
Amino Acid Metabolism in Apicomplexan Parasites



{kind=link}
{kind=link}
Abstract
1. Introduction
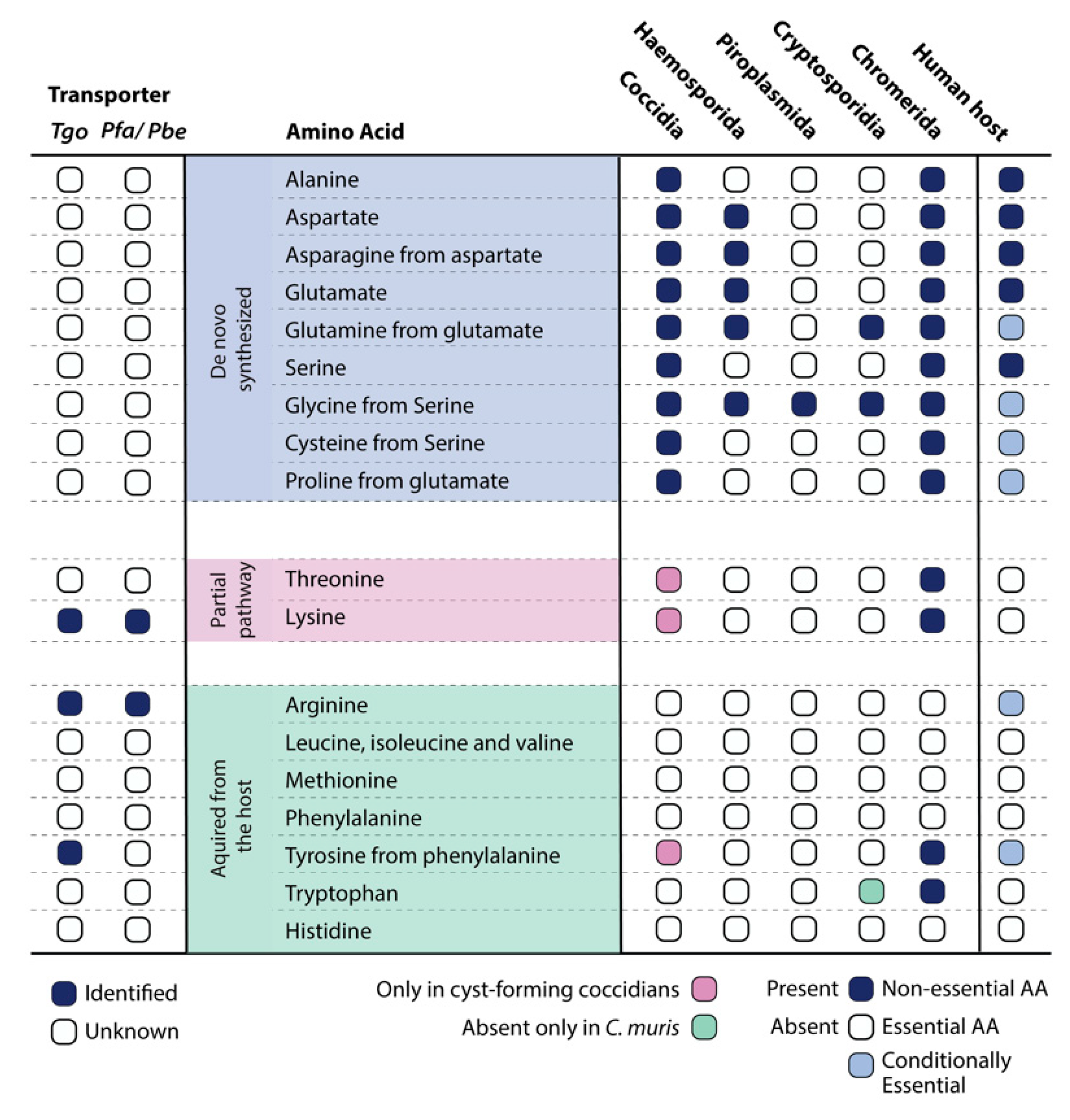
2. Amino Acids That Can Be Synthesized De Novo
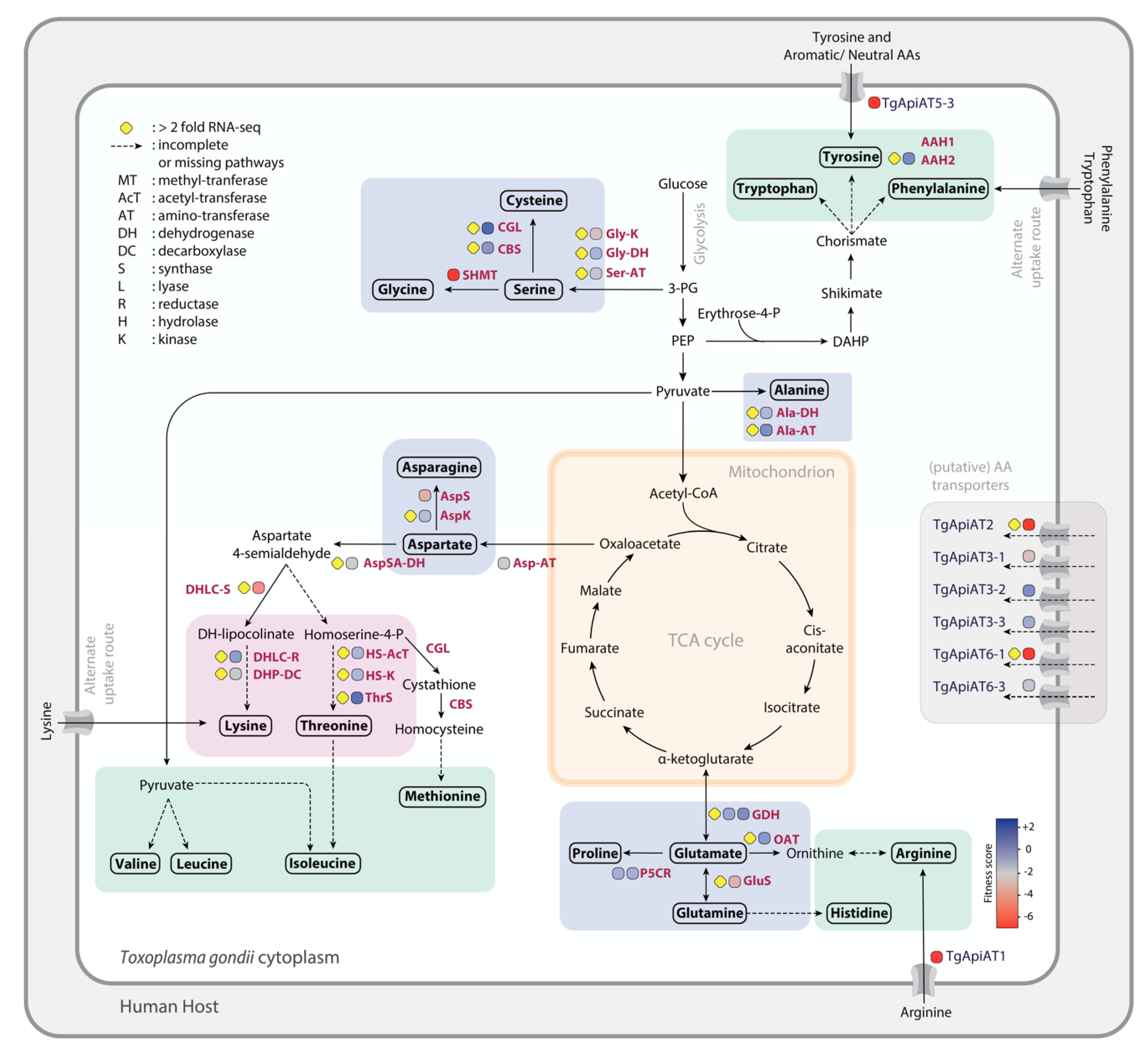
2.1. Alanine
2.2. Asparagine and Aspartate
2.3. Glutamate and Glutamine
2.4. Glycine, Serine, and Cysteine
2.5. Proline
3. Essential Amino Acids with an Incomplete De Novo Synthesis Pathway
3.1. Threonine
3.2. Lysine
4. Essential Amino Acids That Must Be Taken Up
4.1. Arginine
4.2. Leucine, Isoleucine, and Valine
4.3. Methionine
4.4. Phenylalanine, Tryptophan, and Tyrosine
4.5. Histidine
5. Non-Proteogenic Amino Acids
6. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Montoya, J.G.; Liesenfeld, O. Toxoplasmosis. Lancet 2004, 363, 1965–1976. [Google Scholar] [CrossRef]
- Djurković-Djaković, O.; Dupouy-Camet, J.; Van der Giessen, J.; Dubey, J.P. Toxoplasmosis: Overview from a One Health perspective. Food Waterborne Parasitol. 2019, 15, e00054. [Google Scholar] [CrossRef]
- Checkley, W.; White, A.C.; Jaganath, D.; Arrowood, M.J.; Chalmers, R.M.; Chen, X.M.; Fayer, R.; Griffiths, J.K.; Guerrant, R.L.; Hedstrom, L.; et al. A review of the global burden, novel diagnostics, therapeutics, and vaccine targets for cryptosporidium. Lancet Infect. Dis. 2015, 15, 85–94. [Google Scholar] [CrossRef]
- Battle, K.E.; Gething, P.W.; Elyazar, I.R.F.; Moyes, C.L.; Sinka, M.E.; Howes, R.E.; Guerra, C.A.; Price, R.N.; Baird, K.J.; Hay, S.I. The Global Public Health Significance of Plasmodium vivax. In Advances in Parasitology; Academic Press: New York, NY, USA, 2012; Volume 80, pp. 1–111. [Google Scholar]
- Havelaar, A.H.; Kirk, M.D.; Torgerson, P.R.; Gibb, H.J.; Hald, T.; Lake, R.J.; Praet, N.; Bellinger, D.C.; de Silva, N.R.; Gargouri, N.; et al. World Health Organization Global Estimates and Regional Comparisons of the Burden of Foodborne Disease in 2010. PLoS Med. 2015, 12, e1001923. [Google Scholar] [CrossRef]
- Torgerson, P.R.; Mastroiacovo, P. The global burden of congenital toxoplasmosis: A systematic review. Bull. World Health Organ. 2013, 91, 501–508. [Google Scholar] [CrossRef]
- Escalante, A.A.; Ayala, F.J. Evolutionary origin of Plasmodium and other Apicomplexa based on rRNA genes. Proc. Natl. Acad. Sci. USA 1995, 92, 5793–5797. [Google Scholar] [CrossRef]
- White, M.W.; Suvorova, E.S. Apicomplexa Cell Cycles: Something Old, Borrowed, Lost, and New. Trends Parasitol. 2018, 34, 759–771. [Google Scholar] [CrossRef]
- Krishnan, A.; Kloehn, J.; Lunghi, M.; Chiappino-Pepe, A.; Waldman, B.S.; Nicolas, D.; Varesio, E.; Hehl, A.; Lourido, S.; Hatzimanikatis, V.; et al. Functional and Computational Genomics Reveal Unprecedented Flexibility in Stage-Specific Toxoplasma Metabolism. Cell Host Microbe 2020, 27, 290–306.e11. [Google Scholar] [CrossRef] [PubMed]
- Aurrecoechea, C.; Barreto, A.; Basenko, E.Y.; Brestelli, J.; Brunk, B.P.; Cade, S.; Crouch, K.; Doherty, R.; Falke, D.; Fischer, S.; et al. EuPathDB: The eukaryotic pathogen genomics database resource. Nucleic Acids Res. 2017, 45, D581–D591. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Chiasson, M.A.; Nursimulu, N.; Hung, S.S.; Wasmuth, J.; Grigg, M.E.; Parkinson, J. Metabolic reconstruction identifies strain-specific regulation of virulence in Toxoplasma gondii. Mol. Syst. Biol. 2013, 9, 708. [Google Scholar] [CrossRef] [PubMed]
- Chiappino-Pepe, A.; Tymoshenko, S.; Ataman, M.; Soldati-Favre, D.; Hatzimanikatis, V. Bioenergetics-based modeling of Plasmodium falciparum metabolism reveals its essential genes, nutritional requirements, and thermodynamic bottlenecks. PLoS Comput. Biol. 2017, 13, e1005397. [Google Scholar] [CrossRef] [PubMed]
- Tymoshenko, S.; Oppenheim, R.D.; Agren, R.; Nielsen, J.; Soldati-Favre, D.; Hatzimanikatis, V. Metabolic Needs and Capabilities of Toxoplasma gondii through Combined Computational and Experimental Analysis. PLoS Comput. Biol. 2015, 11, e1004261. [Google Scholar] [CrossRef]
- Stanway, R.R.; Bushell, E.; Chiappino-Pepe, A.; Roques, M.; Soldati-Favre, D.; Janse, C.J.; Hatzimanikatis, V.; Billker, O.; Heussler, V.T. Genome Scale Identification of Essential Metabolic Processes for Targeting the Plasmodium Liver Stage. Cell 2019, 179, 1112–1128. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Wang, C.; Otto, T.D.; Oberstaller, J.; Liao, X.; Adapa, S.R.; Udenze, K.; Bronner, I.F.; Casandra, D.; Mayho, M.; et al. Uncovering the essential genes of the human malaria parasite Plasmodium falciparum by saturation mutagenesis. Science 2018, 360, eaap7847. [Google Scholar] [CrossRef] [PubMed]
- Bushell, E.; Gomes, A.R.; Sanderson, T.; Anar, B.; Girling, G.; Herd, C.; Metcalf, T.; Modrzynska, K.; Schwach, F.; Martin, R.E.; et al. Functional Profiling of a Plasmodium Genome Reveals an Abundance of Essential Genes. Cell 2017, 170, 260–272.e8. [Google Scholar] [CrossRef] [PubMed]
- Sidik, S.M.; Huet, D.; Ganesan, S.M.; Huynh, M.-H.H.; Wang, T.; Nasamu, A.S.; Thiru, P.; Saeij, J.P.J.; Carruthers, V.B.; Niles, J.C.; et al. A Genome-wide CRISPR Screen in Toxoplasma Identifies Essential Apicomplexan Genes. Cell 2016, 166, 1423–1435.e12. [Google Scholar] [CrossRef]
- Hehl, A.B.; Basso, W.U.; Lippuner, C.; Ramakrishnan, C.; Okoniewski, M.; Walker, R.A.; Grigg, M.E.; Smith, N.C.; Deplazes, P. Asexual expansion of Toxoplasma gondii merozoites is distinct from tachyzoites and entails expression of non-overlapping gene families to attach, invade, and replicate within feline enterocytes. BMC Genom. 2015, 16, 66. [Google Scholar] [CrossRef]
- Otto, T.D.; Böhme, U.; Jackson, A.P.; Hunt, M.; Franke-Fayard, B.; Hoeijmakers, W.A.M.; Religa, A.A.; Robertson, L.; Sanders, M.; Ogun, S.A.; et al. A comprehensive evaluation of rodent malaria parasite genomes and gene expression. BMC Biol. 2014, 12, 86. [Google Scholar] [CrossRef]
- Caldelari, R.; Dogga, S.; Schmid, M.W.; Franke-Fayard, B.; Janse, C.J.; Soldati-Favre, D.; Heussler, V. Transcriptome analysis of Plasmodium berghei during exo-erythrocytic development. Malar. J. 2019, 18, 330. [Google Scholar] [CrossRef]
- Teng, R.; Junankar, P.R.; Bubb, W.A.; Rae, C.; Mercier, P.; Kirk, K. Metabolite profiling of the intraerythrocytic malaria parasite Plasmodium falciparum by 1H NMR spectroscopy. NMR Biomed. 2009, 22, 292–302. [Google Scholar] [CrossRef]
- Teng, R.; Lehane, A.M.; Winterberg, M.; Shafik, S.H.; Summers, R.L.; Martin, R.E.; Van Schalkwyk, D.A.; Junankar, P.R.; Kirk, K. 1H-NMR metabolite profiles of different strains of Plasmodium falciparum. Biosci. Rep. 2014, 34, 685–699. [Google Scholar] [CrossRef] [PubMed]
- Vo Duy, S.; Besteiro, S.; Berry, L.; Perigaud, C.; Bressolle, F.; Vial, H.J.; Lefebvre-Tournier, I. A quantitative liquid chromatography tandem mass spectrometry method for metabolomic analysis of Plasmodium falciparum lipid related metabolites. Anal. Chim. Acta 2012, 739, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Barylyuk, K.; Koreny, L.; Ke, H.; Butterworth, S.; Crook, O.M.; Lassadi, I.; Gupta, V.; Tromer, E.; Mourier, T.; Stevens, T.J.; et al. A Comprehensive Subcellular Atlas of the Toxoplasma Proteome via hyperLOPIT Provides Spatial Context for Protein Functions. Cell Host Microbe 2020, 28, 752–766.e9. [Google Scholar] [CrossRef] [PubMed]
- Parker, K.E.R.; Fairweather, S.J.; Rajendran, E.; Blume, M.; McConville, M.J.; Bröer, S.; Kirk, K.; van Dooren, G.G. The tyrosine transporter of Toxoplasma gondii is a member of the newly defined apicomplexan amino acid transporter (ApiAT) family. PLoS Pathog. 2019, 15, e1007577. [Google Scholar] [CrossRef]
- Rajendran, E.; Hapuarachchi, S.V.; Miller, C.M.; Fairweather, S.J.; Cai, Y.; Smith, N.C.; Cockburn, I.A.; Broër, S.; Kirk, K.; Van Dooren, G.G. Cationic amino acid transporters play key roles in the survival and transmission of apicomplexan parasites. Nat. Commun. 2017, 8, 1–13. [Google Scholar] [CrossRef]
- Dou, Z.; McGovern, O.L.; Di Cristina, M.; Carruthers, V.B. Toxoplasma gondii Ingests and Digests Host Cytosolic Proteins. MBio 2014, 5, e01188-14. [Google Scholar] [CrossRef]
- Burgess, S.L.; Gilchrist, C.A.; Lynn, T.C.; Petri, W.A. Parasitic Protozoa and Interactions with the Host Intestinal Microbiota. Infect. Immun. 2017, 85, e00101-17. [Google Scholar] [CrossRef]
- O’Hara, S.P.; Chen, X.-M. The cell biology of cryptosporidium infection. Microbes Infect. 2011, 13, 721–730. [Google Scholar] [CrossRef]
- Mehlhorn, H.; Schein, E. The Piroplasms: Life Cycle and Sexual Stages. Adv. Parasitol. 1985, 23, 37–103. [Google Scholar]
- Jalovecka, M.; Hajdusek, O.; Sojka, D.; Kopacek, P.; Malandrin, L. The Complexity of Piroplasms Life Cycles. Front. Cell. Infect. Microbiol. 2018, 8, 248. [Google Scholar] [CrossRef]
- MacRae, J.I.; Sheiner, L.; Nahid, A.; Tonkin, C.; Striepen, B.; McConville, M.J. Mitochondrial metabolism of glucose and glutamine is required for intracellular growth of Toxoplasma gondii. Cell Host Microbe 2012, 12, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Cava, F.; Lam, H.; De Pedro, M.A.; Waldor, M.K. Emerging knowledge of regulatory roles of d-amino acids in bacteria. Cell. Mol. Life Sci. 2011, 68, 817–831. [Google Scholar] [CrossRef] [PubMed]
- Cobbold, S.A.; Martin, R.E.; Kirk, K. Methionine transport in the malaria parasite Plasmodium falciparum. Int. J. Parasitol. 2011, 41, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Wrenger, C.; Müller, I.B.; Schifferdecker, A.J.; Jain, R.; Jordanova, R.; Groves, M.R. Specific inhibition of the aspartate aminotransferase of Plasmodium falciparum. J. Mol. Biol. 2011, 405, 956–971. [Google Scholar] [CrossRef]
- Nagaraj, V.A.; Mukhi, D.; Sathishkumar, V.; Subramani, P.A.; Ghosh, S.K.; Pandey, R.R.; Shetty, M.C.; Padmanaban, G. Asparagine requirement in Plasmodium berghei as a target to prevent malaria transmission and liver infections. Nat. Commun. 2015, 6, 1–13. [Google Scholar] [CrossRef]
- Lunev, S.; Bosch, S.S.; Batista, F.D.A.; Wrenger, C.; Groves, M.R. Crystal structure of truncated aspartate transcarbamoylase from Plasmodium falciparum. Acta Crystallogr. Sect. Struct. Biol. Commun. 2016, 72, 523–533. [Google Scholar] [CrossRef]
- Booker, M.L.; Bastos, C.M.; Kramer, M.L.; Barker, R.H.; Skerlj, R.; Sidhu, A.B.; Deng, X.; Celatka, C.; Cortese, J.F.; Guerrero Bravo, J.E.; et al. Novel inhibitors of Plasmodium falciparum dihydroorotate dehydrogenase with anti-malarial activity in the mouse model. J. Biol. Chem. 2010, 285, 33054–33064. [Google Scholar] [CrossRef]
- Phillips, M.A.; Lotharius, J.; Marsh, K.; White, J.; Dayan, A.; White, K.L.; Njoroge, J.W.; El Mazouni, F.; Lao, Y.; Kokkonda, S.; et al. A long-duration dihydroorotate dehydrogenase inhibitor (DSM265) for prevention and treatment of malaria. Sci. Transl. Med. 2015, 7, 296ra111. [Google Scholar] [CrossRef]
- Blume, M.; Nitzsche, R.; Sternberg, U.; Gerlic, M.; Masters, S.L.; Gupta, N.; McConville, M.J. A Toxoplasma gondii gluconeogenic enzyme contributes to robust central carbon metabolism and is essential for replication and virulence. Cell Host Microbe 2015, 18, 210–220. [Google Scholar] [CrossRef]
- Nitzsche, R.; Zagoriy, V.; Lucius, R.; Gupta, N. Metabolic cooperation of glucose and glutamine is essential for the lytic cycle of obligate intracellular parasite Toxoplasma gondii. J. Biol. Chem. 2016, 291, 126–141. [Google Scholar] [CrossRef]
- Oppenheim, R.D.; Creek, D.J.; Macrae, J.I.; Modrzynska, K.K.; Pino, P.; Limenitakis, J.; Polonais, V.; Seeber, F.; Barrett, M.P.; Billker, O.; et al. BCKDH: The missing link in apicomplexan mitochondrial metabolism is required for full virulence of Toxoplasma gondii and Plasmodium berghei. PLoS Pathog. 2014, 10, e1004263. [Google Scholar] [CrossRef] [PubMed]
- Blume, M.; Rodriguez-Contreras, D.; Landfear, S.; Fleige, T.; Soldati-Favre, D.; Lucius, R.; Gupta, N. Host-derived glucose and its transporter in the obligate intracellular pathogen Toxoplasma gondii are dispensable by glutaminolysis. Proc. Natl. Acad. Sci. USA 2009, 106, 12998–13003. [Google Scholar] [CrossRef] [PubMed]
- MacRae, J.I.; Dixon, M.W.A.; Dearnley, M.K.; Chua, H.H.; Chambers, J.M.; Kenny, S.; Bottova, I.; Tilley, L.; McConville, M.J. Mitochondrial metabolism of sexual and asexual blood stages of the malaria parasite Plasmodium falciparum. BMC Biol. 2013, 11, 67. [Google Scholar] [CrossRef] [PubMed]
- Meireles, P.; Mendes, A.M.; Aroeira, R.I.; Mounce, B.C.; Vignuzzi, M.; Staines, H.M.; Prudêncio, M. Uptake and metabolism of arginine impact Plasmodium development in the liver. Sci. Rep. 2017, 7, 4072. [Google Scholar] [CrossRef] [PubMed]
- Narvaez-Ortiz, H.Y.; Lopez, A.J.; Gupta, N.; Zimmermann, B.H. A CTP synthase undergoing stage-specific spatial expression is essential for the survival of the intracellular parasite Toxoplasma gondii. Front. Cell. Infect. Microbiol. 2018, 8, 83. [Google Scholar] [CrossRef] [PubMed]
- Kloehn, J.; Oppenheim, R.D.; Siddiqui, G.; De Bock, P.J.; Kumar Dogga, S.; Coute, Y.; Hakimi, M.A.; Creek, D.J.; Soldati-Favre, D. Multi-omics analysis delineates the distinct functions of sub-cellular acetyl-CoA pools in Toxoplasma gondii. BMC Biol. 2020, 18, 67. [Google Scholar] [CrossRef]
- Erler, H.; Ren, B.; Gupta, N.; Beitz, E. The intracellular parasite Toxoplasma gondii harbors three druggable FNT-type formate and l-lactate transporters in the plasma membrane. J. Biol. Chem. 2018, 293, 17622–17630. [Google Scholar] [CrossRef]
- Krishnan, A.; Kloehn, J.; Lunghi, M.; Soldati-Favre, D. Vitamin and cofactor acquisition in apicomplexans: Synthesis versus salvage. J. Biol. Chem. 2020, 295, 701–714. [Google Scholar] [CrossRef]
- Alfadhli, S.; Rathod, P.K. Gene organization of a Plasmodium falciparum serine hydroxymethyltransferase and its functional expression in Escherichia coli. Mol. Biochem. Parasitol. 2000, 110, 283–291. [Google Scholar] [CrossRef]
- Sopitthummakhun, K.; Thongpanchang, C.; Vilaivan, T.; Yuthavong, Y.; Chaiyen, P.; Leartsakulpanich, U. Plasmodium serine hydroxymethyltransferase as a potential anti-malarial target: Inhibition studies using improved methods for enzyme production and assay. Malar. J. 2012, 11, 194. [Google Scholar] [CrossRef]
- Xia, N.; Ye, S.; Liang, X.; Chen, P.; Zhou, Y.; Fang, R.; Zhao, J.; Gupta, N.; Yang, S.; Yuan, J.; et al. Pyruvate homeostasis as a determinant of parasite growth and metabolic plasticity in Toxoplasma gondii. MBio 2019, 10, 2020. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, A.; Floyd, K.; Key, M.; Dameron, C.; Rees, K.C.; Brock Thornton, L.; Whitehead, D.C.; Hamza, I.; Dou, Z. Toxoplasma gondii requires its plant-like heme biosynthesis pathway for infection. PLoS Pathog. 2020, 16, e1008499. [Google Scholar] [CrossRef] [PubMed]
- Nagaraj, V.A.; Sundaram, B.; Varadarajan, N.M.; Subramani, P.A.; Kalappa, D.M.; Ghosh, S.K.; Padmanaban, G. Malaria Parasite-Synthesized Heme is Essential in the Mosquito and Liver Stages and Complements Host Heme in the Blood Stages of Infection. PLoS Pathog. 2013, 9, e1003522. [Google Scholar] [CrossRef] [PubMed]
- Ke, H.; Sigala, P.A.; Miura, K.; Morrisey, J.M.; Mather, M.W.; Crowley, J.R.; Henderson, J.P.; Goldberg, D.E.; Long, C.A.; Vaidya, A.B. The heme biosynthesis pathway is essential for Plasmodium falciparum development in mosquito stage but not in blood stages. J. Biol. Chem. 2014, 289, 34827–34837. [Google Scholar] [CrossRef] [PubMed]
- Sigala, P.A.; Crowley, J.R.; Henderson, J.P.; Goldberg, D.E. Deconvoluting heme biosynthesis to target blood-stage malaria parasites. Elife 2015, 4, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Gafan, C.; Wilson, J.; Berger, L.C.; Berger, B.J. Characterization of the ornithine aminotransferase from Plasmodium falciparum. Mol. Biochem. Parasitol. 2001, 118, 1–10. [Google Scholar] [CrossRef]
- Jortzik, E.; Fritz-Wolf, K.; Sturm, N.; Hipp, M.; Rahlfs, S.; Becker, K. Redox regulation of Plasmodium falciparum ornithine δ-aminotransferase. J. Mol. Biol. 2010, 402, 445–459. [Google Scholar] [CrossRef]
- Radtke, L.; Kletter, T.; Kreibich, J.; Niedersen, L.; Nitzsche, R.; Gupta, N. Toxoplasma gondii can synthesize the amino acids otherwise essential for its mammalian host cells. Abstract book, Molecular Parasitology Meeting. 2019; Unpublished. [Google Scholar]
- Hudson, A.O.; Singh, B.K.; Leustek, T.; Gilvarg, C. An LL-diaminopimelate aminotransferase defines a novel variant of the lysine biosynthesis pathway in plants. Plant Physiol. 2006, 140, 292–301. [Google Scholar] [CrossRef]
- Rajendran, E.; Clark, M.; Goulart, C.; Steinhöfel, B.; Tjhin, E.T.; Smith, N.C.; Kirk, K.; van Dooren, G.G. Substrate-mediated regulation of the arginine transporter of Toxoplasma gondii. bioRxiv 2019, 798967. [Google Scholar] [CrossRef]
- Morris, S.M. Arginine: Beyond protein. Am. J. Clin. Nutr. 2006, 83, 508S–512S. [Google Scholar] [CrossRef]
- Seabra, S.H.; De Souza, W.; Damatta, R.A. Toxoplasma gondii exposes phosphatidylserine inducing a TGF-β 1 autocrine effect orchestrating macrophage evasion. Biochem. Biophys. Res. Commun. 2004, 324, 744–752. [Google Scholar] [CrossRef] [PubMed]
- Butcher, B.A.; Fox, B.A.; Rommereim, L.M.; Kim, S.G.; Maurer, K.J.; Yarovinsky, F.; Herbert, D.R.; Bzik, D.J.; Denkers, E.Y. Toxoplasma gondii rhoptry kinase rop16 activates stat3 and stat6 resulting in cytokine inhibition and arginase-1-dependent growth control. PLoS Pathog. 2011, 7, e1002236. [Google Scholar] [CrossRef] [PubMed]
- Fox, B.A.; Gigley, J.P.; Bzik, D.J. Toxoplasma gondii lacks the enzymes required for de novo arginine biosynthesis and arginine starvation triggers cyst formation. Int. J. Parasitol. 2004, 34, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Cobbold, S.A.; Llinás, M.; Kirk, K. Sequestration and metabolism of host cell arginine by the intraerythrocytic malaria parasite Plasmodium falciparum. Cell. Microbiol. 2016, 18, 820–830. [Google Scholar] [CrossRef] [PubMed]
- Meireles, P.; Brás, D.; Fontinha, D.; Chora, Â.F.; Serre, K.; Mendes, A.M.; Prudêncio, M. Elimination of Hepatic Rodent Plasmodium Parasites by Amino Acid Supplementation. iScience 2020, 23, 101781. [Google Scholar] [CrossRef] [PubMed]
- Limenitakis, J.; Oppenheim, R.D.; Creek, D.J.; Foth, B.J.; Barrett, M.P.; Soldati-Favre, D. The 2-methylcitrate cycle is implicated in the detoxification of propionate in Toxoplasma gondii. Mol. Microbiol. 2013, 87, 894–908. [Google Scholar] [CrossRef]
- Martin, R.E.; Kirk, K. Transport of the essential nutrient isoleucine in human erythrocytes infected with the malaria parasite Plasmodium falciparum. Blood 2007, 109, 2217–2224. [Google Scholar] [CrossRef]
- Prior, K.F.; Middleton, B.; Owolabi, A.T.; Westwood, M.L.; Holland, J.; Blackman, M.; Skene, D.J.; Reece, S.E. An essential amino acid synchronises malaria parasite 1 development with daily host rhythms. bioRxiv 2020. [Google Scholar] [CrossRef]
- Babbitt, S.E.; Altenhofen, L.; Cobbold, S.A.; Istvan, E.S.; Fennellf, C.; Doerig, C.; Llinás, M.; Goldberg, D.E. Plasmodium falciparum responds to amino acid starvation by entering into a hibernatory state. Proc. Natl. Acad. Sci. USA 2012, 109, E3278–E3287. [Google Scholar] [CrossRef]
- McLean, K.J.; Jacobs-Lorena, M. The response of Plasmodium falciparum to isoleucine withdrawal is dependent on the stage of progression through the intraerythrocytic cell cycle. Malar. J. 2020, 19, 1–9. [Google Scholar] [CrossRef]
- Behnke, M.S.; Khan, A.; David Sibley, L. Genetic mapping reveals that sinefungin resistance in Toxoplasma gondii is controlled by a putative amino acid transporter locus that can be used as a negative selectable marker. Eukaryot. Cell 2015, 14, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.T.; Harmon, S.; O’Malley, K.L.; Sibley, L.D. Reassessment of the role of aromatic amino acid hydroxylases and the effect of infection by Toxoplasma gondii on host dopamine. Infect. Immun. 2015, 83, 1039–1047. [Google Scholar] [CrossRef] [PubMed]
- Marino, N.D.; Boothroyd, J.C. Toxoplasma growth in vitro is dependent on exogenous tyrosine and is independent of AAH2 even in tyrosine-limiting conditions. Exp. Parasitol. 2017, 176, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.T.; Verma, S.K.; Dubey, J.P.; Sibley, L.D. The aromatic amino acid hydroxylase genes AAH1 and AAH2 in Toxoplasma gondii contribute to transmission in the cat. PLoS Pathog. 2017, 13, e1006272. [Google Scholar] [CrossRef] [PubMed]
- Naemat, A.; Elsheikha, H.M.; Boitor, R.A.; Notingher, I. Tracing amino acid exchange during host-pathogen interaction by combined stable-isotope time-resolved Raman spectral imaging. Sci. Rep. 2016, 6, 20811. [Google Scholar] [CrossRef]
- Fujigaki, S.; Saito, K.; Takemura, M.; Maekawa, N.; Yamada, Y.; Wada, H.; Seishima, M. L-tryptophan-L-kynurenine pathway metabolism accelerated by Toxoplasma gondii infection is abolished in gamma interferon-gene-deficient mice: Cross-regulation between inducible nitric oxide synthase and indoleamine-2,3-dioxygenase. Infect. Immun. 2002, 70, 779–786. [Google Scholar] [CrossRef]
- Ufermann, C.-M.; Domröse, A.; Babel, T.; Tersteegen, A.; Cengiz, S.C.; Eller, S.K.; Spekker-Bosker, K.; Sorg, U.R.; Förster, I.; Däubener, W. Indoleamine 2,3-Dioxygenase Activity during Acute Toxoplasmosis and the Suppressed T Cell Proliferation in Mice. Front. Cell. Infect. Microbiol. 2019, 9, 184. [Google Scholar] [CrossRef]
- Olson, W.J.; Martorelli Di Genova, B.; Gallego-Lopez, G.; Dawson, A.R.; Stevenson, D.; Amador-Noguez, D.; Knoll, L.J. Dual metabolomic profiling uncovers Toxoplasma manipulation of the host metabolome and the discovery of a novel parasite metabolic capability. PLoS Pathog. 2020, 16, e1008432. [Google Scholar] [CrossRef]
- Dos Santos, L.M.; Commodaro, A.G.; Vasquez, A.R.R.; Kohlhoff, M.; de Paula Guerra, D.A.; Coimbra, R.S.; Martins-Filho, O.A.; Teixeira-Carvalho, A.; Rizzo, L.V.; Vieira, L.Q.; et al. Intestinal microbiota regulates tryptophan metabolism following oral infection with Toxoplasma gondii. Parasite Immunol. 2020, 42, e12720. [Google Scholar]
- Tetsutani, K.; To, H.; Torii, M.; Hisaeda, H.; Himeno, K. Malaria parasite induces tryptophan-related immune suppression in mice. Parasitology 2007, 134, 923–930. [Google Scholar] [CrossRef]
- Moffett, J.R.; Namboodiri, M.A. Tryptophan and the immune response. Immunol. Cell Biol. 2003, 81, 247–265. [Google Scholar] [CrossRef] [PubMed]
- Gomes, P.S.; Bhardwaj, J.; Rivera-Correa, J.; Freire-De-Lima, C.G.; Morrot, A. Immune escape strategies of malaria parasites. Front. Microbiol. 2016, 7, 1617. [Google Scholar] [CrossRef] [PubMed]
- Wagner, I.; Musso, H. New Naturally Occurring Amino Acids. Angew. Chemie Int. Ed. Engl. 1983, 22, 816–828. [Google Scholar] [CrossRef]
- Matz, J.M.; Watanabe, M.; Falade, M.; Tohge, T.; Hoefgen, R.; Correspondence, K.M. Plasmodium Para-Aminobenzoate Synthesis and Salvage Resolve Avoidance of Folate Competition and Adaptation to Host Diet. Cell Rep. 2019, 26, 356–363.e4. [Google Scholar] [CrossRef] [PubMed]
- Mather, M.W.; Ke, H. Para-Aminobenzoate Synthesis versus Salvage in Malaria Parasites. Trends Parasitol. 2019, 35, 176–178. [Google Scholar] [CrossRef]
- Hunter, E.S.; Paight, C.; Lane, C.E. Metabolic Contributions of an Alphaproteobacterial Endosymbiont in the Apicomplexan Cardiosporidium cionae. Front. Microbiol. 2020, 11, 3014. [Google Scholar] [CrossRef]
- Goldberg, D.E. Complex nature of malaria parasite hemoglobin degradation. Proc. Natl. Acad. Sci. USA 2013, 110, 5283–5284. [Google Scholar] [CrossRef]
- Gavigan, C.S.; Dalton, J.P.; Bell, A. The role of aminopeptidases in haemoglobin degradation in Plasmodium falciparum-infected erythrocytes. Mol. Biochem. Parasitol. 2001, 117, 37–48. [Google Scholar] [CrossRef]
- Lee, A.H.; Dhingra, S.K.; Lewis, I.A.; Singh, M.K.; Siriwardana, A.; Dalal, S.; Rubiano, K.; Klein, M.S.; Baska, K.S.; Krishna, S.; et al. Evidence for Regulation of Hemoglobin Metabolism and Intracellular Ionic Flux by the Plasmodium falciparum Chloroquine Resistance Transporter. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Pino, P.; Aeby, E.; Foth, B.J.; Sheiner, L.; Soldati, T.; Schneider, A.; Soldati-Favre, D. Mitochondrial translation in absence of local tRNA aminoacylation and methionyl tRNAMet formylation in Apicomplexa. Mol. Microbiol. 2010, 76, 706–718. [Google Scholar] [CrossRef]
- Jain, V.; Yogavel, M.; Kikuchi, H.; Oshima, Y.; Hariguchi, N.; Matsumoto, M.; Goel, P.; Touquet, B.; Jumani, R.S.; Tacchini-Cottier, F.; et al. Targeting prolyl-tRNA synthetase to accelerate drug discovery against malaria, leishmaniasis, toxoplasmosis, cryptosporidiosis, and coccidiosis. Structure 2017, 25, 1495–1505. [Google Scholar] [CrossRef] [PubMed]
- Palencia, A.; Liu, R.J.; Lukarska, M.; Gut, J.; Bougdour, A.; Touquet, B.; Wang, E.-D.; Li, X.; Alley, M.R.K.; Freund, Y.R.; et al. Cryptosporidium and Toxoplasma parasites are inhibited by a benzoxaborole targeting leucyl-tRNA synthetase. Antimicrob. Agents Chemother. 2016, 60, 5817–5827. [Google Scholar] [CrossRef] [PubMed]
- Cowell, A.N.; Winzeler, E.A. Advances in omics-based methods to identify novel targets for malaria and other parasitic protozoan infections. Genome Med. 2019, 11, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Moreira, A.S.; Estato, V.; Malvar, D.C.; Sanches, G.S.; Gomes, F.; Tibirica, E.; Daniel-Ribeiro, C.T.; Carvalho, L.J.M. L-arginine supplementation and thromboxane synthase inhibition increases cerebral blood flow in experimental cerebral malaria. Sci. Rep. 2019, 9, 13621. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krishnan, A.; Soldati-Favre, D. Amino Acid Metabolism in Apicomplexan Parasites. Metabolites 2021, 11, 61. https://doi.org/10.3390/metabo11020061
Krishnan A, Soldati-Favre D. Amino Acid Metabolism in Apicomplexan Parasites. Metabolites. 2021; 11(2):61. https://doi.org/10.3390/metabo11020061
Chicago/Turabian StyleKrishnan, Aarti, and Dominique Soldati-Favre. 2021. "Amino Acid Metabolism in Apicomplexan Parasites" Metabolites 11, no. 2: 61. https://doi.org/10.3390/metabo11020061
APA StyleKrishnan, A., & Soldati-Favre, D. (2021). Amino Acid Metabolism in Apicomplexan Parasites. Metabolites, 11(2), 61. https://doi.org/10.3390/metabo11020061