
Implications of Glycosaminoglycans on Viral Zoonotic Diseases



Abstract
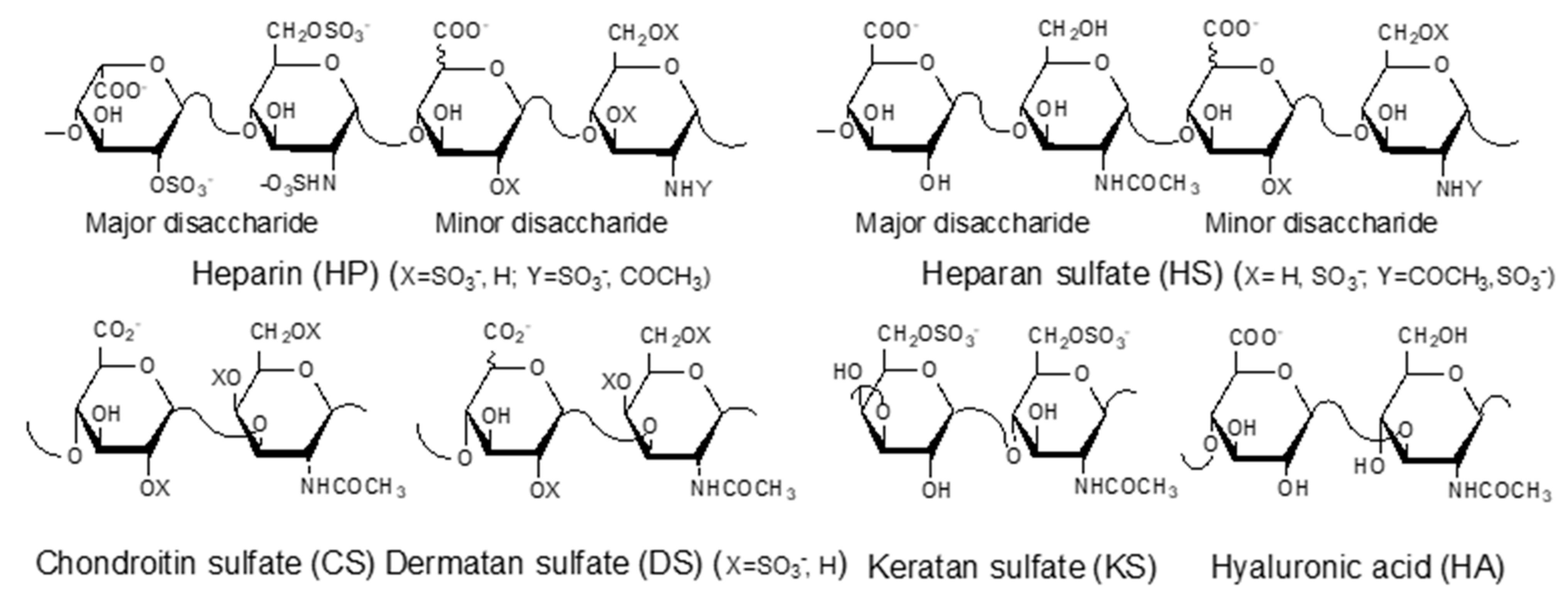
:1. Introduction
2. Zoonotic Diseases
2.1. Filovirus—Ebola
2.2. Henipavirus—Hendra and Nipah
2.3. Coronavirus—SARS, MERS, and COVID-19
2.4. Lentivirus—AIDS
2.5. Flaviviruses—Dengue, Encephalitis from Ticks, Japanese Encephalitis, Zika
2.6. Orthohepevirus—Hepatitis E
2.7. Lyssavirus—Rabies
3. Wet Markets and Bushmeat
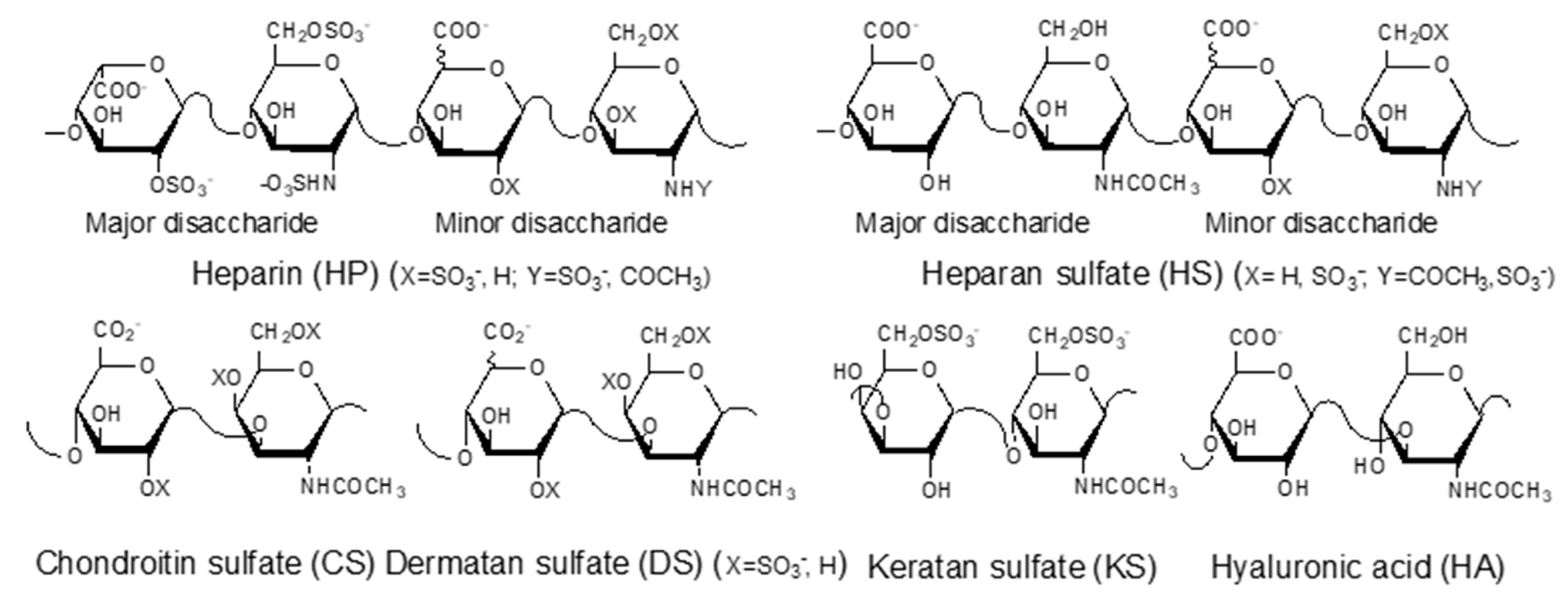
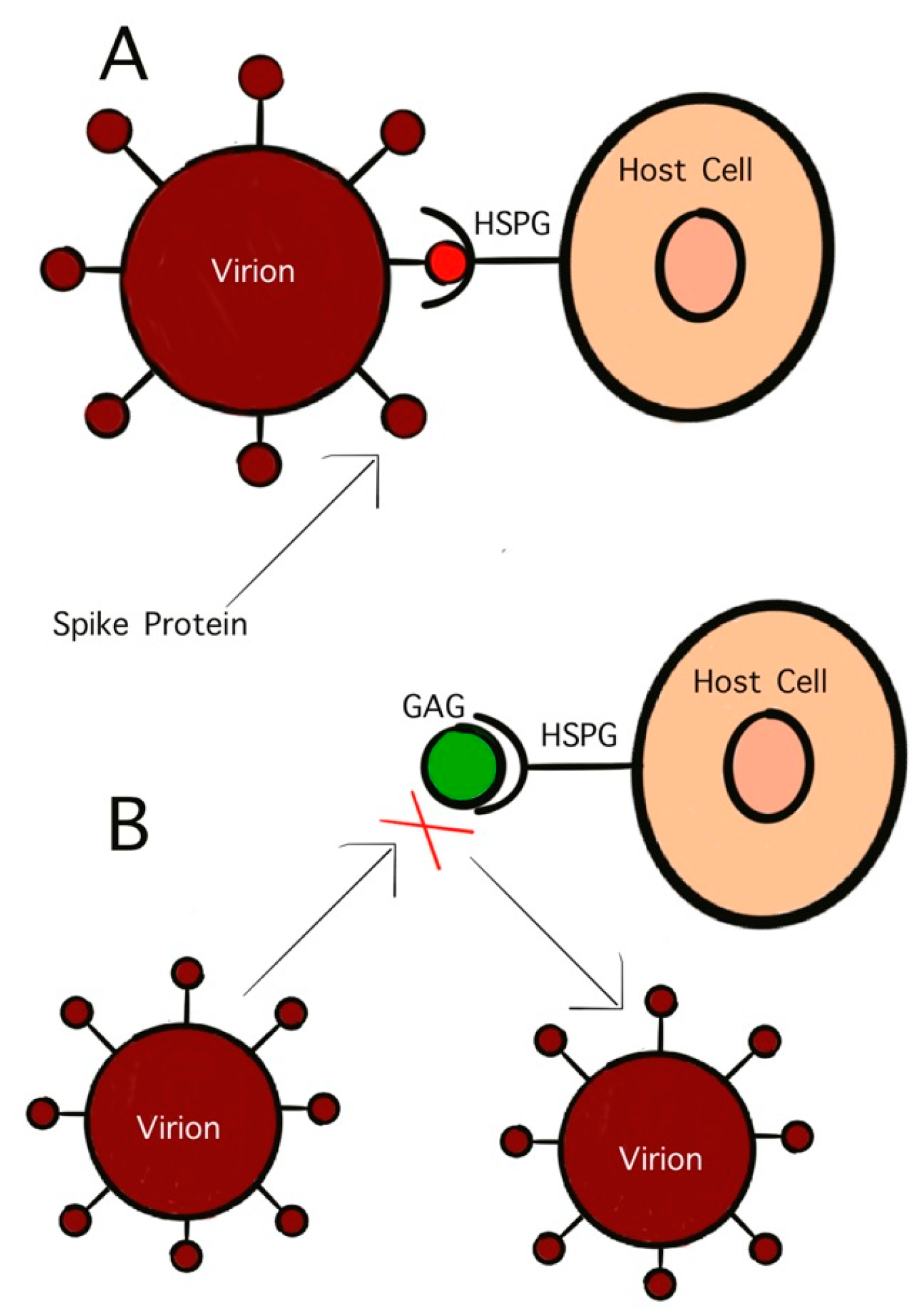
4. Roles of GAGs in Pathogenesis of Viral Zoonotic Diseases
4.1. Ebola
4.2. SARS, MERS, and COVID-19
4.3. AIDS
4.4. Dengue, Encephalitis from Ticks, West Nile Virus, Zika
4.5. Hepatitis E
4.6. Rabies
5. Potential Prophylactic and Therapeutic Applications of GAGs in the Treatments of Viral Zoonotic Diseases
6. Conclusions and Future Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Royce, K.; Fu, F. Mathematically Modeling Spillovers of an Emerging Infectious Zoonosis with an Intermediate Host. PLoS ONE 2020, 15, e0237780. [Google Scholar] [CrossRef]
- Beran, G.W. Handbook of Zoonoses, Section A: Bacterial, Rickettsial, Chalamydial, and Mycotic; Handbook of Zoonoses, Section B: Viral, 2nd ed.; CRC Press: Boca Raton, FL, USA, 1994. [Google Scholar]
- Lei, R.; Qiu, R. A Strategy to Prevent and Control Zoonoses? Hastings Cent. Rep. 2020, 50, 73–74. [Google Scholar] [CrossRef]
- Santana, C. COVID-19, Other Zoonotic Diseases and Wildlife Conservation. Hist. Philos. Life Sci. 2020, 42, 45. [Google Scholar] [CrossRef]
- Cai, Y.; Yu, S.; Chi, X.; Radoshitzky, S.R.; Kuhn, J.H.; Berger, E.A. An Immunotoxin Targeting Ebola Virus Glycoprotein Inhibits Ebola Virus Production from Infected Cells. PLoS ONE 2021, 16, e0245024. [Google Scholar] [CrossRef] [PubMed]
- Sompayrac, L.; Sompayrac, L. How Pathogenic Viruses Think: Making Sense of Virology, 2nd ed.; Jones & Bartlett Learning: Burlington, MA, USA, 2013; ISBN 978-1-4496-4580-9. [Google Scholar]
- Escudero-Pérez, B.; Ruibal, P.; Rottstegge, M.; Lüdtke, A.; Port, J.R.; Hartmann, K.; Gómez-Medina, S.; Müller-Guhl, J.; Nelson, E.V.; Krasemann, S.; et al. Comparative Pathogenesis of Ebola Virus and Reston Virus Infection in Humanized Mice. JCI Insight 2019, 4, e126070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williamson, K.M.; Wheeler, S.; Kerr, J.; Bennett, J.; Freeman, P.; Kohlhagen, J.; Peel, A.J.; Eby, P.; Merritt, T.; Housen, T.; et al. Hendra in the Hunter Valley. One Health 2020, 10, 100162. [Google Scholar] [CrossRef] [PubMed]
- Soman Pillai, V.; Krishna, G.; Valiya Veettil, M. Nipah Virus: Past Outbreaks and Future Containment. Viruses 2020, 12, 465. [Google Scholar] [CrossRef] [Green Version]
- Harmon, A. Middle East Respiratory Syndrome; Salem Press Encycl. Health: Hackensack, NJ, USA, 2020. [Google Scholar]
- Misra, S.; Hascall, V.C.; Markwald, R.R.; Ghatak, S. Interactions between Hyaluronan and Its Receptors (CD44, RHAMM) Regulate the Activities of Inflammation and Cancer. Front. Immunol. 2015, 6, 201. [Google Scholar] [CrossRef] [Green Version]
- Wadstrom, T.; Ljungh, A. Glycosaminoglycan-Binding Microbial Proteins in Tissue Adhesion and Invasion: Key Events in Microbial Pathogenicity. J. Med. Microbiol. 1999, 48, 223–233. [Google Scholar] [CrossRef]
- Kim, S.Y.; Jin, W.; Sood, A.; Montgomery, D.W.; Grant, O.C.; Fuster, M.M.; Fu, L.; Dordick, J.S.; Woods, R.J.; Zhang, F.; et al. Characterization of Heparin and Severe Acute Respiratory Syndrome-Related Coronavirus 2 (SARS-CoV-2) Spike Glycoprotein Binding Interactions. Antiviral Res. 2020, 181, 104873. [Google Scholar] [CrossRef]
- Clausen, T.M.; Sandoval, D.R.; Spliid, C.B.; Pihl, J.; Perrett, H.R.; Painter, C.D.; Narayanan, A.; Majowicz, S.A.; Kwong, E.M.; McVicar, R.N.; et al. SARS-CoV-2 Infection Depends on Cellular Heparan Sulfate and ACE2. Cell 2020, 183, 1043–1057. [Google Scholar] [CrossRef]
- Harrop, H.A.; Rider, C.C. Heparin and Its Derivatives Bind to HIV-1 Recombinant Envelope Glycoproteins, Rather than to Recombinant HIV-1 Receptor, CD4. Glycobiology 1998, 8, 131–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueiredo, L.T.M. Emergent Arboviruses in Brazil. Rev. Soc. Bras. Med. Trop. 2007, 40, 224–229. [Google Scholar] [CrossRef] [Green Version]
- O’Hearn, A.; Wang, M.; Cheng, H.; Lear-Rooney, C.M.; Koning, K.; Rumschlag-Booms, E.; Varhegyi, E.; Olinger, G.; Rong, L. Role of EXT1 and Glycosaminoglycans in the Early Stage of Filovirus Entry. J. Virol. 2015, 89, 5441–5449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozlovskaya, L.I.; Osolodkin, D.I.; Shevtsova, A.S.; Romanova, L.I.; Rogova, Y.V.; Dzhivanian, T.I.; Lyapustin, V.N.; Pivanova, G.P.; Gmyl, A.P.; Palyulin, V.A.; et al. GAG-Binding Variants of Tick-Borne Encephalitis Virus. Virology 2010, 398, 262–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathieu, C.; Dhondt, K.P.; Châlons, M.; Mély, S.; Raoul, H.; Negre, D.; Cosset, F.-L.; Gerlier, D.; Vivès, R.R.; Horvat, B. Heparan Sulfate-Dependent Enhancement of Henipavirus Infection. mBio 2015, 6, e02427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalia, M.; Chandra, V.; Rahman, S.A.; Sehgal, D.; Jameel, S. Heparan Sulfate Proteoglycans Are Required for Cellular Binding of the Hepatitis E Virus ORF2 Capsid Protein and for Viral Infection. J. Virol. 2009, 83, 12714–12724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinheiro, F.P.; Freitas, R.B.; Travassos da Rosa, J.F.; Gabbay, Y.B.; Mello, W.A.; LeDuc, J.W. An Outbreak of Mayaro Virus Disease in Belterra, Brazil. I. Clinical and Virological Findings. Am. J. Trop. Med. Hyg. 1981, 30, 674–681. [Google Scholar] [CrossRef]
- Sasaki, M.; Anindita, P.D.; Ito, N.; Sugiyama, M.; Carr, M.; Fukuhara, H.; Ose, T.; Maenaka, K.; Takada, A.; Hall, W.W.; et al. The Role of Heparan Sulfate Proteoglycans as an Attachment Factor for Rabies Virus Entry and Infection. J. Infect. Dis. 2018, 217, 1740–1749. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.; Hall, R.A.; Lobigs, M. Common E Protein Determinants for Attenuation of Glycosaminoglycan-Binding Variants of Japanese Encephalitis and West Nile Viruses. J. Virol. 2004, 78, 8271–8280. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Zhao, J.; Liu, X.; Fraser, K.; Lin, L.; Zhang, X.; Zhang, F.; Dordick, J.S.; Linhardt, R.J. Interaction of Zika Virus Envelope Protein with Glycosaminoglycans. Biochemistry 2017, 56, 1151–1162. [Google Scholar] [CrossRef]
- Menozzi, F.D.; Bischoff, R.; Fort, E.; Brennan, M.J.; Locht, C. Molecular Characterization of the Mycobacterial Heparin-Binding Hemagglutinin, a Mycobacterial Adhesin. Proc. Natl. Acad. Sci. USA 1998, 95, 12625–12630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castaneda-Roldan, E.I.; Avelino-Flores, F.; Dall’Agnol, M.; Freer, E.; Cedillo, L.; Dornand, J.; Giron, J.A. Adherence of Brucella to Human Epithelial Cells and Macrophages Is Mediated by Sialic Acid Residues. Cell. Microbiol. 2004, 6, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Freitag, C.M.; Strijbis, K.; van Putten, J.P.M. Host Cell Binding of the Flagellar Tip Protein of Campylobacter jejuni. Cell. Microbiol. 2017, 19, e12714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dabo, S.M.; Confer, A.W.; Anderson, B.E.; Gupta, S. Bartonella Henselae Pap31, an Extracellular Matrix Adhesin, Binds the Fibronectin Repeat III13 Module. Infect. Immun. 2006, 74, 2513–2521. [Google Scholar] [CrossRef] [Green Version]
- Ching, A.T.C.; Fávaro, R.D.; Lima, S.S.; Chaves, A.d.A.M.; de Lima, M.A.; Nader, H.B.; Abreu, P.A.E.; Ho, P.L. Lepstospira Interrogans Shotgun Phage Display Identified LigB as a Heparin-Binding Protein. Biochem. Biophys. Res. Commun. 2012, 427, 774–779. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Domínguez, C.; Vázquez-Boland, J.A.; Carrasco-Marín, E.; López-Mato, P.; Leyva-Cobián, F. Host Cell Heparan Sulfate Proteoglycans Mediate Attachment and Entry of Listeria Monocytogenes, and the Listerial Surface Protein ActA Is Involved in Heparan Sulfate Receptor Recognition. Infect. Immun. 1997, 65, 78–88. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-P.; Bhowmick, R.; Coburn, J.; Leong, J.M. Host Cell Heparan Sulfate Glycosaminoglycans Are Ligands for OspF-Related Proteins of the Lyme Disease Spirochete: GAG Binding by B. Burgdorferi OspF-Related Proteins. Cell. Microbiol. 2015, 17, 1464–1476. [Google Scholar] [CrossRef] [Green Version]
- Rostand, K.S.; Esko, J.D. Microbial Adherence to and Invasion through Proteoglycans. Infect. Immun. 1997, 65, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Harper, M.; Boyce, J.D.; Adler, B. Pasteurella Multocida Pathogenesis: 125 Years after Pasteur: Pasteurella Multocida Pathogenesis. FEMS Microbiol. Lett. 2006, 265, 1–10. [Google Scholar] [CrossRef]
- Yamashita, S.; Lukacik, P.; Barnard, T.J.; Noinaj, N.; Felek, S.; Tsang, T.M.; Krukonis, E.S.; Hinnebusch, B.J.; Buchanan, S.K. Structural Insights into Ail-Mediated Adhesion in Yersinia Pestis. Structure 2011, 19, 1672–1682. [Google Scholar] [CrossRef] [Green Version]
- Ludington, J.G.; Ward, H.D. The Cryptosporidium Parvum C-Type Lectin CpClec Mediates Infection of Intestinal Epithelial Cells via Interactions with Sulfated Proteoglycans. Infect. Immun. 2016, 84, 1593–1602. [Google Scholar] [CrossRef] [Green Version]
- Pathuri, P.; Nguyen, E.T.; Svärd, S.G.; Luecke, H. Apo and Calcium-Bound Crystal Structures of Alpha-11 Giardin, an Unusual Annexin from Giardia Lamblia. J. Mol. Biol. 2007, 368, 493–508. [Google Scholar] [CrossRef] [Green Version]
- Sinnis, P.; Coppi, A.; Toida, T.; Toyoda, H.; Kinoshita-Toyoda, A.; Xie, J.; Kemp, M.M.; Linhardt, R.J. Mosquito Heparan Sulfate and Its Potential Role in Malaria Infection and Transmission. J. Biol. Chem. 2007, 282, 25376–25384. [Google Scholar] [CrossRef] [Green Version]
- Gong, H.; Kobayashi, K.; Sugi, T.; Takemae, H.; Kurokawa, H.; Horimoto, T.; Akashi, H.; Kato, K. A Novel PAN/Apple Domain-Containing Protein from Toxoplasma Gondii: Characterization and Receptor Identification. PLoS ONE 2012, 7, e30169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, A.; Sawesi, O.; Pettersson, U.; Dagälv, A.; Kjellén, L.; Lundén, A.; Åbrink, M. Serglycin Proteoglycans Limit Enteropathy in Trichinella Spiralis-Infected Mice. BMC Immunol. 2016, 17, 15. [Google Scholar] [CrossRef] [PubMed]
- Preston, R. Crisis in the Red Zone: The Story of the Deadliest Ebola Outbreak in History, and of the Viruses to Come; Random House: New York, NY, USA, 2019. [Google Scholar]
- Tipton, T.R.W.; Hall, Y.; Bore, J.A.; White, A.; Sibley, L.S.; Sarfas, C.; Yuki, Y.; Martin, M.; Longet, S.; Mellors, J.; et al. Characterisation of the T-Cell Response to Ebola Virus Glycoprotein amongst Survivors of the 2013–16 West Africa Epidemic. Nat. Commun. 2021, 12, 1153. [Google Scholar] [CrossRef] [PubMed]
- Oldstone, M.B.A.; Oldstone, M.R. Ebola’s Curse: 2013–2016 Outbreak in West Africa; Academic Press: London, UK, 2017; ISBN 978-0-12-813889-2. [Google Scholar]
- Escaffre, O.; Juelich, T.L.; Freiberg, A.N. Polyphenylene Carboxymethylene (PPCM) in Vitro Antiviral Efficacy against Ebola Virus in the Context of a Sexually Transmitted Infection. Antiviral Res. 2019, 170, 104567. [Google Scholar] [CrossRef]
- Coltart, C.E.M.; Lindsey, B.; Ghinai, I.; Johnson, A.M.; Heymann, D.L. The Ebola Outbreak, 2013–2016: Old Lessons for New Epidemics. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160297. [Google Scholar] [CrossRef] [PubMed]
- Quammen, D. Spillover: Animal Infections and the Next Human Pandemic, 1st ed.; W.W. Norton & Co.: New York, NY, USA, 2012; ISBN 978-0-393-06680-7. [Google Scholar]
- Preston, R. The Hot Zone, 1st ed.; Random House: New York, NY, USA, 1994; ISBN 0-679-43094-6. [Google Scholar]
- Prasad, A.N.; Woolsey, C.; Geisbert, J.B.; Agans, K.N.; Borisevich, V.; Deer, D.J.; Mire, C.E.; Cross, R.W.; Fenton, K.A.; Broder, C.C.; et al. Resistance of Cynomolgus Monkeys to Nipah and Hendra Virus Disease Is Associated With Cell-Mediated and Humoral Immunity. J. Infect. Dis. 2020, 221, S436–S447. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization Hendra Virus Infection. Available online: https://www.who.int/health-topics/hendra-virus-disease#tab=tab_1 (accessed on 13 October 2021).
- Dang, H.V.; Chan, Y.-P.; Park, Y.-J.; Snijder, J.; Da Silva, S.C.; Vu, B.; Yan, L.; Feng, Y.-R.; Rockx, B.; Geisbert, T.W.; et al. An Antibody against the F Glycoprotein Inhibits Nipah and Hendra Virus Infections. Nat. Struct. Mol. Biol. 2019, 26, 980–987. [Google Scholar] [CrossRef] [PubMed]
- Boardman, W.S.J.; Baker, M.L.; Boyd, V.; Crameri, G.; Peck, G.R.; Reardon, T.; Smith, I.G.; Caraguel, C.G.B.; Prowse, T.A.A. Seroprevalence of Three Paramyxoviruses; Hendra Virus, Tioman Virus, Cedar Virus and a Rhabdovirus, Australian Bat Lyssavirus, in a Range Expanding Fruit Bat, the Grey-Headed Flying Fox (Pteropus Poliocephalus). PLoS ONE 2020, 15, e0232339. [Google Scholar] [CrossRef] [PubMed]
- Woon, A.P.; Boyd, V.; Todd, S.; Smith, I.; Klein, R.; Woodhouse, I.B.; Riddell, S.; Crameri, G.; Bingham, J.; Wang, L.-F.; et al. Acute Experimental Infection of Bats and Ferrets with Hendra Virus: Insights into the Early Host Response of the Reservoir Host and Susceptible Model Species. PLoS Pathog. 2020, 16, e1008412. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Song, Y.; Chen, Y.; Wu, N.; Xu, J.; Sun, C.; Zhang, J.; Weng, T.; Zhang, Z.; Wu, Z.; et al. Molecular Architecture of the SARS-CoV-2 Virus. Cell 2020, 183, 730–738.e13. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, N. MERS Coronovirus Has Probably Been Present in Bats for Many Years, Research Shows. BMJ 2013, 347, f6141. [Google Scholar] [CrossRef] [PubMed]
- Brook, C.E.; Boots, M.; Chandran, K.; Dobson, A.P.; Drosten, C.; Graham, A.L.; Grenfell, B.T.; Müller, M.A.; Ng, M.; Wang, L.-F.; et al. Accelerated Viral Dynamics in Bat Cell Lines, with Implications for Zoonotic Emergence. eLife 2020, 9, e48401. [Google Scholar] [CrossRef]
- Pavlovich, S.S.; Lovett, S.P.; Koroleva, G.; Guito, J.C.; Arnold, C.E.; Nagle, E.R.; Kulcsar, K.; Lee, A.; Thibaud-Nissen, F.; Hume, A.J.; et al. The Egyptian Rousette Genome Reveals Unexpected Features of Bat Antiviral Immunity. Cell 2018, 173, 1098–1110.e18. [Google Scholar] [CrossRef] [Green Version]
- Schountz, T.; Baker, M.L.; Butler, J.; Munster, V. Immunological Control of Viral Infections in Bats and the Emergence of Viruses Highly Pathogenic to Humans. Front. Immunol. 2017, 8, 1098. [Google Scholar] [CrossRef]
- Yan, L.; Song, Y.; Xia, K.; He, P.; Zhang, F.; Chen, S.; Pouliot, R.; Weiss, D.J.; Tandon, R.; Bates, J.T.; et al. Heparan Sulfates from Bat and Human Lung and Their Binding to the Spike Protein of SARS-CoV-2 Virus. Carbohydr. Polym. 2021, 260, 117797. [Google Scholar] [CrossRef]
- Schmitt, K.; Curlin, J.; Kumar, D.M.; Remling-Mulder, L.; Feely, S.; Stenglein, M.; O’Connor, S.; Marx, P.; Akkina, R. SIV Progenitor Evolution toward HIV: A Humanized Mouse Surrogate Model for SIVsm Adaptation toward HIV-2. J. Med. Primatol. 2018, 47, 298–301. [Google Scholar] [CrossRef]
- Flaviviridae_Viral Hemorrhagic Fevers (VHFs). Available online: https://www.cdc.gov/vhf/virus-families/flaviviridae.html (accessed on 13 October 2021).
- Kim, S.Y. Glycosaminoglycans in Infectious Diseases. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/brv.12034 (accessed on 13 October 2021).
- Chen, Y.; Maguire, T.; Hileman, R.E.; Fromm, J.R.; Esko, J.D.; Linhardt, R.J.; Marks, R.M. Dengue Virus Infectivity Depends on Envelope Protein Binding to Target Cell Heparan Sulfate. Nat. Med. 1997, 3, 866–871. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, P.F.C.; Travassos da Rosa, A.P.A.; Rodrigues, S.G.; Travassos da Rosa, E.S.; Dégallier, N.; Travassos da Rosa, J.F.S. Inadequate Management of Natural Ecosystem in the Brazilian Amazon Region Results in the Emergence and Reemergence of Arboviruses. Cad. Saúde Pública 2001, 17, S155–S164. [Google Scholar] [CrossRef]
- Akaberi, D.; Båhlström, A.; Chinthakindi, P.K.; Nyman, T.; Sandström, A.; Järhult, J.D.; Palanisamy, N.; Lundkvist, Å.; Lennerstrand, J. Targeting the NS2B-NS3 Protease of Tick-Borne Encephalitis Virus with Pan-Flaviviral Protease Inhibitors. Antiviral Res. 2021, 190, 105074. [Google Scholar] [CrossRef] [PubMed]
- Thongtan, T.; Cheepsunthorn, P.; Chaiworakul, V.; Rattanarungsan, C.; Wikan, N.; Smith, D.R. Highly Permissive Infection of Microglial Cells by Japanese Encephalitis Virus: A Possible Role as a Viral Reservoir. Microbes Infect. 2010, 12, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Yan, R.; Li, Z.; Tang, X.; Zhou, Y.; He, H. Economic and Disease Burden of Japanese Encephalitis in Zhejiang Province, 2013–2018. PLoS Negl. Trop. Dis. 2021, 15, e0009505. [Google Scholar] [CrossRef]
- Su, Q.; Xie, Z.-X.; He, F.; Liu, Z.-C.; Song, X.-J.; Zhao, F.-C.; Li, D.; Che, F.-Y. Adults with Severe Japanese Encephalitis: A Retrospective Analysis of 9 Cases in Linyi, China. Neurol. Sci. 2021, 42, 2811–2817. [Google Scholar] [CrossRef]
- Syed, S.F.; Zhao, Q.; Umer, M.; Alagawany, M.; Ujjan, I.A.; Soomro, F.; Bangulzai, N.; Baloch, A.H.; Abd El-Hack, M.; Zhou, E.-M.; et al. Past, Present and Future of Hepatitis E Virus Infection: Zoonotic Perspectives. Microb. Pathog. 2018, 119, 103–108. [Google Scholar] [CrossRef]
- Reuter, G.; Boros, Á.; Pankovics, P. Review of Hepatitis E Virus in Rats: Evident Risk of Species Orthohepevirus C to Human Zoonotic Infection and Disease. Viruses 2020, 12, 1148. [Google Scholar] [CrossRef] [PubMed]
- Hepatitis E Questions and Answers for Health Professionals. Available online: https://www.cdc.gov/hepatitis/hev/hevfaq.htm (accessed on 13 October 2021).
- Wang, B.; Meng, X.-J. Hepatitis E Virus: Host Tropism and Zoonotic Infection. Curr. Opin. Microbiol. 2021, 59, 8–15. [Google Scholar] [CrossRef]
- Hatami, H. History of Rabies in Traditional Medicine’s Resources and Iranian Research Studies: On the CccasiOn of the World Rabies Day (September 28, 2012). Int. J. Prev. Med. 2012, 3, 593–595. [Google Scholar]
- Reddy, R.V.C.; Mohana Subramanian, B.; Surendra, K.S.N.L.; Babu, R.P.A.; Rana, S.K.; Manjari, K.S.; Srinivasan, V.A. Rabies Virus Isolates of India—Simultaneous Existence of Two Distinct Evolutionary Lineages. Infect. Genet. Evol. 2014, 27, 163–172. [Google Scholar] [CrossRef]
- Williamson, J.G. Rabies: Symptoms, Treatment and Prevention; Virology Research Progress series; Nova Science Publishers: Hauppauge, NY, USA, 2010; ISBN 978-1-61122-332-3. [Google Scholar]
- Magouras, I.; Brookes, V.J.; Jori, F.; Martin, A.; Pfeiffer, D.U.; Dürr, S. Emerging Zoonotic Diseases: Should We Rethink the Animal–Human Interface? Front. Vet. Sci. 2020, 7, 582743. [Google Scholar] [CrossRef] [PubMed]
- Saylors, K.E.; Mouiche, M.M.; Lucas, A.; McIver, D.J.; Matsida, A.; Clary, C.; Maptue, V.T.; Euren, J.D.; LeBreton, M.; Tamoufe, U. Market Characteristics and Zoonotic Disease Risk Perception in Cameroon Bushmeat Markets. Soc. Sci. Med. 2021, 268, 113358. [Google Scholar] [CrossRef]
- Naguib, M.M.; Li, R.; Ling, J.; Grace, D.; Nguyen-Viet, H.; Lindahl, J.F. Live and Wet Markets: Food Access versus the Risk of Disease Emergence. Trends Microbiol. 2021, S0966842X21000433. [Google Scholar] [CrossRef] [PubMed]
- Kieny, M.P.; Salama, P. WHO R&D Blueprint: A Global Coordination Mechanism for R&D Preparedness. Lancet 2017, 389, 2469–2470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, H.; Lear-Rooney, C.M.; Johansen, L.; Varhegyi, E.; Chen, Z.W.; Olinger, G.G.; Rong, L. Inhibition of Ebola and Marburg Virus Entry by G Protein-Coupled Receptor Antagonists. J. Virol. 2015, 89, 9932–9938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvador, B.; Sexton, N.R.; Carrion, R.; Nunneley, J.; Patterson, J.L.; Steffen, I.; Lu, K.; Muench, M.O.; Lembo, D.; Simmons, G. Filoviruses Utilize Glycosaminoglycans for Their Attachment to Target Cells. J. Virol. 2013, 87, 3295–3304. [Google Scholar] [CrossRef] [Green Version]
- Tamhankar, M.; Gerhardt, D.M.; Bennett, R.S.; Murphy, N.; Jahrling, P.B.; Patterson, J.L. Heparan Sulfate Is an Important Mediator of Ebola Virus Infection in Polarized Epithelial Cells. Virol. J. 2018, 15, 135. [Google Scholar] [CrossRef] [Green Version]
- Buijsers, B.; Yanginlar, C.; Maciej-Hulme, M.L.; de Mast, Q.; van der Vlag, J. Beneficial Non-Anticoagulant Mechanisms Underlying Heparin Treatment of COVID-19 Patients. EBioMedicine 2020, 59, 102969. [Google Scholar] [CrossRef]
- Liu, L.; Chopra, P.; Li, X.; Bouwman, K.M.; Tompkins, S.M.; Wolfert, M.A.; de Vries, R.P.; Boons, G.-J. Heparan Sulfate Proteoglycans as Attachment Factor for SARS-CoV-2. ACS Cent. Sci. 2021, 7, 1009–1018. [Google Scholar] [CrossRef]
- Gozzo, L.; Viale, P.; Longo, L.; Vitale, D.C.; Drago, F. The Potential Role of Heparin in Patients With COVID-19: Beyond the Anticoagulant Effect. A Review. Front. Pharmacol. 2020, 11, 1307. [Google Scholar] [CrossRef]
- Lertjuthaporn, S.; Cicala, C.; Van Ryk, D.; Liu, M.; Yolitz, J.; Wei, D.; Nawaz, F.; Doyle, A.; Horowitch, B.; Park, C.; et al. Select Gp120 V2 Domain Specific Antibodies Derived from HIV and SIV Infection and Vaccination Inhibit Gp120 Binding to A4β7. PLoS Pathog. 2018, 14, e1007278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, C.W.; Sam, I.-C.; Chong, W.L.; Lee, V.S.; Chan, Y.F. Polysulfonate Suramin Inhibits Zika Virus Infection. Antiviral Res. 2017, 143, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Hyatt, J.G.; Prévost, S.; Devos, J.M.; Mycroft-West, C.J.; Skidmore, M.A.; Winter, A. Molecular Changes in Dengue Envelope Protein Domain III upon Interaction with Glycosaminoglycans. Pathogens 2020, 9, 935. [Google Scholar] [CrossRef]
- Watterson, D.; Kobe, B.; Young, P.R. Residues in Domain III of the Dengue Virus Envelope Glycoprotein Involved in Cell-Surface Glycosaminoglycan Binding. J. Gen. Virol. 2012, 93, 72–82. [Google Scholar] [CrossRef]
- Park, J.; Kang, T.-B.; Lim, J.-H.; Won, H.-S. Molecular Targeting of VEGF with a Suramin Fragment–DOCA Conjugate by Mimicking the Action of Low Molecular Weight Heparins. Biomolecules 2020, 11, 46. [Google Scholar] [CrossRef] [PubMed]
- Basavannacharya, C.; Vasudevan, S.G. Suramin Inhibits Helicase Activity of NS3 Protein of Dengue Virus in a Fluorescence-Based High Throughput Assay Format. Biochem. Biophys. Res. Commun. 2014, 453, 539–544. [Google Scholar] [CrossRef]
- Lee, E.; Lobigs, M. Mechanism of Virulence Attenuation of Glycosaminoglycan-Binding Variants of Japanese Encephalitis Virus and Murray Valley Encephalitis Virus. J. Virol. 2002, 76, 4901–4911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiou, S.-S.; Liu, H.; Chuang, C.-K.; Lin, C.-C.; Chen, W.-J. Fitness of Japanese Encephalitis Virus to Neuro-2a Cells Is Determined by Interactions of the Viral Envelope Protein with Highly Sulfated Glycosaminoglycans on the Cell Surface. J. Med. Virol. 2005, 76, 583–592. [Google Scholar] [CrossRef]
- Kim, S.Y.; Koetzner, C.A.; Payne, A.F.; Nierode, G.J.; Yu, Y.; Wang, R.; Barr, E.; Dordick, J.S.; Kramer, L.D.; Zhang, F.; et al. Glycosaminoglycan Compositional Analysis of Relevant Tissues in Zika Virus Pathogenesis and in Vitro Evaluation of Heparin as an Antiviral against Zika Virus Infection. Biochemistry 2019, 58, 1155–1166. [Google Scholar] [CrossRef] [PubMed]
- Leonova, G.N.; Belikov, S.I. Effect of Glycosaminoglycans on Pathogenic Properties Far-Eastern Tick-Borne Encephalitis Virus. Bull. Exp. Biol. Med. 2019, 167, 482–485. [Google Scholar] [CrossRef]
- Rincón, V.; Corredor, A.; Martínez-Gutíerrez, M.; Castellanos, J.E. Fluorometric Cell-ELISA for Quantifying Rabies Infection and Heparin Inhibition. J. Virol. Methods 2005, 127, 33–39. [Google Scholar] [CrossRef]
- Jinno, A.; Park, P.W. Role of Glycosaminoglycans in Infectious Disease. In Glycosaminoglycans; Balagurunathan, K., Nakato, H., Desai, U.R., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2015; Volume 1229, pp. 567–585. ISBN 978-1-4939-1713-6. [Google Scholar]
- Park, P.W. Glycosaminoglycans and Infection. Front. Biosci. 2016, 21, 1260–1277. [Google Scholar] [CrossRef] [Green Version]
- Simon, E.J.; Linstedt, A.D. Site-Specific Glycosylation of Ebola Virus Glycoprotein by Human Polypeptide GalNAc-Transferase 1 Induces Cell Adhesion Defects. J. Biol. Chem. 2018, 293, 19866–19873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- New Dengue Hemorrhagic Fever Findings from School of Medicine Discussed (Suramin Inhibits Helicase Activity of NS3 Protein of Dengue Virus in a Fluorescence-Based High Throughput Assay Format). Malar. Wkly. Atlanta GA 2015, 4.
- Mire, C.E.; Chan, Y.-P.; Borisevich, V.; Cross, R.W.; Yan, L.; Agans, K.N.; Dang, H.V.; Veesler, D.; Fenton, K.A.; Geisbert, T.W.; et al. A Cross-Reactive Humanized Monoclonal Antibody Targeting Fusion Glycoprotein Function Protects Ferrets Against Lethal Nipah Virus and Hendra Virus Infection. J. Infect. Dis. 2020, 221, S471–S479. [Google Scholar] [CrossRef]
- Bradel-Tretheway, B.G.; Zamora, J.L.R.; Stone, J.A.; Liu, Q.; Li, J.; Aguilar, H.C. Nipah and Hendra Virus Glycoproteins Induce Comparable Homologous but Distinct Heterologous Fusion Phenotypes. J. Virol. 2019, 93, e00577-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalbhor, M.S.; Bhowmick, S.; Alanazi, A.M.; Patil, P.C.; Islam, M.A. Multi-Step Molecular Docking and Dynamics Simulation-Based Screening of Large Antiviral Specific Chemical Libraries for Identification of Nipah Virus Glycoprotein Inhibitors. Biophys. Chem. 2021, 270, 106537. [Google Scholar] [CrossRef]
- Dong, J.; Cross, R.W.; Doyle, M.P.; Kose, N.; Mousa, J.J.; Annand, E.J.; Borisevich, V.; Agans, K.N.; Sutton, R.; Nargi, R.; et al. Potent Henipavirus Neutralization by Antibodies Recognizing Diverse Sites on Hendra and Nipah Virus Receptor Binding Protein. Cell 2020, 183, 1536–1550.e17. [Google Scholar] [CrossRef]
- Sun, B.; Jia, L.; Liang, B.; Chen, Q.; Liu, D. Phylogeography, Transmission, and Viral Proteins of Nipah Virus. Virol. Sin. 2018, 33, 385–393. [Google Scholar] [CrossRef] [Green Version]
- Khusro, A. Hendra Virus Infection in Horses: A Review on Emerging Mystery Paramyxovirus. J. Equine Vet. Sci. 2020, 91, 103149. [Google Scholar] [CrossRef]
- Das, S.K. The Pathophysiology, Diagnosis and Treatment of Corona Virus Disease 2019 (COVID-19). Indian J. Clin. Biochem. 2020, 35, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, E.; Hernandez-Vargas, E.A. (Eds.) Chapter 5—Modeling Ebola Virus Infection. In Modeling and Control of Infectious Diseases in the Host; Academic Press: London, UK, 2019; pp. 85–103. ISBN 978-0-12-813052-0. [Google Scholar]
- Salgado-Benvindo, C.; Thaler, M.; Tas, A.; Ogando, N.S.; Bredenbeek, P.J.; Ninaber, D.K.; Wang, Y.; Hiemstra, P.S.; Snijder, E.J. Suramin Inhibits SARS-CoV-2 Infection in Cell Culture by Interfering with Early Steps of the Replication Cycle. Antimicrob. Agents Chemother. 2020, 64, 11. [Google Scholar] [CrossRef]
- Bauer, S.; Jin, W.; Zhang, F.; Linhardt, R.J. The Application of Seaweed Polysaccharides and Their Derived Products with Potential for the Treatment of Alzheimer’s Disease. Mar. Drugs 2021, 19, 89. [Google Scholar] [CrossRef] [PubMed]
- Zoepfl, M.; Dwivedi, R.; Taylor, M.C.; Pomin, V.H.; McVoy, M.A. Antiviral Activities of Four Marine Sulfated Glycans against Adenovirus and Human Cytomegalovirus. Antiviral Res. 2021, 190, 105077. [Google Scholar] [CrossRef]
- Luo, Z.; Tian, D.; Zhou, M.; Xiao, W.; Zhang, Y.; Li, M.; Sui, B.; Wang, W.; Guan, H.; Chen, H.; et al. λ-Carrageenan P32 Is a Potent Inhibitor of Rabies Virus Infection. PLoS ONE 2015, 10, e0140586. [Google Scholar] [CrossRef] [PubMed]
- Rentsch, C.T.; Beckman, J.A.; Tomlinson, L.; Gellad, W.F.; Alcorn, C.; Kidwai-Khan, F.; Skanderson, M.; Brittain, E.; King, J.T.; Ho, Y.-L.; et al. Early Initiation of Prophylactic Anticoagulation for Prevention of Coronavirus Disease 2019 Mortality in Patients Admitted to Hospital in the United States: Cohort Study. BMJ 2021, 372, n311. [Google Scholar] [CrossRef]
- Pawlowski, C.; Venkatakrishnan, A.; Kirkup, C.; Berner, G.; Puranik, A.; O’Horo, J.C.; Badley, A.D.; Soundararajan, V. Enoxaparin Is Associated with Lower Rates of Mortality than Unfractionated Heparin in Hospitalized COVID-19 Patients. EClinicalMedicine 2021, 33, 100774. [Google Scholar] [CrossRef]
- Hernando, P.J.; Dedola, S.; Marín, M.J.; Field, R.A. Recent Developments in the Use of Glyconanoparticles and Related Quantum Dots for the Detection of Lectins, Viruses, Bacteria and Cancer Cells. Front. Chem. 2021, 9, 668509. [Google Scholar] [CrossRef]

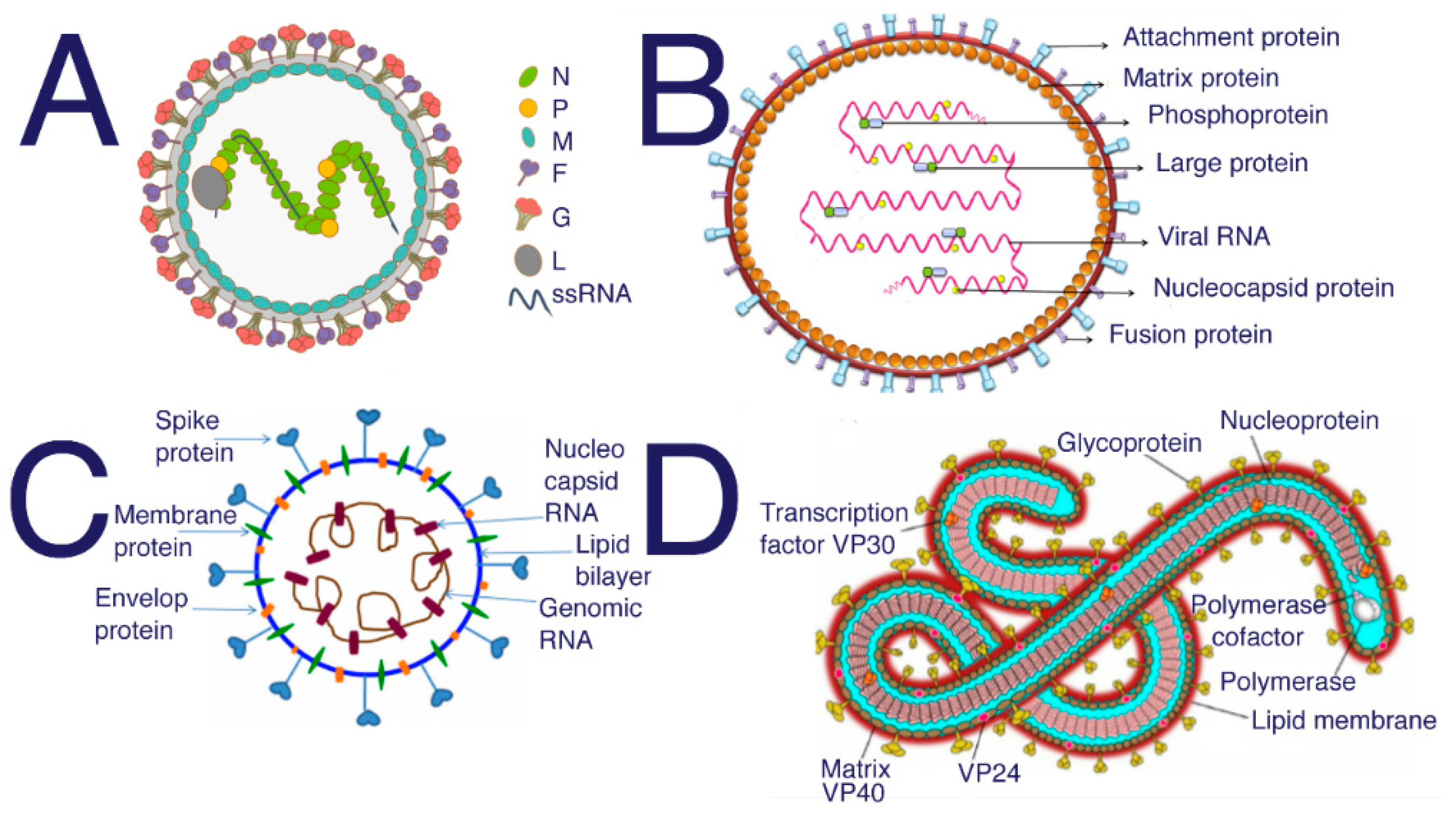

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Location | Virus/Strain |
|---|---|---|
| 1976 | Zaire (Now DRC) | Ebola Zaire |
| 1976 | Sudan (Now South Sudan) | Ebola Sudan |
| 1976 | U.K. | Ebola Sudan |
| 1989 | Philippines | Ebola Reston |
| 1990 | U.S. | Ebola Reston |
| 1994 | Gabon | Ebola Zaire |
| 1994 | Mackay, Australia | Hendra |
| 1994 | Brisbane, Australia | Hendra |
| 1995 | DRC | Ebola Zaire |
| 1996 | South Africa | Ebola Zaire |
| 1996 | Russia | Ebola Zaire |
| 1998 | Malaysia | Nipah |
| 1999 | Cairns, Australia | Hendra |
| 2000 | Uganda | Ebola Zaire |
| 2001 | Gabon | Ebola Zaire |
| 2001 | Bangladesh | Nipah |
| 2002 | Republic of Congo | Ebola Zaire |
| 2002 | Bangladesh | Nipah |
| 2003 | Bangladesh | Nipah |
| 2003 | Guangdong, China | SARS |
| 2004 | Sudan (Now South Sudan) | Ebola Sudan |
| 2004 | Russia | Ebola Zaire |
| 2004 | Cairns, Australia | Hendra |
| 2004 | Townsville, Australia | Hendra |
| 2004 | Bangladesh | Nipah |
| 2005 | Bangladesh | Nipah |
| 2006 | Peachester, Australia | Hendra |
| 2006 | Muwillumbah, Australia | Hendra |
| 2006 | Bangladesh | Nipah |
| 2007 | DRC | Ebola Zaire |
| 2007 | Clifton Beach, Australia | Hendra |
| 2007 | Peachester, Australia | Hendra |
| 2007 | Bangladesh | Nipah |
| 2008 | Philippines | Ebola Reston |
| 2008 | Proserpine, Australia | Hendra |
| 2008 | Brisbane, Australia | Hendra |
| 2008 | Bangladesh | Nipah |
| 2009 | Bangladesh | Nipah |
| 2010 | Bangladesh | Nipah |
| 2011 | Uganda | Ebola Sudan |
| 2011 | Bangladesh | Nipah |
| 2012 | DRC | Ebola Sudan |
| 2012 | Bangladesh | Nipah |
| 2012 | Jordan | MERS |
| 2012 | Saudi Arabia | MERS |
| 2013 | West Africa | Ebola Zaire |
| 2013 | Bangladesh | Nipah |
| 2014 | Bangladesh | Nipah |
| 2015 | Bangladesh | Nipah |
| 2015 | Republic of Korea | MERS |
| 2016 | Bangladesh | Nipah |
| 2017 | Bangladesh | Nipah |
| 2018 | DRC | Ebola Bundibugyo |
| 2018 | India | Nipah |
| 2019 | Hunter Valley, Australia | Hendra |
| 2019 | Wuhan, China | SARS-Covid 19 |
| Disease | Pathogen | Animal | Insect Vector | GAG-Binding Protein | Reference |
|---|---|---|---|---|---|
| VIRAL | |||||
| AIDS | HIV Lentivirus | Chimps | None | GP-120 | [15] |
| Bird flu | Influenza A H5N1 | Waterfowl | None | - | - |
| Chikungunya | Alphavirus | [16] | |||
| Covid/MERS/SARS | Coronavirus | Bats, civets/camels | None | Spike glycoprotein | [11,12] |
| Dengue fever | Flavivirus | Monkeys | Mosquito | Envelope protein | |
| Ebola | Filovirus | Bats | None | Filoviral glycoprotein | [17] |
| Encephalitis from ticks | Flavivirus | Tick | Envelope protein | [18] | |
| Hendra | Hendra Henipavirus | Bats | None | Ephrin-B2 and -B3 | [19] |
| Hepatitis E | Orthohepevirus HEV | Rats | None | ORF2 capsid protein | [20] |
| Japanese encephalitis | Flavivirus | Bats/pigs | Mosquitos | ||
| Louping ill | Flavivirus | Sheep | Tick | - | - |
| Lymphocytic choriomeningitis | Arenavirus | Rodents | None | - | - |
| Mayaro | Alphavirus Togaviridae | Monkeys | Mosquitos | [21] | |
| Nipah | Nipah Henipavirus | Bats | None | Ephrin-B2 and -B3 | [19] |
| Orf infection | Para poxvirus | Sheep | None | - | - |
| Rabies | Lyssavirus | Bats | Attachment factor | [22] | |
| Saint Louis encephalitis | Flavivirus | ||||
| Swine flu | Virus, Influenza A H1N1 | Swine | None | - | - |
| Venezuelan equine encephalitis | Alphavirus | ||||
| West Nile virus | Flavivirus | Mosquitos | Envelope protein | [23] | |
| Yellow fever | Flavivirus | ||||
| Zika | Flavivirus | Monkeys | Mosquitos | Envelope protein | [24] |
| BACTERIAL | |||||
| Anthrax | Bacillus anthracis | Hoofed animals | None | - | - |
| Bovine tuberculosis | Mycobacterium bovis | Cattle | None | Heparin-binding hemagglutinin | [25] |
| Brucellosis | Brucella sp. | Cows, goats, sheep | None | Unknown | [26] |
| Campylobacter infection | Campylobacter sp. | None | FliD protein | [27] | |
| Cat scratch fever | Bartonella henselae | Cats | None | Pap31 | [28] |
| Erysipeloid | Erysipelothrix rhusiopathiae | Fish, birds, mammals | None | - | - |
| Glanders | Burkholderia mallei | None | - | - | |
| Leptospirosis | Leptospira sp. | Cattle | None | LigB adhesin | [29] |
| Listeria infection | Listeria monocytogenes | Ruminants, sheep | None | Surface protein ActA | [30] |
| Lyme disease | Borrelia burgdorferi | Deer | Tick | OspF-related proteins, adhesion BBK32, adhesion DbpA | [31] |
| Parrot fever | Chlamydia psittaci | Parrots and other birds | None | Unknown | [32] |
| Pasteurellosis | Pasteurella multocida | Domestic animals | None | OmpA β-barrel ion channel protein | [33] |
| Plague | Yersinia pestis | Rats and rodents | Flea | Ail outer membrane protein | [34] |
| Q fever | Coxiella burnetii | Sheep, goats, cattle | None | - | - |
| Rocky Mountain spotted fever | Rickettsia rickettsii | Rodents, dogs | Tick | Unknown | [32] |
| Tularemia | Francisella tularensis | Rodents, rabbits | Tick or deerfly | ||
| Zoonotic diphtheria | Corynebacterium diphtheria | Dogs | None | - | - |
| FUNGAL | |||||
| Ringworm | Tinea corporis | Domestic animal species | None | - | - |
| PARASITIC | |||||
| Cryptosporidiosis | Cryptosporidium sp. | Calves and lambs | None | Mucin-like glycoprotein, CpClec | [35] |
| Giardiasis | Giardia lamblia | Domestic and wild mammals | None | Alpha-11 Giardin Annexin | [36] |
| Malaria | Plasmodium falciparum | Nonhuman primates | Mosquito | Circumsporozoite protein | [37] |
| Toxocariasis | Toxocara canis or T. cati | Dogs/Cats | None | - | - |
| Toxoplasmosis | Toxoplasma gondii | Cats | None | Protein of 104 kDa (P104) Microneme-2 (MIC2) | [38] |
| Trichinellosis | Trichinella sp. | Pigs | None | Unknown | [39] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bauer, S.; Zhang, F.; Linhardt, R.J. Implications of Glycosaminoglycans on Viral Zoonotic Diseases. Diseases 2021, 9, 85. https://doi.org/10.3390/diseases9040085
Bauer S, Zhang F, Linhardt RJ. Implications of Glycosaminoglycans on Viral Zoonotic Diseases. Diseases. 2021; 9(4):85. https://doi.org/10.3390/diseases9040085
Chicago/Turabian StyleBauer, Sarah, Fuming Zhang, and Robert J. Linhardt. 2021. "Implications of Glycosaminoglycans on Viral Zoonotic Diseases" Diseases 9, no. 4: 85. https://doi.org/10.3390/diseases9040085
APA StyleBauer, S., Zhang, F., & Linhardt, R. J. (2021). Implications of Glycosaminoglycans on Viral Zoonotic Diseases. Diseases, 9(4), 85. https://doi.org/10.3390/diseases9040085