Bacterial Pathogen Occurrence and Persistence in Livestock Mortality Biopiles

Abstract
:1. Introduction
2. Materials and Methodology
3. Results
3.1. Slaughterhouse-Residual Media
3.2. Sawdust and Biopile Media
3.3. Soil Media
3.4. Liquid Media
3.5. Persistent Pathogens and Escherichia coli NAR
4. Discussion
4.1. Input Media
4.2. Sawdust and Biopile Media
4.3. Soil Media
4.4. Persistence in Solid Media
4.5. Liquid Media
4.6. Persistent Pathogens and Escherichia coli NAR
5. Summary
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
References
- Whitman, W.B.; Coleman, D.C.; Wiebe, W.J. Prokaryotes: The unseen majority. Proc. Natl. Acad. Sci. USA 1998, 85, 6578–6583. [Google Scholar] [CrossRef]
- Stecher, B.; Hardt, W.-D. The role of microbiota in infectious disease. Trends Microbiol. 2008, 16, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Prescott, L.M.; Harley, J.P.; Klein, D.A. Microbiology, 3rd ed.; Wm. C. Brown Publishers: Dubuque, IA, USA, 1996. [Google Scholar]
- Park, S.Y.; Woodward, C.L.; Kubena, L.F.; Nisbet, D.J.; Birkhold, S.G.; Ricke, S.C. Environmental dissemination of foodborne Salmonella in preharvest poultry production: Reservoirs, critical factors, and research strategies. Crit. Rev. Environ. Sci. Technol. 2008, 38, 73–111. [Google Scholar] [CrossRef]
- Lemunier, M.; Francou, C.; Rousseaux, S.; Houot, S.; Dantigny, P.; Piveteau, P.; Guzzo, J. Long-term survival of pathogenic and sanitation indicator bacteria in experimental biowaste composts. Appl. Environ. Microbiol. 2005, 71, 5779–5786. [Google Scholar] [CrossRef] [PubMed]
- Sobsey, M.D.; Khatib, L.A.; Hill, V.R.; Alocilja, E.; Pillai, S. Pathogens in animal wastes and the impacts of waste management practices on their survival, transport and fate. In Animal Agriculture and the Environment: National Center for Manure and Animal Waste Management White Papers; Rice, J.M., Caldwell, D.F., Humenik, F.J., Eds.; ASABE: St. Joseph, MI, USA, 2006; pp. 609–666. [Google Scholar]
- Turpin, P.E.; Maycroft, K.A.; Rowlands, C.L.; Wellington, E.M.H. Viable but non-culturable Salmonellas in soil. J. Appl. Bacteriol. 1993, 74, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, F.A.; Groves, S.J.; Chambers, B.J. Pathogen survival during livestock manure storage and following land application. Bioresour. Technol. 2005, 96, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Conboy, M.J.; Goss, M.J. Identification of an assemblage of indicator organisms to assess timing and source of bacterial contamination in groundwater. Water Air Soil Pollut. 2000, 129, 101–118. [Google Scholar] [CrossRef]
- Frey, S.D. Spatial distribution of soil organisms. In Soil Microbiology, Ecology, and Biochemistry; Paul, E.A., Ed.; Elsevier: Burlington, MA, USA, 2007; pp. 283–300. [Google Scholar]
- Dapilly, N.; Neyrat, A. Etude des microorganismes pathogènes dans les composts. Rapport de recherche bibliographique CREED. 1999. Available online: http://portailbiblio.ipsl-edu.com/gediweb5/;jsessionid=660489F566F7422719A267FF4E439BA9?0&docId=90141&page=home&displayTab=2&blocOngletId=15645 (accessed on 26 September 2017).
- Gale, P. Risks to farm animals from pathogens in composted catering waste containing meat. Vet. Rec. 2004, 155, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.E., Jr.; Millner, P.D.; Jakubowski, W.; Goldstein, N.; Rynk, R. (Eds.) Contemporary Perspectives on Infectious Disease Agents in Sewage Sludge and Manure; The JG Press, Inc.: Emmaus, UK, 2005. [Google Scholar]
- Van Elsas, J.D.; Ling, T.; Finlay, R.D.; Killham, K.; Trevors, J.T. Microbial interactions in soil. In Modern Soil Microbiology; van Elsas, J.D., Jansson, J.K., Trevors, J.T., Eds.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2007; pp. 177–210. [Google Scholar]
- Islam, M.; Doyle, M.P.; Phatak, S.C.; Millner, P.; Jiang, X. Survival of Escherichia coli O157:H7 in soil and on carrots and onions grown in fields treated with contaminated manure composts or irrigation water. Food Microbiol. 2005, 22, 63–70. [Google Scholar] [CrossRef]
- Jamieson, R.C.; Joy, D.M.; Lee, H.; Kostaschuk, R.; Gordon, R.J. Persistence of enteric bacteria in alluvial streams. J. Environ. Eng. Sci. 2004, 3, 203–212. [Google Scholar] [CrossRef]
- Wilkinson, K.G. The biosecurity of on-farm mortality composting. J. Appl. Microbiol. 2007, 102, 609–618. [Google Scholar] [CrossRef] [PubMed]
- CCME (Canadian Council for Ministers of the Environment). Guidelines for Compost Quality; Report No. PN 1340; CCME: Winnipeg, MB, Canada, 2005. [Google Scholar]
- Jamieson, R.C. An Investigation of the Fate and Transport of Enteric Bacteria in Alluvial Streams. Ph.D. Thesis, University of Guelph, Guelph, ON, Canada, May 2005. [Google Scholar]
- Ahring, B.K. Perspectives for anaerobic digestion. Adv. Biochem. Eng. Biotechnol. 2003, 81, 1–30. [Google Scholar] [PubMed]
- Noonan, M.J.; Harman, H.; Keeley, G.M. An alternative method for reducing numbers of fecal coliform bacteria in slaughterhouse effluent. In Alternative Waste Treatment Systems; Bhamidimarri, R., Ed.; Elsevier Science Publishing Co., Inc.: New York, NY, USA, 1988; pp. 219–229. [Google Scholar]
- Yan, T.; Sadowsky, M.M. Determining sources of fecal bacteria in waterways. Environ. Monit. Assess. 2007, 129, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Health Canada. Guidelines for Canadian Drinking Water Quality: Summary Table. 2008. Available online: http://www.hc-sc.gc.ca/ewh-semt/alt_formats/hecs-sesc/pdf/pubs/water-eau/sum_guide-res_recom/summary-sommaire-eng.pdf (accessed on 1 October 2008).
- CCME (Canadian Council for Ministers of the Environment). Canadian Water Quality Guidelines for the Protection of Agricultural Water Uses: Summary Table. 2005. Available online: http://ceqg-rcqe.ccme.ca/ (accessed on 1 October 2008).
- WHO (World Health Organization). Guidelines for the Safe Use of Wastewater, Excreta, and Greywater; Wastewater Use in Agriculture; WHO: Geneva, Switzerland, 2006; Volume 2. [Google Scholar]
- Michitsch, R.; Gordon, R.; Jamieson, R. A system to evaluate livestock mortality biopiles. J. Appl. Agric. Res. 2010, 3, 85–101. [Google Scholar]
- Webb, K.T.; Thompson, R.L.; Beke, G.J.; Nowland, J.L. Soils of Colchester County, Nova Scotia: Nova Scotia Soil Survey; Report No. 19. Land Resource Research Centre Contribution No. 85-45; Minister of Supply and Services Canada: Ottawa, ON, Canada, 1991. [Google Scholar]
- Michitsch, R.; Jamieson, R.; Gordon, R.; Stratton, G.; Lake, C. Bacterial pathogen indicator transport from livestock mortality biopiles. J. Environ. Qual. 2011, 44, 1355–1365. [Google Scholar] [CrossRef] [PubMed]
- HACH Company. The Use of Indicator Organisms to Assess Public Water Safety. Technical Information Series—Booklet No. 13. 2000. Available online: http://www.hach.com/fmmimghach?/CODE%3AL7015547%7C1 (accessed on 1 December 2007).
- USEPA. USEPA Method 1603: Results of the Interlaboratory Validation of EPA Method 1603 (Modified mTEC) for E. coli in Wastewater Effluent; ID# EPA-821-R-04-009; United States Environmental Protection Agency, Office of Water: Washington, DC, USA, 2004.
- USEPA. USEPA Method 1600: Enterococci in Water by Membrane Filtration Using Membrane-Enteroccus Indoxyl-β-d-glucoside Agar (mEI); ID# EPA-821-R-04-023; United States Environmental Protection Agency, Office of Water: Washington, DC, USA, 2005.
- Eaton, A.D.; Clesceri, L.S.; Greenberg, A.E. Standard Methods for the Examination of Water and Wastewater; APHA: Washington, DC, USA; AWWA: Denver, CO, USA; WEF: Alexandria, VA, USA, 1995. [Google Scholar]
- Michitsch, R.C. Bacterial Pathogen Fate in Slaughterhouse-residual Biopiles. Ph.D. Thesis, Dalhousie University, Halifax, NS, Canada, July 2009. [Google Scholar]
- Zuberer, D.A. Recovery and enumeration of viable bacteria. In Methods of Soil Analysis Part 2: Microbiological and Biochemical Properties; Bigham, J.M., Mickelson, S.H., Eds.; Soil Science Society of America, Inc.: Madison, WI, USA, 1994; pp. 127–129. [Google Scholar]
- Andrews, W.H.; Hamack, T.S.E. Microbiological methods. In Official Methods of Analysis of AOAC International; Horwitz, W., Latimer, G.W., Jr., Eds.; Association of Official Analytical Chemists International: Gaithersburg, MD, USA, 2005. [Google Scholar]
- McSpadden-Gardener, B.B. Statistical analyses of microbiological and environmental data. In Modern Soil Microbiology; van Elsas, J.D., Jansson, J.K., Trevors, J.T., Eds.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2007; pp. 553–583. [Google Scholar]
- SAS. The SAS System for Windows: Release 9.1.3; SAS Institute: Cary, NC, USA, 2008. [Google Scholar]
- Holt, J.G. (Ed.) Bergey’s Manual of Determinative Bacteriology; Williams and Wilkins: Baltimore, MD, USA, 1984; Available online: https://archive.org/details/bergeysmanualofd1957amer (assessed on 26 September 2017).
- Holt, J.G. (Ed.) Bergey’s Manual of Systematic Bacteriology; Williams and Wilkins: Philadelphia, PA, USA, 1984; Available online: https://searchworks.stanford.edu/view/318937 (assessed on 26 September 2017).
- Dworkin, M. (Ed.) The Prokaryotes: A Handbook on the Biology of Bacteria, 3rd ed.; Springer Science & Business Media: New York, NY, USA, 2006; Volume 1–7. [Google Scholar]
- Winfield, M.D.; Groisman, E.A. Role of nonhost environments in the lifestyles of Salmonella and Escherichia coli. Appl. Environ. Microbiol. 2003, 69, 3687–3694. [Google Scholar] [CrossRef] [PubMed]
- Namura, M.; Hijikata, T.; Miyanaga, K.; Tanji, Y. Detection of Escherichia coli with fluorescent labeled phages that have a broad host range to E. coli in sewage water. Biotechnol. Prog. 2008, 24, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Hazen, T. Fecal coliforms as indicators in tropical waters: A review. Environ. Toxicol. 1988, 3, 461–477. [Google Scholar] [CrossRef]
- Nybroe, O.; Brandt, K.K.; Nicolaisen, M.H.; Sörensen, J. Methods to detect and quantify bacteria in soil. In Modern Soil Microbiology; van Elsas, J.D., Jansson, J.K., Trevors, J.T., Eds.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2007; pp. 283–316. [Google Scholar]
- Hartz, A.; Cuvelier, M.; Nowosielski, K.; Bonilla, T.D.; Green, M.; Esiobu, N.; McCorquodale, D.S.; Rogerson, A. Survival potential of Escherichia coli and Enterococci in subtropical beach sand: Implications for water quality managers. J. Environ. Qual. 2008, 37, 898–905. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Reuter, G.; Inglis, D.; Larney, F.J.; Alexander, T.W.; Guan, J.; Stanford, K.; Xu, Y.; McAllister, T.A. A biosecure composting system for disposal of cattle carcasses and manure following infectious disease outbreak. J. Environ. Qual. 2009, 38, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Kandeler, E. Physiological and biochemical methods for studying soil biota and their function. In Soil Microbiology, Ecology, and Biochemistry; Paul, E.A., Ed.; Elsevier: Burlington, MA, USA, 2007; pp. 53–83. [Google Scholar]
- Joy, D.; Lee, H.; Reaume, C.; Whiteley, H.; Zelin, S. Microbial contamination of subsurface tile drainage water from field application of liquid manure. Can. Agric. Eng. 1998, 40, 153–160. [Google Scholar]
- Dean, D.M.; Foran, M.E. The effect of farm liquid waste application on tile drainage. J. Soil Water Conserv. 1992, 5, 368–369. [Google Scholar]
- Harvey, R.W.; Harms, H. Tracers in groundwater: Use of microorganisms and microspheres. In Encyclopedia of Environmental Microbiology; Bitton, G., Ed.; Wiley-Interscience: New York, NY, USA, 2002; pp. 3194–3202. [Google Scholar]
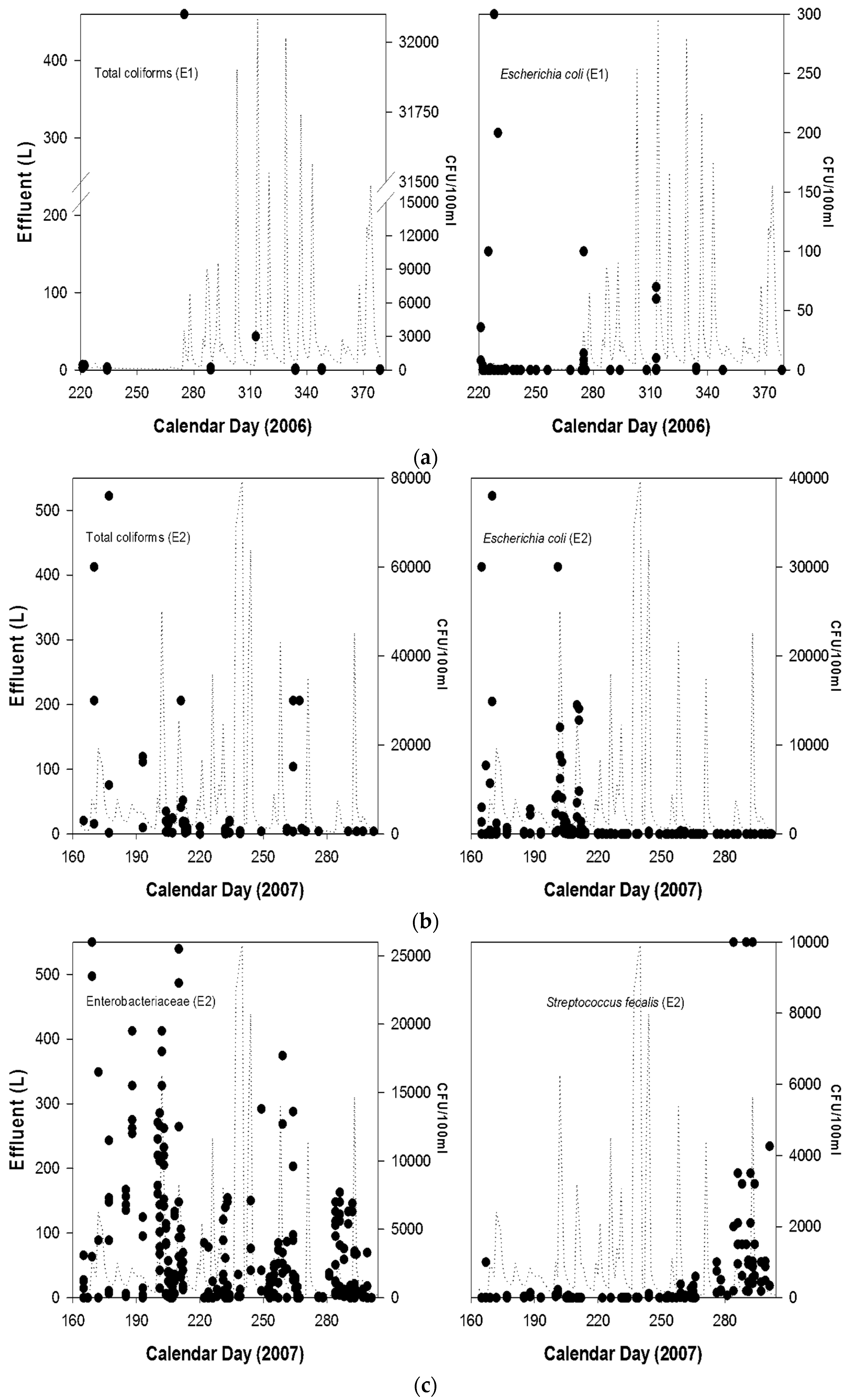
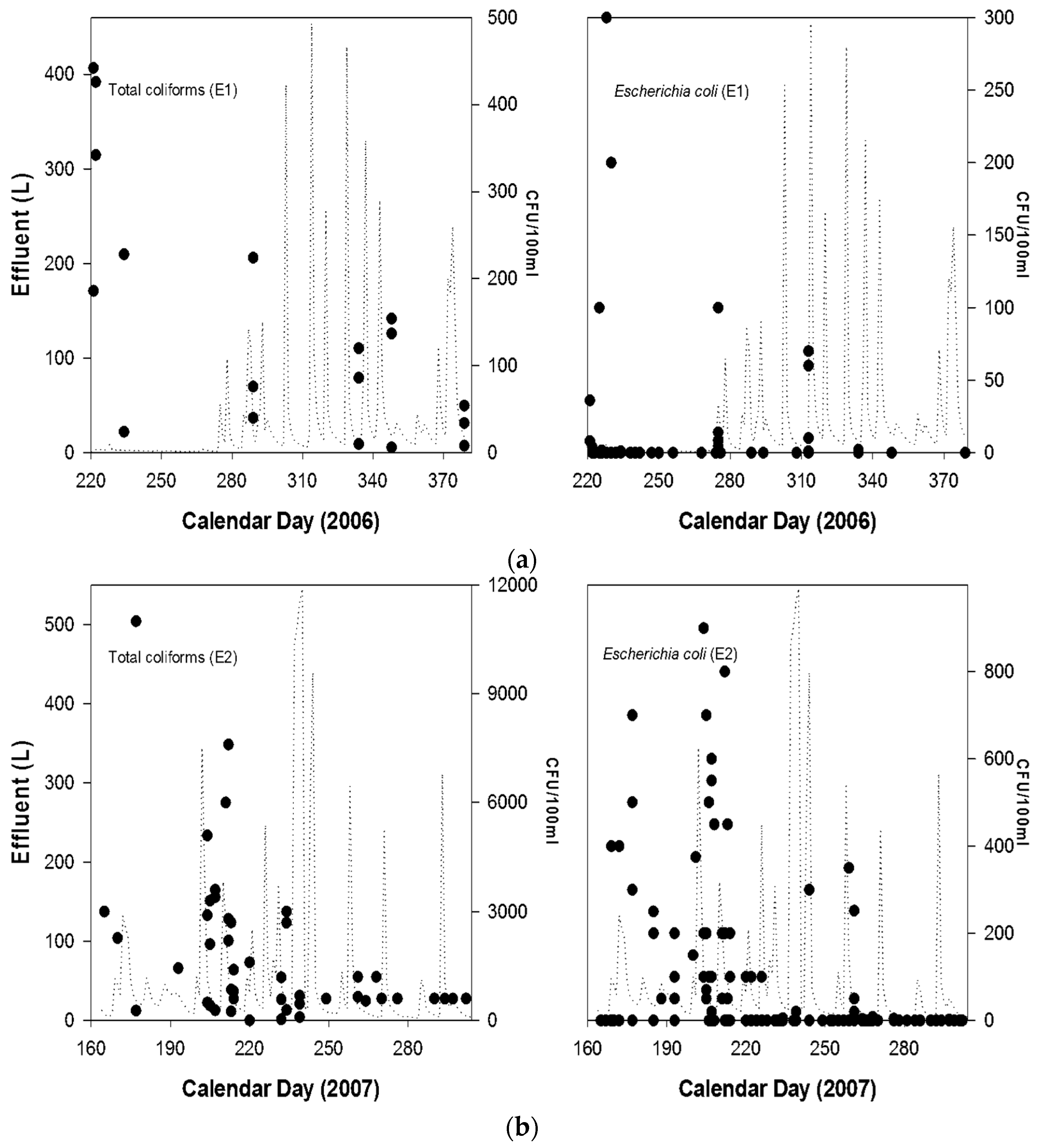

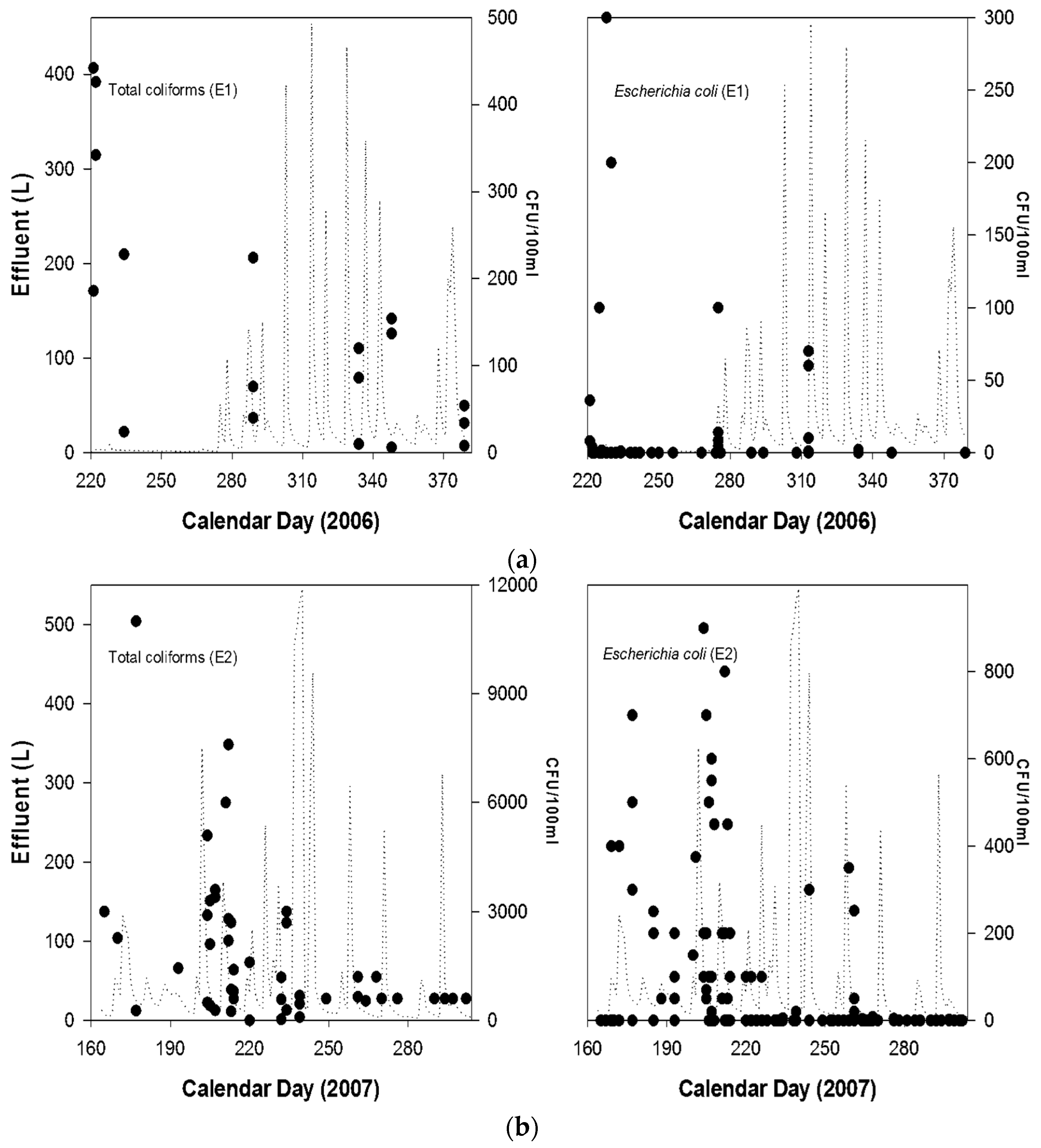
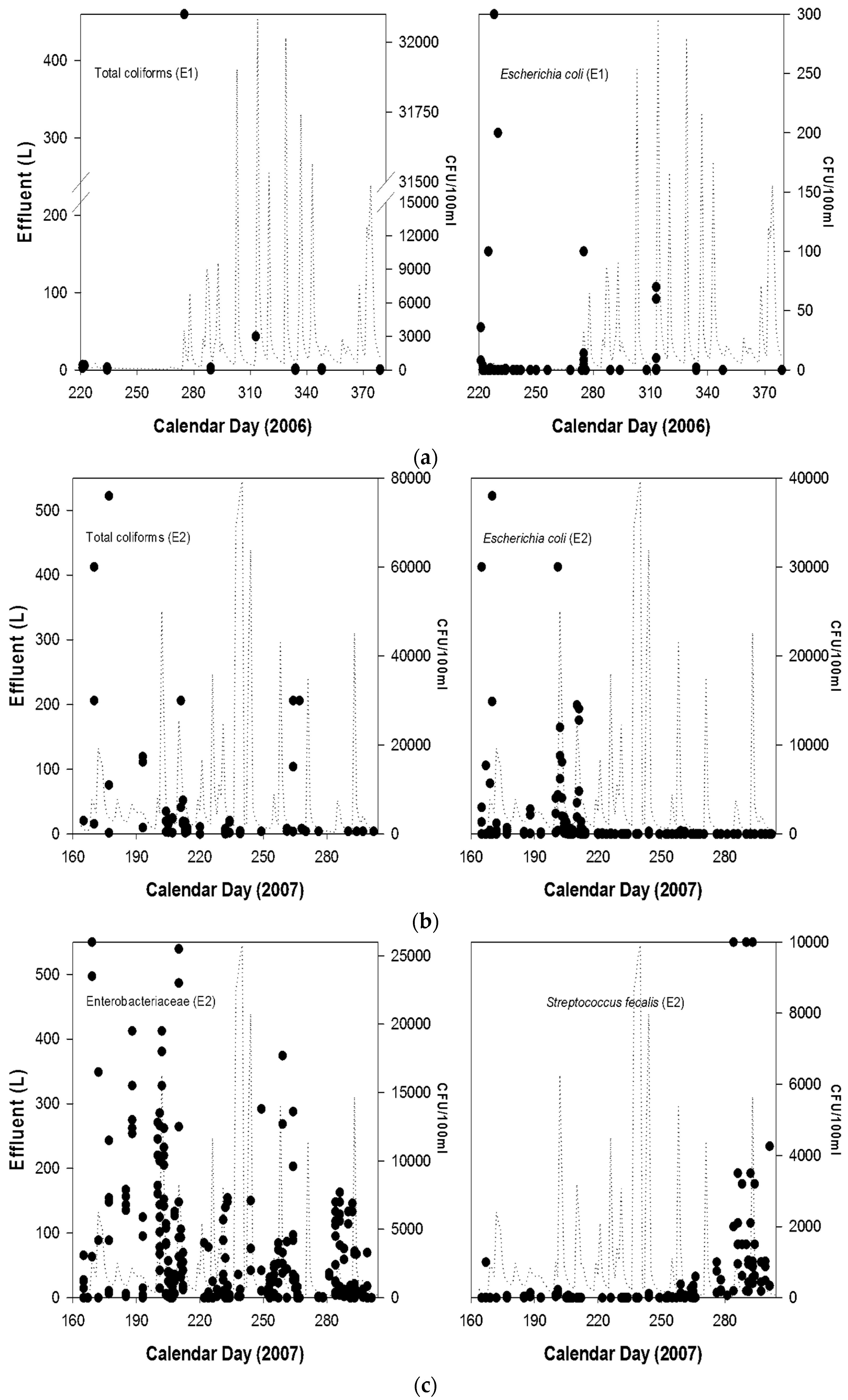{kind=link}
{kind=link}
| Time | n | TC † (cfu·g−1) | EC (cfu·g−1) | EB (cfu·g−1) | SF (cfu·g−1) |
|---|---|---|---|---|---|
| Sawdust/Biopile | |||||
| Initial ‡ | 2 | 4.8 × 103 ± 5.8 × 102 | 0 ± 0 | - | - |
| Final | 27 | 3.0 × 101 ± 1.5 × 102 | 0 ± 0 | 8.1 × 101 ± 4.2 × 102 | 0 ± 0 |
| Soil | |||||
| Initial | 2 | 1.0 × 102 ± 1.4 × 102 | 0 ± 0 | - | - |
| 10 cm § | 18 | 1.6 × 102 ± 5.0 × 102 | 0 ± 0 | 3.3 × 102 ± 8.2 × 102 | 0 ± 0 |
| 20 cm § | 18 | 5.3 × 102 ± 1.5 × 103 | 0 ± 0 | 5.9 × 102 ± 1.7 × 103 | 0 ± 0 |
| Slaughterhouse residuals | |||||
| Initial | 5 | 5.1 × 103 ± 3.5 × 103 | 6.3 × 102 ± 1.3 × 103 | - | - |
| Time | n | TC † (cfu·g−1) | EC (cfu·g−1) | EB (cfu·g−1) | SF (cfu·g−1) |
|---|---|---|---|---|---|
| Sawdust/Biopile | |||||
| Initial ‡ | 4 | 0 ± 0 | 0 ± 0 | 2.2 × 103 ± 2.2 × 103 | 1 ± 1 |
| Final | 18 | 4.8 × 102 ± 1.4 × 103 | 0 ± 0 | 1.8 × 103 ± 2.1 × 103 | 9.7 × 101 ± 1.1 × 102 |
| Soil | |||||
| Initial | 5 | 1.0 × 105 ± 1.4 × 105 | 0 ± 0 | 8.0 × 105 ± 1.1 × 106 | 0 ± 0 |
| 10 cm § | 9 | 1.1 × 104 ± 3.3 × 104 | 0 ± 0 | 1.4 × 105 ± 3.0 × 105 | 0 ± 0 |
| 20 cm § | 9 | 0 ± 0 | 0 ± 0 | 3.0 × 105 ± 7.5 × 105 | 0 ± 0 |
| Slaughterhouse residuals | |||||
| Initial | 6 | 1.5 × 107 ± 2.7 × 107 | 4.3 × 105 ± 7.0 × 105 | 5.1 × 108 ± 7.8 × 108 | 9.1 × 101 ± 1.4 × 102 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michitsch, R.; Gordon, R.; Jamieson, R.; Stratton, G. Bacterial Pathogen Occurrence and Persistence in Livestock Mortality Biopiles. Resources 2017, 6, 49. https://doi.org/10.3390/resources6040049
Michitsch R, Gordon R, Jamieson R, Stratton G. Bacterial Pathogen Occurrence and Persistence in Livestock Mortality Biopiles. Resources. 2017; 6(4):49. https://doi.org/10.3390/resources6040049
Chicago/Turabian StyleMichitsch, Robert, Robert Gordon, Rob Jamieson, and Glenn Stratton. 2017. "Bacterial Pathogen Occurrence and Persistence in Livestock Mortality Biopiles" Resources 6, no. 4: 49. https://doi.org/10.3390/resources6040049
APA StyleMichitsch, R., Gordon, R., Jamieson, R., & Stratton, G. (2017). Bacterial Pathogen Occurrence and Persistence in Livestock Mortality Biopiles. Resources, 6(4), 49. https://doi.org/10.3390/resources6040049