1. Introduction
Maternal depression is a general term referring to a spectrum of depressive conditions that typically occur during pregnancy and up to 12 months postpartum [
1]. It is also a critical period of fetal and child development, and maternal disorders can have far-reaching impacts on offspring [
2,
3]. Maternal depression is a known risk factor for preterm birth, low birth weight, and intrauterine growth restriction [
4,
5]. These complications are reported to be the leading cause of death in children under five years of age [
6,
7].
Many studies have examined the effects of prenatal maternal stress, anxiety, and depression on child development, impacting cognitive, behavioral, social, mood, language, and attachment outcomes [
8,
9,
10]. Children of depressed mothers also face higher rates of cognitive, social, and mood disorders later in life [
11,
12]. Distinguishing the effects of untreated prenatal depression from those of other factors is challenging. Animal models, with their controlled environments, help us better understand the adverse effects of maternal stress and depression on offspring development.
The most frequently used antidepressant drugs are the second-generation antidepressants such as selective serotonin (5-HT) reuptake inhibitors (SSRIs) [
13,
14]. However, concerns have been raised about their use during pregnancy and the postpartum period because SSRIs easily cross the placenta and are present in breast milk, and fetuses and infants may have difficulty metabolizing SSRIs efficiently [
15,
16,
17]. SSRIs may inadvertently affect serotonin signaling in offspring, leading to doubts about whether prescribing SSRIs to pregnant and postpartum women is safe for offspring. Several studies suggest that prenatal SSRI exposure has developmental consequences (enhancement of anxiety states) in both rats and human offspring [
18,
19].
Mirtazapine, a third-generation antidepressant, according to a recent meta-study, belongs to a small group of the most effective antidepressants [
20] and is recommended for use during pregnancy. Mirtazapine is an antagonist of α
2-adrenergic inhibitory, 5-HT
2, and 5-HT
3 receptors [
21]. However, insufficient data are available to support its safe use during pregnancy [
22].
In this study, we focused on the possible effects of maternal depression and its pharmacological treatment during gestation on the neuronal development and behavior of offspring in an early stage of postnatal development (up to 2 weeks). As it is involved in learning memory, cognitive function, and emotion regulation, the hippocampus is among the key brain regions affected by the pathogenesis of depression [
23,
24,
25]. Therefore, we focused on possible alterations in hippocampal excitability.
As an animal model of maternal depression, we have used pregestational exposure to chronic unpredictable stress (CUS) [
17,
26,
27]. We had previously reported that the CUS dams showed behavioral features resembling the symptoms of depression, as well as impaired hippocampal neurogenesis. Furthermore, we have shown that the behavioral and neuroanatomical abnormalities in the CUS dams were reversed by chronic antidepressant treatment [
28]. CUS dams therefore possess constructive and predictive validity as a model of prenatal depression. In this work, we tested whether the previously observed alterations are reflected in altered hippocampal excitability. We compared the hippocampal excitability of offspring from four experimental groups: (i) dams not exposed to CUS and treated with vehicle; (ii) dams not exposed to CUS and treated with mirtazapine; (iii) dams exposed to CUS and treated with vehicle; and (iv) dams exposed to CUS and treated with mirtazapine.
The primary goal of our study was to test the hypothesis that the maternal stress and/or alteration in maternal monoamine neurotransmission during gestation affect the intrinsic properties of the embryonal glutamate neurons, or their progenitors. This alteration in the intrinsic properties of the embryonal neurons may subsequently affect their maturation and functioning in in vivo or in vitro conditions.
2. Results
2.1. Effects of Pregestational Stress and Prenatal Mirtazapine on the Electrophysiological Characteristics of Cultivated Pyramidal Neurons Isolated from Newborn Offspring Hippocampi
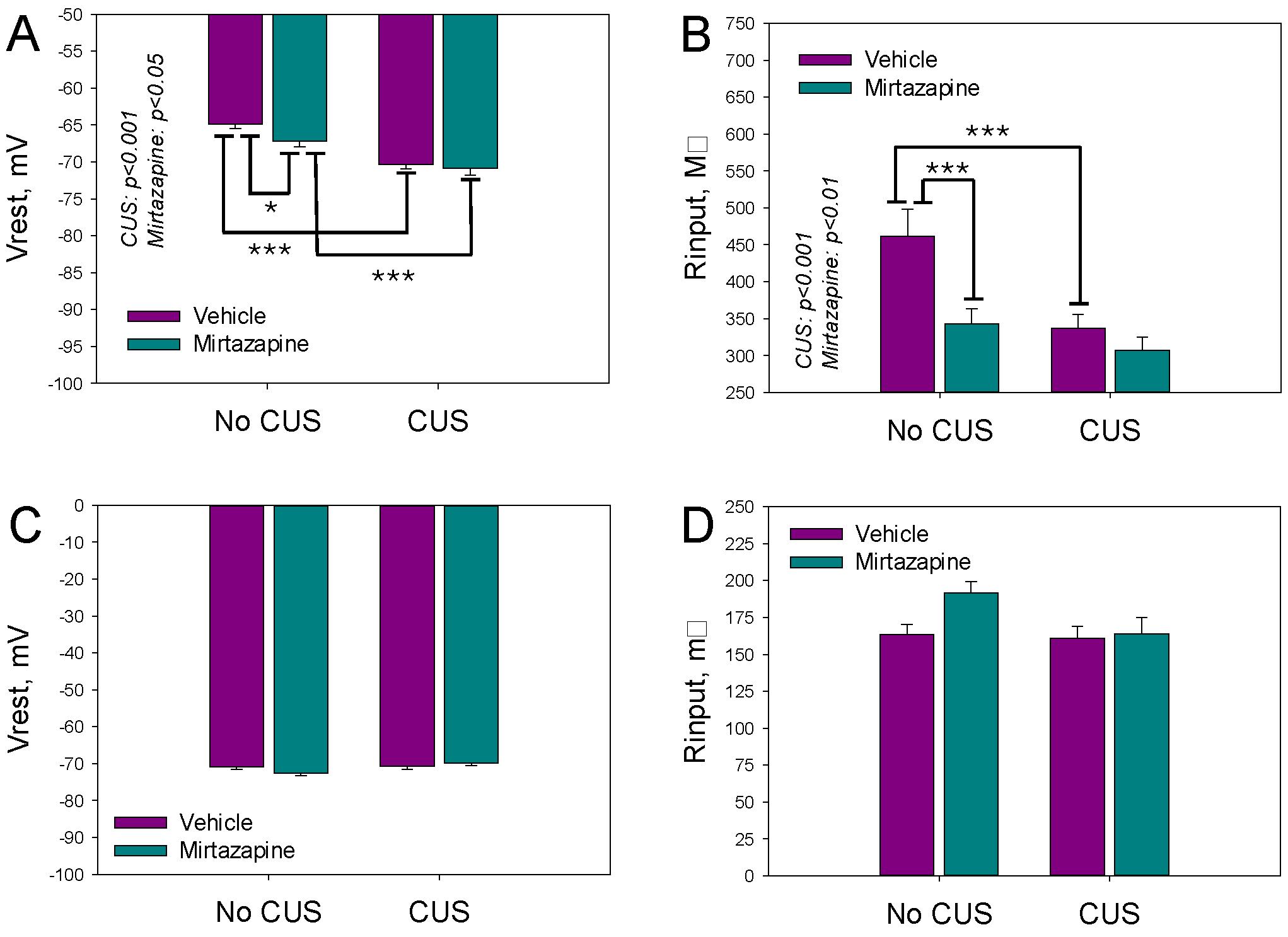
The resting membrane potentials (V
rest) and input resistance (R
inp) reflect the expression of voltage-gated ion channels in respective cells. V
rest and R
inp of the cultivated neurons isolated from the hippocampi of the newborn offspring of the dams exposed to pregestational CUS, prenatal mirtazapine treatment, or their combination are shown in
Figure 1A. The Two-way ANOVA revealed a significant effect of CUS and perinatal mirtazapine on V
rest (F
1,280 = 38.85,
p < 0.001, and F
1,280 = 4.04,
p = 0.045, respectively) and R
inp (F
1,279 = 11.57,
p < 0.001, and F
1,279 = 10.02,
p = 0.002, respectively). The interaction between these factors for V
rest and R
inp was not statistically significant.
Observed differences were diminished during 11–13 days of postnatal development, as shown in
Figure 1C,D. According to the two-way ANOVA results, maternal stress and mirtazapine treatment did not significantly affect V
rest or R
inp. The interactions between these factors were also not statistically significant.
The waveform of a single action potential (AP) activated by a brief depolarizing current pulse was characterized by the threshold (
Figure 2A) and peak (
Figure 2B) voltages, latency time (
Figure 2C), width of action potential (
Figure 2D), rise time (
Figure 2E), and minimal current needed to evoke a single action potential (
Figure 2F). CUS depressed excitability by increasing the voltage threshold for AP generation. Treatment by mirtazapine facilitated it by decreasing both the voltage threshold and minimal current amplitude needed for an activation of an AP.
For the monitored parameters, a significant effect of CUS on the peak voltage of the AP (F1,278 = 9.35 p = 0.002; two-way ANOVA) and its rise time (F1,278 = 9.67, p < 0.002; two-way ANOVA) was detected; a significant effect of maternal mirtazapine treatment on the threshold (F1,278 = 7.99, p = 0.005; two-way ANOVA) and peak (F1,278 = 11.28, p < 0.001; two-way ANOVA) voltages and latency times was detected (F1,278 = 4.13, p = 0.04; two-way ANOVA); and a significant interaction between these factors concerning the voltage threshold (F1,278 = 49.59, p < 0.001; two-way ANOVA), rise time (F1,278 = 13.27, p < 0.001; two-way ANOVA), and the minimal current needed to induce a single action potential was detected (F1,278 = 14.23, p < 0.001; two-way ANOVA).
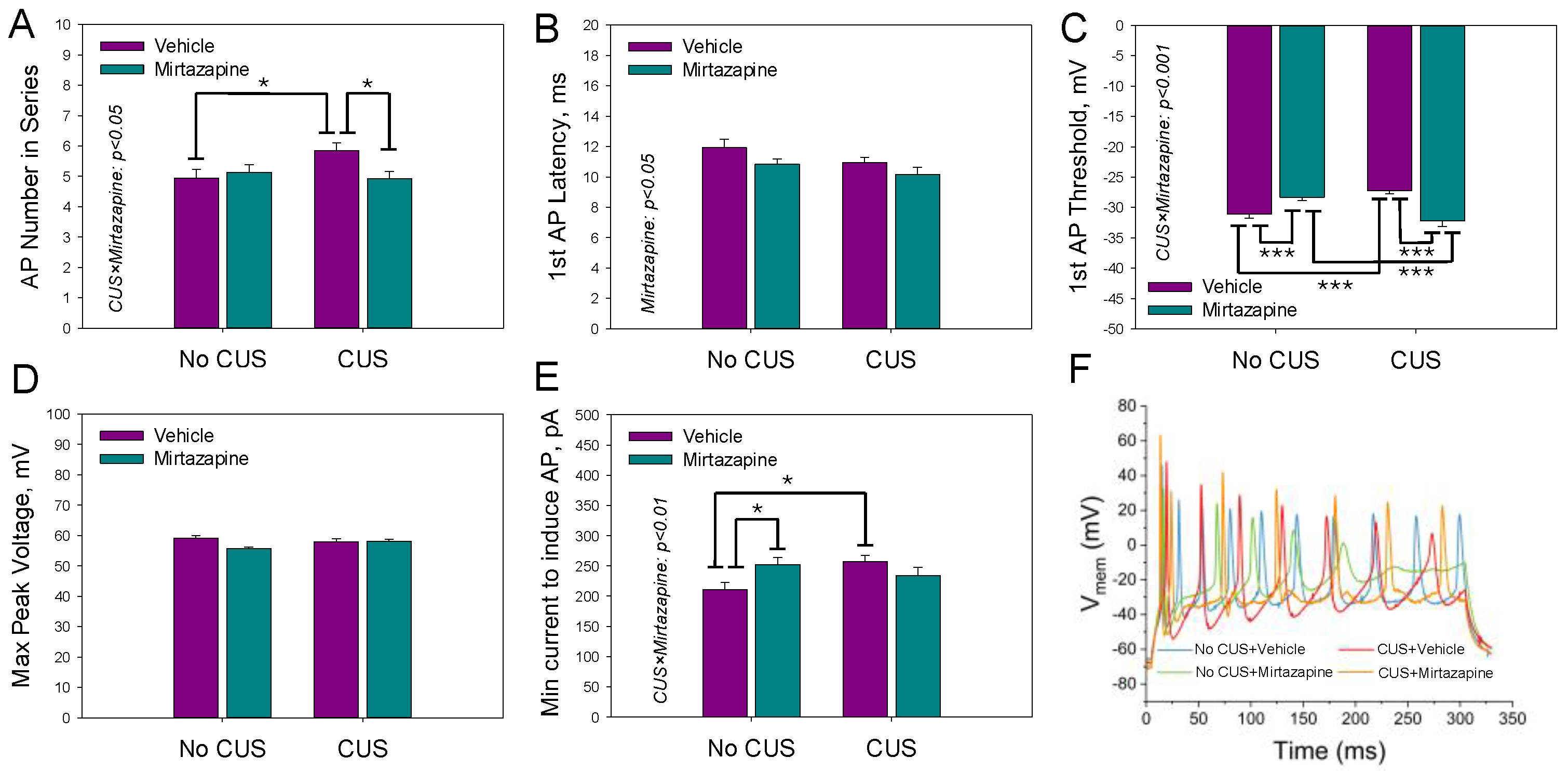
The parameters characterizing the AP series (three or more APs generated during a depolarizing current pulse) activated by the second suprathreshold current pulse were analyzed in the same experimental groups are shown in
Figure 3.
The two-way ANOVA revealed a statistically significant effect of maternal mirtazapine on the first AP latency time (F
1,225 = 4.93,
p = 0.03;
Figure 3B) and a significant interaction between maternal stress and mirtazapine with respect to the number of APs within the series (F
1,225 = 4.46,
p = 0.04;
Figure 3A), first AP threshold voltage (F
1,225 = 33.65,
p < 0.001;
Figure 3C), and minimal current required to induce the series (F
1,225 = 7.52,
p = 0.007;
Figure 3E). Again, CUS suppressed excitability by increasing both the voltage threshold for the generation of the first AP and minimal current amplitude needed for the generation of the first AP series. Mirtazapine rectified both changes.
Figure 4 illustrates the effects of maternal stress, mirtazapine, and their combination on the percentage of neurons in primary cultures with AP series observed during depolarizing current pulses with amplitudes between 100 and 400 pA. A significant interaction effect between maternal stress and mirtazapine was detected during depolarizing pulses of 300 (F
1,19 = 4.77,
p = 0.04), 350 (F
1,19 = 5.14,
p = 0.04), and 400 pA (F
1,19 = 6.44,
p = 0.02). CUS increased excitability at higher depolarizing pulses and mirtazapine treatment suppressed this effect.
2.2. Effects of Pregestational Stress and Prenatal Mirtazapine on the Electrophysiological Characteristics of Pyramidal Neurons in Hippocampal Slices Isolated from 11–13-Day-Old Offspring
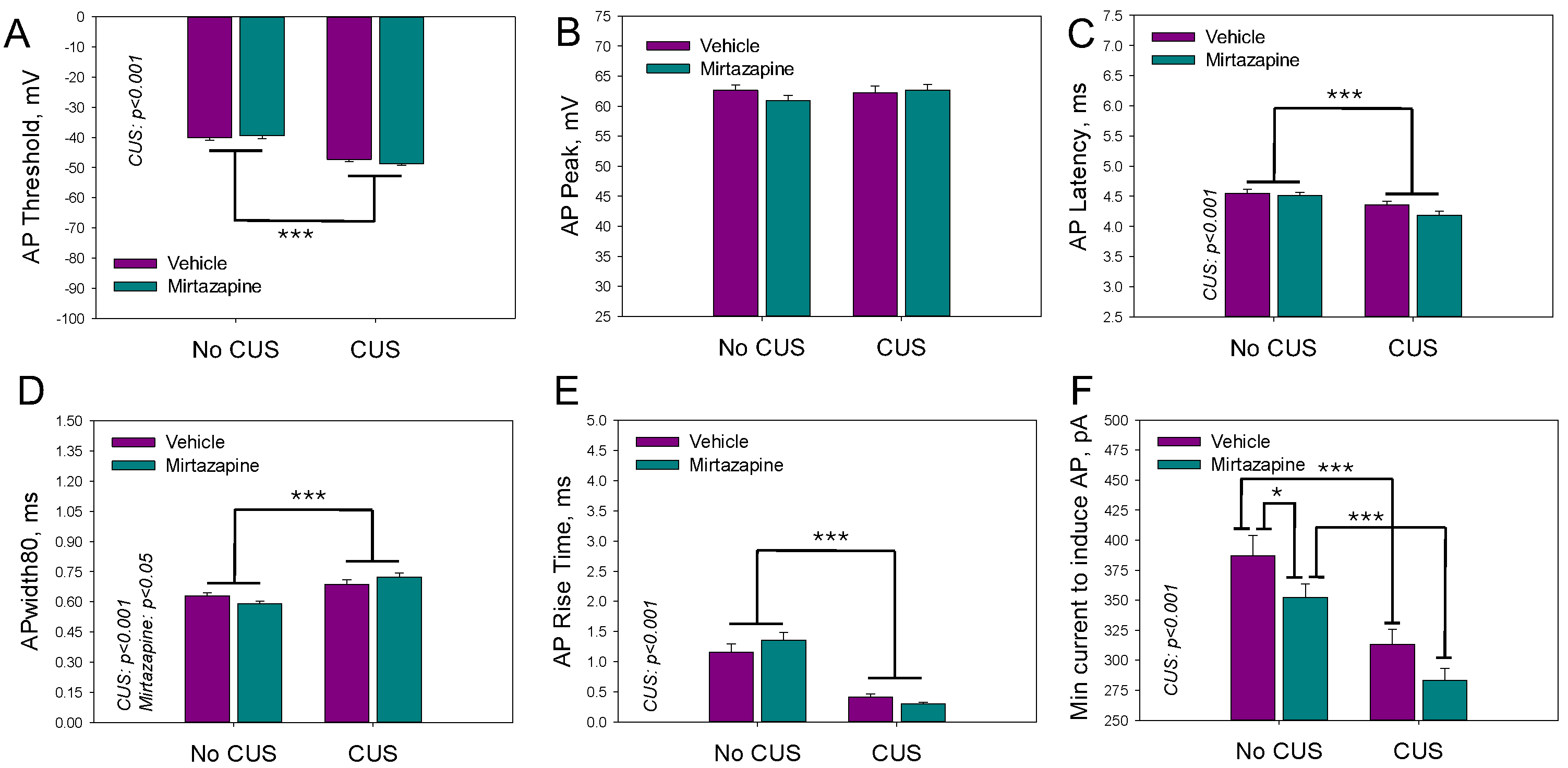
Individual AP parameters of the pyramidal neurons in the slices isolated from the hippocampi of 11–13-day-old offspring of the dams that experienced pregestational stress, prenatal mirtazapine treatment, or their combination are shown in
Figure 5. The two-way ANOVA revealed a significant effect of maternal stress on the threshold voltage (F
1,189 = 89.92,
p < 0.001), latency time (F
1,189 = 17.75,
p < 0.001), width of action potential (F
1,189 = 28.15,
p < 0.001), rise time (F
1,189 = 59.21,
p < 0.001), and minimal current needed to induce a series of action potentials (F
1,189 = 28.05,
p < 0.001). With respect to the width of the action potential, there was also a significant interaction effect between maternal stress and mirtazapine treatment (F
1,189 = 4.53,
p = 0.04).
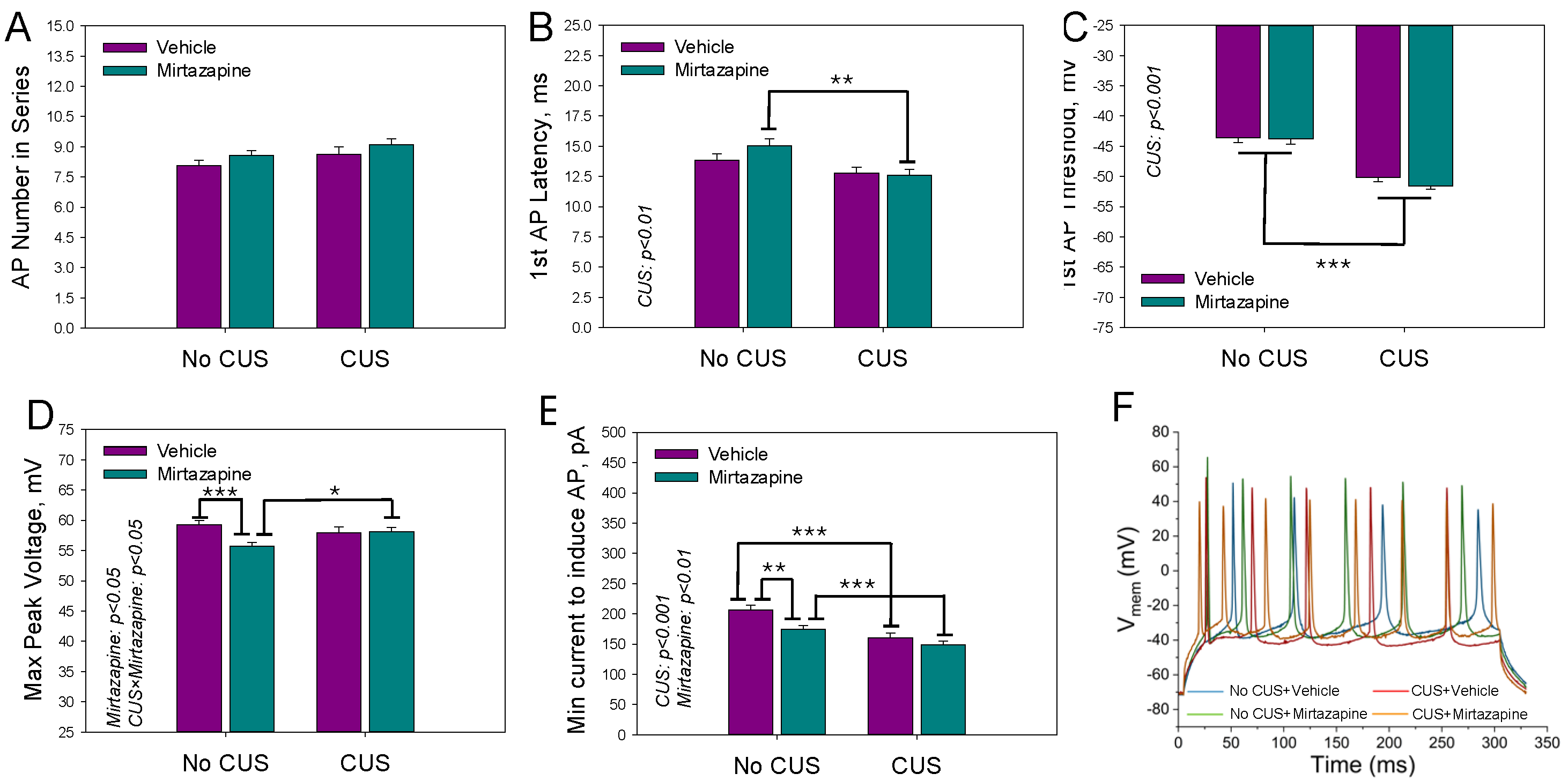
Figure 6 shows the mean number of action potentials within the depolarization current-induced series (A), latency time (B) and threshold voltage (C) of the first AP, maximal peak voltage within the series of APs (D), and minimal current needed to induce the series (E).
We observed a significant effect of maternal stress on the latency time (F1,195 = 9.73, p < 0.002; two-way ANOVA) and threshold voltage (F1,195 = 79.40, p < 0.001; two-way ANOVA) of the first AP and the minimal current needed to induce the series (F1,195 = 25.42, p < 0.001; two-way ANOVA), a significant effect of maternal mirtazapine on the peak voltage within the series of APs (F1,195 = 4.49, p < 0.04; two-way ANOVA) and the minimal current needed to induce the series (F1,195 = 9.06, p < 0.003; two-way ANOVA), and significant interactions between these factors with respect to the peak voltage (F1,195 = 5.47, p < 0.02; two-way ANOVA).
The effects of maternal stress, mirtazapine, and their combination on the percentage of neurons in hippocampal slices with AP series observed during depolarizing current pulses with amplitudes between 100 and 400 pA are shown above in
Figure 4B. A significant effect of maternal stress was detected during depolarizing pulses of 250 (F
1,18 = 5.79,
p = 0.03), 300 (F
1,18 = 26.30,
p < 0.001), 350 (F
1,18 = 21.86,
p < 0.001), and 400 pA (F
1,18 = 8.53,
p = 0.01).
2.3. Spontaneous Activity of Pyramidal Neurons in Hippocampal Slices Isolated from 11–13-Day-Old Offspring
The spontaneous activity of hippocampal neurons was measured only in the slices. On PNDs 11–13, of all the measured cells, 25% of the neurons in the offspring of nonstressed vehicle-treated dams, 22% of the neurons in the nonstressed mirtazapine-treated group, 35% of the neurons in the CUS-exposed vehicle-treated group, and up to 54% of the neurons in the CUS-exposed mirtazapine-treated group manifested spontaneous activity. The parameters of spontaneous activity, i.e., the average number of fired action potentials, the mean instantaneous frequency, and the firing frequency of action potentials, were evaluated, and no significant differences were found.
3. Discussion
The effects of depression on offspring development and how antidepressant treatment during pregnancy influences such changes are still open questions. Animal models offer the possibility to precisely control the pregestational and gestational states of the mother and pharmacotherapy. The offspring of treated mothers can be monitored under defined conditions for a longer period of time. Moreover, it is possible to combine behavioral, molecular, and electrophysiological analyses in the same settings [
27].
In this study, we used a well-established pregestational chronic unpredictable stress (CUS) rat model of maternal depression [
21,
29]. As a tested antidepressant, a nonselective alpha 2-adrenoceptor antagonist, mirtazapine, which has antidepressant activity in major depression, was used. In this model, we recently reported behavioral changes in offspring at juvenile and adolescent ages [
21].
CUS and/or mirtazapine significantly affected several parameters of hippocampal excitability in cultured primary neurons, whereas another set of parameters was not changed. CUS suppresses the excitability, as manifested in hyperpolarized resting membrane potential, increased the voltage threshold for an action potential (AP), and increased the minimal depolarizing current pulse amplitude for activation of both single AP and AP series. Similar effects were caused by mirtazapine when it was administered to dams not exposed to CUS: the resting membrane potential was hyperpolarized, the AP threshold and latency were increased, the minimal depolarizing current pulse amplitude for the activation of a single AP was increased, and the AP rise time was prolonged. Mirtazapine administered to CUS-exposed dams had a positive effect on excitability in selected CUS-affected parameters: it lowered the voltage threshold for the generation of a single AP and AP series and lowered the minimal depolarizing current pulse amplitude for the activation of a single AP. Taken together, the administration of mirtazapine to control dams had minor effects on the hippocampal excitability of offspring. Pregestational CUS, however, suppressed the hippocampal excitability of offspring to a moderate extent. The treatment of CUS-exposed dams with mirtazapine partly compensated for the observed changes in hippocampal excitability.
Approximately two weeks of maturation of the hippocampus under native conditions resulted in a different pattern of excitability alterations observed in acute slices. CUS mostly facilitates excitability by lowering both the threshold for AP activation and the minimal current amplitude needed to activate a single AP and AP series, shortening the AP latency and accelerating the AP rise time. The negative effects of mirtazapine administration to healthy control offspring almost disappeared during the first weeks of postnatal development. It was restricted to lowering the minimal current amplitude needed to activate a single AP and AP series (facilitation of excitability) and lowering the AP peak voltage in the AP series (negative effect). Mirtazapine administered to CUS-exposed dams facilitated the excitability of offspring in a majority of the analyzed parameters. The effect of CUS was opposite to that observed in primary cultures but remained minor to moderate. Mirtazapine treatment did not compensate for these changes.
While numerous studies have analyzed the behavioral and/or metabolic effects of maternal stress on offspring, alterations in neuronal excitability measured at the level of individual neurons are less frequently investigated. Grigoryan and Segal [
30] analyzed the excitability of hippocampal neurons in a primary culture established from the offspring of dams exposed to stress during gestation on the day of birth. They reported no differences in the resting membrane potential, input resistance, or AP threshold voltage between the stressed and nonstressed groups. Nevertheless, they reported reduced GABAergic inhibition, which resulted in a greater frequency of network activity in cultured neurons [
30]. In the prenatally stressed group, a reduction in inhibitory tone was also confirmed in hippocampal slices of 2–3-week-old rats from the same cohort. These findings support our findings that maternal stress may facilitate the hippocampal excitability of offspring in early life, whereas they contrast with our findings in primary cultures. This difference may arise from a different stress paradigm and the time of its application. The stress paradigm applied during gestation might have had a stronger effect at the time of birth. Another study in primary hippocampal culture [
27] reported a negative effect of pregestational stress on AP generation, which aligns with the findings of this study. Furthermore, maternal CUS decreased the expression of 5-HT and serotonin transporters. Moreover, it increased the 5-HT level in the hippocampi of 20-day-old embryos [
31], which is also in line with the decreased excitability of hippocampal neurons just after birth observed in this work.
Experiments on acute slices reflect not only two weeks of neuronal development in a native environment but also altered developmental interactions with mothers and siblings. Indeed, the results we obtained from the slices differ from those obtained from the primary cultures. The effect of the administration of mirtazapine to nonstressed dams was markedly reduced in slices and restricted to only two parameters. The significant change in the resting membrane potential observed in cultures disappeared, and several parameters of excitability were increased in offspring from CUS-exposed mothers. These results parallel those of several other studies in which a pregestational maternal stress model was used. The antidepressant venlafaxine decreased the number of mature hippocampal neurons in juvenile (31 days) offspring of the control but not CUS dams, and this effect disappeared in adolescent (47 days) and adult (75 days) offspring [
32]. In adult male rats, pregestational maternal stress reduces neurogenesis in hippocampal granule cells, and perinatal treatment with another antidepressant, fluoxetine, reverses this effect [
33].
Pregestational stress decreased the metabolism of the inhibitory neurotransmitter GABA in the hippocampus of 30-day-old offspring, as documented by MRI scans [
34], in line with a model used by Grigoryan and Segal [
30] and in line with the increased excitability observed in our model in slices. The same results were obtained in our model by Makova et al. [
35].
The effects of mirtazapine reported in this study may also be partly explained by its action as an antagonist of 5-HT
2 and 5-HT
3 receptors. While the activation of 5-HT
2, 5-HT
3, or 5-HT
4 receptors results in the depolarization of the membrane potential and increased neuronal excitability [
36,
37,
38], the activation of 5-HT
1A receptors causes hyperpolarization and decreased excitability [
36,
39,
40].
In the present study, we observed spontaneous activity in hippocampal neurons from all four offspring groups, with no significant differences in quantitative parameters of AP firing between the groups. However, the percentage of spontaneously active neurons was increased in CUS group. In our earlier work [
27], we reported increased levels of spontaneous activity in neurons from primary hippocampal cultures in the stressed group. Our findings can be interpreted in terms of altered GABAergic and glutamatergic systems, which enhance spontaneous action potential firing [
8].
The primary limitation of the present study is the fact that the offspring in each group (those of nonstressed vehicle treated dams, nonstressed mirtazapine treated dams, stressed vehicle treated dams, and stressed mirtazapine treated dams) were assessed as a whole, without distinguishing between males and females.
4. Materials and Methods
4.1. Animals
Female and male Wistar rats (initial weight 200–220 g; 2–3 months old) were obtained from the Department of Toxicology and Laboratory Animals Breeding, Centre of Experimental Medicine of the Slovak Academy of Sciences, Dobra Voda, Slovak Republic. All the animals were housed (38 × 59 × 25 cm3 large cages) under standard laboratory conditions (temperature: 22 ± 2 °C, humidity: 55 ± 10%) with a 12 h light/12 h dark cycle (lights on at 7.00 a.m.). Pelleted food and tap water were available ad libitum. The State Veterinary and Food Administration of the Slovak Republic approved all the experimental procedures (Approval No. 3334-5/2020-220 from 11 December 2020). The rats were handled according to the Guide for the Care and Use of Laboratory Animals (N.R.C., 1996) and the European Communities Council Directive of September 22, 2010 (2010/63/EU, 74). In accordance with the Directive, the animal environment was enriched, e.g., by providing hiding places and tissue paper for exploration and nest building.
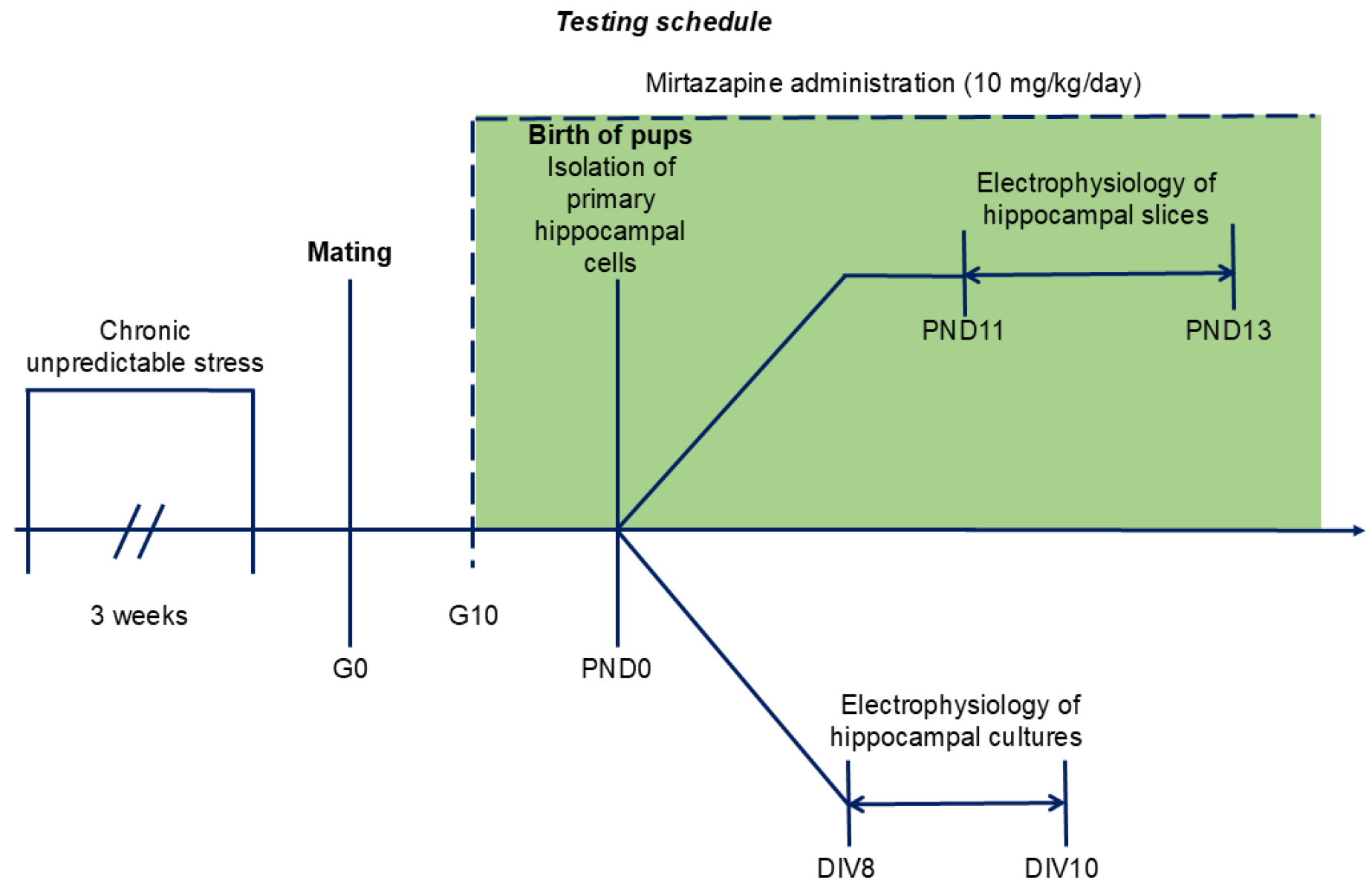
4.2. Experimental Design
The timing of the experiments is shown in
Figure 7.
4.3. Chronic Unpredictable Stress Schedule
After one week of adaptation, the female rats (n = 20) were randomly allocated to one of the following two groups: no chronic unpredictable stress (NO CUS; n = 10) and chronic unpredictable stress (CUS; n = 10). Animals from the stressed group were randomly exposed to 6 distinct stressors for three consecutive weeks: (1) exposure to damp bedding (12 h); (2) food deprivation (24 h); (3) predator stress–clothing with cat odor (10 h); (4) water deprivation (12 h); (5) cage decline at a 45° angle (6 h); and (6) Strobe light flicker lights (12 h). After the stress procedure, female rats from both groups were mated with males (3:1). At gestational day 0 (G0), each pregnant rat was housed separately. The G0 day was detected by the presence of sperm in vaginal smears.
4.4. Mirtazapine Treatment
On the tenth day of gestation, the dams from each group (NO CUS and CUS) were randomly divided into two sub-groups: vehicle (VEH) and mirtazapine (MIR), with five animals in each sub-group: NO CUS+VEH, NO CUS+MIR, CUS+VEH, and CUS+MIR. Mirtazapine (Mikrochem Trade s.r.o.; M = 265.36 g/mol) was dissolved in citric acid and administered orally via a wafer biscuit (size of 1 cm
3) to pregnant rats at a clinically relevant dose of 10 mg/kg/day (
Figure 7). The dose used in this experiment was chosen on the basis of previous results [
21]. The female rats in the control groups received a 1 cm
3 wafer biscuit once daily, which was filled with vehicle.
4.5. Primary Hippocampal Cell Culture
Primary hippocampal cultures were prepared on PND 0 according to the methods of Reichova et al. [
41]. For each primary culture preparation, three newborn pups were randomly selected from each litter of each dam from the groups NO CUS+VEH, NO CUS+MIR, CUS+VEH, and CUS+MIR. The animals were euthanized by cutting off the head using a special guillotine designed for killing laboratory rodents. The brains were dissected in ice-cold Hank’s balanced salt solution (HBSS; Gibco, Jenks, OK, USA) supplemented with 1% antibiotic-antimycotic (Sigma–Aldrich, Darmstadt, Germany) and 0.3 M HEPES (Sigma–Aldrich, Darmstadt, Germany) under a stereomicroscope (Optika Microscopes, Ponteranica, Italy). Hippocampal tissues were dissociated for 20 min at 37 °C with an enzymatic solution containing HBSS, 0.1% trypsin (Gibco, Jenks, OK, USA), and 0.1 mg/mL DNAse I (Roche, Mannheim, Germany). The cells were subsequently resuspended and plated on 24-well plates containing coverslips (Sarstedt, Nümbrecht, Germany) coated with 10 µg/mL poly-D-lysine (Sigma–Aldrich, Darmstadt, Germany) at a density of 0.8–1 × 10
5 cells/mL in RPMI medium (Gibco, Jenks, OK, USA) containing 10% fetal bovine serum (Sigma–Aldrich, Darmstadt, Germany). After 3 h of cell culture at 37 °C in a 5% CO
2 incubator, the medium was replaced with selective growth medium (Neurobasal A; Gibco, Jenks, OK, USA) supplemented with 1% antibiotic–antimycotic (Sigma–Aldrich, Darmstadt, Germany), 1% glutamine (Sigma–Aldrich, Darmstadt, Germany), and 2% B27 supplement (Gibco, Jenks, OK, USA). This was assigned as day 1 in vitro (DIV 1). At DIV5, 50% of the fresh growth medium was replaced. Cultured hippocampal neurons with pyramidal-like morphology were used for electrophysiology experiments at DIV 8–10. The excitability of at least 30 neurons were recorded from each culture. The exact numbers of the neurons/recordings from the hippocampal cultures are provided in the legends to the figures.
4.6. Acute Hippocampal Slices
Acute hippocampal slices were prepared on PNDs 11–13. Three were randomly selected from each litter of each dam from the groups NO CUS+VEH, NO CUS+MIR, CUS+VEH, and CUS+MIR. The animals were euthanized by cutting off the head using a special guillotine designed for killing laboratory rodents. The brains were cut on a vibratome (Campden Instruments, Loughborough, England) into 300 µm thick slices in ice-cold artificial cerebrospinal fluid (ACSF) containing the following (in mM): 125 NaCl, 3 KCl, 25 NaHCO3, 1.25 NaH2PO4, 2 CaCl2, 1 MgCl2, and 10 D-glucose (all chemicals were purchased from Sigma–Aldrich, Darmstadt, Germany). ACSF was oxygenated with 95% O2 and 5% CO2, pH 7.4. Before recording, the slices were incubated for 60 min at 37 °C and then for 30 min at room temperature in oxygenated ACSF solution. The exact numbers of the neurons/recordings from the hippocampal slices are provided in the legends to the figures.
4.7. Electrophysiological Experiments
All electrophysiology measurements were performed via the whole-cell current clamp configuration of the patch-clamp technique. The signals were recorded via a HEKA EPC10 amplifier (HEKA Electronics, Reutlingen, Germany) controlled by Patchmaster v2x73.3 (HEKA Electronics, Reutlingen, Germany) software. The data were analyzed via Fitmaster v2x73.3 (HEKA Electronics, Reutlingen, Germany) and Origin 2022 Academic (OriginLab Co., Northampton, MA, USA) software. The parameters of an action potential (AP) series were extracted from the experimental traces with custom code via the open-source Electrophysiological Feature Extraction Library (eFEL,
https://github.com/BlueBrain/eFEL, accessed on 25 March 2025).
Patch electrodes were pulled from borosilicate glass with 3–4 MΩ resistance. The extracellular solution in primary hippocampal cultures contained the following (in mM): 130 NaCl; 3 KCl; 10 HEPES; 2 CaCl2; 1 MgCl2; 10 D-glucose; pH 7.4 (with NaOH), and the intracellular solution contained the following (in mM): 120 K-gluconate; 20 KCl; 2 MgCl2; 2 Na2ATP; 0.25 Na2GTP; 10 HEPES; pH 7.4 (with KOH) (all chemicals from Sigma–Aldrich, Darmstadt, Germany). The extracellular solution for acute neuronal slices was ASCF solution oxygenated with 95% O2 and 5% CO2, pH 7.4, and the intracellular solution was the same as that for the primary hippocampal cultures. The osmolarity of an intracellular solution was approximately 290–300 mOsm/L (Osmomat 030-Gonotec, Berlin, Germany). The osmolarity of an extracellular solution was adjusted by adding sucrose to a final osmolarity, which was 2–3 mOsm/L lower than the osmolarity of the corresponding intracellular solution.
The actual resting membrane potential (Vrest) was read immediately after establishing the whole-cell configuration and switching to current-clamp mode. During all other experiments, the resting membrane potential was held at −70 mV. The input resistance (Rinp) was measured by a series of 250 ms long hyperpolarizing current pulses with the amplitude decreasing from −70 mV with a step of −25 pA. The membrane potential at the end of each current pulse was read and the Rinp value was calculated from the fit of the voltage–current relationship by the Ohm equation. Single APs were activated by a series of 5 ms long depolarizing current pulses with the amplitude increasing with a step of +50 pA. A series of APs were activated by a series of 300 ms long depolarizing current pulses with the amplitude increasing with a step of +50 pA. Spontaneous activity was measured for 5 min from each neuron’s resting membrane potential.
4.8. Statistical Analyses
The statistical analyses were performed via GraphPad Prism 8.3 software. Individual recordings from individual neurons were used as data points for the statistical analysis. Outliers were identified and excluded via GraphPad PRISM’s ROUT method (Q = 1%). Neurons, considered as an outlet according to one parameter, were excluded from the assessment of all other characteristics. The data were analyzed by a two-way analysis of variance (ANOVA) with treatment (mirtazapine) and conditions (stress vs. nonstress) as the main factors, followed by the Bonferroni post hoc test. The results are expressed as the mean ± standard error of the mean (SEM), and a value of p ≤ 0.05 was considered statistically significant.


 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}