Abstract
The amphibian Xenopus has long been a comparative model system of choice for a number of different biological research areas, including immunology. Specifically, the evolutionary distance between amphibians and mammals, including humans, allows for the study of both species-specific adaptations, as well as conserved features of the immune system. Furthermore, the Xenopus genus includes species with multiple levels of polyploidy, thereby providing a unique model to study whole genome duplication and its effects thereof on individual genes. To better exploit this amphibian model, the development and innovative applications of novel research tools have been a priority. In this regard, recent advances in adapting the transgenesis approach to Xenopus have allowed for in vivo studies of the impact of loss and gain of function of specific genes at the level of the whole organism, further enhancing the potential uses of Xenopus as an important biomedical model system. This review highlights some of the major uses and applications of the Xenopus model.
1. Introduction
For several decades, the African amphibian species of the Xenopus genus, Xenopus laevis and X. tropicalis have been the model system of choice for developmental, embryological, and immunological studies (reviewed in [1,2]). In addition, Xenopus is emerging as an excellent model system for toxicology, neurobiology, regenerative biology, and other areas of biomedical science [3]. Indeed, the use of Xenopus in various areas of biomedical research including tumor biology, transplantation biology, self-tolerance, and auto immunity is steadily increasing and has significantly contributed to advances in these fields. Importantly, these advances have gone hand-in-hand with innovation and adaptation of new techniques. This development of novel research tools has proved critical to further empowering Xenopus as a model system and keeping it at the forefront of biomedical research.
A key advantageous feature of Xenopus is its external development free of maternal influence that allows easy access to experimental manipulation during all stages of development. The transparency of tadpoles permits easy visualization of certain organs, such as the thymus. Moreover, Xenopus are easy to maintain and they breed all year, with each mating producing a high number of progeny. Furthermore, the recently completed whole genome sequencing and annotations of X. tropicalis [4], and more recently its sister species X. laevis [5], has revealed a remarkable conservation of gene synteny with the human genome [4]. The availability of this genetic information has greatly facilitated the identification of novel, potentially immunologically relevant genes. In addition, the availability of major histocompatibility (MHC)-defined X. laevis animal inbred strains, gynogenetic clones, and cell lines, coupled with a powerful and continuously improving reverse genetics approach provide the basic tools for addressing fundamental questions about the ontogeny and phylogeny of the immune system [6]. This review will focus on some of these features that makes Xenopus an ideal comparative model system for biomedical and immunological studies, with particular focus on some recent advances in reverse genetics approaches aimed specifically at the study of in vivo loss and gain of function of immunologically-relevant genes.
2. Xenopus as a Model for Biomedical Research
The immune system of Xenopus is one of the most comprehensively studied outside of mammals and chicken (reviewed in [1,2]), and the wealth of information already available on this topic, combined with established experimental systems, provides a solid basis to further developing this unique biomedical model. Importantly, the evolutionary and genetic distance of Xenopus from humans permits one to distinguish species-specific adaptations from more conserved features of both the innate and adaptive immune systems. The immune system of Xenopus is fundamentally similar to those of mammals. Although Xenopus lacks the functional equivalent of mammalian lymph nodes, it has a thymus, a spleen, and localized leukocyte accumulation in in the periphery of the liver, the kidneys, and along the intestine [2]. Moreover, most cell types of lymphoid origin present in mammals are also found in Xenopus including B cells, T cells, and NK cells, as well as important hallmarks of adaptive immunity like recombination activated genes (RAG)-mediated rearranging immunoglobulin receptors (Ig) and T cell receptors (TCR), MHC class I and II genes, and activation-induced cytidine deaminase (AID) [1].
2.1. Xenopus as a Model to Study Immune Tolerance
Xenopus like other anuran amphibians have two physiologically and phenotypically distinct larval and adult life stages. The immune system at each of these stages also exhibits significant differences as well as similarities. The main developmental steps of the Xenopus immune system are summarized in Table 1. For example, although larvae have B and T cells, their total larval Ig and TCR repertoire is less heterogeneous than in adults, and overall both B and T cell responses are weaker (reviewed in [7,8]). Larvae are immunocompetent and capable of rejecting MHC mismatched skin grafts [9]. However, adults are also able to reject skin grafts from MHC identical but minor histocompatibility (H) locus disparate donors, whereas larvae typically tolerate such minor H-locus incompatible grafts [10]. This form of larval-induced allotolerance is antigen-specific and persists in the animal after metamorphosis as demonstrated by permanent survival of a second graft from the same minor H-antigen mismatched donor [11]. However, although allotolerance is established, there is still lymphocyte infiltration in the allograft, something that is not observed with autografts. Moreover, splenocytes co-cultured with irradiated donor splenocytes undergo proliferation [12]. This suggests that the allograft is recognized by recipient immune cells as non-self. Such immune allorecognition without rejection implies some state of anergy, adoptive tolerance, and/or the influence of regulatory cells such as regulatory T cells (Treg), which in mammals represent an subset of CD4 T cells critical for suppressing or modulating immune responses.

Table 1.
Summary of the main developmental steps of the Xenopus immune system.
| Young tadpoles | Stage 35–40, 5–8 mm long, 2–7 days post-fertilization |
|
| Pre-metamorphic tadpoles | Stage 50–57, 30–50 mm long, 2–6 weeks post-fertilization |
|
| Metamorph | Stage 58–66, 40–60 mm long, 6–8 weeks post-fertilization |
|
| Young adults | 40–60 mm long, 3–12 months-old |
|
During metamorphosis the immune system undergoes extensive remodeling and a significant reduction in both thymocytes and splenic lymphocytes occurs during this stage [2]. This lymphocyte depletion is followed by a second wave of precursor cell immigration as well as T cell differentiation and renewal [13]. These distinct periods of thymocyte differentiation (first during embryogenesis and later during metamorphosis) take place in different environments as the whole organism undergoes remodeling and differential gene expression [2]. Indeed, at this time many new adult-type proteins are expressed that could potentially be recognized as non-self by the larval immune system. As such, an additional negative selection process is likely to occur to achieve new self-tolerance. Furthermore, MHC class I expression is differentially regulated with sub optimal expression of classical MHC class Ia (class Ia) genes in the pre-metamorphic larval stage [14,15]. Using the anti-class Ia monoclonal antibody (mAb), TB17, MHC class Ia surface expression is first detected, at low levels, during pre-metamorphic developmental stages 56–58 and subsequently increases during metamorphosis [16]. Similarly larval and adult stages differ in their MHC class II protein expression. In tadpoles, class II surface expression is observed on the thymic epithelium, (but not on thymocytes) and on B cells and macrophages with no detectable class II protein expression on peripheral T cells [17]. By comparison, all adult lymphocytes stain positive with MHC class II specific mAbs. Hence, for both MHC class I and class II molecules there is a sharp difference in expression pattern between larvae and adults suggesting that different T cell selection processes occur during the two different life stages. This is important as it provides a unique model to study new potential links between the regulation of MHC expression and immune tolerance. In this context, another attribute of the Xenopus model worth mentioning is the relative ease with which metamorphosis can be experimentally accelerated with the synthetic thyroid hormone (e.g., T4), or delayed with goitrogen (e.g., sodium perchlorate), added in the water of pre-metamorphic tadpoles [16]. Additionally, T cell deficient Xenopus can be generated by thymectomy at early developmental stages (the thymus is easily visible and accessible in the transparent tadpoles) resulting in an adult frog with no peripheral T cells and severely deficient T cell responses (e.g., they fail to reject MHC-disparate grafts) [18].
2.1.1. Xenopus as a Model to Study Skin Graft Tolerance and Rejection
One of the advantages of using Xenopus as a biomedical model is the availability of MHC-defined inbred strains and clones as well as MHC-defined transplantable lymphoid tumor cell lines (Table 2). These inbred clones, strains, and tumor cell lines provide the basis for a powerful transplantation system to study the immune response during skin allografting and tumor transplantation in vivo. Several fibroblast cell lines derived from these strains and clones have also been established (Table 2). These resources (inbred strains and clones) also permit classic adoptive cell transfer as well as cells and tissue transplantation between larvae and adults [19].
A first group of MHC-defined animals include genetically defined MHC homozygote inbreed animals such as the J (j/j MHC haplotype [20]) and the F (f/f MHC haplotype [21]) strains. The J strain was inbred for 25 generations in Japan followed by a further six generations in our Xenopus research resource. Skin grafting among J siblings frogs are not rejected, indicating that this strain is truly inbred. Although the F strain is MHC homozygous, it is not fully inbred and minor H Ag are still segregating. The preservation of these various animal strains require a continuous maintenance of good founder animals and several other strains (i.e., G, R) are currently threatened because of a difficulty in maintaining a sufficient number of breeders. The conservation of these strains is vitally important. For example, J and F strains have proved extremely useful for the X. laevis full genome sequence project [22].
Table 2.
Main X. laevis strains and clones with their respective cell lines.
| Definition of X. laevis strains | Name (MHC genotype) | Cell lines |
|---|---|---|
| Partially inbred, MHC homozygous strains | F (f/f) | Thymic lymphoid tumor cell lines |
| ||
| J (j/j) | Fibroblast cell lines | |
| ||
| Isogenetic laevis/gilli (LG) clones. | LG-15 (a/c) | Thymic lymphoid tumor cell line |
| Same heterozygous MHC but different minor H loci |
| |
| LG-6 (a/c) | Fibroblast cell line | |
|
A second group of MHC defined animals are isogenetic clones that are interspecies hybrids between X. laevis and X. gilli (LG; [23]), in which progenies are maintained by gynogenesis. LG females have the unique ability to produce a mixture of large eggs that are diploid and smaller eggs that are aneuploid. Normal fertilization of diploid eggs will produce triploid animals, while aneuploid eggs are sterile. It is possible, however, to produce diploid clonal progenies from LG females by gynogenesis. During this process, the diploid LG eggs are activated by UV-irradiated sperm from an X. laevis outbred male and thus the spermatozoid DNA does not contribute any genetic material to the offspring. Several different isogenetic LG clones have been MHC typed and include strains that have identical heterozygous MHC haplotypes (LG-6, LG-15, and LG-7, a/c haplotype) but differ at several minor histocompatibility (H) loci. Other strains are MHC-disparate including the LG-6 (a/c), LG-3 (b/d), and LG-5 (b/c) [1]. These clonal animals provide a genetic system that permits the study of allograft rejection against both major and minor H-antigens. Skin grafting in Xenopus is a well-established and straightforward technique and rejection is easily measured visually by the destruction of silvery iridophores in the graft (Figure 1) and described in detail in [24].
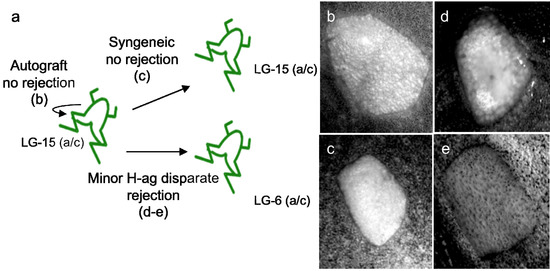
Figure 1.
X. laevis skin graft transplantation system. (a) Schematic representation of the X. laevis major and minor H-antigen skin transplantation system. MHC-typed LG clones with the same MHC haplotype but different minor H-antigens (LG-15 and LG-6) provide a system to study allograft rejection. Examples of skin graft rejection (b)–(e). (b) Autograft from the same LG-15 individual two months post-transplantation. (c) Isograft from one LG-15 individual to another LG-15, three months post-transplantation. In both (b) and (c) no rejection is observed and the entire graft appears silvery due to iridophores. In (d) a LG-6 graft to a LG-15 recipient is beginning to show rejection 10 days after transplantation, (loss of iridophores). In addition, in (e) full rejection has occurred 35 days post-transplantation and the entire graft appears dull.
Figure 1.
X. laevis skin graft transplantation system. (a) Schematic representation of the X. laevis major and minor H-antigen skin transplantation system. MHC-typed LG clones with the same MHC haplotype but different minor H-antigens (LG-15 and LG-6) provide a system to study allograft rejection. Examples of skin graft rejection (b)–(e). (b) Autograft from the same LG-15 individual two months post-transplantation. (c) Isograft from one LG-15 individual to another LG-15, three months post-transplantation. In both (b) and (c) no rejection is observed and the entire graft appears silvery due to iridophores. In (d) a LG-6 graft to a LG-15 recipient is beginning to show rejection 10 days after transplantation, (loss of iridophores). In addition, in (e) full rejection has occurred 35 days post-transplantation and the entire graft appears dull.
2.1.1.1. The Xenopus Immuno-Cancer Model
Xenopus has proven to be instrumental for exploring novel and innovative approaches for cancer immunotherapy. The high degree of functional conservation of the Xenopus and human immune systems including anti-tumor immune effector cell, MHC class Ia, and non-classical MHC class Ib (class Ib) molecules makes it a highly relevant and attractive non-mammalian comparative model (reviewed in [1,2,19,25,26]). In fact, X. laevis represents the only amphibian species in which true lymphoid tumors have been discovered and tumorigenic transplantable tumor cell lines derived from these spontaneously occurring thymic tumors were established during the early 1990s [27,28]. A number of these tumor cell lines were derived from the partially inbred F frogs (ff-2 and B3B7); while others originated from cloned LG-15 frogs (15/0 and 15/40). These tumor cell lines are non-adherent, grow continuously at 27 °C with a generation time of 18–24 h, display a T/B dual phenotype reminiscent of rare lymphocytic leukemia, and can be easily traced in vivo and ex vivo by the expression of the immature thymocyte marker CTX [1,29].
The distinct tumor cell lines differ in their tumorigenicity [25,30]. For example, the 15/0 tumor, derived from the LG-15 clone, is highly tumorigenic and grows in both MHC compatible adults and in larvae that share at least one MHC haplotype with the tumor (a/c) (Figure 2a,b). In contrast, the ff-2 does not grow in adults, regardless of genetic background and it only grows in larvae that share at least one MHC haplotype (f). Notably, these different tumor cell lines differ in their expression of MHC class Ia as well as class Ib proteins [28,31,32]. This has particular bearing as both class Ia and class Ib MHC molecules play important roles in immune surveillance and tumorigenesis [33]. Recently, using short hairpin RNA (shRNA), we have generated a number of different 15/0 transformed cell lines with either a global MHC class Ib silencing or specific silencing of defined class Ib molecules. Using this approach, we were able to demonstrated that a global down regulation of class Ib on 15/0 tumor cells leads to augmented resistance to unconventional cytotoxic T cell-mediated killing and higher susceptibility to NK-like mediated killing, and resulted in increased tumorigenicity in vivo [31].
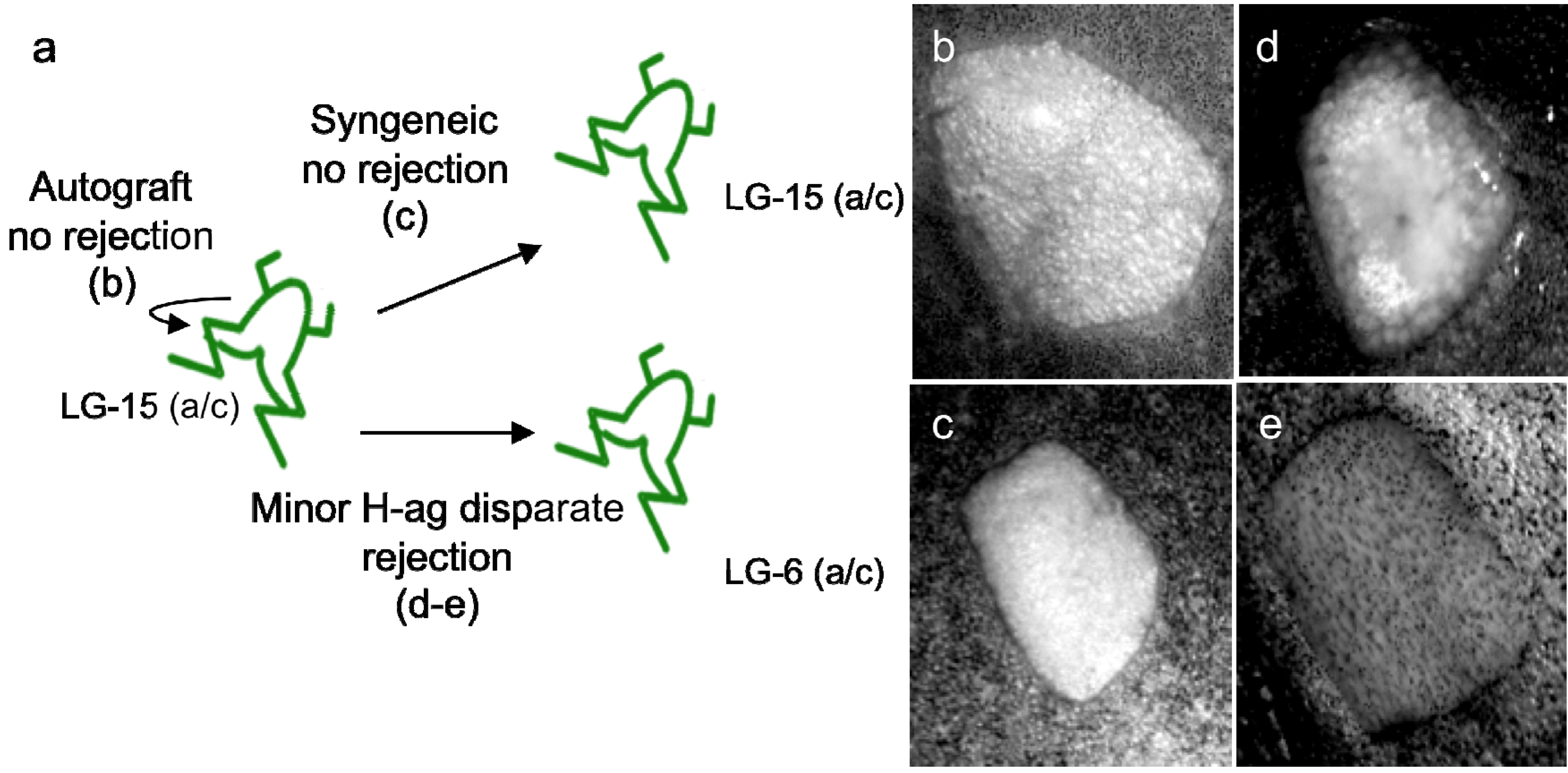
Figure 2.
15/0 tumor growth in LG-15 X. laevis tadpoles. (a) and (b) 15/0 tumor cells (100,000 viable cells) were injected intra-peritoneal into developmental stage 53 (25 days post fertilization) LG-15 tadpoles and the tumor was allowed to establish. An example of tumor growth in one individual over three weeks is shown in (a) and (b); (c) 15/0 tumor cells (100,000 viable cells) were embedded in collagen are grafted under the transparent skin on each dorsal side of a three week-old (developmental stage 53) LG-15 tadpoles and the tumor was allowed to establish. Two 15/0 tumor grafts 10 days post grafting is shown. Arrows indicate areas of tumor growth.
Figure 2.
15/0 tumor growth in LG-15 X. laevis tadpoles. (a) and (b) 15/0 tumor cells (100,000 viable cells) were injected intra-peritoneal into developmental stage 53 (25 days post fertilization) LG-15 tadpoles and the tumor was allowed to establish. An example of tumor growth in one individual over three weeks is shown in (a) and (b); (c) 15/0 tumor cells (100,000 viable cells) were embedded in collagen are grafted under the transparent skin on each dorsal side of a three week-old (developmental stage 53) LG-15 tadpoles and the tumor was allowed to establish. Two 15/0 tumor grafts 10 days post grafting is shown. Arrows indicate areas of tumor growth.
These experiments point to the critical role that class Ia and class Ib regulation or rather dysregulation plays during immune evasion and tumorigenesis. Although a number of studies have shown that class Ia and class Ib dysregulation plays key roles during tumorigenesis and that expression of certain class Ib genes have prognostic value, the underlying mechanisms governing class Ib immune functions are still poorly understood [34,35]. Notably, in this instance as well, the distinct immunocompetence of larvae and adults provides unique approaches to investigate, in vivo, the possible influence of the immune system on the selection of different tumors. For example, since class Ia function is suboptimal in tadpoles, the involvement of class Ib molecules in larval anti-tumor immune response may be different from that of class Ia competent adults. We have recently developed a novel semi-solid tumor assay to visualize in real-time immune responses within the tumor tissue. This consists in embedding 15/0 tumor cells in a collagen matrix, and then to graft the semi-solid tumor under the dorsal transparent skin of an LG-15 tadpole recipient (Figure 2c). Hence, Xenopus is a suitable model for further elucidating specific fundamental mechanisms underlying the observed role of class Ib molecules in tumor immunity, thereby contributing to the innovation and design of more efficient immunotherapeutic approaches to control cancer.
In summary, the availability of MHC-defined clones, inbred MHC homozygote animal strains, and cell lines coupled with a well-established transplantable tumor model, provides a unique and powerful system for studying many aspects of the immune system. All of these animal strains, cell-lines, and tools are maintained and available through the NIH funded X. laevis research resource for immunobiology. This facility also provides a number of different reagents including mAbs, DNA probes, cDNA, and genomic libraries. More information about these tools can be found at [6].
3. Reverse Genetics
Transgenesis in Xenopus is well established and has been successfully implemented by developmental biologists for close to three decades. The Xenopus embryo has numerous advantages including its large size, high amenability to surgical manipulations, external development, and rapid development (a tadpole develops within a few days after fertilization). Moreover, the early development, patterning and morphogenesis of the developing embryo are well defined. Over the years, many different strategies to generate transgenic Xenopus have been developed and optimized and the techniques used are continuously evolving and expanding (reviewed in [36]). Each of these strategies is dependent on specific molecular and technical features, and each offers its own set of advantages and potential drawbacks. In many cases, the method of choice ultimately depends on the goal and applications of the particular study. The initial and most broadly used technique for generating transgenic Xenopus is based on restriction-mediated integration (REMI) and has proven to be highly effective in generating non-mosaic founder animals [37,38]. The REMI method relies upon incorporation of a linear DNA template into the host genome prior to cell division and requires integration of the transgene into the sperm nuclei, which are then transplanted into unfertilized eggs using an infusion pump, thereby generating a transgenic embryo.
3.1. Optimized Transgenesis for Immunological Studies in Xenopus
Since one of the main advantages of Xenopus as a model for biomedical research is the availability of MHC defined cloned animal strains (i.e., LG clones discussed in detail in Section 2.1.1), the generation of MHC-defined, rather than outbred transgenic strains for immunological studies, was imperative. However, the REMI approach is not suitable when it comes to generating transgenic X. laevis MHC-defined isogeneic clones, as these LG clones are generated by gynogenesis where spermatozoids do not contribute any genetic material to the progeny. To date, several techniques that are not dependent on integration of the transgene into the sperm nucleus are currently in use, including the phiC31 integrase [39], transposon-mediated integration [40], and the I-SecI meganuclease [41,42,43]. All three techniques have been shown to be successful in generating transgenic Xenopus although with varying degrees of efficiency. These three different techniques are briefly discussed in the following section.
The phiC31 integration system is based on bacteriophage integrase-mediated integration of the transgene into the target genome. The bacteriophage phiC31 encodes an integrase enzyme that catalyzes recombination between a 39 bp long phage attachment site (attP) and a 34 bp long bacterial attachment site (attB). The phiC31 integrase enzyme is relatively promiscuous and recognizes so-called pseudo-attachment sites that are present in many different organisms [39,44,45]. These pseudo-attachment sites can share as little as 24% homology with the phage attP site and still allow successful recombination. Hence, transgenesis using the phiC31 takes advantage of the attB site to mediate integration of the transgene into pseudo-attP sites in the Xenopus genome. The integrase mRNA and the plasmid carrying the transgene flanked by attB sites are co-injected into the fertilized embryo. To minimize chromatin-positioning effects, and prevent distant enhancers from acting on the transgene promoter region, the transgene is also flanked by HS4 insulator sequences [44].
Transposon-mediated integration is based on co-injection of the fertilized embryo with the transposase enzyme transcript and a plasmid containing the transgene flanked by terminal-repeat elements recognized by the transposase. One advantage of the transposon-mediated approach is that the integrated DNA does not include any plasmid vector sequences and due to the precise integration of the transgene it is easy to determine the site of integration using anchor based PCR [46]. However, a drawback of this approach is that the use of transposable elements commonly results in animals with a high degree of mosaic expression of the transgene. In addition, the transgene integration from generation to generation is not very stable. The most commonly used transposons in Xenopus transgenesis are the Sleeping Beauty (SB, [40]) and the Tol2 transposon systems [47]. Several other transposable elements that have been used successfully in other systems could potentially be applied to Xenopus. These include the amphibian-derived transposable element, Frog Prince, which was reconstructed from an inactivated transposable element in the frog, Rana pipiens [48].
The transgenic method using I-Sec1 meganuclease was originally developed in the medaka fish (Oryzias latipes) and later adapted to Xenopus [41,43]. I-SecI, originally isolated from Saccharomyces cerevisiae, is a commercially available meganuclease with an 18 bp recognition sequence. To achieve integration in the genome, the transgene is flanked by the 18 bp recognition sequence. The plasmid is digested with the I SecI meganuclease using standard techniques and the whole mixture is injected into the fertilized egg. The precise mechanism by which the I SecI meganuclease mediates transgene integration is currently unknown as there are no known endogenous I-SecI sites in either the X. laevis or the X. tropicalis genomes.
These last three transgenic techniques were successfully applied to isogenetic LG-6 and LG-15 Xenopus clones, albeit with different efficiency and varying degree of mosaicism [49]. In fact, the I-SecI meganuclease method clearly resulted in the highest transgenesis efficiency (35%–50%) compared to 12%–35% and 30%–40% for SB transposase and phiC31 integrase, respectively. In addition I-SecI meganuclease resulted in the highest survival rate at hatching compared to the two other methods. Importantly, the I-SecI meganuclease mediated transgenesis consistently resulted in strong and uniform non-mosaic expression of the fluorescent GFP reporter gene in more than 80% of F0 animals (Figure 3). Furthermore, a 100% germ line transmission of the transgene to F1 progeny as a result of gynogenesis was observed [49]. The optimization of these different transgenesis techniques and the successful generation of transgenic LG clones now makes it possible to perform immunological assays such as transplantation studies and adoptive transfer on a transgenic background.
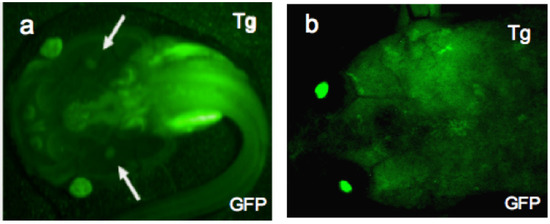
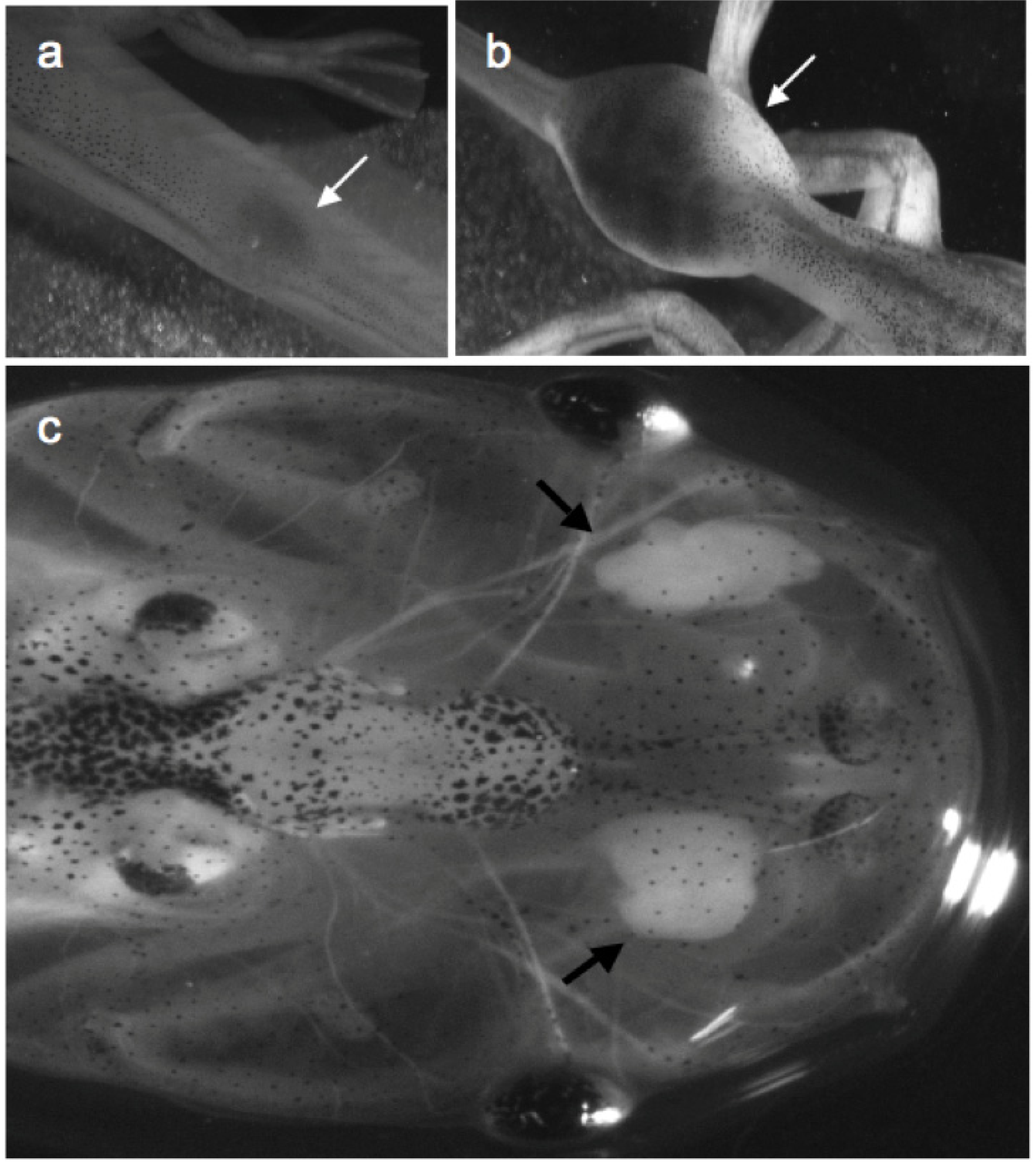
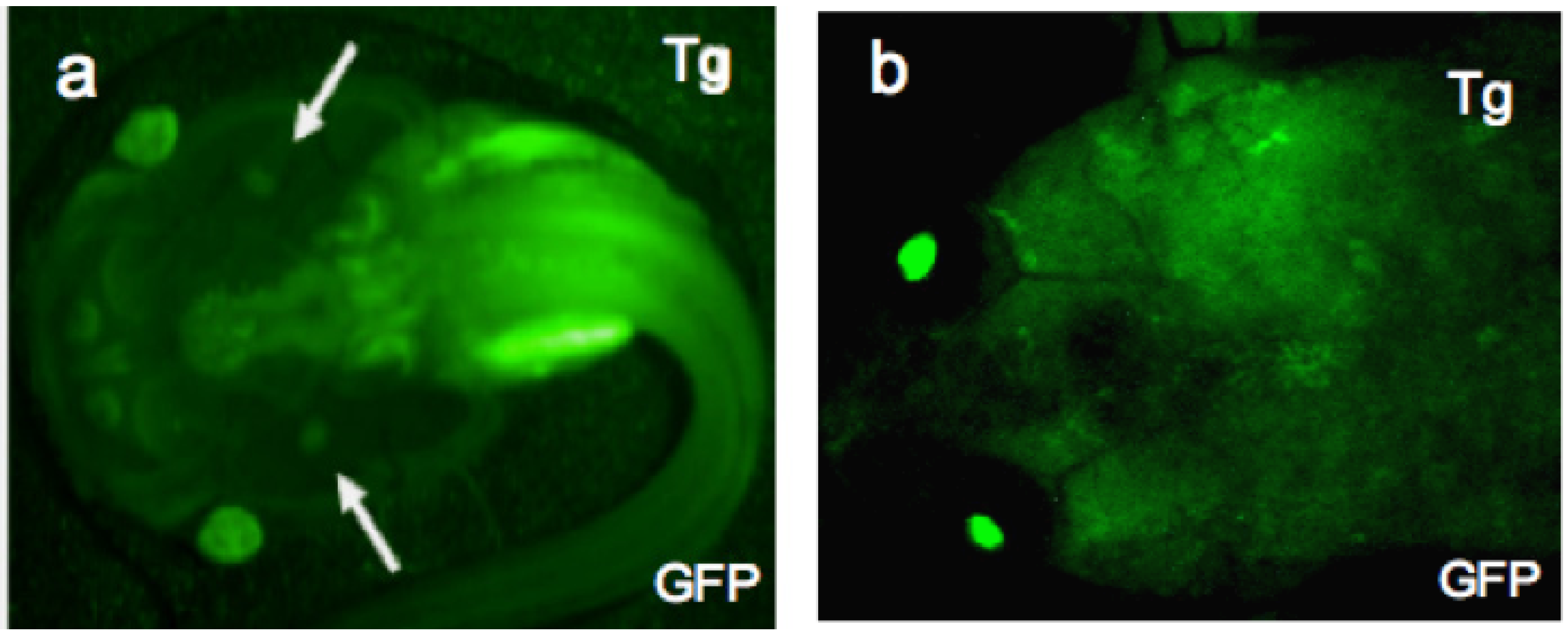
Figure 3.
LG-15 frogs expressing the transgene GFP. Strong and uniform expression of the transgene, GFP is achieved in F0 LG-15 animals using I-SecI meganuclease mediated transgenesis (a) One-month-old larva (developmental stage 56) and (b) one-year-old adult were screened for stable integration of the transgene, GFP using a fluorescence stereomicroscope. Arrows in (a) indicate thymi.
Figure 3.
LG-15 frogs expressing the transgene GFP. Strong and uniform expression of the transgene, GFP is achieved in F0 LG-15 animals using I-SecI meganuclease mediated transgenesis (a) One-month-old larva (developmental stage 56) and (b) one-year-old adult were screened for stable integration of the transgene, GFP using a fluorescence stereomicroscope. Arrows in (a) indicate thymi.
3.1.1. Recent Advances in Loss-of-Function Approaches
As Xenopus is ideally suited for high-throughput analysis of gene function, the need to further develop reliable loss-of-function technologies has been recognized, and recently various methodologies of gene silencing have been successfully applied to Xenopus. These include RNA interference (RNAi) [50], transcriptional activator-like effector nucleases (TALENs) [51,52], and zinc finger proteases [53]. Vertebrates RNAi technologies are based on small hairpin RNAs (shRNAs) with homology to the target gene. A construct encoding the shRNA under a RNA polymerase III promoter (e.g., H1 or U6) is integrated into the host genome, and upon transcription, the product is loaded into the RNA-induced silencing complex (RISC). Subsequently the sense-strand RNA is degraded whereas the anti-sense RNA strand directs RISC to its complementary mRNA target sequence resulting in targeted gene silencing [50]. As proof of concept, efficient shRNA-mediated silencing has recently been achieved in X. laevis LG clones by combining RNAi with the I-SceI meganuclease-mediated transgenesis, providing a convenient and efficient alternative to morpholinos for loss-of-function studies in both tadpoles and adults [54]. Using this approach, β2-microglobulin (b2m), a critical component for proper surface expression of both class Ia and the majority of class Ib molecules, was efficiently knocked down. A high level of knockdown was observed in both LG and outbred F1 animals. However, LG clones exhibited a higher degree of b2m knockdown (~60% compared to ~40%; [54]). Moreover, transgenic LG animals had lower knockdown variability among individuals compared to outbred frogs. This is most likely due to the out-crossing of outbreed male and female transgenic founders with different levels of knockdown. The higher penetrance of RNAi observed in the F1 generation of LG animals could offer an advantage for functional studies and the generation of these animals by gynogenesis bypasses the need for out-crossing and subsequent screening of F1 progeny. Importantly, even partial silencing (~60% KD) of b2m had a measurable functional effect. It resulted in reduced MHC class Ia protein surface expression, fewer circulating CD8+ T cells and significantly delayed skin graft rejection. These results clearly demonstrate the feasibility of this reverse genetic approach to study the effect of gene modulation in vivo. Additionally, functional studies using this RNAi-based transgenesis method are applicable to the F0 generation. This is shown by the fact that b2m silencing increased susceptibility of transgenic F0 larvae to viral infection [54]. The possibility of conducting functional studies in the F0 generation is important as it significantly accelerates the timeline of investigations.
Besides using shRNA to silence genes, two additional reverse genetics approaches have recently been successfully applied to Xenopus: transcriptional activator-like effector nucleases (TALENs) and zinc finger nucleases (ZFN) [51,52,53]. TALENs consist of a TAL effector DNA binding domains that can be engineered to specifically target any sequence of interest in the host genome. The DNA binding domain is fused to the catalytic domain of the endonuclease FokI. The binding of two adjacent TALENs allows dimerization of the FokI endonuclease domains, which then create double stranded breaks in the target sequence. These breaks are frequently inaccurately repaired creating insertion or deletion mutations disrupting the gene expression. Similarly, ZFN consist of a DNA binding domain (zinc fingers), which can be engineered to bind any sequence, fused to the nonspecific cleavage domain of FokI. Collectively, these different reverse genetic approaches are emerging as potential alternatives to antisense morpholino oligonucleotides for performing loss-of-function experiments in Xenopus.
4. The Xenopus Genus: Genetic Resources and Polyploidy
By far, the two best-known and most studied species of Xenopus are the pseudotetraploid X. laevis and its sister species X. tropicalis, which is the only true diploid species in the Xenopus genus. Each of these species offers unique advantages and complements each other providing a strong platform upon which to further build a powerful non-mammalian biomedical model. However, the existence of other polyoploid Xenopus species strengthens this system by providing a unique and comprehensive model to study many aspects of genomic regulation of the immune system (see Section 4.1.1 below).
4.1. Genomic Resources
Xenopus has a key phylogenetic position connecting mammals with vertebrates of more ancient origin, such as bony and cartilaginous fish, and the availability of X. tropicalis and X. laevis genomes will drastically advance the understanding of vertebrate genome evolution. Based primarily on the diploid and relatively small genome size (~1.7 Gbp), X. tropicalis was the first amphibian selected for whole genome sequencing and a draft version of the genome was published in 2010 [4]. A high level of synteny with human genomes was reported and more then half of all gene models could be associated with a human, mouse, and/or chicken orthologs. Moreover, a large number of X. tropicalis orthologs of identified human disease genes were found [4]. Currently the whole genome sequencing and annotation of X. laevis using a male of the MHC-homozygous inbreed X. laevis J strain of our colony is ongoing and both genomes can be accessed via the X. laevis and X. tropicalis biology and genomics resources website [22]. Despite some initial trepidation as to whether or not the allotetraploid nature of the X. laevis genome would allow for separate assembly of the alloalleles in the genome, recent technological advances made this possible and the results of the X. laevis genome sequencing project has exceeded expectations. As mentioned, the use of the partially inbred F strain to generate F2 progeny from heterozygous J/F parents will help the assignment of paralogs to scaffolds and subsequently chromosomes.
Furthermore, by taking advantage of the large scale genomic sequence data to compare X. laevis and X. tropicalis gene homologs it is possible to analyze a gene complement relatively soon after a rediplodization event. It is estimated that X. laevis and X. tropicalis coding DNA regions are ~90% identical which is comparable to human and mice, which are 85% identical, consistent with the fact that the two species diverged from a common ancestor ~65 million years ago, which is approximately the same time frame as that separating primates and rodents. Additionally >1 million X. laevis and >1.5 million X. tropicalis express sequence tags (EST) have been sequenced and different EST libraries where the EST are grouped into putative gene entries are available from the NCBI UniGene website [55]. In short, the availability of Xenopus genetic information has opened a new dimension of information greatly facilitating the identification of novel and potentially immune relevant genes.
4.1.1. Natural and Artificial Polyploidy in the Xenopus Genus
The Xenopus genus is one of few extant vertebrate groups that contain natural polyploid species and the only one with a distinct series of well-defined relatively recently derived species (1–40 Ma, [56]), ranging from diploid (2n) to dodecaploid (12n) species. This range of species provides additional platforms for further studies of many important aspects of immunity. Polyploidization in Xenopus is inferred to have occurred multiple times during the course of evolution spanning a period of 50–80 million years, with speciation occurring through a combination of bifurcating- and reticulate-speciation via allopolyploidization [57,58]. Compared to bifurcating speciation where one ancestor gives rise to two descendants with the same number of chromosomes, reticulating speciation via allopolyploidization occurs by inter-species hybridization and fusion of the two genomes resulting in a new descendant with, at least in the early stages of evolution, a duplicated genome. The Xenopus genus is comprised of more than 20 species of frogs including a single true diploid species, Xenopus (Silurana) tropicalis, and a series of tetraploid species, including X. laevis, (4n; X. gilli, X. clivii, X. largeni, X. fraseri, X. pygmaeus, X. borealis, X. mulleri, and S. epitropicalis), octoploid (8n; X. wittei, X. vestitus, X. andrei, X. bombaensi, and X. amieti) and at least two dodecaploid species (12n; X. longipes and X. ruwenzoriensis) (reviewed in [57]). These different Xenopus species, with varying levels of polyploidy, provide a unique opportunity to study the effects of whole genome duplication on genome evolution and the subsequent fate of individual genes such as redundancy, neofunctionalization, subfunctionalization, and pseudogenization. Importantly, polyploidy animals can be experimentally generated in the laboratory by subjecting diploid eggs from isogenetic clones to hydrostatic pressure or cold shock during the last meiotic division [23].
Xenopus also provides an excellent model for the study of rediploidization following whole genome-wide duplication as the X. laevis allotetraploidization event is considerably more recent (~40 Ma) compared to the vertebrate-specific (>500 Ma [58]) and teleost-specific duplication events (~350 Ma), while being substantially longer then the average lifespan of a duplicated genes in a diploid background. By comparing X. tropicalis and X. laevis genes, it has been estimated that around 50%–70% of the genes in X. laevis have been diploidized and are now expressed as single copy genes. Although there does not appear to be an enrichment either to maintain duplicate gene copies or towards diploidization for any specific functional category of genes [59,60,61], it is interesting to note that genes involved in antigen-specific adaptive immunity such as MHC and Ig heavy chain have undergone rapid diploidization [62,63]. Specifically, studies of duplicated immune genes after whole genome duplication using Xenopus polyploidy species will provide insight into the emergence of the mammalian adaptive immune system.
5. Concluding Remarks
Xenopus has many advantages that make it an ideal and diverse non-mammalian comparative model system suitable for many aspects of biomedical sciences. These advantages include the developmental transition from larval to adults, multiple naturally polyploidy species, a powerful skin graft/tumor transplantation system and a well established, reverse genetics approach to in vivo study of loss and gain of function. In addition, with the advances in genome sequencing a large amount of genetic information is available. Taken together, Xenopus provides a unique model for investigating both fundamental concepts of ontogeny and evolution of the immune system, as well as more applied aspects of biomedical research.
Acknowledgments
We would like to thank Nicolas Cohen and Leon Grayfer, and Nikesha Haynes for critical reading of the manuscript.
Conflict of Interest
The authors declare no conflict of interest.
References
- Robert, J.; Ohta, Y. Comparative and developmental study of the immune system in Xenopus. Dev. Dyn. 2009, 238, 1249–1270. [Google Scholar] [CrossRef]
- Du Pasquier, L.; Schwager, J.; Flajnik, M.F. The immune system of Xenopus. Annu. Rev. Immunol. 1989, 7, 251–275. [Google Scholar] [CrossRef]
- Khokha, M.K. Xenopus white papers and resources: Folding functional genomics and genetics into the frog. Genesis 2012, 50, 133–142. [Google Scholar] [CrossRef]
- Hellsten, U.; Harland, R.M.; Gilchrist, M.J.; Hendrix, D.; Jurka, J.; Kapitonov, V.; Ovcharenko, I.; Putnam, N.H.; Shu, S.; Taher, L.; et al. The genome of the western clawed frog Xenopus tropicalis. Science 2010, 328, 633–636. [Google Scholar] [CrossRef]
- Xenopus laevis Genome Resources Web Page. Available online: http://www.xenbase.org/genomes/static/laevis.jsp (accessed on 9 July 2013).
- University of Rochester Medical Center Web Page. Xenopus laevis Research Resource for Immunobiology. Available online: http://www.urmc.rochester.edu/mbi/resources/Xenopus (accessed on 9 July 2013).
- Hsu, E. Mutation, selection, and memory in B lymphocytes of exothermic vertebrates. Immunol. Rev. 1998, 162, 25–36. [Google Scholar] [CrossRef]
- Du Pasquier, L.; Robert, J.; Courtet, M.; Mussmann, R. B-cell development in the amphibian Xenopus. Immunol. Rev. 2000, 175, 201–213. [Google Scholar] [CrossRef]
- Barlow, E.H.; Cohen, N. The thymus dependency of transplantation allotolerance in the metamorphosing frog Xenopus laevis. Transplantation 1983, 35, 612–619. [Google Scholar] [CrossRef]
- Nedelkovska, H.; Robert, J. Comparative Study of Skin Graft Tolerance and Rejection in the Frog Xenopus laevis. Skin Graft–Indications, Applications and Current Research; Spear, S., Ed.; InTech: New York, NY, USA, 2011. Available online: http://www.intechopen.com/books/skin-grafts-indications-applications-and-current-research/comparative-study-of-skin-graft-tolerance-and-rejection-in-the-frog-Xenopus-laevis (accessed on 10 July 2013).
- Chardonnens, X.; Du Pasquier, L. Induction of skin allograft tolerance during metamorphosis of the toad Xenopus laevis: A possible model for studying generation of self tolerance to histocompatibility antigens. Eur. J. Immunol. 1973, 3, 569–573. [Google Scholar] [CrossRef]
- Horton, J.D.; Horton, T.L.; Ritchie, P. Incomplete tolerance induced in Xenopus by larval tissue allografting: Evidence from immunohistology and mixed leucocyte culture. Dev. Comp. Immunol. 1993, 17, 249–262. [Google Scholar] [CrossRef]
- Turpen, J.B.; Smith, P.B. Precursor immigration and thymocyte succession during larval development and metamorphosis in Xenopus. J. Immunol. 1989, 142, 41–47. [Google Scholar]
- Flajnik, M.F.; Kaufman, J.F.; Hsu, E.; Manes, M.; Parisot, R.; Du Pasquier, L. Major histocompatibility complex-encoded class I molecules are absent in immunologically competent Xenopus before metamorphosis. J. Immunol. 1986, 137, 3891–3899. [Google Scholar]
- Salter-Cid, L.; Nonaka, M.; Flajnik, M.F. Expression of MHC class Ia and class Ib during ontogeny: High expression in epithelia and coregulation of class Ia and lmp7 genes. J. Immunol. 1998, 160, 2853–2861. [Google Scholar]
- Rollins-Smith, L.A.; Flajnik, M.F.; Blair, P.J.; Davis, A.T.; Green, W.F. Involvement of thyroid hormones in the expression of MHC class I antigens during ontogeny in Xenopus. Dev. Immunol. 1997, 5, 133–144. [Google Scholar] [CrossRef]
- Du Pasquier, L.; Flajnik, M.F. Expression of MHC class II antigens during Xenopus development. Dev. Immunol. 1990, 1, 85–95. [Google Scholar] [CrossRef]
- Horton, J.D.; Horton, T.L.; Dzialo, R.; Gravenor, I.; Minter, R.; Ritchie, P.; Gartland, L.; Watson, M.D.; Cooper, M.D. T-cell and natural killer cell development in thymectomized Xenopus. Immunol. Rev. 1998, 166, 245–258. [Google Scholar] [CrossRef]
- Robert, J.; Gantress, J.; Cohen, N.; Maniero, G.D. Xenopus as an experimental model for studying evolution of hsp—Immune system interactions. Methods 2004, 32, 42–53. [Google Scholar] [CrossRef]
- Du Pasquier, L.; Chardonnens, C. Genetic aspects of the tolerance to allografts induced at metamorphosis in the toad Xenopus laevis. Immunogenetics 1975, 2, 431–440. [Google Scholar] [CrossRef]
- Tochinai, S.K.; Katagiri, C. Complete abrogation of immune responses to skin allografts and rabbit erythrocytes in the early thymectomized Xenopus. Dev. Growth Differ. 1975, 17, 383–394. [Google Scholar]
- Xenbase Home Page. Available online: http://www.xenbase.org (accessed on 9 July 2013).
- Kobel, H.R.; Du Pasquier, L. Production of large clones of histocompatible, fully identical clawed toads (Xenopus). Immunogenetics 1975, 2, 7–91. [Google Scholar]
- Nedelkovska, H.; Cruz-Luna, T.; McPherson, P.; Robert, J. Comparative in vivo study of gp96 adjuvanticity in the frog Xenopus laevis. J. Vis. Exp. 2010, 43, 2026:1–2026:5. [Google Scholar]
- Robert, J.; Cohen, N. Evolution of immune surveillance and tumor immunity: Studies in Xenopus. Immunol. Rev. 1998, 166, 231–243. [Google Scholar] [CrossRef]
- Goyos, A.; Robert, J. Tumorigenesis and anti-tumor immune responses in Xenopus. Front. Biosci. 2009, 14, 167–176. [Google Scholar] [CrossRef]
- Du Pasquier, L.; Robert, J. In vitro growth of thymic tumor cell lines from Xenopus. Dev. Immunol. 1992, 2, 295–307. [Google Scholar] [CrossRef]
- Robert, J.; Guiet, C.; Du Pasquier, L. Lymphoid tumors of Xenopus laevis with different capacities for growth in larvae and adults. Dev. Immunol. 1994, 3, 297–307. [Google Scholar] [CrossRef]
- Robert, J.; Cohen, N. Ontogeny of CTX expression in Xenopus. Dev. Comp. Immunol. 1998, 22, 605–612. [Google Scholar] [CrossRef]
- Robert, J.; Guiet, C.; Du Pasquier, L. Ontogeny of the alloimmune response against a transplanted tumor in Xenopus laevis. Differentiation 1995, 59, 135–144. [Google Scholar] [CrossRef]
- Goyos, A.; Guselnikov, S.; Chida, A.S.; Sniderhan, L.F.; Maggirwar, S.B.; Nedelkovska, H.; Robert, J. Involvement of nonclassical MHC class Ib molecules in heat shock protein-mediated anti-tumor responses. Eur. J. Immunol. 2007, 37, 1494–1501. [Google Scholar] [CrossRef]
- Goyos, A.; Ohta, Y.; Guselnikov, S.; Robert, J. Novel nonclassical MHC class Ib genes associated with CD8 T cell development and thymic tumors. Mol. Immunol. 2009, 46, 1775–1786. [Google Scholar] [CrossRef]
- Zitvogel, L.; Tesniere, A.; Kroemer, G. Cancer despite immunosurveillance: Immunoselection and immunosubversion. Nat. Rev. Immunol. 2006, 6, 715–727. [Google Scholar] [CrossRef]
- Yie, S.M.; Yang, H.; Ye, S.R.; Li, K.; Dong, D.D.; Lin, X.M. Expression of human leucocyte antigen G (HLA-G) is associated with prognosis in non-small cell lung cancer. Lung Cancer 2007, 58, 267–274. [Google Scholar]
- De Kruijf, E.M.; Sajet, A.; van Nes, J.G.; Natanov, R.; Putter, H.; Smit, V.T.; Liefers, G.J.; van den Elsen, P.J.; van de Velde, C.J.; Kuppen, P.J. HLA-E and HLA-G expression in classical HLA class I-negative tumors is of prognostic value for clinical outcome of early breast cancer patients. J. Immunol. 2010, 185, 7452–7459. [Google Scholar] [CrossRef]
- Chesneau, A.; Sachs, L.M.; Chai, N.; Chen, Y.; Du Pasquier, L.; Loeber, J.; Pollet, N.; Reilly, M.; Weeks, D.L.; Bronchain, O.J. Transgenesis procedures in Xenopus. Biol. Cell 2008, 100, 503–521. [Google Scholar] [CrossRef]
- Kroll, K.L.; Amaya, E. Transgenic Xenopus embryos from sperm nuclear transplantations reveal FGF signaling requirements during gastrulation. Development 1996, 122, 3173–3183. [Google Scholar]
- Amaya, E.; Kroll, K.L. A method for generating transgenic frog embryos. MethodsMol. Biol. 1999, 97, 393–414. [Google Scholar]
- Allen, B.G.; Weeks, D.L. Transgenic Xenopus laevis embryos can be generated using phiC31 integrase. Nat. Methods 2005, 2, 975–979. [Google Scholar] [CrossRef]
- Sinzelle, L.; Vallin, J.; Coen, L.; Chesneau, A.; Du Pasquier, D.; Pollet, N.; Demeneix, B.; Mazabraud, A. Generation of trangenic Xenopus laevis using the Sleeping Beauty transposon system. Transgenic Res. 2006, 15, 751–760. [Google Scholar] [CrossRef]
- Ogino, H.; McConnell, W.B.; Grainger, R.M. High-throughput transgenesis in Xenopus using I-SceI meganuclease. Nat. Protoc. 2006, 1, 1703–1710. [Google Scholar] [CrossRef]
- Ogino, H.; McConnell, W.B.; Grainger, R.M. Highly efficient transgenesis in Xenopus tropicalis using I-SceI meganuclease. Mech. Dev. 2006, 123, 103–113. [Google Scholar] [CrossRef]
- Pan, F.C.; Chen, Y.; Loeber, J.; Henningfeld, K.; Pieler, T. I-SceI meganuclease-mediated transgenesis in Xenopus. Dev. Dyn. 2006, 235, 247–252. [Google Scholar] [CrossRef]
- Allen, B.G.; Weeks, D.L. Bacteriophage phiC31 integrase mediated transgenesis in Xenopus laevis for protein expression at endogenous levels. Methods Mol. Biol. 2009, 518, 113–122. [Google Scholar] [CrossRef]
- Groth, A.C.; Olivares, E.C.; Thyagarajan, B.; Calos, M.P. A phage integrase directs efficient site-specific integration in human cells. Proc. Natl. Acad. Sci. USA 2000, 97, 5995–6000. [Google Scholar] [CrossRef]
- Yergeau, D.A.; Kelley, C.M.; Zhu, H.; Kuliyev, E.; Mead, P.E. Transposon transgenesis in Xenopus. Methods 2010, 51, 92–100. [Google Scholar] [CrossRef]
- Hamlet, M.R.; Yergeau, D.A.; Kuliyev, E.; Takeda, M.; Taira, M.; Kawakami, K.; Mead, P.E. Tol2 transposon-mediated transgenesis in Xenopus tropicalis. Genesis 2006, 44, 438–445. [Google Scholar] [CrossRef]
- Miskey, C.; Izsvak, Z.; Plasterk, R.H.; Ivics, Z. The Frog Prince: A reconstructed transposon from Rana pipiens with high transpositional activity in vertebrate cells. Nucleic Acids Res. 2003, 31, 6873–6881. [Google Scholar] [CrossRef]
- Nedelkovska, H.; Robert, J. Optimized transgenesis in Xenopus laevis/gilli isogenetic clones for immunological studies. Genesis 2012, 50, 300–306. [Google Scholar] [CrossRef]
- Sifuentes-Romero, I.; Milton, S.L.; Garcia-Gasca, A. Post-transcriptional gene silencing by RNA interference in non-mammalian vertebrate systems: Where do we stand? Mutat. Res. 2011, 728, 158–171. [Google Scholar] [CrossRef]
- Lei, Y.; Guo, X.; Liu, Y.; Cao, Y.; Deng, Y.; Chen, X.; Cheng, C.H.; Dawid, I.B.; Chen, Y.; Zhao, H. Efficient targeted gene disruption in Xenopus embryos using engineered transcription activator-like effector nucleases (TALENs). Proc. Natl. Acad. Sci. USA 2012, 109, 17484–17489. [Google Scholar]
- Ishibashi, S.; Cliffe, R.; Amaya, E. Highly efficient bi-allelic mutation rates using TALENs in Xenopus tropicalis. Biol. Open 2012, 1, 1273–1276. [Google Scholar] [CrossRef]
- Young, J.J.; Cherone, J.M.; Doyon, Y.; Ankoudinova, I.; Faraji, F.M.; Lee, A.H.; Ngo, C.; Guschin, D.Y.; Paschon, D.E.; Miller, J.C.; et al. Efficient targeted gene disruption in the soma and germ line of the frog Xenopus tropicalis using engineered zinc-finger nucleases. Proc. Natl. Acad. Sci 2011, 108, 7052–7057. [Google Scholar] [CrossRef]
- Nedelkovska, H.; Edholm, E.S.; Haynes, N.; Robert, J. Effective RNAi-mediated beta2-microglobulin loss of function by transgenesis in Xenopus laevis. Biol. Open 2013, 2, 335–342. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information (NCBI) Web Page. UniGene. Available online: http://www.ncbi.nlm.nih.gov/unigene (accessed on 9 July 2013).
- Kobel, H.R.; Tinsley, R.C. Xenopus Species and Ecology 2: The Extant Species. In The Biology of Xenopus; Clarendon: Oxford, UK, 1996; pp. 9–31. [Google Scholar]
- Evans, B.J. Genome evolution and speciation genetics of clawed frogs (Xenopus and Silurana). Front. Biosci. 2008, 13, 4687–4706. [Google Scholar] [CrossRef]
- Kobel, H.R.; Du Pasquier, L. Genetics of polyploidy Xenopus. Trends Genet. 1986, 2, 310–315. [Google Scholar] [CrossRef]
- Flajnik, M.F.; Kasahara, M. Comparative genomics of the MHC: Glimpses into the evolution of the adaptive immune system. Immunity 2001, 15, 351–362. [Google Scholar] [CrossRef]
- Hellsten, U.; Khokha, M.K.; Grammer, T.C.; Harland, R.M.; Richardson, P.; Rokhsar, D.S. Accelerated gene evolution and subfunctionalization in the pseudotetraploid frog Xenopus laevis. BMC Biol 2007, 5. [Google Scholar] [CrossRef]
- Hughes, M.K.; Hughes, A.L. Evolution of duplicate genes in a tetraploid animal, Xenopus laevis. Mol. Biol. Evol. 1993, 10, 1360–1369. [Google Scholar]
- Shum, B.P.; Avila, D.; Du Pasquier, L.; Kasahara, M.; Flajnik, M.F. Isolation of a classical MHC class I cDNA from an amphibian. Evidence for only one class I locus in the Xenopus MHC. J. Immunol. 1993, 151, 5376–5386. [Google Scholar]
- Courtet, M.; Flajnik, M.; Du Pasquier, L. Major histocompatibility complex and immunoglobulin loci visualized by in situ hybridization on Xenopus chromosomes. Dev. Comp. Immunol. 2001, 25, 149–157. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).