Valproic Acid Downregulates Cytokine Expression in Human Macrophages Infected with Dengue Virus


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Dengue Virus
2.2. Peripheral Blood Mononuclear Cell (PBMC) Purification
2.3. In Vitro Generation of Monocyte-Derived Macrophages (MDMs)
2.4. Cell Viability Assay
2.5. Treatment with HDAC Inhibitors and Cytokine Quantification
2.6. Cytokine mRNA Quantification
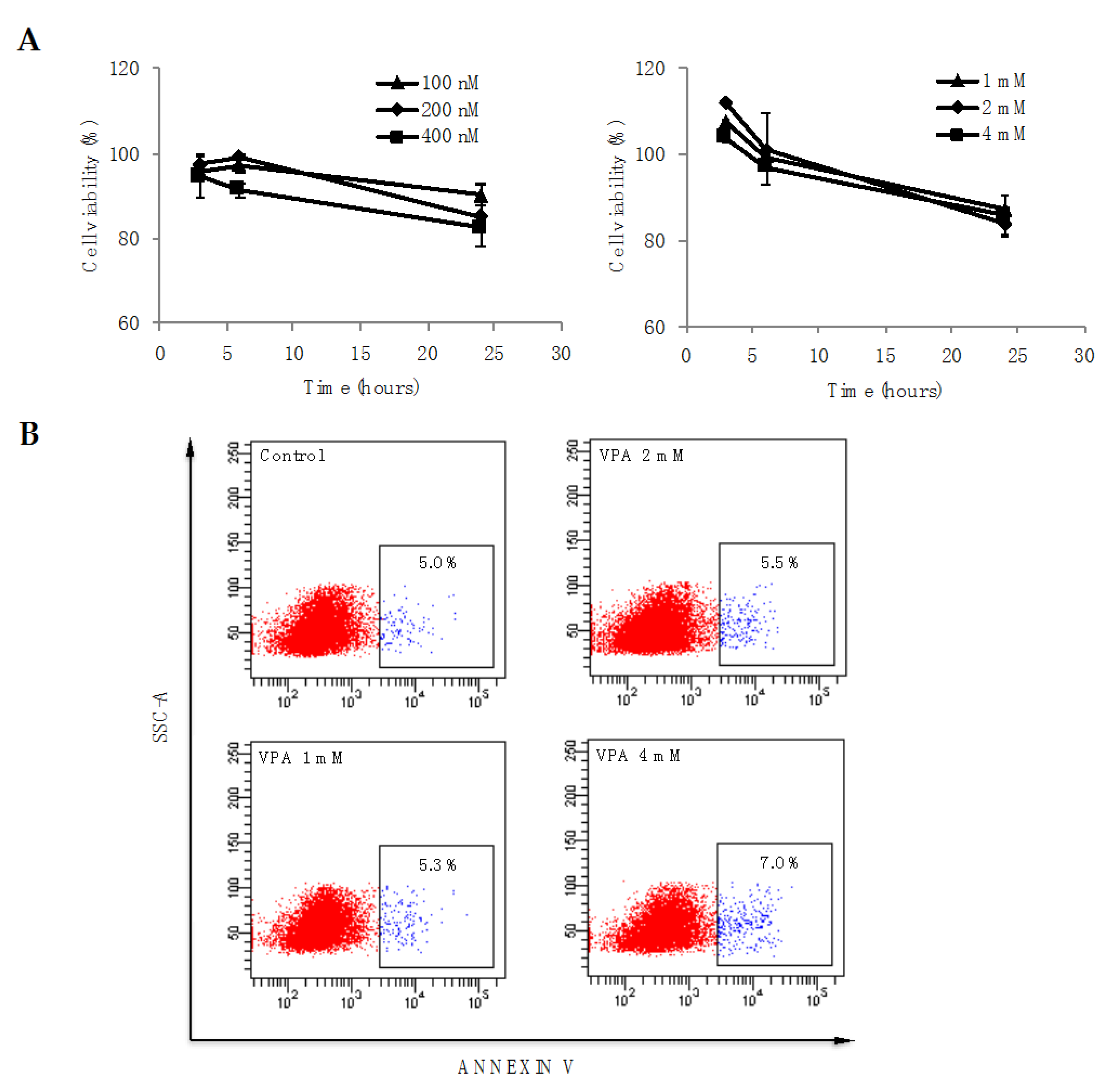
2.7. Apoptosis Assay
2.8. Statistical Analysis
3. Results
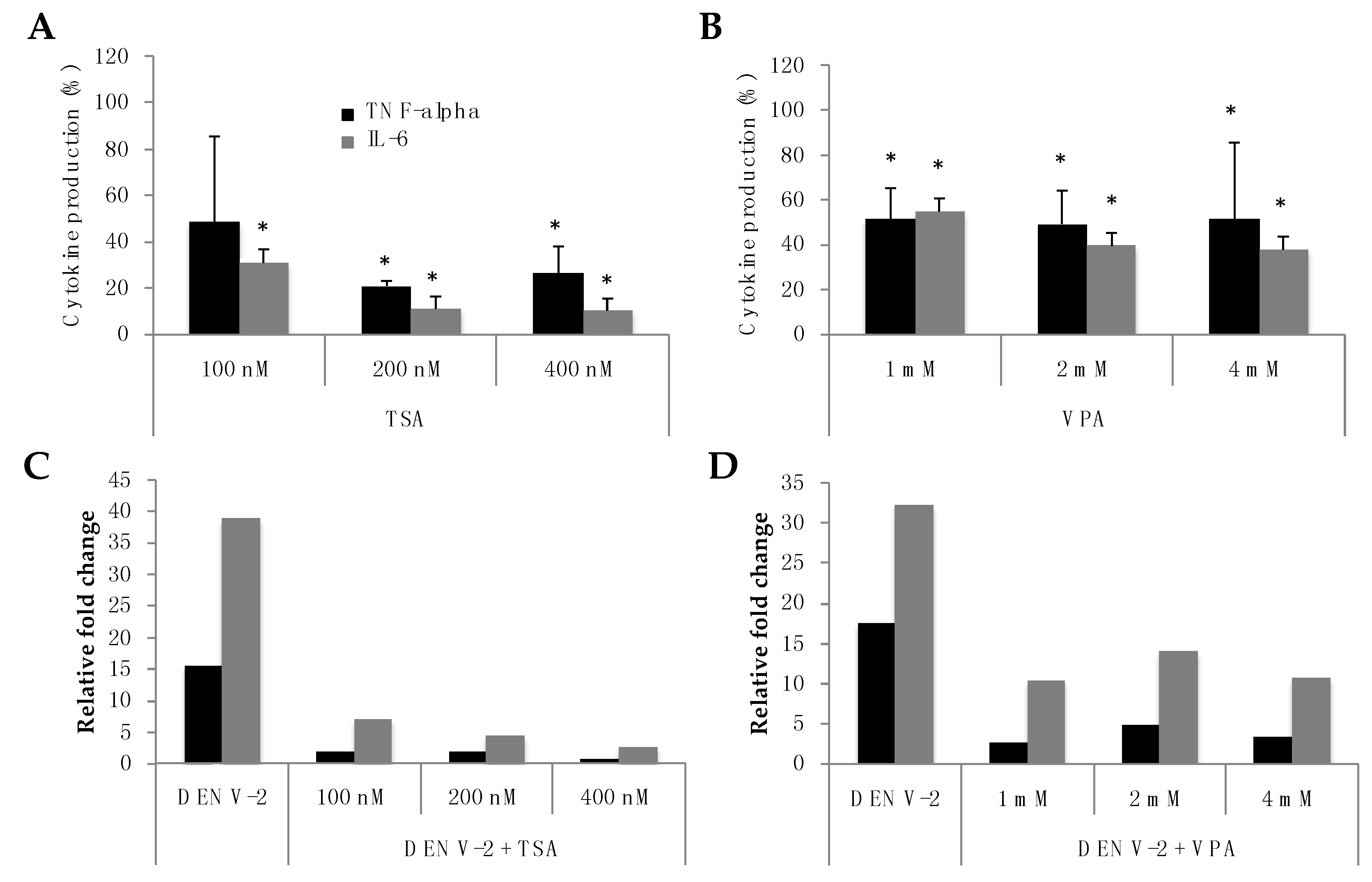
3.1. HDAC Inhibition in PBMCs Infected with DENV-2 Induces a Decreased TNF-Alpha and IL-6 Production
3.2. HDAC Inhibition Have Not a Significant Effect in Cell Viability
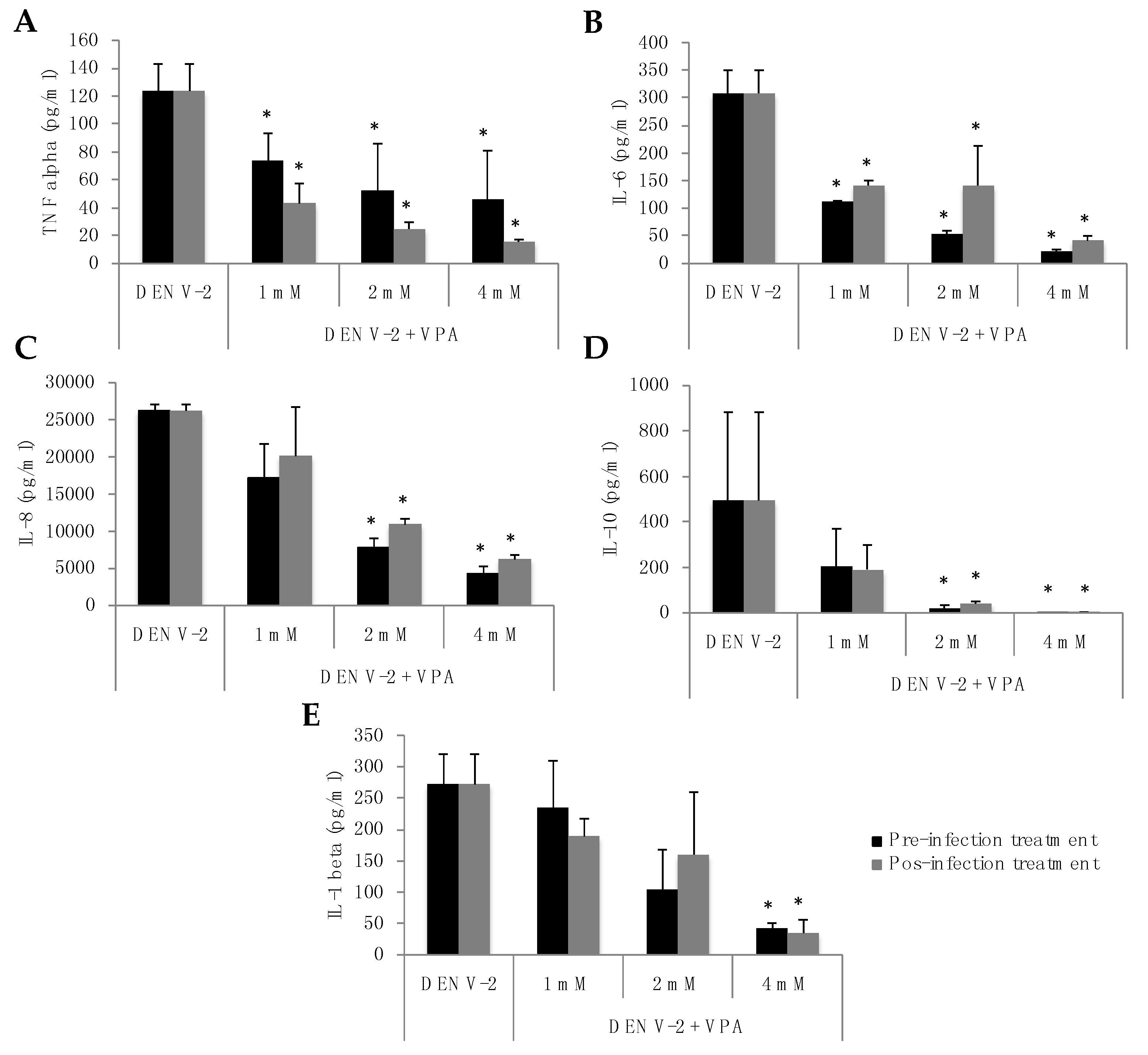
3.3. VPA Treatment of MDMs Infected with DENV-2 Induces a Significant Decrease in Virus-Induced Inflammatory Cytokine Production
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Shepard, D.S.; Undurraga, E.A.; Halasa, Y.A.; Stanaway, J.D. The global economic burden of dengue: A systematic analysis. Lancet Infect. Dis. 2016, 16, 935–941. [Google Scholar] [CrossRef]
- Guzman, M.G.; Harris, E. Dengue. Lancet 2015, 385, 453–465. [Google Scholar] [CrossRef]
- Guzman, M.G.; Gubler, D.J.; Izquierdo, A.; Martinez, E.; Halstead, S.B. Dengue infection. Nat. Rev. Dis. Prim. 2016, 2, 16055. [Google Scholar] [CrossRef] [PubMed]
- Flipse, J.; Smit, J.M. The complexity of a dengue vaccine: A review of the human antibody response. PLoS Negl. Trop. Dis. 2015, 9, e0003749. [Google Scholar] [CrossRef] [PubMed]
- Guy, B.; Jackson, N. Dengue vaccine: Hypotheses to understand CYD-TDV-induced protection. Nat. Rev. Microbiol. 2016, 14, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Das Gupta, K.; Shakespear, M.R.; Iyer, A.; Fairlie, D.P.; Sweet, M.J. Histone deacetylases in monocyte/macrophage development, activation and metabolism: Refining hdac targets for inflammatory and infectious diseases. Clin. Transl. Immunol. 2016, 5, e62. [Google Scholar] [CrossRef] [PubMed]
- Roger, T.; Lugrin, J.; Le Roy, D.; Goy, G.; Mombelli, M.; Koessler, T.; Ding, X.C.; Chanson, A.L.; Reymond, M.K.; Miconnet, I.; et al. Histone deacetylase inhibitors impair innate immune responses to toll-like receptor agonists and to infection. Blood 2011, 117, 1205–1217. [Google Scholar] [CrossRef] [PubMed]
- Hull, E.E.; Montgomery, M.R.; Leyva, K.J. Hdac inhibitors as epigenetic regulators of the immune system: Impacts on cancer therapy and inflammatory diseases. BioMed Res. Int. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Huber, L.C.; Brock, M.; Hemmatazad, H.; Giger, O.T.; Moritz, F.; Trenkmann, M.; Distler, J.H.; Gay, R.E.; Kolling, C.; Moch, H.; et al. Histone deacetylase/acetylase activity in total synovial tissue derived from rheumatoid arthritis and osteoarthritis patients. Arthritis Rheum. 2007, 56, 1087–1093. [Google Scholar] [CrossRef] [PubMed]
- Gunawardhana, L.P.; Gibson, P.G.; Simpson, J.L.; Powell, H.; Baines, K.J. Activity and expression of histone acetylases and deacetylases in inflammatory phenotypes of asthma. Clin. Exp. Allergy 2014, 44, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Owens, M.J.; Nemeroff, C.B. Pharmacology of valproate. Psychopharmacol. Bull. 2003, 37, 17–24. [Google Scholar] [PubMed]
- Gottlicher, M.; Minucci, S.; Zhu, P.; Kramer, O.H.; Schimpf, A.; Giavara, S.; Sleeman, J.P.; Lo Coco, F.; Nervi, C.; Pelicci, P.G.; et al. Valproic acid defines a novel class of hdac inhibitors inducing differentiation of transformed cells. EMBO J. 2001, 20, 6969–6978. [Google Scholar] [CrossRef] [PubMed]
- Grabiec, A.M.; Tak, P.P.; Reedquist, K.A. Function of histone deacetylase inhibitors in inflammation. Crit. Rev. Immunol. 2011, 31, 233–263. [Google Scholar] [CrossRef] [PubMed]
- Velandia-Romero, M.L.; Acosta-Losada, O.; Castellanos, J.E. In vivo infection by a neuroinvasive neurovirulent dengue virus. J. Neurovirol. 2012, 18, 374–387. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Wang, S.Y. Activation of terminally differentiated human monocytes/macrophages by dengue virus: Productive infection, hierarchical production of innate cytokines and chemokines, and the synergistic effect of lipopolysaccharide. J. Virol. 2002, 76, 9877–9887. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Delgado, F.G.; Pérez-Acosta, M.; Castellanos, J.E. Descripción de un modelo de infección in vitro con virus dengue empleando células mononucleares humanas de sangre periférica. Iatreia 2014, 27, 267–277. [Google Scholar]
- Pérez-Acosta, M.; Delgado, F.G.; Castellanos, J.E. Soluble ST2 does not regulate TNF- and IL-6 production in dengue virus-infected human monocytes. ISRN Trop. Med. 2013, 2013. [Google Scholar] [CrossRef]
- Castelo-Branco, G.; Stridh, P.; Guerreiro-Cacais, A.O.; Adzemovic, M.Z.; Falcao, A.M.; Marta, M.; Berglund, R.; Gillett, A.; Hamza, K.H.; Lassmann, H.; et al. Acute treatment with valproic acid and l-thyroxine ameliorates clinical signs of experimental autoimmune encephalomyelitis and prevents brain pathology in da rats. Neurobiol. Dis. 2014, 71, 220–233. [Google Scholar] [CrossRef] [PubMed]
- Cantley, M.D.; Fairlie, D.P.; Bartold, P.M.; Marino, V.; Gupta, P.K.; Haynes, D.R. Inhibiting histone deacetylase 1 suppresses both inflammation and bone loss in arthritis. Rheumatol. Oxf. 2015, 54, 1713–1723. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, Z.Y.; Fauser, U.; Schluesener, H.J. Valproic acid attenuates inflammation in experimental autoimmune neuritis. Cell. Mol. Life Sci. 2008, 65, 4055–4065. [Google Scholar] [CrossRef] [PubMed]
- Cetinkaya, M.; Cansev, M.; Cekmez, F.; Tayman, C.; Canpolat, F.E.; Kafa, I.M.; Yaylagul, E.O.; Kramer, B.W.; Sarici, S.U. Protective effects of valproic acid, a histone deacetylase inhibitor, against hyperoxic lung injury in a neonatal rat model. PLoS ONE 2015, 10, e0126028. [Google Scholar] [CrossRef] [PubMed]
- Costalonga, E.C.; Silva, F.M.; Noronha, I.L. Valproic acid prevents renal dysfunction and inflammation in the ischemia-reperfusion injury model. BioMed Res. Int. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Armeanu, S.; Pathil, A.; Venturelli, S.; Mascagni, P.; Weiss, T.S.; Gottlicher, M.; Gregor, M.; Lauer, U.M.; Bitzer, M. Apoptosis on hepatoma cells but not on primary hepatocytes by histone deacetylase inhibitors valproate and itf2357. J. Hepatol. 2005, 42, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Kalanxhi, E.; Risberg, K.; Barua, I.S.; Dueland, S.; Waagene, S.; Andersen, S.N.; Pettersen, S.J.; Lindvall, J.M.; Redalen, K.R.; Flatmark, K.; et al. Induction of apoptosis in intestinal toxicity to a histone deacetylase inhibitor in a phase i study with pelvic radiotherapy. Cancer Res. Treat. 2017, 49, 374–386. [Google Scholar] [CrossRef] [PubMed]
- Sweet, M.J.; Shakespear, M.R.; Kamal, N.A.; Fairlie, D.P. Hdac inhibitors: Modulating leukocyte differentiation, survival, proliferation and inflammation. Immunol. Cell Biol. 2012, 90, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Grabiec, A.M.; Potempa, J. Epigenetic regulation in bacterial infections: Targeting histone deacetylases. Crit. Rev. Microbiol. 2018, 44, 336–350. [Google Scholar] [CrossRef] [PubMed]
- Nusinzon, I.; Horvath, C.M. Positive and negative regulation of the innate antiviral response and beta interferon gene expression by deacetylation. Mo. Cell. Biol. 2006, 26, 3106–3113. [Google Scholar] [CrossRef] [PubMed]
- Strahl, B.D.; Allis, C.D. The language of covalent histone modifications. Nature 2000, 403, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Kori, Y.; Sidoli, S.; Yuan, Z.F.; Lund, P.J.; Zhao, X.; Garcia, B.A. Proteome-wide acetylation dynamics in human cells. Sci. Rep. 2017, 7, 10296. [Google Scholar] [CrossRef] [PubMed]
- Spange, S.; Wagner, T.; Heinzel, T.; Kramer, O.H. Acetylation of non-histone proteins modulates cellular signalling at multiple levels. Int. J. Biochem. Cell Biol. 2009, 41, 185–198. [Google Scholar] [CrossRef] [PubMed]



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delgado, F.G.; Cárdenas, P.; Castellanos, J.E. Valproic Acid Downregulates Cytokine Expression in Human Macrophages Infected with Dengue Virus. Diseases 2018, 6, 59. https://doi.org/10.3390/diseases6030059
Delgado FG, Cárdenas P, Castellanos JE. Valproic Acid Downregulates Cytokine Expression in Human Macrophages Infected with Dengue Virus. Diseases. 2018; 6(3):59. https://doi.org/10.3390/diseases6030059
Chicago/Turabian StyleDelgado, Félix G., Paola Cárdenas, and Jaime E. Castellanos. 2018. "Valproic Acid Downregulates Cytokine Expression in Human Macrophages Infected with Dengue Virus" Diseases 6, no. 3: 59. https://doi.org/10.3390/diseases6030059
APA StyleDelgado, F. G., Cárdenas, P., & Castellanos, J. E. (2018). Valproic Acid Downregulates Cytokine Expression in Human Macrophages Infected with Dengue Virus. Diseases, 6(3), 59. https://doi.org/10.3390/diseases6030059