5-Fluorouracil Conversion Pathway Mutations in Gastric Cancer

 ,
,  ,
,  ,
,  , and
, and
Abstract
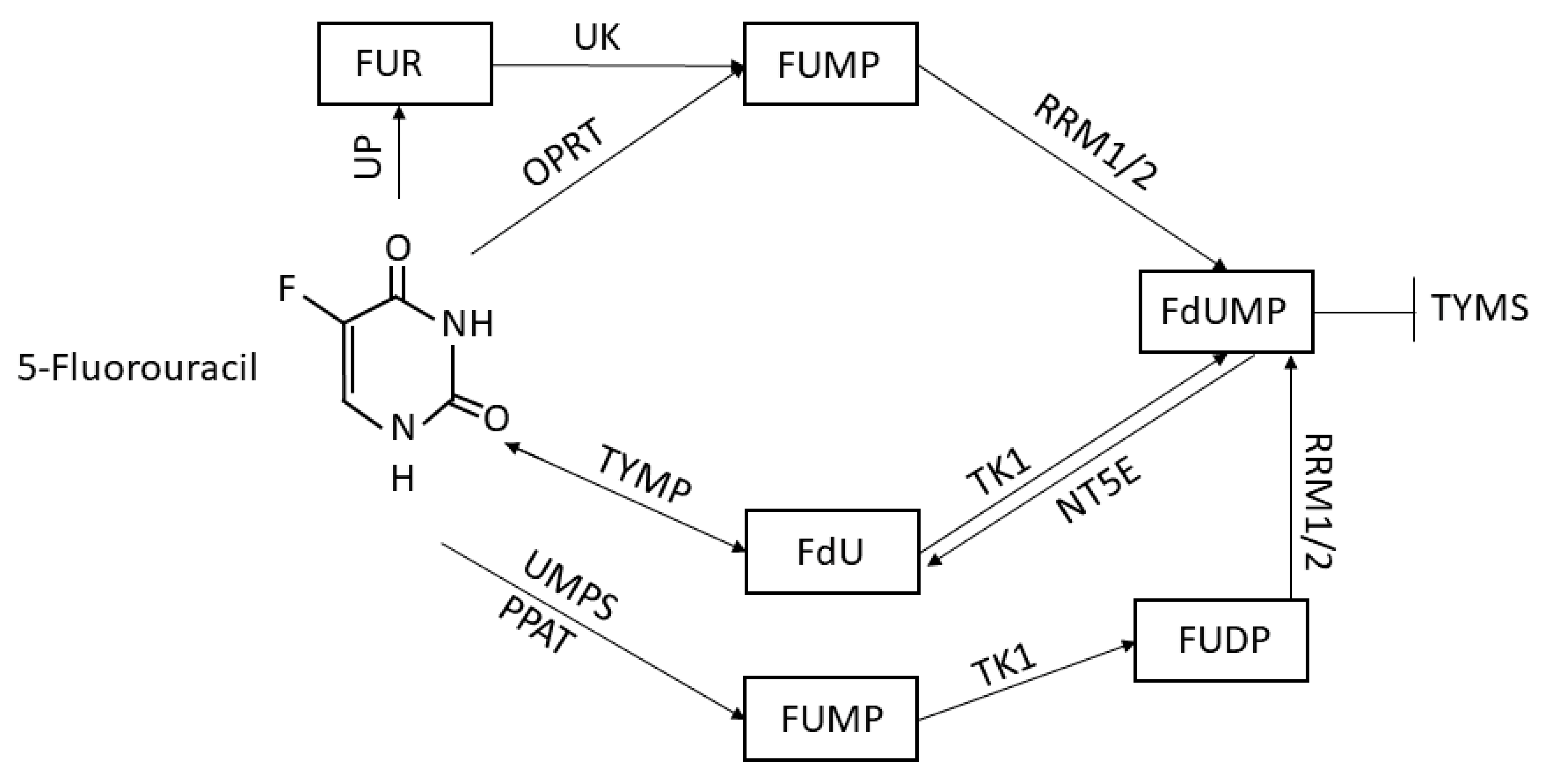
:1. Introduction
2. Materials and Methods
2.1. Analysis of Genomic Data
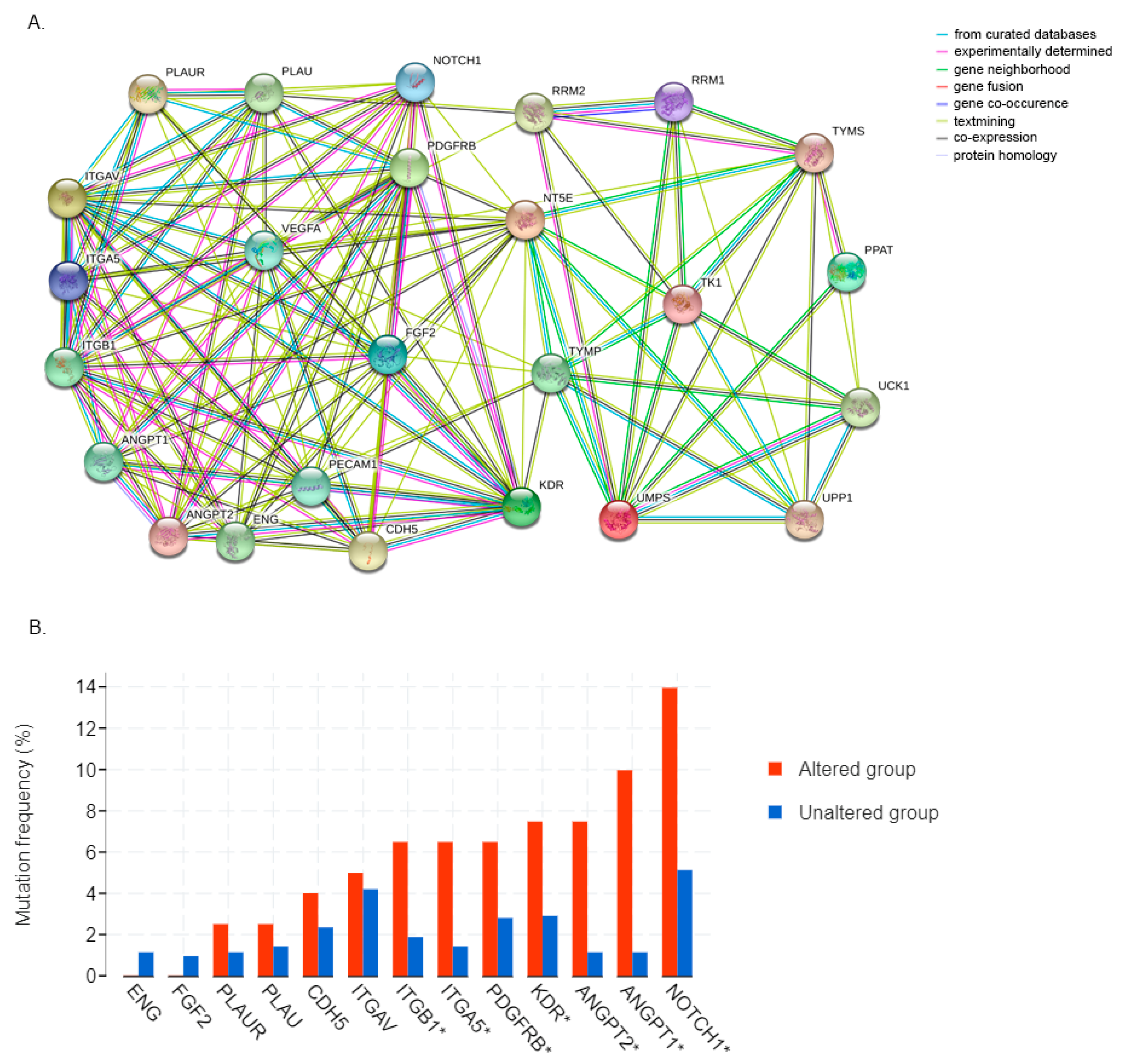
2.2. Protein–Protein Interaction Analysis
2.3. TYMP Expression Analysis
2.4. Statistical Analysis
3. Results
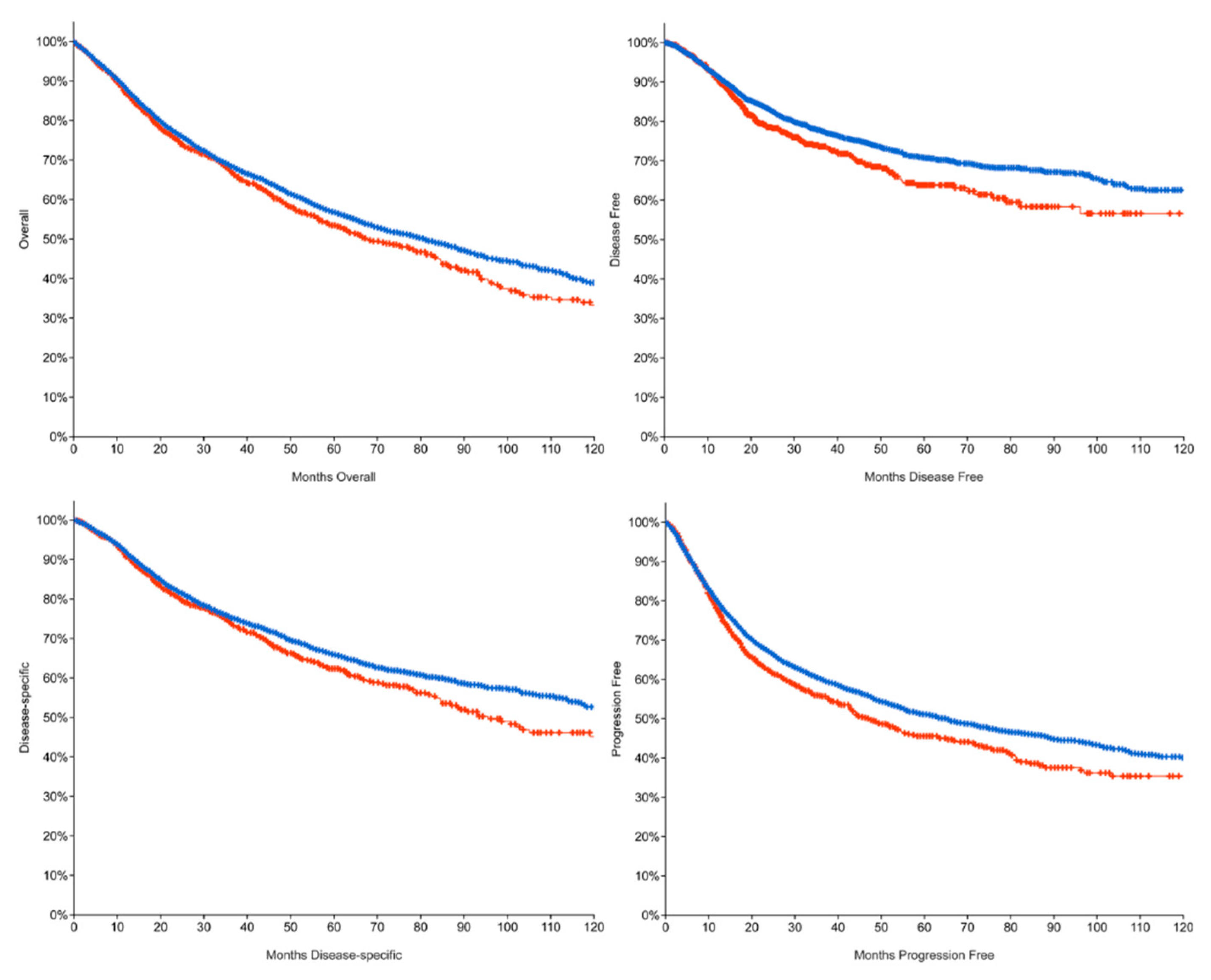
3.1. 5FU System Mutations Lead to a Poor Prognosis
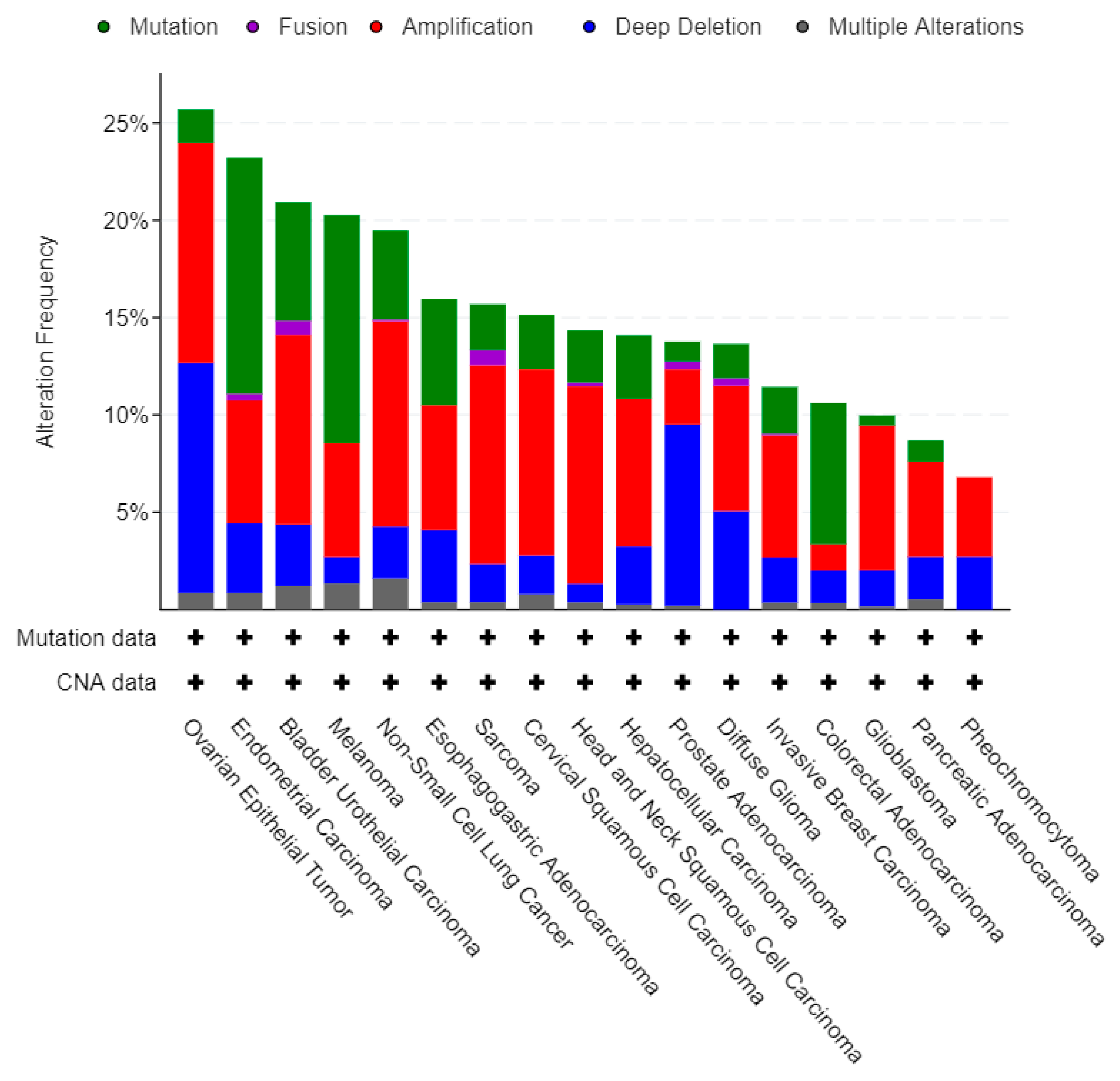
3.2. 5FU Conversion System Mutation Frequency among Cancers
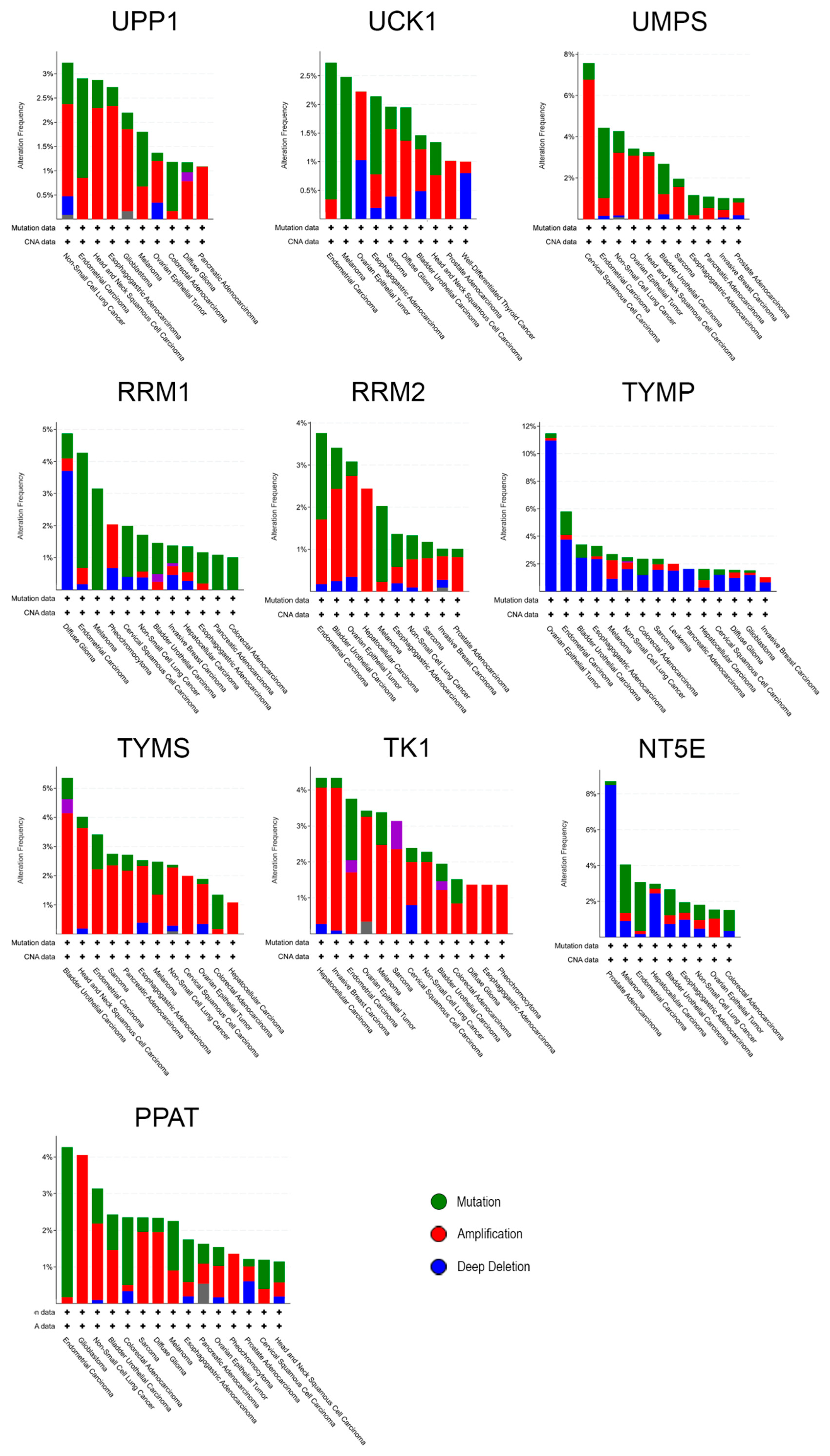
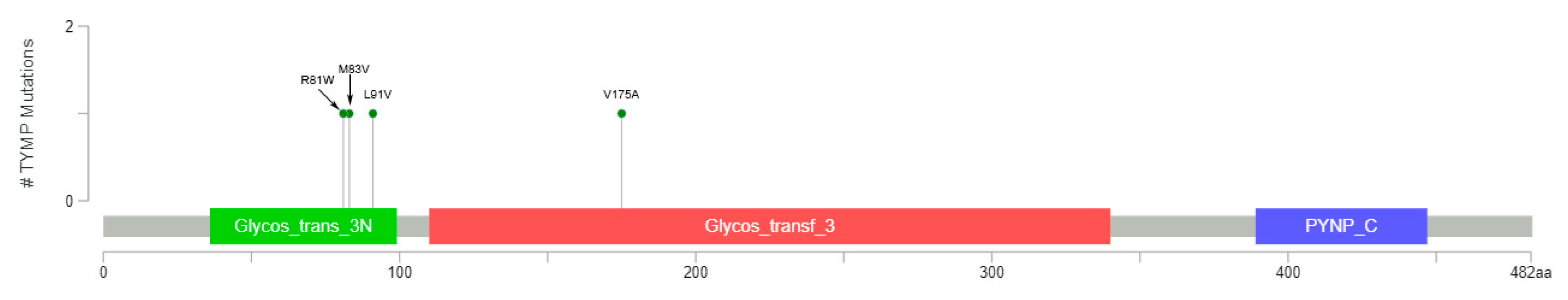
3.3. Individual Analysis of Mutation Frequency
3.4. 5FU Conversion System Mutations Analysis in GC
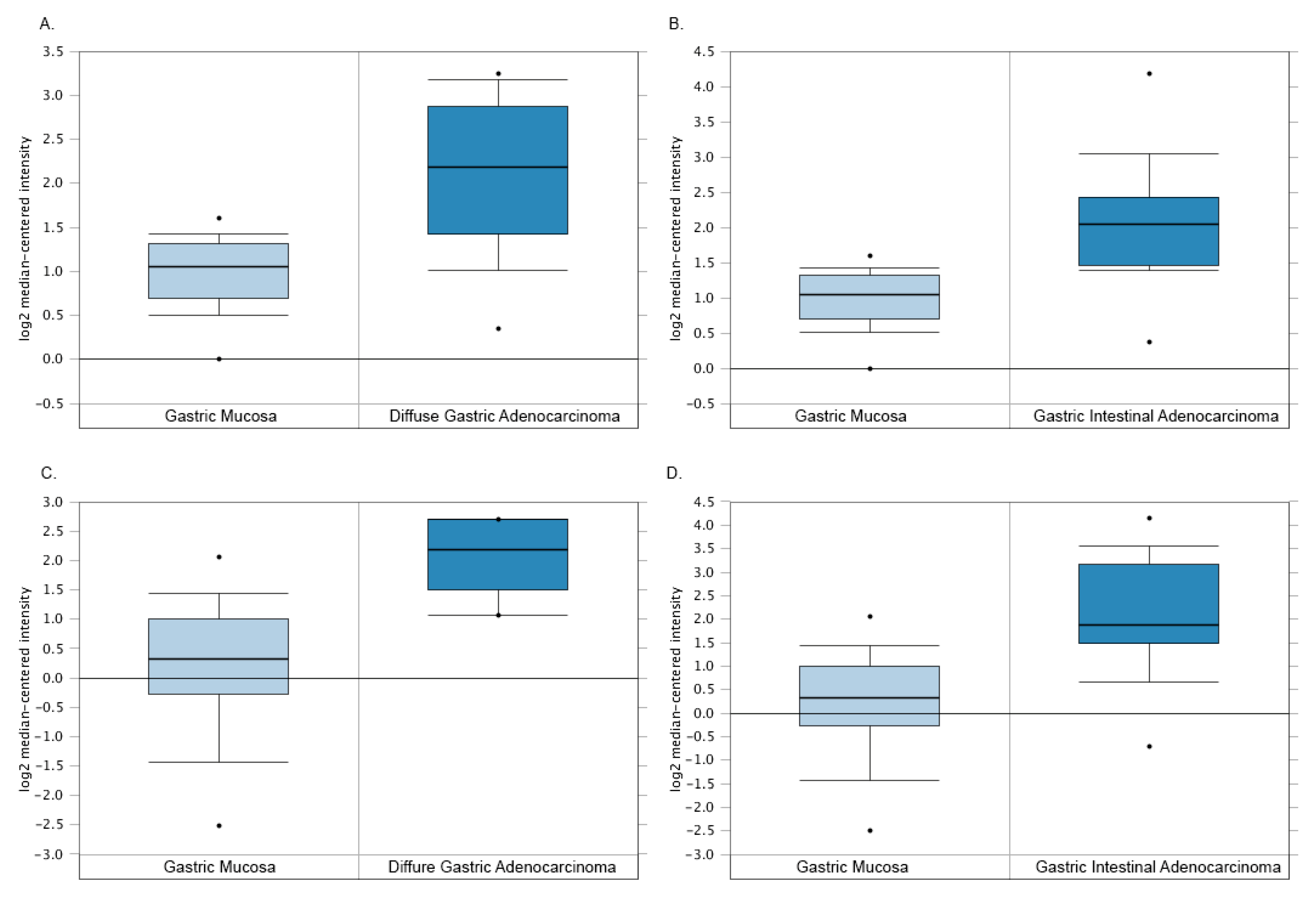
3.5. The Paradoxical TYMP Expression in GC
3.6. 5FU Conversion System Is Associated with Tumor Angiogenesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Biagioni, A.; Skalamera, I.; Peri, S.; Schiavone, N.; Cianchi, F.; Giommoni, E.; Magnelli, L.; Papucci, L. Update on gastric cancer treatments and gene therapies. Cancer Metastasis Rev. 2019, 38, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Heidelberger, C.; Chaudhuri, N.K.; Danneberg, P.; Mooren, D.; Griesbach, L.; Duschinsky, R.; Schnitzer, R.J.; Pleven, E.; Scheiner, J. Fluorinated pyrimidines, a new class of tumour-inhibitory compounds. Nature 1957, 179, 663–666. [Google Scholar] [CrossRef]
- Longley, D.B.; Harkin, D.P.; Johnston, P.G. 5-fluorouracil: Mechanisms of action and clinical strategies. Nat. Rev. Cancer 2003, 3, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Mori, R.; Yoshida, K.; Futamura, M.; Suetsugu, T.; Shizu, K.; Tanahashi, T.; Tanaka, Y.; Matsuhashi, N.; Yamaguchi, K. The inhibition of thymidine phosphorylase can reverse acquired 5FU-resistance in gastric cancer cells. Gastric Cancer 2019, 22, 497–505. [Google Scholar] [CrossRef] [Green Version]
- Ooyama, A.; Takechi, T.; Toda, E.; Nagase, H.; Okayama, Y.; Kitazato, K.; Sugimoto, Y.; Oka, T.; Fukushima, M. Gene expression analysis using human cancer xenografts to identify novel predictive marker genes for the efficacy of 5-fluorouracil-based drugs. Cancer Sci. 2006, 97, 510–522. [Google Scholar] [CrossRef]
- Griffith, M.; Mwenifumbo, J.C.; Cheung, P.Y.; Paul, J.E.; Pugh, T.J.; Tang, M.J.; Chittaranjan, S.; Morin, R.D.; Asano, J.K.; Ally, A.A.; et al. Novel mRNA isoforms and mutations of uridine monophosphate synthetase and 5-fluorouracil resistance in colorectal cancer. Pharmacogenomics J. 2013, 13, 148–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gustavsson, B.; Carlsson, G.; Machover, D.; Petrelli, N.; Roth, A.; Schmoll, H.-J.; Tveit, K.-M.; Gibson, F. A Review of the Evolution of Systemic Chemotherapy in the Management of Colorectal Cancer. Clin. Color. Cancer 2015, 14, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.-H.; Dong, H.; Zhao, F.; Tang, J.; Chen, X.; Ding, J.; Men, H.-T.; Luo, W.-X.; Du, Y.; Ge, J.; et al. The upregulation of dihydropyrimidine dehydrogenase in liver is involved in acquired resistance to 5-fluorouracil. Eur. J. Cancer 2013, 49, 1752–1760. [Google Scholar] [CrossRef]
- Sara, J.D.; Kaur, J.; Khodadadi, R.; Rehman, M.; Lobo, R.; Chakrabarti, S.; Herrmann, J.; Lerman, A.; Grothey, A. 5-fluorouracil and cardiotoxicity: A review. Ther. Adv. Med. Oncol. 2018, 10, 175883591878014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordier, P.-Y.; Nau, A.; Ciccolini, J.; Oliver, M.; Mercier, C.; Lacarelle, B.; Peytel, E. 5-FU-induced neurotoxicity in cancer patients with profound DPD deficiency syndrome: A report of two cases. Cancer Chemother. Pharmacol. 2011, 68, 823–826. [Google Scholar] [CrossRef] [PubMed]
- Chintala, L.; Vaka, S.; Baranda, J.; Williamson, S.K. Capecitabine versus 5-fluorouracil in colorectal cancer: Where are we now? Oncol. Rev. 2011, 5, 129–140. [Google Scholar] [CrossRef]
- Schöffski, P. The modulated oral fluoropyrimidine prodrug S-1, and its use in gastrointestinal cancer and other solid tumors. Anticancer Drugs 2004, 15, 85–106. [Google Scholar] [CrossRef]
- Kobayakawa, M.; Kojima, Y. Tegafur/gimeracil/oteracil (S-1) approved for the treatment of advanced gastric cancer in adults when given in combination with cisplatin: A review comparing it with other fluoropyrimidine-based therapies. Onco Targets Ther. 2011, 4, 193–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakata, K.-I.; Someya, M.; Matsumoto, Y.; Tauchi, H.; Kai, M.; Toyota, M.; Takagi, M.; Hareyama, M.; Fukushima, M. Gimeracil, an inhibitor of dihydropyrimidine dehydrogenase, inhibits the early step in homologous recombination. Cancer Sci. 2011, 102, 1712–1716. [Google Scholar] [CrossRef] [PubMed]
- Schilsky, R.L.; Kindler, H.L. Eniluracil: An irreversible inhibitor of dihydropyrimidine dehydrogenase. Expert Opin. Investig. Drugs 2000, 9, 1635–1649. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, E.; Tominaga, K.; Kuwamura, H.; Watanabe, T.; Fujiwara, Y.; Oshitani, N.; Higuchi, K.; Arakawa, T. Synergistic antitumor effect of combined 5-fluorouracil (5-FU) with 5-chloro-2,4-dihydroxypyridine on 5-FU-resistant gastric cancer cells: Possible role of a dihydropyrimidine dehydrogenase-independent mechanism. J. Gastroenterol. 2007, 42, 816–822. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, M.S.; Stojanov, P.; Polak, P.; Kryukov, G.V.; Cibulskis, K.; Sivachenko, A.; Carter, S.L.; Stewart, C.; Mermel, C.H.; Roberts, S.A.; et al. Mutational heterogeneity in cancer and the search for new cancer-associated genes. Nature 2013, 499, 214–218. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Ajani, J.A.; Buyse, M.; Lichinitser, M.; Gorbunova, V.; Bodoky, G.; Douillard, J.Y.; Cascinu, S.; Heinemann, V.; Zaucha, R.; Carrato, A.; et al. Combination of cisplatin/S-1 in the treatment of patients with advanced gastric or gastroesophageal adenocarcinoma: Results of noninferiority and safety analyses compared with cisplatin/5-fluorouracil in the First-Line Advanced Gastric Cancer Study. Eur. J. Cancer 2013, 49, 3616–3624. [Google Scholar] [CrossRef]
- Van Cutsem, E.; Moiseyenko, V.M.; Tjulandin, S.; Majlis, A.; Constenla, M.; Boni, C.; Rodrigues, A.; Fodor, M.; Chao, Y.; Voznyi, E.; et al. Phase III study of docetaxel and cisplatin plus fluorouracil compared with cisplatin and fluorouracil as first-line therapy for advanced gastric cancer: A report of the V325 Study Group. J. Clin. Oncol. 2006, 24, 4991–4997. [Google Scholar] [CrossRef]
- Al-Batran, S.-E.; Homann, N.; Pauligk, C.; Goetze, T.O.; Meiler, J.; Kasper, S.; Kopp, H.-G.; Mayer, F.; Haag, G.M.; Luley, K.; et al. Perioperative chemotherapy with fluorouracil plus leucovorin, oxaliplatin, and docetaxel versus fluorouracil or capecitabine plus cisplatin and epirubicin for locally advanced, resectable gastric or gastro-oesophageal junction adenocarcinoma (FLOT4): A randomised, phase 2/3 trial. Lancet 2019, 393, 1948–1957. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, C.S.; Tomasek, J.; Yong, C.J.; Dumitru, F.; Passalacqua, R.; Goswami, C.; Safran, H.; Dos Santos, L.V.; Aprile, G.; Ferry, D.R.; et al. Ramucirumab monotherapy for previously treated advanced gastric or gastro-oesophageal junction adenocarcinoma (REGARD): An international, randomised, multicentre, placebo-controlled, phase 3 trial. Lancet 2014, 383, 31–39. [Google Scholar] [CrossRef]
- Wilke, H.; Muro, K.; Van Cutsem, E.; Oh, S.-C.; Bodoky, G.; Shimada, Y.; Hironaka, S.; Sugimoto, N.; Lipatov, O.; Kim, T.-Y.; et al. Ramucirumab plus paclitaxel versus placebo plus paclitaxel in patients with previously treated advanced gastric or gastro-oesophageal junction adenocarcinoma (RAINBOW): A double-blind, randomised phase 3 trial. Lancet Oncol. 2014, 15, 1224–1235. [Google Scholar] [CrossRef]
- Maugeri-Saccà, M.; Pizzuti, L.; Sergi, D.; Barba, M.; Belli, F.; Fattoruso, S.; Giannarelli, D.; Amodio, A.; Boggia, S.; Vici, P.; et al. FOLFIRI as a second-line therapy in patients with docetaxel-pretreated gastric cancer: A historical cohort. J. Exp. Clin. Cancer Res. 2013, 32, 67. [Google Scholar] [CrossRef]
- Laurén, P. The Two Histological Main Types of Gastric Carcinoma: Diffuse and so-called Intestinal-type Carcinoma: An Attempt at a Histo-Clinical Classification. Acta Pathol. Microbiol. Scand. 1965, 64, 31–49. [Google Scholar] [CrossRef] [PubMed]
- Hadjipanayis, A.; Potapova, O.; Pantazi, A. The Cancer Genome Atlas Research Network Comprehensive molecular characterization of gastric adenocarcinoma. Nature 2014, 513, 202–209. [Google Scholar] [CrossRef] [Green Version]
- Lei, Z.; Tan, I.B.; Das, K.; Deng, N.; Zouridis, H.; Pattison, S.; Chua, C.; Feng, Z.; Guan, Y.K.; Ooi, C.H.; et al. Identification of molecular subtypes of gastric cancer with different responses to PI3-kinase inhibitors and 5-fluorouracil. Gastroenterology 2013, 145, 554–565. [Google Scholar] [CrossRef]
- Tan, P.; Yeoh, K.-G. Genetics and Molecular Pathogenesis of Gastric Adenocarcinoma. Gastroenterology 2015, 149, 1153–1162. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, l1. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.Y.; Lim, J.Y.; Cheong, J.H.; Park, Y.-Y.; Yoon, S.-L.; Kim, S.M.; Kim, S.-B.; Kim, H.; Hong, S.W.; Park, Y.N.; et al. Gene expression signature-based prognostic risk score in gastric cancer. Clin. Cancer Res. 2011, 17, 1850–1857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Errico, M.; de Rinaldis, E.; Blasi, M.F.; Viti, V.; Falchetti, M.; Calcagnile, A.; Sera, F.; Saieva, C.; Ottini, L.; Palli, D.; et al. Genome-wide expression profile of sporadic gastric cancers with microsatellite instability. Eur. J. Cancer 2009, 45, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, D.R.; Yu, J.; Shanker, K.; Deshpande, N.; Varambally, R.; Ghosh, D.; Barrette, T.; Pandey, A.; Chinnaiyan, A.M. ONCOMINE: A cancer microarray database and integrated data-mining platform. Neoplasia 2004, 6, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Murata, D.; Endo, Y.; Obata, T.; Sakamoto, K.; Syouji, Y.; Kadohira, M.; Matsuda, A.; Sasaki, T. A crucial role of uridine/cytidine kinase 2 in antitumor activity of 3′-ethynyl nucleosides. Drug Metab. Dispos. 2004, 32, 1178–1182. [Google Scholar] [CrossRef] [Green Version]
- Evrard, A.; Cuq, P.; Ciccolini, J.; Vian, L.; Cano, J.P. Increased cytotoxicity and bystander effect of 5-fluorouracil and 5-deoxy-5-fluorouridine in human colorectal cancer cells transfected with thymidine phosphorylase. Br. J. Cancer 1999, 80, 1726–1733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varghese, V.; Magnani, L.; Harada-Shoji, N.; Mauri, F.; Szydlo, R.M.; Yao, S.; Lam, E.W.-F.; Kenny, L.M. FOXM1 modulates 5-FU resistance in colorectal cancer through regulating TYMS expression. Sci. Rep. 2019, 9, 1505. [Google Scholar] [CrossRef] [Green Version]
- Jia, F.; Teer, J.K.; Knepper, T.C.; Lee, J.K.; Zhou, H.-H.; He, Y.-J.; McLeod, H.L. Discordance of Somatic Mutations Between Asian and Caucasian Patient Populations with Gastric Cancer. Mol. Diagn. Ther. 2017, 21, 179–185. [Google Scholar] [CrossRef]
- Kim, H.W.; Kim, J.-H.; Lim, B.J.; Kim, H.; Kim, H.; Park, J.J.; Youn, Y.H.; Park, H.; Noh, S.H.; Kim, J.W.; et al. Sex Disparity in Gastric Cancer: Female Sex is a Poor Prognostic Factor for Advanced Gastric Cancer. Ann. Surg. Oncol. 2016, 23, 4344–4351. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, C.; Wang, Q.; Li, Z.; Lin, J.; Wang, H. Differences in Stage of Cancer at Diagnosis, Treatment, and Survival by Race and Ethnicity Among Leading Cancer Types. JAMA Netw. Open 2020, 3, e202950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakiuchi, M.; Nishizawa, T.; Ueda, H.; Gotoh, K.; Tanaka, A.; Hayashi, A.; Yamamoto, S.; Tatsuno, K.; Katoh, H.; Watanabe, Y.; et al. Recurrent gain-of-function mutations of RHOA in diffuse-type gastric carcinoma. Nat. Genet. 2014, 46, 583–587. [Google Scholar] [CrossRef]
- Wang, K.; Kan, J.; Yuen, S.T.; Shi, S.T.; Chu, K.M.; Law, S.; Chan, T.L.; Kan, Z.; Chan, A.S.Y.; Tsui, W.Y.; et al. Exome sequencing identifies frequent mutation of ARID1A in molecular subtypes of gastric cancer. Nat. Genet. 2011, 43, 1219–1223. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Yuen, S.T.; Xu, J.; Lee, S.P.; Yan, H.H.N.; Shi, S.T.; Siu, H.C.; Deng, S.; Chu, K.M.; Law, S.; et al. Whole-genome sequencing and comprehensive molecular profiling identify new driver mutations in gastric cancer. Nat. Genet. 2014, 46, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Ellrott, K.; Bailey, M.H.; Saksena, G.; Covington, K.R.; Kandoth, C.; Stewart, C.; Hess, J.; Ma, S.; Chiotti, K.E.; McLellan, M.; et al. Scalable Open Science Approach for Mutation Calling of Tumor Exomes Using Multiple Genomic Pipelines. Cell Syst. 2018, 6, 271–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoadley, K.A.; Yau, C.; Hinoue, T.; Wolf, D.M.; Lazar, A.J.; Drill, E.; Shen, R.; Taylor, A.M.; Cherniack, A.D.; Thorsson, V.; et al. Cell-of-Origin Patterns Dominate the Molecular Classification of 10,000 Tumors from 33 Types of Cancer. Cell 2018, 173, 291–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, A.M.; Shih, J.; Ha, G.; Gao, G.F.; Zhang, X.; Berger, A.C.; Schumacher, S.E.; Wang, C.; Hu, H.; Liu, J.; et al. Genomic and Functional Approaches to Understanding Cancer Aneuploidy. Cancer Cell 2018, 33, 676–689. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Lichtenberg, T.; Hoadley, K.A.; Poisson, L.M.; Lazar, A.J.; Cherniack, A.D.; Kovatich, A.J.; Benz, C.C.; Levine, D.A.; Lee, A.V.; et al. An Integrated TCGA Pan-Cancer Clinical Data Resource to Drive High-Quality Survival Outcome Analytics. Cell 2018, 173, 400–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Vega, F.; Mina, M.; Armenia, J.; Chatila, W.K.; Luna, A.; La, K.C.; Dimitriadoy, S.; Liu, D.L.; Kantheti, H.S.; Saghafinia, S.; et al. Oncogenic Signaling Pathways in The Cancer Genome Atlas. Cell 2018, 173, 321–337. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.; Liang, W.-W.; Foltz, S.M.; Mutharasu, G.; Jayasinghe, R.G.; Cao, S.; Liao, W.-W.; Reynolds, S.M.; Wyczalkowski, M.A.; Yao, L.; et al. Driver Fusions and Their Implications in the Development and Treatment of Human Cancers. Cell Rep. 2018, 23, 227–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, H.; Konishi, K.; Kaji, M.; Maeda, K.; Yabushita, K.; Miwa, A. Correlation between expression levels of thymidine phosphorylase (dThdPase) and clinical features in human gastric carcinoma. Hepatogastroenterology 2002, 49, 882–886. [Google Scholar]
- Tabata, S.; Yamamoto, M.; Goto, H.; Hirayama, A.; Ohishi, M.; Kuramoto, T.; Mitsuhashi, A.; Ikeda, R.; Haraguchi, M.; Kawahara, K.; et al. Thymidine Catabolism as a Metabolic Strategy for Cancer Survival. Cell Rep. 2017, 19, 1313–1321. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Tang, S.; Wang, Y.; Xu, S.; Yu, J.; Zhi, X.; Ou, Z.; Yang, J.; Zhou, P.; Shao, Z. Ecto-5′-nucleotidase (CD73) promotes tumor angiogenesis. Clin. Exp. Metastasis 2013, 30, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, C.T.U.; Martins, N.N.F.; da Silva Oliveira, K.C.; Almeida, C.M.; Pinheiro, T.M.; Gigek, C.O.; de Araújo Cavallero, S.R.; Assumpção, P.P.; Smith, M.A.C.; Burbano, R.R.; et al. Liquid biopsy provides new insights into gastric cancer. Oncotarget 2018, 9, 15144–15156. [Google Scholar] [CrossRef] [Green Version]
- Györffy, B.; Surowiak, P.; Kiesslich, O.; Denkert, C.; Schäfer, R.; Dietel, M.; Lage, H. Gene expression profiling of 30 cancer cell lines predicts resistance towards 11 anticancer drugs at clinically achieved concentrations. Int. J. Cancer 2006, 118, 1699–1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawahara, A.; Hattori, S.; Akiba, J.; Nakashima, K.; Taira, T.; Watari, K.; Hosoi, F.; Uba, M.; Basaki, Y.; Koufuji, K.; et al. Infiltration of thymidine phosphorylase-positive macrophages is closely associated with tumor angiogenesis and survival in intestinal type gastric cancer. Oncol. Rep. 2010, 24, 405–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Zheng, Z.; Shin, Y.K.; Kim, K.-Y.; Rha, S.Y.; Noh, S.H.; Chung, H.C.; Jeung, H.-C. Angiogenic factor thymidine phosphorylase associates with angiogenesis and lymphangiogenesis in the intestinal-type gastric cancer. Pathology 2014, 46, 316–324. [Google Scholar] [CrossRef]
- Yu, E.J.; Lee, Y.; Rha, S.Y.; Kim, T.S.; Chung, H.C.; Oh, B.K.; Yang, W.I.; Noh, S.H.; Jeung, H.-C. Angiogenic factor thymidine phosphorylase increases cancer cell invasion activity in patients with gastric adenocarcinoma. Mol. Cancer Res. 2008, 6, 1554–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bronckaers, A.; Gago, F.; Balzarini, J.; Liekens, S. The dual role of thymidine phosphorylase in cancer development and chemotherapy. Med. Res. Rev. 2009, 29, 903–953. [Google Scholar] [CrossRef]
- Seftor, R.E.B.; Hess, A.R.; Seftor, E.A.; Kirschmann, D.A.; Hardy, K.M.; Margaryan, N.V.; Hendrix, M.J.C. Tumor Cell Vasculogenic Mimicry. Am. J. Pathol. 2012, 181, 1115–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garone, C.; Tadesse, S.; Hirano, M. Clinical and genetic spectrum of mitochondrial neurogastrointestinal encephalomyopathy. Brain 2011, 134, 3326–3332. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Median Months Overall (OS) | Median Months Disease-Free (DF) | Median Months Progression-Free (PFS) | Median Months Disease-Specific (DSS) | |
|---|---|---|---|---|
| Altered group | 67.46 | NA | 46.98 | 95.57 |
| Unaltered group | 80.74 | NA | 65.00 | 152.02 |
| Gene | Protein | Mutation Frequency | # Mutations | Missense | Truncating | Inframe | SMF |
|---|---|---|---|---|---|---|---|
| UPP1 | Uridine phosphorylase 1 | 2.6% (35) | 3 | 2 | 1 | 0 | 0.4% |
| UCK1 | Uridine-cytidine kinase 1 | 2.6% (35) | 8 | 6 | 2 | 0 | 1.2% |
| UMPS | Uridine monophosphate synthetase * | 1.4% (18) | 6 | 2 | 4 | 0 | 1.0% |
| RRM1 | Ribonucleotide reductase 1 | 1.4% (18) | 5 | 5 | 0 | 0 | 1.1% |
| RRM2 | Ribonucleotide reductase 2 | 1.4% (18) | 5 | 4 | 1 | 0 | 0.8% |
| TYMP | Thymidine phosphorylase | 2.8% (37) | 4 | 4 | 0 | 0 | 0.7% |
| TYMS | Thymidylate synthase | 1.4% (18) | 1 | 1 | 0 | 0 | 0.2% |
| TK1 | Thymidine kinase 1 | 1.4% (18) | 1 | 1 | 0 | 0 | 0.1% |
| NT5E | Ecto-5′-nucleotidase | 2.5% (33) | 6 | 4 | 1 | 1 | 0.7% |
| PPAT | Phosphoribosyl pyrophosphate amidotransferase | 2.2% (29) | 7 | 5 | 2 | 0 | 1.3% |
| N = 46 | 34 (73.9%) | 11 (23.9%) | 1 (2.2%) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biagioni, A.; Staderini, F.; Peri, S.; Versienti, G.; Schiavone, N.; Cianchi, F.; Papucci, L.; Magnelli, L. 5-Fluorouracil Conversion Pathway Mutations in Gastric Cancer. Biology 2020, 9, 265. https://doi.org/10.3390/biology9090265
Biagioni A, Staderini F, Peri S, Versienti G, Schiavone N, Cianchi F, Papucci L, Magnelli L. 5-Fluorouracil Conversion Pathway Mutations in Gastric Cancer. Biology. 2020; 9(9):265. https://doi.org/10.3390/biology9090265
Chicago/Turabian StyleBiagioni, Alessio, Fabio Staderini, Sara Peri, Giampaolo Versienti, Nicola Schiavone, Fabio Cianchi, Laura Papucci, and Lucia Magnelli. 2020. "5-Fluorouracil Conversion Pathway Mutations in Gastric Cancer" Biology 9, no. 9: 265. https://doi.org/10.3390/biology9090265
APA StyleBiagioni, A., Staderini, F., Peri, S., Versienti, G., Schiavone, N., Cianchi, F., Papucci, L., & Magnelli, L. (2020). 5-Fluorouracil Conversion Pathway Mutations in Gastric Cancer. Biology, 9(9), 265. https://doi.org/10.3390/biology9090265