
Caesalpinia sappan L. Ameliorates Scopolamine-Induced Memory Deficits in Mice via the cAMP/PKA/CREB/BDNF Pathway



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Chemicals and Reagents
2.3. Drugs
2.4. Experimental Animals
2.5. CSW Extract Preparation
2.6. TLC-Densitometry Analysis
2.7. Test Solution Preparation
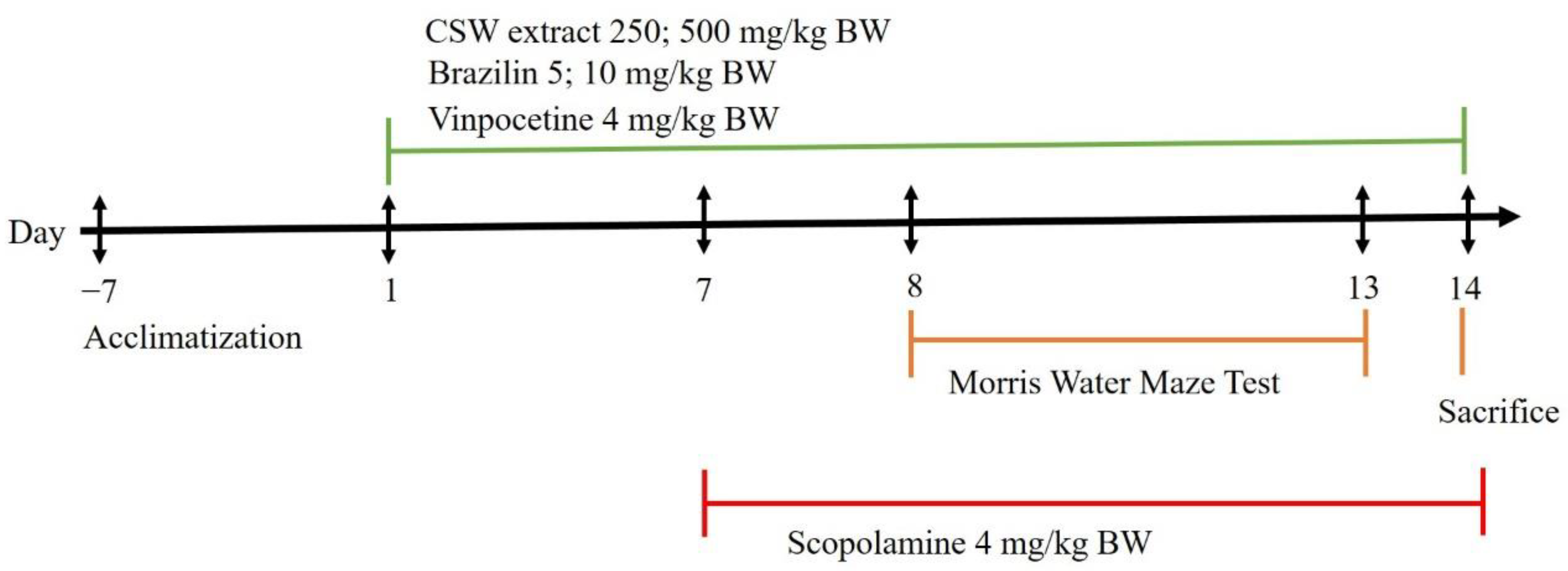
2.8. Experimental Design
2.9. Morris Water Maze Test
2.10. Hippocampus Collection
2.11. cAMP Level Analysis
2.12. PKA Activity Analysis
2.13. Protein Isolation
2.14. Western Blot Analysis
2.15. Statistical Analysis
3. Results
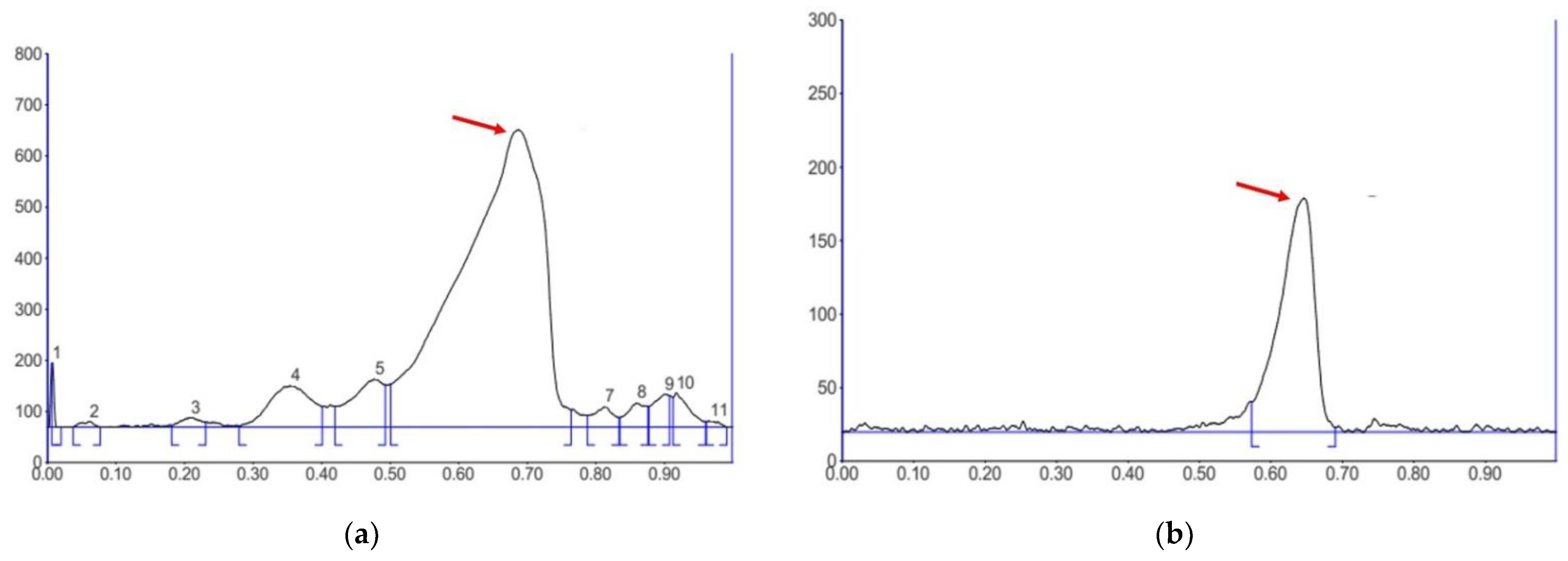
3.1. Determination of Brazilin Content in the CSW Extract
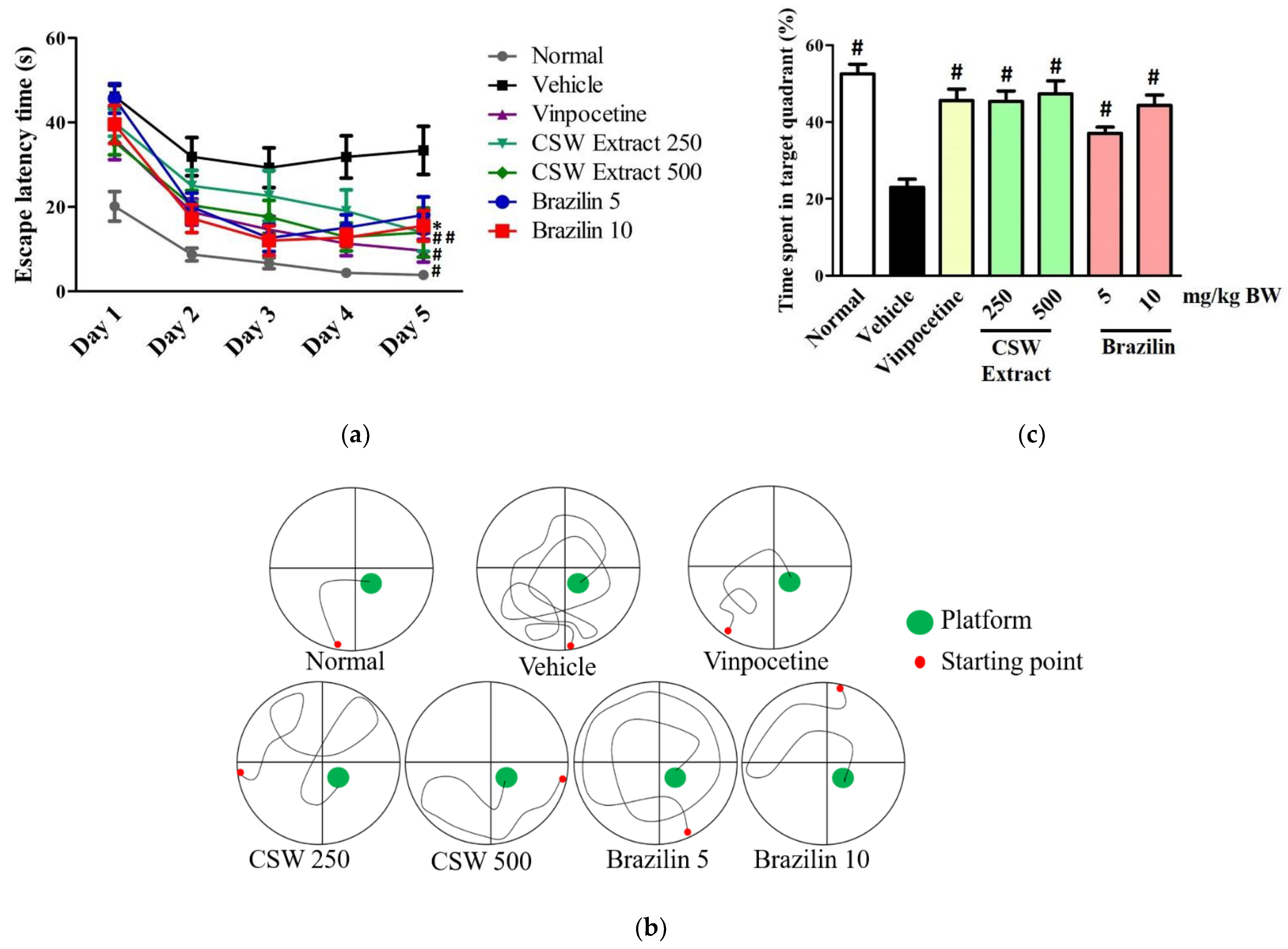
3.2. CSW Extract and Brazilin Improved Spatial Memory in the MWM Test
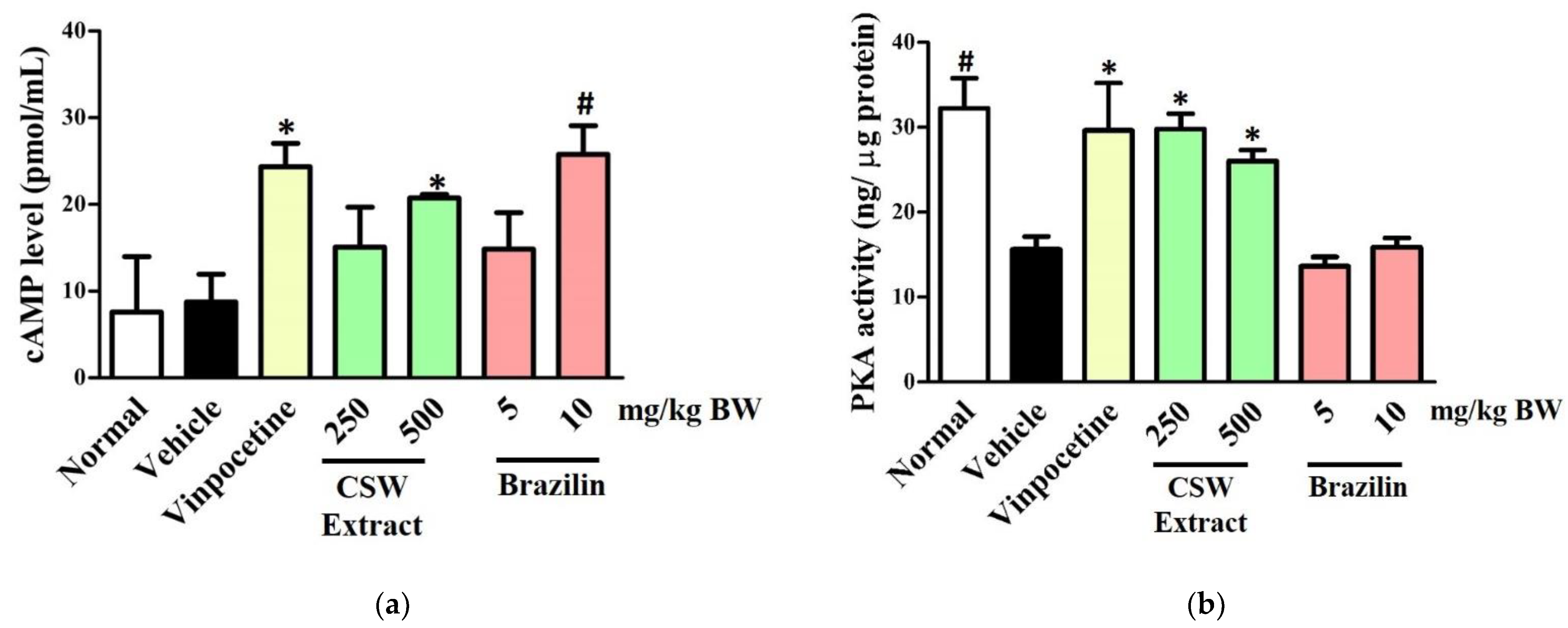
3.3. CSW Extract and Brazilin Increased cAMP Levels and PKA Activity in the Hippocampus
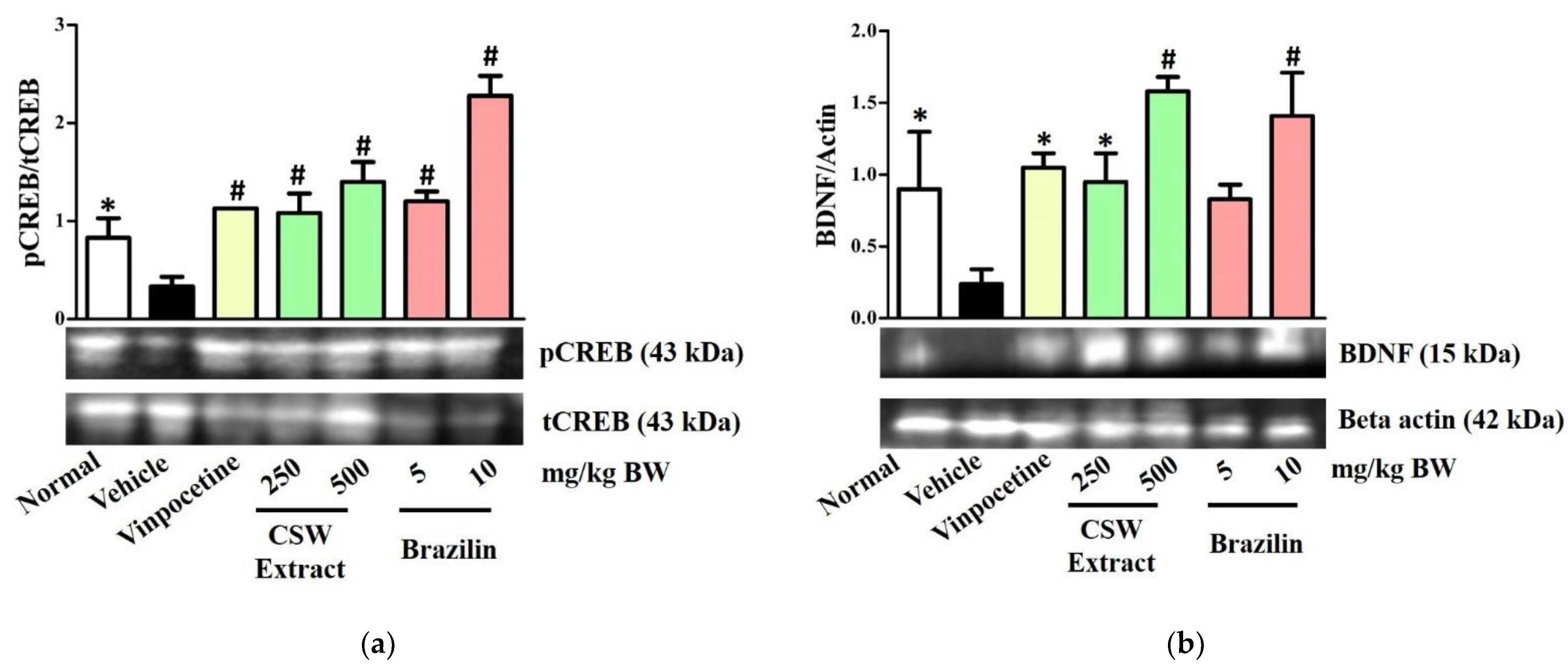
3.4. CSW Extract and Brazilin Increased the Protein Expression Level of pCREB and BDNF in the Hippocampus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rizzi, L.; Rosset, I.; Roriz-cruz, M. Global Epidemiology of Dementia: Alzheimer’s and Vascular Types. BioMed Res. Int. 2014, 2014, 908915. [Google Scholar] [CrossRef]
- Richardson, C.; Stephan, B.C.M.; Robinson, L.; Brayne, C.; Matthews, F.E. Two-Decade Change in Prevalence of Cognitive Impairment in the UK. Eur. J. Epidemiol. 2019, 34, 1085–1092. [Google Scholar] [CrossRef]
- Li, W.; Sun, L.; Xiao, S. Prevalence, Incidence, Influence Factors and Cognitive Characteristics of Amnestic Mild Cognitive Impairment Among Older Adult: A 1-Year Follow-Up Study in China. Front. Psychiatry 2020, 11, 75. [Google Scholar] [CrossRef]
- Deture, M.A.; Dickson, D.W. The Neuropathological Diagnosis of Alzheimer’s Disease. Mol. Neurodegener. 2019, 14, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Xie, N.; Tang, B.; Li, R.; Shen, Y. Alzheimer’s Disease: From Genetic Variants to the Distinct Pathological Mechanisms. Front. Mol. Neurosci. 2017, 10, 319. [Google Scholar] [CrossRef]
- Kent, S.A.; Spires, T.L.; Claire, J. The Physiological Roles of Tau and Aβ: Implications for Alzheimer’s Disease Pathology and Therapeutics. Acta Neuropathol. 2020, 140, 417–447. [Google Scholar] [CrossRef]
- Sigurdsson, T.; Duvarci, S. Hippocampal-Prefrontal Interactions in Cognition, Behavior and Psychiatric Disease. Front. Syst. Neurosci. 2016, 9, 190. [Google Scholar] [CrossRef]
- Jin, J.; Maren, S. Prefrontal-Hippocampal Interactions in Memory and Emotion. Front. Syst. Neurosci. 2015, 9, 170. [Google Scholar] [CrossRef] [PubMed]
- Stanciu, G.D.; Luca, A.; Rusu, R.N.; Bild, V.; Chiriac, S.I.B.; Solcan, C.; Bild, W.; Ababei, D.C. Alzheimer’s Disease Pharmacotherapy in Relation to Cholinergic System Involvement. Biomolecules 2020, 10, 40. [Google Scholar] [CrossRef] [PubMed]
- Moss, D.E. Improving Anti-Neurodegenerative Benefits of Acetylcholinesterase Inhibitors in Alzheimer’s Disease: Are Irreversible Inhibitors the Future? Int. J. Mol. Sci. 2020, 21, 3438. [Google Scholar] [CrossRef]
- Mehta, M.; Adem, A.; Sabbagh, M. New Acetylcholinesterase Inhibitors for Alzheimer’s Disease. Int. J. Alzheimer’s Dis. 2012, 2012, 728983. [Google Scholar] [CrossRef]
- Mufson, E.J.; Counts, S.E.; Perez, S.E.; Ginsberg, S.D. Cholinergic System During the Progression of Alzheimer’s Disease: Therapeutic Implications. Expert Rev. Neurother. 2009, 8, 1703–1718. [Google Scholar] [CrossRef]
- Wu, Y.; Li, Z.; Huang, Y.; Wu, D.; Luo, H. Novel Phosphodiesterase Inhibitors for Cognitive Improvement in Alzheimer’s Disease. J. Med. Chem. 2018, 61, 5467–5483. [Google Scholar] [CrossRef]
- Conti, M.; Richter, W. Phosphodiesterases and Cyclic Nucleotide Signaling in the CNS. In Cyclic Nucleotide Phosphodiesterases in the Central Nervous System; Brandon, J., West, N.J., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2014; pp. 1–46. [Google Scholar]
- Kumar, A.; Sharma, V.; Pal, V.; Kaundal, M. Herbs to Curb Cyclic Nucleotide Phosphodiesterase and Their Potential Role in Alzheimer’s Disease. Mech. Ageing Dev. 2015, 149, 75–87. [Google Scholar] [CrossRef]
- Rahimi, R.; Ghiasi, S.; Azimi, H.; Fakhari, S.; Abdollahi, M. Cytokine A Review of the Herbal Phosphodiesterase Inhibitors: Future Perspective of New Drugs. Cytokine 2010, 49, 123–129. [Google Scholar] [CrossRef]
- Froestl, W.; Muhs, A.; Pfeifer, A. Cognitive Enhancers (Nootropics) Part 2: Drugs Interacting with Enzymes Update 2014. J. Alzheimer Dis. 2014, 42, 1–68. [Google Scholar] [CrossRef]
- Rina, O.; Ibrahim, S.; Dharma, A.; Afrizal, W.C.U.; Widodo, Y.R. Stabilities Natural Colorant of Sappan Wood (Caesalpinia sappan L.) for Food and Beverages in Various pH, Temperature and Matrices of Food. Int. J. ChemTech Res. 2017, 10, 98–103. [Google Scholar]
- Rahmawati, F. Kajian Potensi “Wedang Uwuh” Sebagai Minuman Fungsional. In Proceedings of the Seminar Wonderfull Indonesia, Yogyakarta, Indonesia, 3 December 2011; pp. 619–631. [Google Scholar]
- Helmi, H.; Fakhrudin, N.; Nurrochmad, A.; Sudarmanto, A.; Ikawati, Z. Caesalpinia sappan L. Wood Is A Potential Source of Natural Phosphodiesterase-1 Inhibitors. Pharmacogn. J. 2020, 12, 1206–1217. [Google Scholar]
- Helmi, H.; Fakhrudin, N.; Nurrochmad, A.; Sudarmanto, B.S.A.; Ikawati, Z. In Vitro and In Silico Studies of Secang Wood (Caesalpinia sappan L.) Extracts and Brazilin as Natural Phosphodiesterase-1 (PDE1) Inhibitor for Herbal Cognitive Enhancer Development. Res. J. Pharm. Technol. 2020, 13, 2269–2274. [Google Scholar] [CrossRef]
- Yuliani, S.; Partadiredja, G. Turmeric (Curcuma longa L.) Extract May Prevent the Deterioration of Spatial Memory and the Deficit of Estimated Total Number of Hippocampal Pyramidal Cells of Trimethyltin-Exposed Rats. Drug Chem. Toxicol. 2018, 41, 62–71. [Google Scholar]
- Tabassum, N.; Rasool, S.; Malik, Z.A.; Ahmad, F. Natural Cognitive Enhancers. J. Pharm. Res. 2012, 5, 153–160. [Google Scholar]
- Krishna, G.; Muralidhara, M. Dietary Supplements as Cognitive Enhancers the Role of Flavonoid-Rich Foods and Their Relevance in Age-Related Neurodegeneration. In Bioactive Nutraceuticals and Dietery Supplements in Neurological and Brain Disease: Prevention and Therapy, 1st ed.; Watson, R.R., Preedy, V.R., Eds.; Academic Press Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Nirmal, N.P.; Panichayupakaranant, P. Antioxidant, Antibacterial and Anti-inflammatory Activities of Standardized Brazilin-Rich Caesalpinia sappan Extract. Pharm. Biol. 2015, 53, 1339–1343. [Google Scholar]
- Nirmal, N.P.; Rajput, M.S.; Prasad, R.G.S.V.; Ahmad, M. Brazilin from Caesalpinia sappan Heartwood and its Pharmacological Activities: A review. Asian Pac. J. Trop. Med. 2015, 8, 421–430. [Google Scholar] [CrossRef]
- Omori, K.; Kotera, J. Overview of PDEs and Their Regulation. Circl. Res. 2007, 100, 309–327. [Google Scholar] [CrossRef] [PubMed]
- Medina, A.E. Therapeutic Utility of Phosphodiesterase Type I Inhibitors in Neurological Conditions. Front. Neurosci. 2011, 5, 2007–2011. [Google Scholar] [CrossRef] [PubMed]
- Mccabe, D.P.; Mcdaniel, M.A.; Hambrick, D.Z. The Relationship Between Working Memory Capacity and Executive Functioning: Evidence for A Common Executive Attention Construct. Neuropsychology 2011, 24, 222–243. [Google Scholar] [CrossRef] [PubMed]
- Sapiurka, M.E. Spatial Navigation and Memory: The Role of the Hippocampus and Neocortical Structures. Ph.D. Thesis, Unniversity of California, San Diego, CA, USA, 2016. [Google Scholar]
- Negrón-oyarzo, I.; Espinoza, N.; Aguilar-Rivera, M.; Fuenzalida, M.; Aboitiz, F.; Fuentealba, P. Coordinated Prefrontal–Hippocampal Activity and Navigation Strategy-Related Prefrontal Firing during Spatial Memory Formation. Proc. Natl. Acad. Sci. USA 2018, 115, 7123–7128. [Google Scholar] [CrossRef]
- Alberini, C.M. Transcription Factor in Long-Term Memory and Synaptic Plasticity. Phys. Rev. 2014, 89, 121–145. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, A.; Giralt, A.; Arumí, H.; Alberch, J.; Pérez-navarro, E. Regulation of Hippocampal cGMP Levels as a Candidate to Treat Cognitive Deficits in Huntington’s Disease. PLoS ONE. 2013, 8, e73664. [Google Scholar] [CrossRef]
- Keravis, T.; Lugnier, C. Cyclic Nucleotide Phosphodiesterase (PDE) Isozymes as Targets of the Intracellular Signaling Network: Benefits of PDE Inhibitors in Various Diseases and Perspectives for Future. Br. J. Pharmacol. 2012, 165, 1288–1305. [Google Scholar] [CrossRef]
- Balmus, I.M.; Ciobica, A. Main Plant Extracts’ Active Properties Effective on Scopolamine-Induced Memory Loss. Am. J. Alzheimers. Dis. Other Demen. 2017, 32, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Prashar, Y.; Gill, N.S.; Kakkar, S. A Review on Medicinal Plants Affecting Amnesia on Scopolamine Induced Model. Pharmatutor Mag. 2014, 2, 20–28. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Helmi, H.; Fakhrudin, N.; Nurrochmad, A.; Ikawati, Z. Caesalpinia sappan L. Ameliorates Scopolamine-Induced Memory Deficits in Mice via the cAMP/PKA/CREB/BDNF Pathway. Sci. Pharm. 2021, 89, 29. https://doi.org/10.3390/scipharm89020029
Helmi H, Fakhrudin N, Nurrochmad A, Ikawati Z. Caesalpinia sappan L. Ameliorates Scopolamine-Induced Memory Deficits in Mice via the cAMP/PKA/CREB/BDNF Pathway. Scientia Pharmaceutica. 2021; 89(2):29. https://doi.org/10.3390/scipharm89020029
Chicago/Turabian StyleHelmi, Helmi, Nanang Fakhrudin, Arief Nurrochmad, and Zullies Ikawati. 2021. "Caesalpinia sappan L. Ameliorates Scopolamine-Induced Memory Deficits in Mice via the cAMP/PKA/CREB/BDNF Pathway" Scientia Pharmaceutica 89, no. 2: 29. https://doi.org/10.3390/scipharm89020029
APA StyleHelmi, H., Fakhrudin, N., Nurrochmad, A., & Ikawati, Z. (2021). Caesalpinia sappan L. Ameliorates Scopolamine-Induced Memory Deficits in Mice via the cAMP/PKA/CREB/BDNF Pathway. Scientia Pharmaceutica, 89(2), 29. https://doi.org/10.3390/scipharm89020029