Plant Defence Mechanisms Are Modulated by the Circadian System



{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
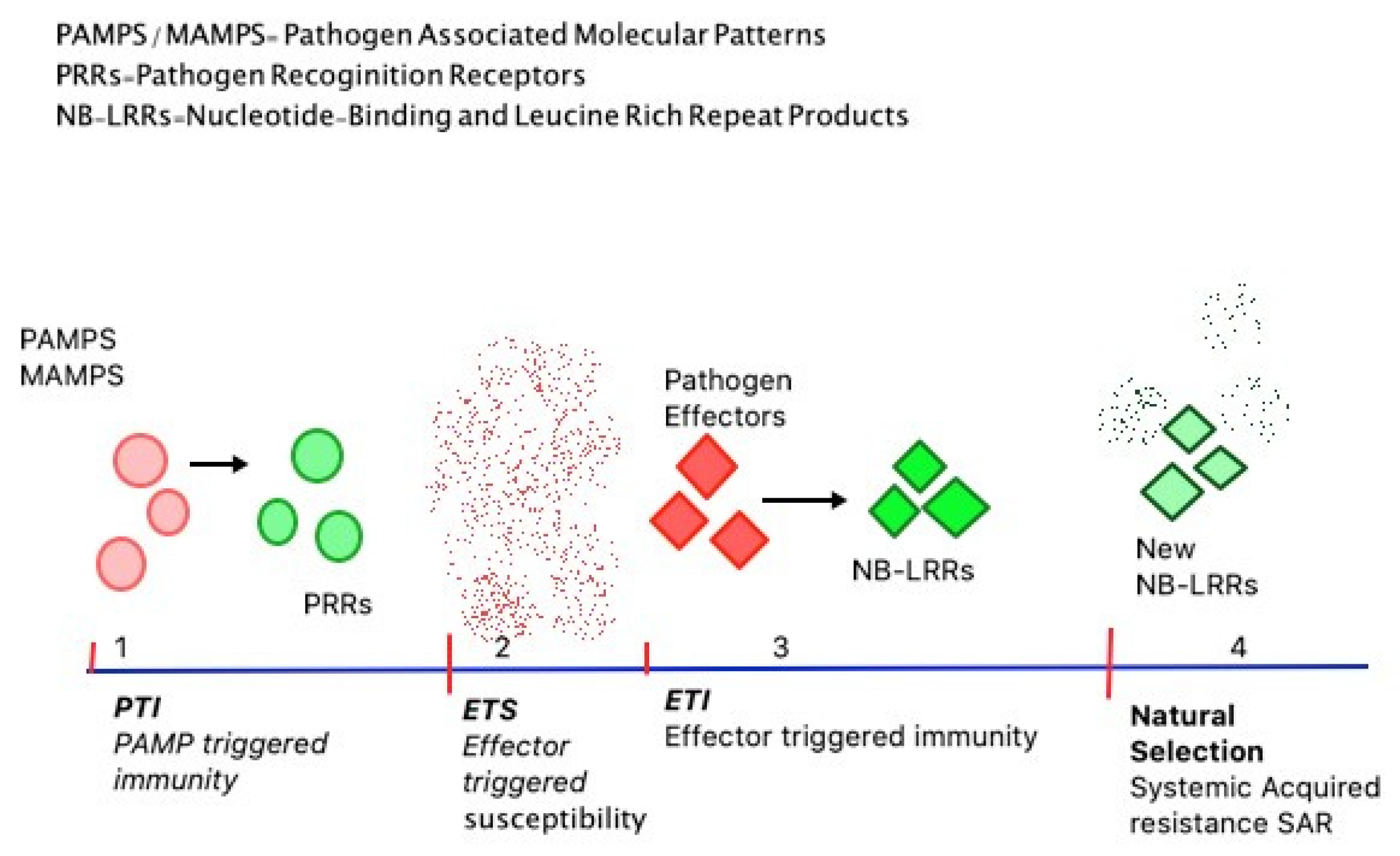
1.1. Plants’ Immune and Defence System
Four Phases of Plants’ Immunity
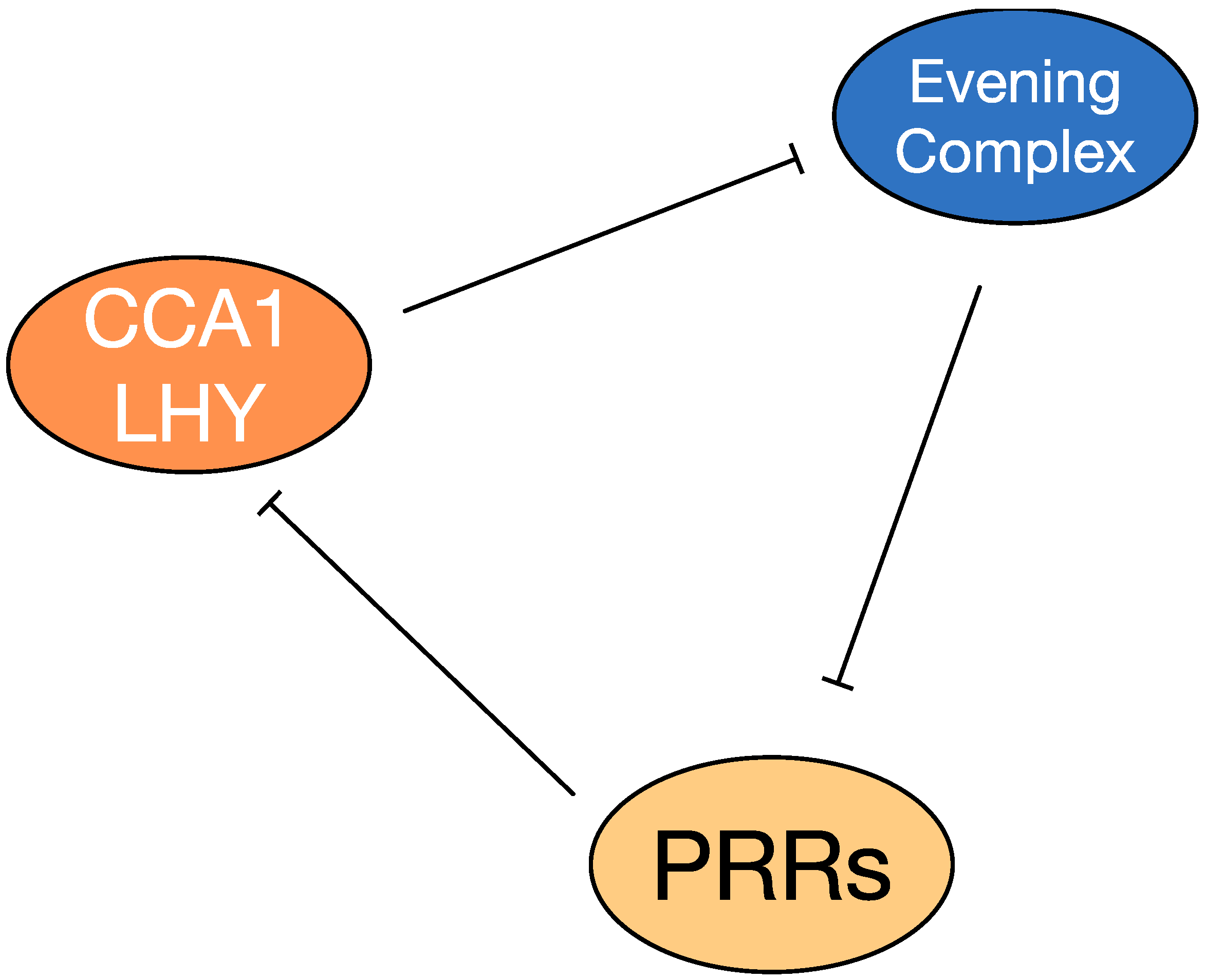
1.2. The Plant Circadian System
2. Presence of Circadian Rhythms in Plant Pathogens
3. Modulation of Plant’s Immune Responses by the Circadian Clock
4. Interactions between the Circadian System, Light, and Plant Defence
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Strange, R.N.; Scott, P.R. Plant Disease: A Threat to Global Food Security. Annu. Rev. Phytopathol. 2005, 43, 83–116. [Google Scholar] [CrossRef] [PubMed]
- Scala, V.; Pucci, N.; Loreti, S. The diagnosis of plant pathogenic bacteria: A state of art. Front. Biosci. 2018, 10, 449–460. [Google Scholar]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [PubMed]
- George Agrios. Plant Pathology; Elseiver: San Diego, CA, USA, 2005; pp. 4–42. [Google Scholar]
- Hua, J. Modulation of plant immunity by light, circadian rhythm, and temperature. Curr. Opin. Plant Biol. 2013, 16, 406–413. [Google Scholar] [CrossRef]
- Fan, G.; Dong, Y.; Deng, M.; Zhao, Z.; Niu, S.; Xu, E. Plant-Pathogen Interaction, Circadian Rhythm, and Hormone-Related Gene Expression Provide Indicators of Phytoplasma Infection in Paulownia fortunei. Int. J. Mol. Sci. 2014, 15, 23141–23162. [Google Scholar] [CrossRef]
- Karapetyan, S.; Dong, X. Redox and the circadian clock in plant immunity: A balancing act. Free. Radic. Biol. Med. 2018, 119, 56–61. [Google Scholar] [CrossRef]
- Lu, H.; McClung, C.R.; Zhang, C. Tick Tock: Circadian Regulation of Plant Innate Immunity. Annu. Rev. Phytopathol. 2017, 55, 287–311. [Google Scholar] [CrossRef]
- Farré, E.M. The regulation of plant growth by the circadian clock. Plant Biol. 2012, 14, 401–410. [Google Scholar] [CrossRef]
- Oakenfull, R.J.; Davis, S.J. Shining a light on the Arabidopsis circadian clock. Plant Cell Environ. 2017, 40, 2571–2585. [Google Scholar] [CrossRef]
- Li, Z.; Bonaldi, K.; Uribe, F.; Pruneda-Paz, J.L. A Localized Pseudomonas syringae Infection Triggers Systemic Clock Responses in Arabidopsis. Curr. Biol. 2018, 28, 630–639.e4. [Google Scholar] [CrossRef]
- Roden, L.C.; Ingle, R.A. Lights, Rhythms, Infection: The Role of Light and the Circadian Clock in Determining the Outcome of Plant–Pathogen Interactions. Plant Cell 2009, 21, 2546–2552. [Google Scholar] [CrossRef] [PubMed]
- McWatters, H.; Devlin, P. Timing in plants—A rhythmic arrangement. FEBS Lett. 2011, 585, 1474–1484. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.A. Retrograde signalling as an informant of circadian timing. New Phytol. 2018, 221, 1749–1753. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.N.G.; Alves, G.S.C.; Van Sluys, M.-A. Plant immunity: Unravelling the complexity of plant responses to biotic stresses. Ann. Bot. 2017, 119, 681–687. [Google Scholar] [CrossRef]
- Westwood, M.L.; O’Donnell, A.J.; De Bekker, C.; Lively, C.M.; Zuk, M.; Reece, S.E. The evolutionary ecology of circadian rhythms in infection. Nat. Ecol. Evol. 2019, 3, 552–560. [Google Scholar] [CrossRef]
- Spoel, S.H.; Dong, X. Making Sense of Hormone Crosstalk during Plant Immune Responses. Cell Host Microbe 2008, 3, 348–351. [Google Scholar] [CrossRef]
- Malik, N.A.A.; Kumar, I.S.; Nadarajah, K. Elicitor and Receptor Molecules: Orchestrators of Plant Defense and Immunity. Int. J. Mol. Sci. 2020, 21, 963. [Google Scholar] [CrossRef]
- Jones, J.; Dangl, J. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef]
- Genoud, T.; Buchala, A.J.; Chua, N.-H.; Métraux, J.-P. Phytochrome signalling modulates the SA-perceptive pathway in Arabidopsis. Plant J. 2002, 31, 87–95. [Google Scholar] [CrossRef]
- Greenberg, J.T.; Yao, N. The role and regulation of programmed cell death in plant-pathogen interactions. Cell. Microbiol. 2004, 6, 201–211. [Google Scholar] [CrossRef]
- Tamaoki, D.; Seo, S.; Yamada, S.; Kano, A.; Miyamoto, A.; Shishido, H.; Miyoshi, S.; Taniguchi, S.; Akimitsu, K.; Gomi, K. Jasmonic acid and salicylic acid activate a common defense system in rice. Plant Signal. Behav. 2013, 8, e24260. [Google Scholar] [CrossRef]
- Loake, G.; Grant, M. Salicylic acid in plant defence—the players and protagonists. Curr. Opin. Plant Biol. 2007, 10, 466–472. [Google Scholar] [CrossRef] [PubMed]
- Bechtold, U.; Karpinski, S.; Mullineaux, P.M. The influence of the light environment and photosynthesis on oxidative signalling responses in plant-biotrophic pathogen interactions. Plant Cell Environ. 2005, 28, 1046–1055. [Google Scholar] [CrossRef]
- Ruan, J.; Zhou, Y.; Zhou, M.; Yan, J.; Khurshid, M.; Weng, W.; Cheng, J.; Zhang, K. Jasmonic Acid Signaling Pathway in Plants. Int. J. Mol. Sci. 2019, 20, 2479. [Google Scholar] [CrossRef] [PubMed]
- McClung, C. Plant Circadian Rhythms. Plant Cell 2006, 18, 792–803. [Google Scholar] [CrossRef]
- Mas, P.; Yanovsky, M.J. Time for circadian rhythms: Plants get synchronized. Curr. Opin. Plant Biol. 2009, 12, 574–579. [Google Scholar] [CrossRef] [PubMed]
- Dodd, A.N.; Salathia, N.; Hall, A.; Kévei, E.; Tóth, R.; Nagy, F.; Hibberd, J.M.; Millar, A.J.; Webb, A.A.R. Plant Circadian Clocks Increase Photosynthesis, Growth, Survival, and Competitive Advantage. Science 2005, 309, 630–633. [Google Scholar] [CrossRef]
- Costantini, C.; Renga, G.; Sellitto, F.; Borghi, M.; Stincardini, C.; Pariano, M.; Zelante, T.; Chiarotti, F.; Bartoli, A.; Mosci, P.; et al. Microbes in the Era of Circadian Medicine. Front. Cell. Infect. Microbiol. 2020, 10, 30. [Google Scholar] [CrossRef]
- Simon, N.M.; Dodd, A.N. A new link between plant metabolism and circadian rhythms? Plant Cell Environ. 2017, 40, 995–996. [Google Scholar] [CrossRef]
- Muranaka, T.; Oyama, T. Monitoring circadian rhythms of individual cells in plants. J. Plant Res. 2017, 131, 15–21. [Google Scholar] [CrossRef]
- Hevia, M.A.; Canessa, P.; Larrondo, L.F. Circadian clocks and the regulation of virulence in fungi: Getting up to speed. Semin. Cell Dev. Biol. 2016, 57, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.; Harmer, S. Wheels within wheels: The plant circadian system. Trends Plant Sci. 2014, 19, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Baudry, A.; Ito, S.; Song, Y.H.; Strait, A.A.; Kiba, T.; Lu, S.; Henriques, R.; Pruneda-Paz, J.L.; Chua, N.-H.; Tobin, E.M.; et al. F-Box Proteins FKF1 and LKP2 Act in Concert with ZEITLUPE to Control Arabidopsis Clock Progression. Plant Cell 2010, 22, 606–622. [Google Scholar] [CrossRef] [PubMed]
- Voitsekhovskaja, O.V. Phytochromes and Other (Photo)Receptors of Information in Plants. Russ. J. Plant Physiol. 2019, 66, 351–364. [Google Scholar] [CrossRef]
- Bendix, C.; Marshall, C.M.; Harmon, F.G. Circadian Clock Genes Universally Control Key Agricultural Traits. Mol. Plant 2015, 8, 1135–1152. [Google Scholar] [CrossRef]
- Tataroglu, O.; Emery, P. Studying circadian rhythms in Drosophila melanogaster. Methods 2014, 68, 140–150. [Google Scholar] [CrossRef]
- Sartor, F.; Eelderink-Chen, Z.; Aronson, B.; Bosman, J.; Hibbert, L.; Dodd, A.; Kovács, Á.; Merrow, M. Are There Circadian Clocks in Non-Photosynthetic Bacteria? Biology 2019, 8, 41. [Google Scholar] [CrossRef]
- Brody, S. Circadian Rhythms in Fungi: Structure/Function/Evolution of Some Clock Components. J. Biol. Rhythm. 2019, 34, 364–379. [Google Scholar] [CrossRef]
- Dunlap, J.C.; Loros, J.; Colot, H.; Mehra, A.; Belden, W.; Shi, M.; Hong, C.; Larrondo, L.; Baker, C.; Chen, C.-H.; et al. A Circadian Clock in Neurospora: How Genes and Proteins Cooperate to Produce a Sustained, Entrainable, and Compensated Biological Oscillator with a Period of about a Day. Cold Spring Harb. Symp. Quant. Biol. 2007, 72, 57–68. [Google Scholar] [CrossRef]
- Bell-Pedersen, D.; Garceau, N.; Loros, J.J. Circadian rhythms in fungi. J. Genet. 1996, 75, 387–401. [Google Scholar] [CrossRef]
- Mishra, S.; Digennaro, P. Root-knot nematodes demonstrate temporal variation in host penetration. J. Nematol. 2020, 52. [Google Scholar] [CrossRef] [PubMed]
- Marsh, E.K.; May, R.C. Caenorhabditis elegans, a Model Organism for Investigating Immunity. Appl. Environ. Microbiol. 2012, 78, 2075–2081. [Google Scholar] [CrossRef] [PubMed]
- Simonetta, S.H.; Migliori, M.L.; Romanowski, A.; Golombek, D.A. Timing of Locomotor Activity Circadian Rhythms in Caenorhabditis elegans. PLOS ONE 2009, 4, e7571. [Google Scholar] [CrossRef]
- Sharma, V.K. On the significance of circadian clocks for insects. J. Indian Inst. Sci. 2002, 83, 3–26. [Google Scholar]
- Meireles-Filho, A.C.A.; Kyriacou, C.P. Circadian rhythms in insect disease vectors. Memórias do Instituto Oswaldo Cruz 2013, 108, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Bhatt, D. The circadian clock and defence signalling in plants. Mol. Plant Pathol. 2014, 16, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Barnaby, J.Y.; Tada, Y.; Li, H.; Tör, M.; Caldelari, D.; Lee, D.-U.; Fu, X.-D.; Dong, X. Timing of plant immune responses by a central circadian regulator. Nat. Cell Biol. 2011, 470, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Goodspeed, D.; Chehab, E.W.; Min-Venditti, A.; Braam, J.; Covington, M.F. Cozzarelli Prize Winner: Arabidopsis synchronizes jasmonate-mediated defense with insect circadian behavior. Proc. Natl. Acad. Sci. USA 2012, 109, 4674–4677. [Google Scholar] [CrossRef]
- Goodspeed, D.; Chehab, E.W.; Covington, M.F.; Braam, J. Circadian control of jasmonates and salicylates. Plant Signal. Behav. 2013, 8, e23123. [Google Scholar] [CrossRef]
- Zheng, S.X.; Xiao, S.; Chye, M. The gene encoding Arabidopsis acyl-CoA-binding protein 3 is pathogen inducible and subject to circadian regulation. J. Exp. Bot. 2012, 63, 2985–3000. [Google Scholar] [CrossRef]
- Zhang, J.; Ren, Z.; Zhou, Y.; Ma, Z.; Ma, Y.; Hou, D.; Xu, Z.; Huang, X. NPR1 and Redox Rhythms: Connections, between Circadian Clock and Plant Immunity. Int. J. Mol. Sci. 2019, 20, 1211. [Google Scholar] [CrossRef] [PubMed]
- Weyman, P.D.; Pan, Z.; Feng, Q.; Gilchrist, D.G.; Bostock, R.M. A Circadian Rhythm-Regulated Tomato Gene Is Induced by Arachidonic Acid and Phythophthora infestans Infection. Plant Physiol. 2005, 140, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wang, M.-M.; Yin, Y.-L.; Onac, E.; Zhou, G.-F.; Peng, S.; Xia, X.-J.; Shi, K.; Yu, J.-Q.; Zhou, Y. RNA-seq analysis reveals the role of red light in resistance against Pseudomonas syringae pv. tomato DC3000 in tomato plants. BMC Genom. 2015, 16, 120. [Google Scholar] [CrossRef] [PubMed]
- Korneli, C.; Danisman, S.; Staiger, D. Differential Control of Pre-Invasive and Post-Invasive Antibacterial Defense by the Arabidopsis Circadian Clock. Plant Cell Physiol. 2014, 55, 1613–1622. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, V.; Meier, S.; Petersen, L.N.; Ingle, R.A.; Roden, L.C. Defence Responses of Arabidopsis thaliana to Infection by Pseudomonas syringae Are Regulated by the Circadian Clock. PLoS ONE 2011, 6, e26968. [Google Scholar] [CrossRef] [PubMed]
- De Leone, M.J.; Hernando, C.E.; Romanowski, A.; Careno, D.A.; Soverna, A.F.; Sun, H.; Bologna, N.G.; Vázquez, M.; Schneeberger, K.; Yanovsky, M.J. Bacterial Infection Disrupts Clock Gene Expression to Attenuate Immune Responses. Curr. Biol. 2020, 30, 1740–1747.e6. [Google Scholar] [CrossRef] [PubMed]
- Griebel, T.; Zeier, J. Light Regulation and Daytime Dependency of Inducible Plant Defenses in Arabidopsis: Phytochrome Signaling Controls Systemic Acquired Resistance Rather Than Local Defense. Plant Physiol. 2008, 147, 790–801. [Google Scholar] [CrossRef]
- Pink, B.; Mueller, M.J.; Berger, S. Light conditions influence specific defence responses in incompatible plant?pathogen interactions: Uncoupling systemic resistance from salicylic acid and PR-1 accumulation. Planta 2004, 219, 673–683. [Google Scholar] [CrossRef]
- Gallé, Á.; Czékus, Z.; Bela, K.; Horváth, E.; Ördög, A.; Csiszár, J.; Poór, P. Plant Glutathione Transferases and Light. Front. Plant Sci. 2019, 9, 1994. [Google Scholar] [CrossRef]
- Chandra-Shekara, A.C.; Gupte, M.; Navarre, D.; Raina, S.; Raina, R.; Klessig, D.; Kachroo, P. Light-dependent hypersensitive response and resistance signaling against Turnip Crinkle Virus in Arabidopsis. Plant J. 2006, 45, 320–334. [Google Scholar] [CrossRef]
- Moghaddam, M.R.B.; Ende, W.V.D. Sweet immunity in the plant circadian regulatory network. J. Exp. Bot. 2013, 64, 1439–1449. [Google Scholar] [CrossRef] [PubMed]
- Martinez, M.; Helm, B. The influence of biological rhythms on host–parasite interactions. Trends Ecol. Evol. 2015, 30, 314–326. [Google Scholar] [CrossRef] [PubMed]
- Seifalian, A.; Hart, A. Circadian Rhythms: Will It Revolutionise the Management of Diseases? J. Lifestyle Med. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rijo-Ferreira, F.; Takahashi, J.S. Genomics of circadian rhythms in health and disease. Genome Med. 2019, 11, 1–16. [Google Scholar] [CrossRef]
- Zhang, R.; Lahens, N.F.; Ballance, H.I.; Hughes, M.E.; HogenEsch, J.B. A circadian gene expression atlas in mammals: Implications for biology and medicine. Proc. Natl. Acad. Sci. USA 2014, 111, 16219–16224. [Google Scholar] [CrossRef]
- Scheiermann, C.; Gibbs, J.; Ince, L.; Loudon, A.S.I. Clocking in to immunity. Nat. Rev. Immunol. 2018, 18, 423–437. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Butt, G.R.; Qayyum, Z.A.; Jones, M.A. Plant Defence Mechanisms Are Modulated by the Circadian System. Biology 2020, 9, 454. https://doi.org/10.3390/biology9120454
Butt GR, Qayyum ZA, Jones MA. Plant Defence Mechanisms Are Modulated by the Circadian System. Biology. 2020; 9(12):454. https://doi.org/10.3390/biology9120454
Chicago/Turabian StyleButt, Ghazala Rauf, Zainab Abdul Qayyum, and Matthew Alan Jones. 2020. "Plant Defence Mechanisms Are Modulated by the Circadian System" Biology 9, no. 12: 454. https://doi.org/10.3390/biology9120454
APA StyleButt, G. R., Qayyum, Z. A., & Jones, M. A. (2020). Plant Defence Mechanisms Are Modulated by the Circadian System. Biology, 9(12), 454. https://doi.org/10.3390/biology9120454