google Scholar] [CrossRef]
- Ding, Z.; Zhou, J.; Tan, N.; Teng, R. Two new cyclic peptides from Drymaria diandra. Planta Med. 2000, 66, 386–388. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Saha, K.; Bhattacharya, S.; Giri, S.; Pal, M.; Saha, B. Studies on antitussive activity of Drymaria cordata Willd. (Caryophyllaceae). J. Ethnopharmacol. 1997, 56, 77–80. [Google Scholar] [CrossRef]
- Hsieh, P.W.; Chang, F.R.; Wu, C.C.; Wu, K.Y.; Li, C.M.; Wang, W.Y.; Gu, L.C.; Wu, Y.C. Selective Inhibition of Collagen-Induced Platelet Aggregation by a Cyclic Peptide from Drymaria diandra. Helv. Chim. Acta 2004, 87, 57–66. [Google Scholar] [CrossRef]
- Hsieh, P.W.; Chang, F.R.; Lee, K.H.; Hwang, T.L.; Chang, S.M.; Wu, Y.C. A new anti-HIV alkaloid, drymaritin, and a new C-glycoside flavonoid, diandraflavone, from Drymaria diandra. J. Nat. Prod. 2004, 67, 1175–1177. [Google Scholar] [CrossRef]
- Patra, S.; Bhattacharya, S.; Bala, A.; Haldar, P.K. Antidiabetic effect of Drymaria cordata leaf against streptozotocin-nicotinamide-induced diabetic albino rats. J. Adv. Pharm. Technol. Res. 2020, 11, 44–52. [Google Scholar] [CrossRef]
- Zhang, Z.; Wallace, M.B.; Feng, J.; Stafford, J.A.; Skene, R.J.; Shi, L.; Lee, B.; Aertgeerts, K.; Jennings, A.; Xu, R.; et al. Design and synthesis of pyrimidinone and pyrimidinedione inhibitors of dipeptidyl peptidase IV. J. Med. Chem. 2011, 54, 510–524. [Google Scholar] [CrossRef]
- Bower, A.M.; Real Hernandez, L.M.; Berhow, M.A.; de Mejia, E.G. Bioactive compounds from culinary herbs inhibit a molecular target for type 2 diabetes management, dipeptidyl peptidase IV. J. Agric. Food Chem. 2014, 62, 6147–6158. [Google Scholar] [CrossRef] [PubMed]
- Morita, H.; Shishido, A.; Matsumoto, T.; Takeya, K.; Itokawa, H.; Hirano, T.; Oka, K. A new immunosuppressive cyclic nonapeptide, cyclolinopeptide B from Linum usitatissimum. Bioorganic Med. Chem. Lett. 1997, 7, 1269–1272. [Google Scholar] [CrossRef]
- Chittepu, V.; Kalhotra, P.; Osorio-Gallardo, T.; Jiménez-Martínez, C.; Torre, R.R.R.; Gallardo-Velazquez, T.; Osorio-Revilla, G. New Molecular Insights into the Inhibition of Dipeptidyl Peptidase-4 by Natural Cyclic Peptide Oxytocin. Molecules 2019, 24, 3887. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Damjanovic, J.; Miao, J.; Lin, Y.S. Cyclic peptides: Backbone rigidification and capability of mimicking motifs at protein-protein interfaces. Phys. Chem. Chem. Phys. 2021, 23, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Poojary, B.; Kumar, K.H.; Belagali, S.L. Synthesis and biological evaluation of pseudostellarin B. Farmaco 2001, 56, 331–334. [Google Scholar] [CrossRef]
- Dahiya, R.; Singh, S.; Varghese Gupta, S.; Sutariya, V.B.; Bhatia, D.; Mourya, R.; Chennupati, S.V.; Sharma, A. First Total Synthesis and Pharmacological Potential of a Plant Based Hexacyclopeptide. Iran. J. Pharm. Res. 2019, 18, 938–947. [Google Scholar] [CrossRef]
- Yang, C.; You, L.; Yin, X.; Liu, Y.; Leng, X.; Wang, W.; Sai, N.; Ni, J. Heterophyllin B Ameliorates Lipopolysaccharide-Induced Inflammation and Oxidative Stress in RAW 264.7 Macrophages by Suppressing the PI3K/Akt Pathways. Molecules 2018, 23, 717. [Google Scholar] [CrossRef] [Green Version]
- Zhuge, F.; Ni, Y.; Nagashimada, M.; Nagata, N.; Xu, L.; Mukaida, N.; Kaneko, S.; Ota, T. DPP-4 Inhibition by Linagliptin Attenuates Obesity-Related Inflammation and Insulin Resistance by Regulating M1/M2 Macrophage Polarization. Diabetes 2016, 65, 2966–2979. [Google Scholar] [CrossRef] [Green Version]
- Arulmozhiraja, S.; Matsuo, N.; Ishitsubo, E.; Okazaki, S.; Shimano, H.; Tokiwa, H. Comparative Binding Analysis of Dipeptidyl Peptidase IV (DPP-4) with Antidiabetic Drugs—An Ab Initio Fragment Molecular Orbital Study. PLoS ONE 2016, 11, e0166275. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.; Dey, S. Molecular Docking Studies of a Cyclic Octapeptide-Cyclosaplin from Sandalwood. Biomolecules 2019, 9, 740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Y.; Wang, M.; Sun, C.; Peng, C.; Zhang, Y.; Li, X. Tunicyclin L, a cyclic peptide from Psammosilene tunicoides: Isolation, characterization, conformational studies and biological activity. Fitoterapia 2020, 145, 104628. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhao, S.; Song, L.; Pu, Y.; Wang, Q.; Zeng, G.; Liu, X.; Bai, M.; Li, S.; Gao, F.; et al. Natural cyclopeptide RA-V inhibits the NF-κB signaling pathway by targeting TAK1. Cell Death Dis. 2018, 9, 715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapovalov, M.V.; Dunbrack, R.L., Jr. A smoothed backbone-dependent rotamer library for proteins derived from adaptive kernel density estimates and regressions. Structure 2011, 19, 844–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Wang, W.; Kollman, P.A.; Case, D.A. Automatic atom type and bond type perception in molecular mechanical calculations. J. Mol. Graph. Model. 2006, 25, 247–260. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N. | Abbr. | Compound | M.W. (g/mol) | Structure Name | BA (kcal/mol) |
|---|---|---|---|---|---|
| 1 | HA | Heterophyllin A | 727.9 | (cyclo)-PVIFGIT-(cyclo) [37] | −9.2 |
| 2 | HB | Heterophyllin B | 778.9 | (cyclo)-GGLPPPIF-(cyclo) | −10.4 |
| 3 | HC | Heterophyllin C | 703.9 | (cyclo)-GPIIPIL-(cyclo) | −8.8 |
| 4 | HJ | Heterophyllin J | 487.5 | (cyclo)-AGPVY-(cyclo) | −8.8 |
| 5 | PA | Pseudostellarin A | 501.6 | (cyclo)-AGPYL-(cyclo) | −8.4 |
| 6 | PB | Pseudostellarin B | 682.8 | (cyclo)-GGGPPFGI-(cyclo) | −9.6 |
| 7 | PC | Pseudostellarin C | 813 | (cyclo)-GTLPSPFL-(cyclo) | −8.5 |
| 8 | PD | Pseudostellarin D | 713.9 | (cyclo)-GPLILGY-(cyclo) | −9.6 |
| 9 | PE | Pseudostellarin E | 878.1 | (cyclo)-GPPLGPVIF-(cyclo) | −9.3 |
| 10 | PH | Pseudostellarin H | 861 | (cyclo)-GTPTPLFF-(cyclo) | −9.4 |
| N. | Abbr. | Compound | M.W. (g/mol) | Structure Name | BA (kcal/mol) |
|---|---|---|---|---|---|
| 1 | CLA | Cyclolinopeptide A | 1040.3 | (cyclo)-ILLPPFFLV-(cyclo) | −9.8 |
| 2 | CLB | Cyclolinopeptide B | 1058.4 | (cyclo)-IMLIPPFFV-(cyclo) | −9.8 |
| 3 | CLC | Cyclolinopeptide C | 1074.4 | (cyclo)-IM(O)LIPPFFV-(cyclo) | −10.0 |
| 4 | CLD | Cyclolinopeptide D | 1064.3 | (cyclo)-LLPFFWIM(O)-(cyclo) | −8.7 |
| 5 | CLE | Cyclolinopeptide E | 977.3 | (cyclo)-IM(O)LVFPLF-(cyclo) | −9.1 |
| 6 | CLF | Cyclolinopeptide F | 1084.4 | (cyclo)-LM(O)PFFWVM(O)-(cyclo) | −9.3 |
| 7 | CLG | Cyclolinopeptide G | 1098.4 | (cyclo)-LM(O)PFFWIM(O)-(cyclo) | −9.3 |
| 8 | CLH | Cyclolinopeptide H | 1082.4 | (cyclo)-LMPFFWIM(O)-(cyclo) | −9.1 |
| 9 | CLI | Cyclolinopeptide I | 1068.4 | (cyclo)-LM(O)PFFWVM-(cyclo) | −9.2 |
| 10 | CLJ | Cyclolinopeptide J | 993.3 | (cyclo)-IM(O2)LVFPLF-(cyclo) | −9.1 |
| N. | Abbr. | Compound | M.W. (g/mol) | Structure Name | BA (kcal/mol) |
|---|---|---|---|---|---|
| 1 | DmA | Drymarin A | 1016.2 | (cyclo)-AFPPPFFVI-(cyclo) | −10.2 |
| 2 | DmB | Drymarin B | 674.8 | (cyclo)-GLPFYP-(cyclo) | −8.9 |
| 3 | DdA | Diandrine A | 747.8 | (cyclo)-GPWPYF-(cyclo) | −9.6 |
| 4 | DdB | Diandrine B | 838 | (cyclo)-GPLPLWSS-(cyclo) | −8.9 |
| 5 | DdC | Diandrine C | 657.7 | (cyclo)-GGPYWP-(cyclo) | −10.7 |
| N | Sequence | S2 Ext | S2 and S1 Pocket | S1′ and Surrounding | Periphery | BA (kcal/mol) | MV (g/mol) |
|---|---|---|---|---|---|---|---|
| 1 | IFGGLPPP | F357 F357π | R669 | Y547 | - | −9.7 | 796.9733 |
| 2 | IFGGLPPPP | S209, R358 E361 | - | Y547 | Y752 | −10.3 | 894.0913 |
| 3 | GGLPPPIF | - | E205, S630, W629π | Y547, K554 | - | −8.9 | 796.9733 |
| 4 | IP | - | Y662 | - | - | −6.6 | 228.2914 |
| 5 | IFP | - | S630, Y662π, Y666π | - | - | −8.4 | 375.4680 |
| 6 | IFPP | - | W629, S630, Y662π, Y666π | Y547 | - | −9.5 | 472.5903 |
| 7 | IFPPP | - | E205, Y662π, Y666π | Y547 | - | −10.0 | 569.7083 |
| 8 | IFPPPP | - | R125, S630, Y662π, Y666π | D545 | Y752 | −10.4 | 666.8263 |
| 9 | IFFPPP | - | R125, E206, S630, Y666π | D545, Y547π | Y752 | −10.8 | 716.8863 |
| 10 | IWWPP | F357π | E206, S630, W629 | D545, Y547, C551 | - | −11.1 | 697.8423 |
| 11 | IFWPP | - | R125, Y666 | Y547, Y547π | - | −9.8 | 658.8053 |
| 12 | IFWPPP | S209 | R125, S630, N710, Y662, W629π | Y547, Y547π | - | −10.5 | 755.9233 |
| 13 | IFWPPPP | - | E205, E206, S630, W629π, Y662π, Y666π | N562 | - | −11.2 | 853.0413 |
| 14 | IFWWPPP | F357π | S630, Y662π, W629π | Y547, C551 | −11.9 | 941.4800 | |
| 15 | IFGPPPP | - | R125, E205, Y662, Y662π, Y666π | D545, K554 | - | −10.7 | 723.8783 |
| 16 | IFGWPPP | - | E205, Y662π, Y666π, W629π | Q527, K554, N562 | - | −12.0 | 812.9753 |
| 17 | IFGGWPPP | - | E206, Y662π, Y666π | D545, Y547, K554 | - | −11.4 | 870.0273 |
| 18 | IFYWPPPP | - | R125, W629π | V546, K554 | Y752 Y48 | −11.3 | 1016.219 |
| 19 | IFPPFPP | - | R125, E205, E206 S630, Y662π, Y666π | W563 | Y752 Y48 | −10.6 | 814.0043 |
| 20 | IFYGPPP | - | E205, E206, S630 Y662, Y662π, Y666π | V546, K554 Y547π, K554 | Y752 | −10.4 | 789.9383 |
| 21 | IFIFPPP | - | R125, E205, S630 Y662π, Y666π | - | Y752 | −10.0 | 716.8863 |
| 22 | GGPYWP | F357π | E206 | Y456, Y547π D556, Y585 | - | −10.4 | 675.7483 |
| DdC | CLC | HB | PB | IFGWPPP | IFWWPPP | |
|---|---|---|---|---|---|---|
| Number of atoms | 48 | 76 | 56 | 49 | 59 | 69 |
| Average RMSD over 1000 frames | 1.611 | 2.43 | 2.514 | 2.118 | 4.214 | 4.582 |
| Average Potential energy | 747.162 | 987.740 | 931.417 | 792.090 | 955.261 | 938.587 |
| Average Kinetic energy | 336.549 | 628.665 | 472.127 | 342.365 | 430.523 | 486.235 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, H.-J.; Tzen, J.T.C. The Potential Role of Cyclopeptides from Pseudostellaria heterophylla, Linum usitatissimum and Drymaria diandra, and Peptides Derived from Heterophyllin B as Dipeptidyl Peptidase IV Inhibitors for the Treatment of Type 2 Diabetes: An In Silico Study. Metabolites 2022, 12, 387. https://doi.org/10.3390/metabo12050387
Liao H-J, Tzen JTC. The Potential Role of Cyclopeptides from Pseudostellaria heterophylla, Linum usitatissimum and Drymaria diandra, and Peptides Derived from Heterophyllin B as Dipeptidyl Peptidase IV Inhibitors for the Treatment of Type 2 Diabetes: An In Silico Study. Metabolites. 2022; 12(5):387. https://doi.org/10.3390/metabo12050387
Chicago/Turabian StyleLiao, Hui-Jun, and Jason T. C. Tzen. 2022. "The Potential Role of Cyclopeptides from Pseudostellaria heterophylla, Linum usitatissimum and Drymaria diandra, and Peptides Derived from Heterophyllin B as Dipeptidyl Peptidase IV Inhibitors for the Treatment of Type 2 Diabetes: An In Silico Study" Metabolites 12, no. 5: 387. https://doi.org/10.3390/metabo12050387
APA StyleLiao, H.-J., & Tzen, J. T. C. (2022). The Potential Role of Cyclopeptides from Pseudostellaria heterophylla, Linum usitatissimum and Drymaria diandra, and Peptides Derived from Heterophyllin B as Dipeptidyl Peptidase IV Inhibitors for the Treatment of Type 2 Diabetes: An In Silico Study. Metabolites, 12(5), 387. https://doi.org/10.3390/metabo12050387