Determining Soil Microbial Communities and Their Influence on Ganoderma Disease Incidences in Oil Palm (Elaeis guineensis) via High-Throughput Sequencing

 , , and
, , and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
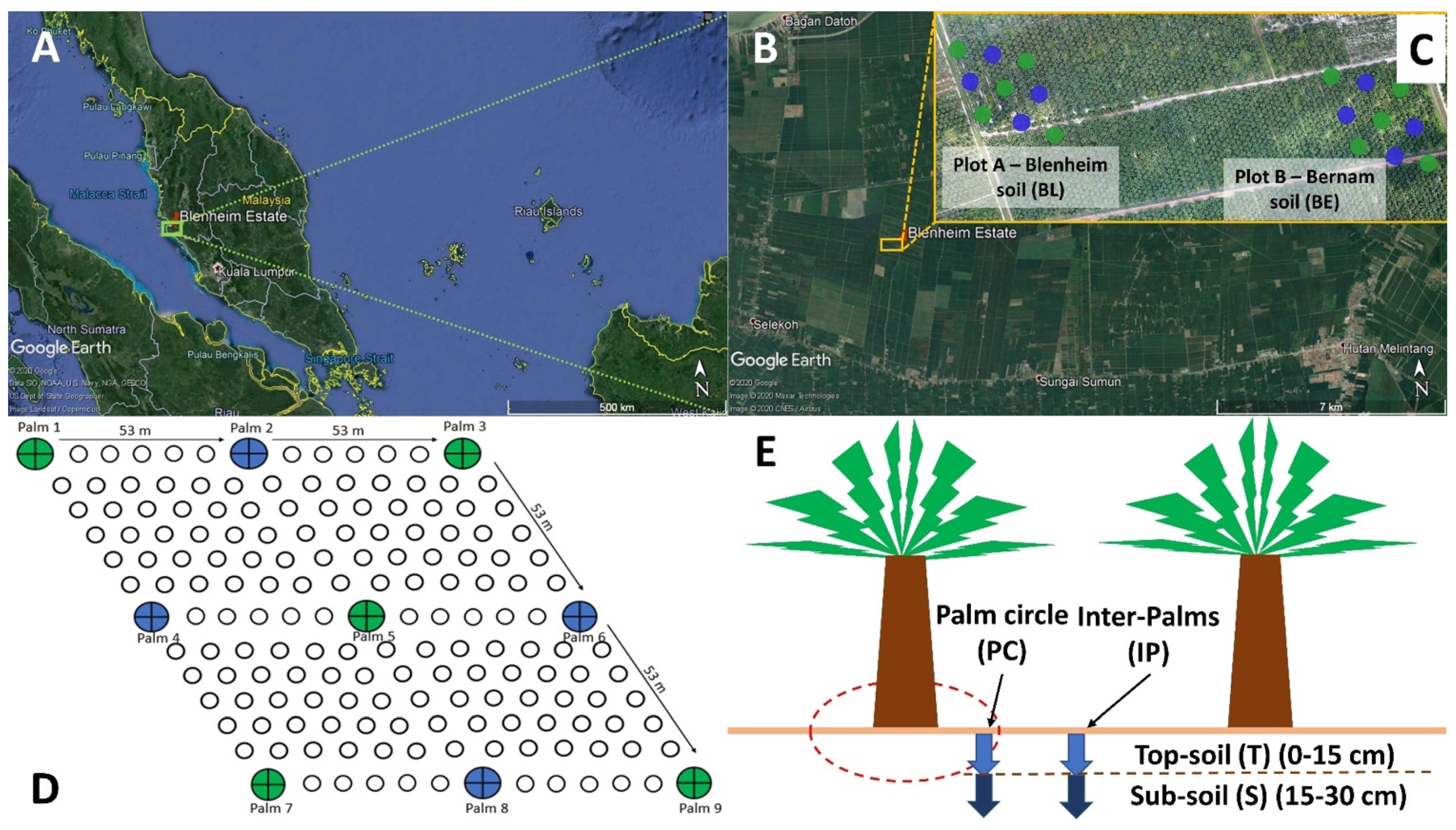
2.1. Site and Soil Collection
2.2. Field BSR Disease Census
2.3. Soil Physicochemical Analyses
2.4. Soil DNA Extraction, Library Preparation, Purification, and MiSeq Sequencing
2.5. Sequence Processing and Analyses
2.6. Statistical and Data Analyses
3. Results
3.1. Ganoderma Incidences in Blenheim and Bernam Soils
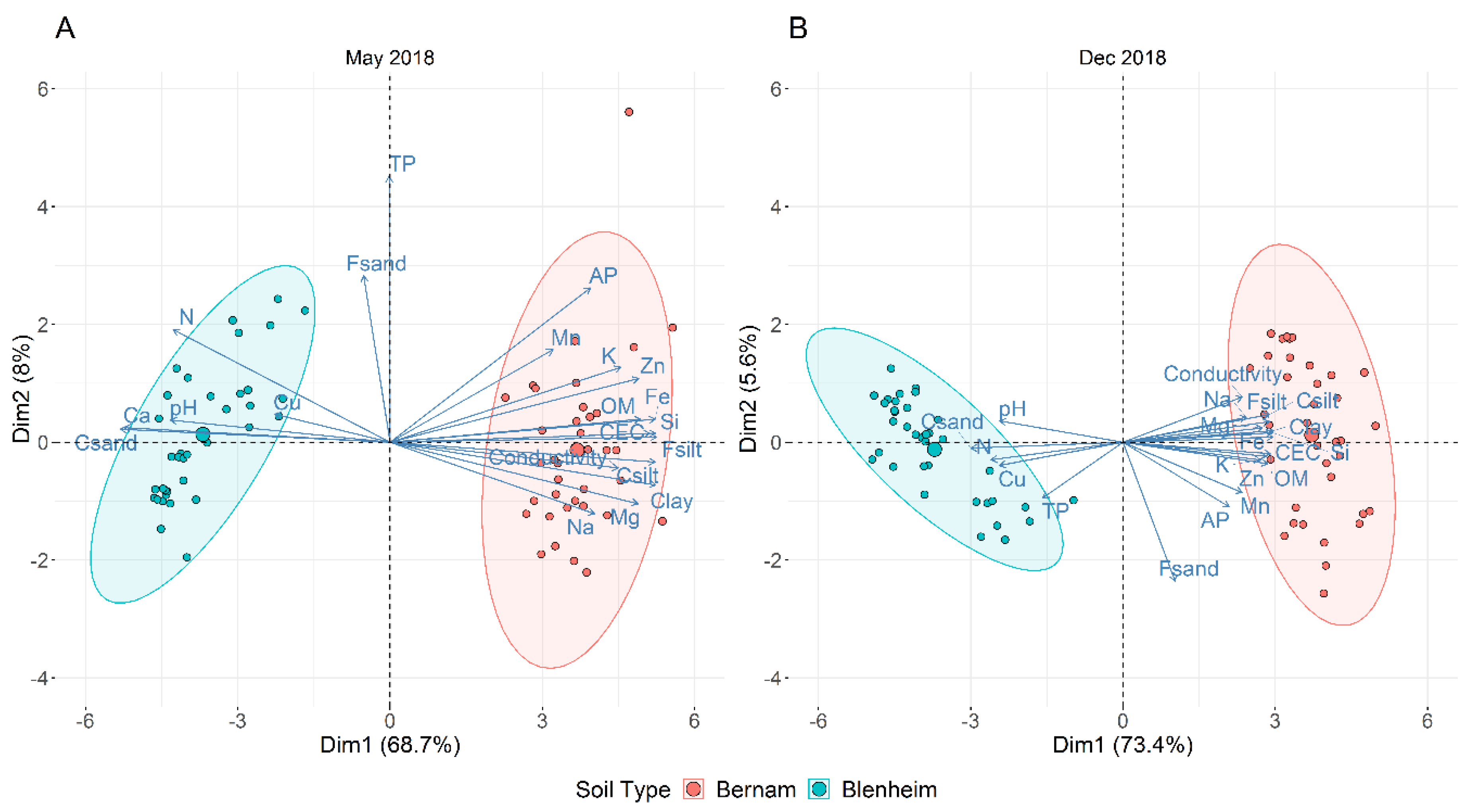
3.2. Physicochemical Analyses of Blenheim and Bernam Soils
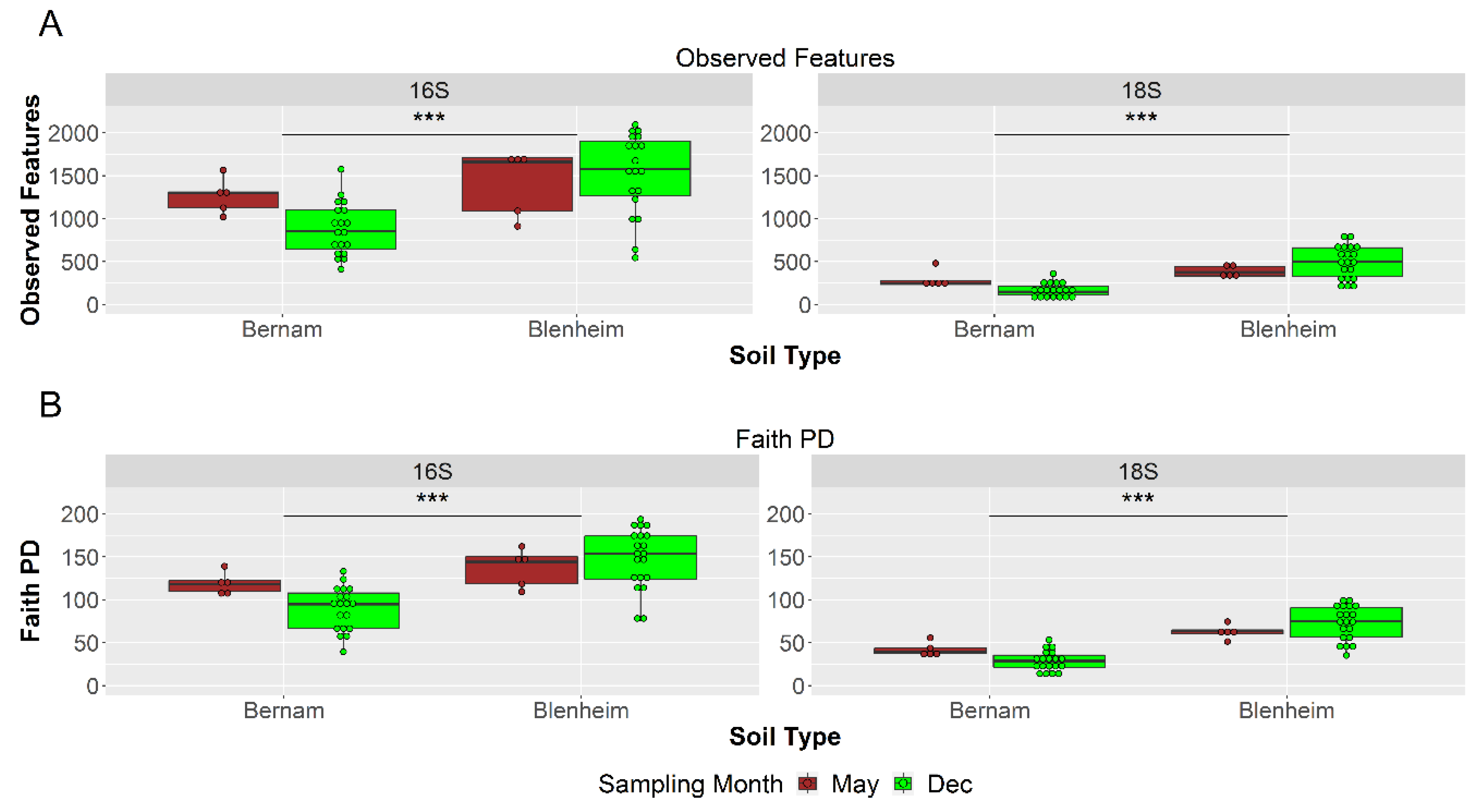
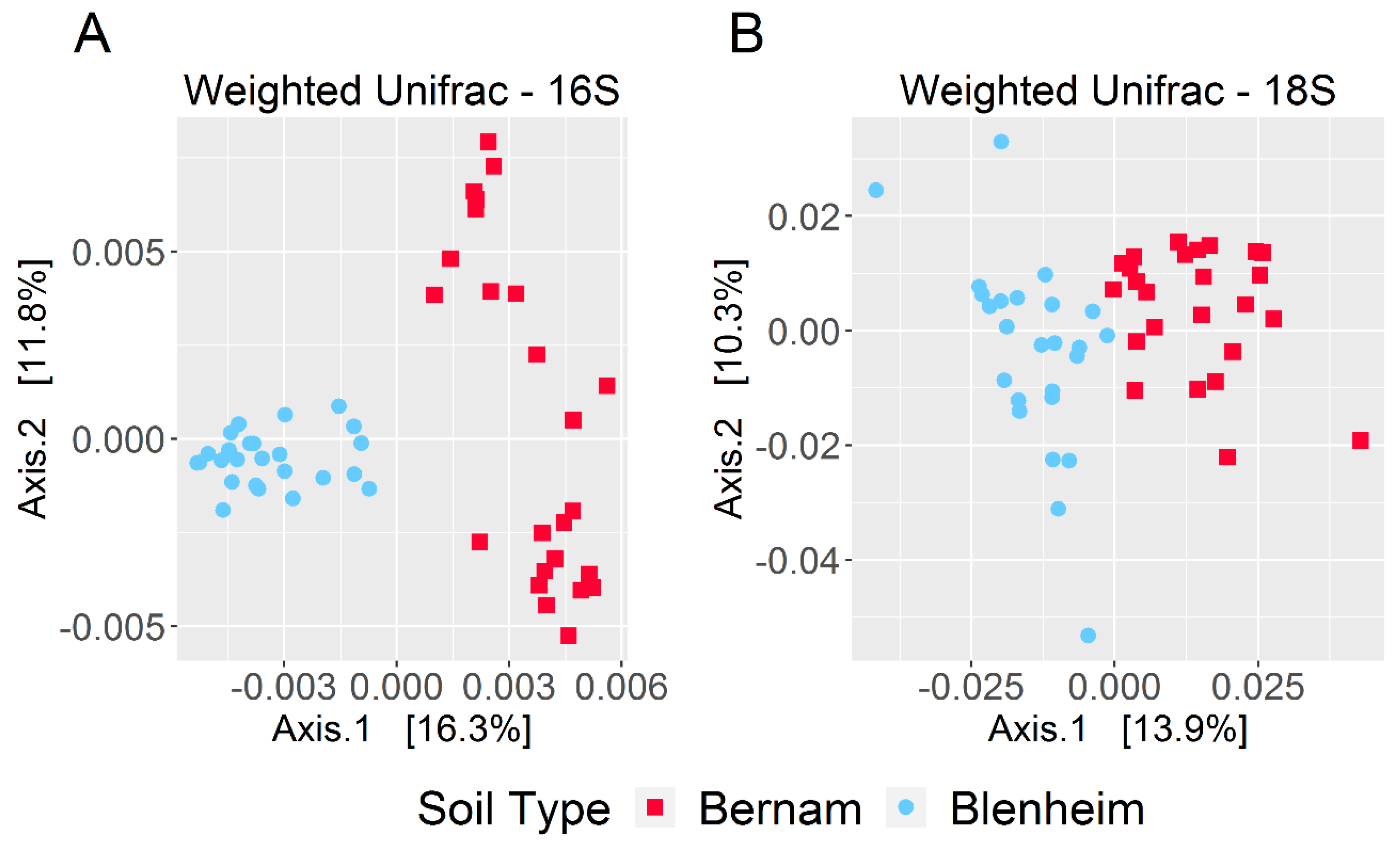
3.3. Prokaryotic and Eukaryotic Richness and Diversity in Blenheim and Bernam Soils
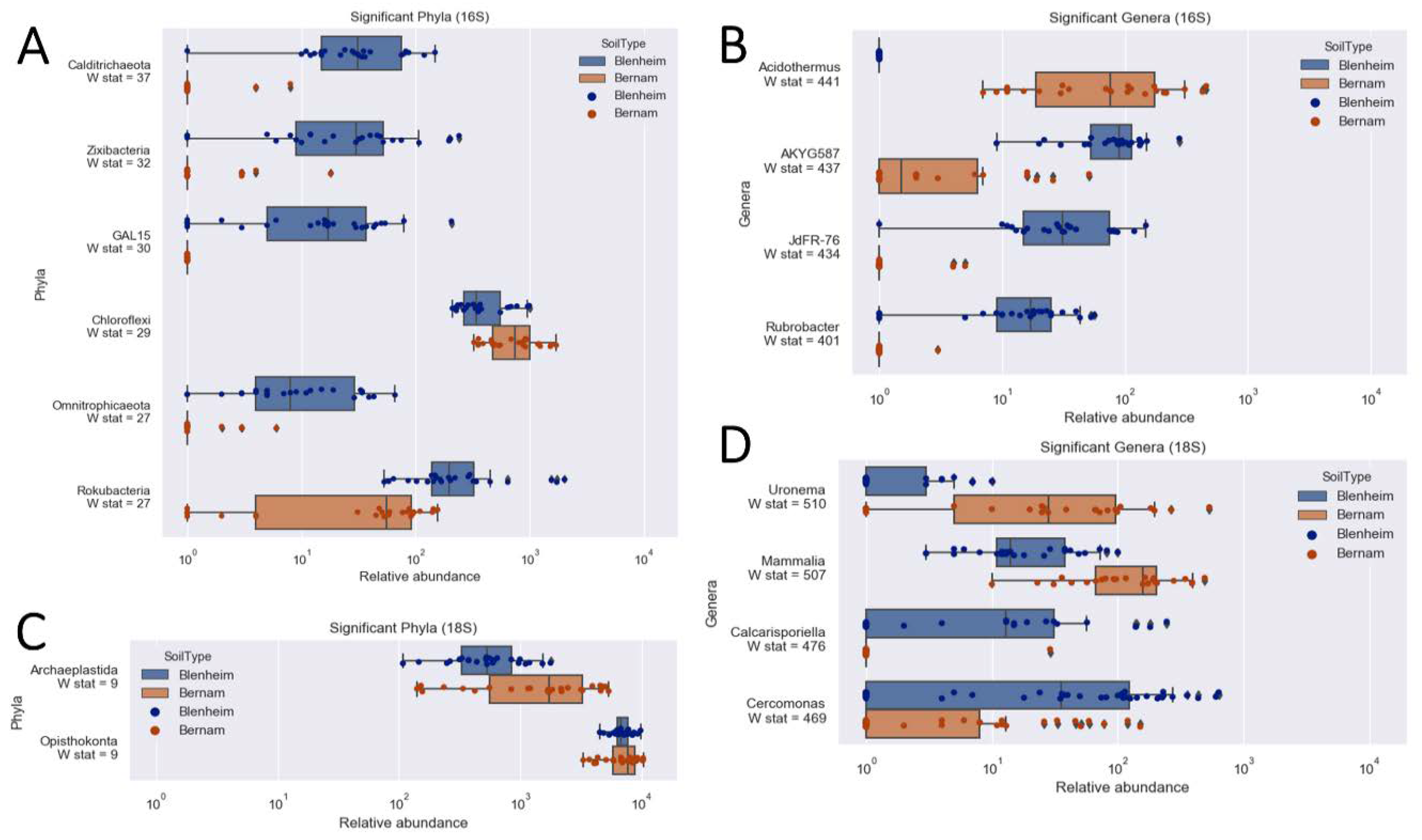
3.4. Prokaryotic Microbial Community at Phylum and Genus Levels
3.5. Eukaryotic Microbial Community Structure at Supergroup and Genus or Class Levels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Murphy, D.J. The future of oil palm as a major global crop: Opportunities and challenges. J. Oil Palm Res. 2014, 26, 1–24. [Google Scholar]
- Woittiez, L.S.; van Wijk, M.T.; Slingerland, M.; van Noordwijk, M.; Giller, K.E. Yield gaps in oil palm: A quantitative review of contributing factors. Eur. J. Agron. 2017, 83, 57–77. [Google Scholar] [CrossRef]
- MPOB. Malaysian Oil Palm Statistics 2018, 38th ed.; Malaysian Palm Oil Board: Bangi, Malaysia, 2019.
- Kushairi, A.; Ong-Abdullah, M.; Nambiappan, B.; Hishamuddin, E.; Bidin, M.N.I.Z.; Ghazalia, R.; Subramaniam, V.; Sundram, S.; Parveez, G.K.A. Oil palm economic performance in Malaysia and R&D progress in 2018. J. Oil Palm Res. 2019, 31, 165–194. [Google Scholar] [CrossRef]
- Corley, R.H.V.; Tinker, P.B. The Oil Palm, 5th ed.; Blackwell Publishing: Oxford, UK, 2016. [Google Scholar]
- Rees, R.W.; Flood, J.; Hasan, Y.; Wills, M.A.; Cooper, R.M. Ganoderma boninense basidiospores in oil palm plantations: Evaluation of their possible role in stem rots of Elaeis guineensis. Plant Pathol. 2012, 61, 567–578. [Google Scholar] [CrossRef]
- Turner, P.D. Oil Palm Diseases and Disorders; Oxford University Press: Oxford, UK, 1981. [Google Scholar]
- Hushiarian, R.; Yusof, N.A.; Dutse, S.W. Detection and control of Ganoderma boninense: Strategies and perspectives. Springerplus 2013, 2, 555. [Google Scholar] [CrossRef]
- Cooper, R.M.; Flood, J.; Rees, R.W. Ganoderma boninense in oil palm plantations: Current thinking on epidemiology, resistance and pathology. Planter 2011, 87, 515–526. [Google Scholar]
- Ariffin, D.; Idris, A.S.; Singh, G. Status of Ganoderma in oil palm. In Ganoderma Diseases of Perennial Crops; Flood, J., Bridge, P.D., Holderness, M., Eds.; CABI Publishing: Wallingford, UK, 2000. [Google Scholar]
- Singh, G. Ganoderma-the scourge of oil palms in the coastal areas. Planter 1991, 67, 421–444. [Google Scholar]
- Naher, L.; Yusuf, U.K.; Ismail, A.; Tan, S.G.; Mondal, M.M.A. Ecological status of Ganoderma and basal stem rot disease of oil palms (Elaeis guineensis Jacq.). Aust. J. Crop Sci. 2013, 7, 1723–1727. [Google Scholar] [CrossRef]
- Chung, G.F. Management of Ganoderma diseases in oil palm plantations. Planter 2011, 87, 325–339. [Google Scholar]
- Norman, K.; Idris, A.S.; Mohd Mazmira, M.M. Prospects in sustainable control of oil palm pests and diseases through the enhancement of ecosystem services—The way forward. J. Oil Palm Res. 2019, 31, 381–393. [Google Scholar] [CrossRef]
- Susanto, A.; Sudharto, P.; Purba, R. Enhancing biological control of basal stem rot disease (Ganoderma boninense) in oil palm plantations. Mycopathologia 2005, 159, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Sapak, Z.; Meon, S.; Ahmad, Z.M.A. Effect of endophytic bacteria on growth and suppression of Ganoderma infected in oil palm. Int. J. Agric. Biol. 2008, 10, 127–132. [Google Scholar]
- Muniroh, M.S.; Nusaibah, S.A.; Vadamalai, G.; Siddique, Y. Proficiency of biocontrol agents as plant growth promoters and hydrolytic enzyme producers in Ganoderma boninense infected oil palm seedlings. Curr. Plant Biol. 2019, 20, 100116. [Google Scholar] [CrossRef]
- Breton, F.; Hasan, Y.; Hariadi; Lubis, Z.; de Franqueville, H. Characterization of parameters for the development of an early screening test for basal stem rot tolerance in oil palm progenies. J. Oil Palm Res. 2006, 24–36. [Google Scholar]
- Idris, A.; Kushairi, A.; Ismail, S.; Ariffin, D. Selection for partial resistance in oil palm progenies to Ganoderma basal stem rot. J. Oil Palm Res. 2004, 16, 12–18. [Google Scholar]
- Durand-Gasselin, T.; Asmady, H.; Flori, A.; Jacquemard, J.C.; Hayun, Z.; Breton, F.; de Franqueville, H. Possible sources of genetic resistance in oil palm (Elaeis guineensis Jacq.) to basal stem rot caused by Ganoderma boninense—Prospects for future breeding. Mycopathologia 2005, 159, 93–100. [Google Scholar] [CrossRef]
- Chaparro, J.M.; Sheflin, A.M.; Manter, D.K.; Vivanco, J.M. Manipulating the soil microbiome to increase soil health and plant fertility. Biol. Fert. Soils 2012, 48, 489–499. [Google Scholar] [CrossRef]
- Tian, L.; Lin, X.; Tian, J.; Ji, L.; Chen, Y.; Tran, L.-S.P.; Tian, C. Research advances of beneficial microbiota associated with crop plants. Int. J. Mol. Sci. 2020, 21, 1792. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef]
- Wei, Z.; Gu, Y.; Friman, V.-P.; Kowalchuk, G.A.; Xu, Y.; Shen, Q.; Jousset, A. Initial soil microbiome composition and functioning predetermine future plant health. Sci. Adv. 2019, 5, eaaw0759. [Google Scholar] [CrossRef]
- Schlatter, D.; Kinkel, L.; Thomashow, L.; Weller, D.; Paulitz, T. Disease suppressive soils: New insights from the soil microbiome. Phytopathology 2017, 107, 1284–1297. [Google Scholar] [CrossRef] [PubMed]
- Janvier, C.; Villeneuve, F.; Alabouvette, C.; Edel-Hermann, V.; Mateille, T.; Steinberg, C. Soil health through soil disease suppression: Which strategy from descriptors to indicators? Soil Biol. Biochem. 2007, 39, 1–23. [Google Scholar] [CrossRef]
- Mendes, R.; Kruijt, M.; de Bruijn, I.; Dekkers, E.; van der Voort, M.; Schneider, J.H.M.; Piceno, Y.M.; DeSantis, T.Z.; Andersen, G.L.; Bakker, P.A.H.M.; et al. Deciphering the rhizosphere microbiome for disease-suppressive bacteria. Science 2011, 332, 1097. [Google Scholar] [CrossRef] [PubMed]
- Ros, M.; Almagro, M.; Fernández, J.A.; Egea-Gilabert, C.; Faz, Á.; Pascual, J.A. Approaches for the discrimination of suppressive soils for Pythium irregulare disease. Appl. Soil Ecol. 2020, 147, 103439. [Google Scholar] [CrossRef]
- Siegel-Hertz, K.; Edel-Hermann, V.; Chapelle, E.; Terrat, S.; Raaijmakers, J.M.; Steinberg, C. Comparative microbiome analysis of a Fusarium wilt suppressive soil and a Fusarium wilt conducive soil from the Châteaurenard region. Front. Microbiol. 2018, 9, 568. [Google Scholar] [CrossRef] [PubMed]
- Schulz, G.; Schneider, D.; Brinkmann, N.; Edy, N.; Daniel, R.; Polle, A.; Scheu, S.; Krashevska, V. Changes in trophic groups of protists with conversion of rainforest into rubber and oil palm plantations. Front. Microbiol. 2019, 10, 240. [Google Scholar] [CrossRef] [PubMed]
- Tin, H.S.; Palaniveloo, K.; Anilik, J.; Vickneswaran, M.; Tashiro, Y.; Vairappan, C.S.; Sakai, K. Impact of land-use change on vertical soil bacterial communities in Sabah. Microb. Ecol. 2018, 75, 459–467. [Google Scholar] [CrossRef]
- Tripathi, B.M.; Edwards, D.P.; Mendes, L.W.; Kim, M.; Dong, K.; Kim, H.; Adams, J.M. The impact of tropical forest logging and oil palm agriculture on the soil microbiome. Mol. Ecol. 2016, 25, 2244–2257. [Google Scholar] [CrossRef]
- Wood, S.A.; Gilbert, J.A.; Leff, J.W.; Fierer, N.; D’Angelo, H.; Bateman, C.; Gedallovich, S.M.; Gillikin, C.M.; Gradoville, M.R.; Mansor, P.; et al. Consequences of tropical forest conversion to oil palm on soil bacterial community and network structure. Soil Biol. Biochem. 2017, 112, 258–268. [Google Scholar] [CrossRef]
- Rao, V.; Lim, C.C.; Chia, C.C.; Teo, K.W. Studies on Ganoderma spread and control. Planter 2003, 79, 367–383. [Google Scholar]
- Paramananthan, S. Soils of Malaysia: Their Characteristics and Identification; Academy of Sciences Malaysia: Kuala Lumpur, Malaysia, 2000; Volume 1, p. 616. [Google Scholar]
- Soil Survey Staff. Keys to Soil Taxonomy—Twelfth Edition; Soil Conservation Service: Washington, DC, USA, 2014. [Google Scholar]
- Strohm, J.H.T. The Influence of Agricultural Tillage Practices on Soil Biodiversity. Master’s Thesis, The University of Guelph, Guelph, ON, Canada, 2015. [Google Scholar]
- Chen, Z.Y.; Goh, Y.K.; Goh, Y.K.; Goh, K.J. Life expectancy of oil palm (Elaeis guineensis) infected by Ganoderma boninense in coastal soils, Malaysia: A case study. Arch. Phytopathol. Pflanzenschutz 2017, 50, 598–612. [Google Scholar] [CrossRef]
- Campbell, C.L.; Madden, L.V. Introduction to Plant Disease Epidemiology; John Wiley & Sons: New York, NY, USA, 1990; p. 532. [Google Scholar]
- Murakami, H.; Tsushima, S.; Kuroyanagi, Y.; Shishido, Y. Reduction of resting spore density of Plasmodiophora brassicae and clubroot disease severity by liming. Soil Sci. Plant Nutr. 2002, 48, 685–691. [Google Scholar] [CrossRef]
- Kome, G.K.; Enang, R.K.; Yerima, B.P.K.; Lontsi, M.G.R. Models relating soil pH measurements in H2O, KCl and CaCl2 for volcanic ash soils of Cameroon. Geoderma Reg. 2018, 14, e00185. [Google Scholar] [CrossRef]
- Behera, S.K.; Suresh, K.; Rao, B.N.; Manoja, K.; Manorama, K. Soil nutrient status and leaf nutrient norms in oil palm (Elaeis guineensis Jacq.) plantations grown in the West Coastal area of India. Commun. Soil Sci. Plant Anal. 2015, 47, 255–262. [Google Scholar] [CrossRef]
- Jones, J.B.J. Laboratory Guide for Conducting Soil Tests and Plant Analysis; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Ng, P.H.C.; Gan, H.H.; Goh, K.J. Soil nutrient changes in Ultisols under oil palm in Johor, Malaysia. J. Oil Palm Environ. 2011, 2, 93–104. [Google Scholar] [CrossRef]
- Moris, N.; Mohinder, S.S. Manual of Laboratory Methods of Chemical Soil Analysis; Rubber Research Institute Malaysia (RRIM): Kuala Lumpur, Malaysia, 1980. [Google Scholar]
- Sharifuddin, H.A.H.; Fauziah, I.; Zaharah, A.R. Technique of soil testing and plant analysis and their utilization for crop production in malaysia. Commun. Soil Sci. Plant Anal. 1990, 21, 1959–1978. [Google Scholar] [CrossRef]
- Lim, H.K. Working Manual for Soil Analysis; Ministry of Agriculture: Kuala Lumpur, Malaysia, 1975. [Google Scholar]
- Burt, R. Soil Survey Laboratory Methods Manual: Soil Survey Investigations Report No. 42; United States Department of Agriculture (USDA)-Natural Resources Conservation Service (NRCS): Washington, DC, USA, 2004; p. 736. [Google Scholar]
- Courty, P.E.; Buée, M.; Tech, J.J.T.; Brulé, D.; Colin, Y.; Leveau, J.H.J.; Uroz, S. Impact of soil pedogenesis on the diversity and composition of fungal communities across the California soil chronosequence of Mendocino. Mycorrhiza 2018, 28, 343–356. [Google Scholar] [CrossRef]
- Herlemann, D.P.R.; Labrenz, M.; Jürgens, K.; Bertilsson, S.; Waniek, J.J.; Andersson, A.F. Transitions in bacterial communities along the 2000 km salinity gradient of the Baltic Sea. ISME J. 2011, 5, 1571–1579. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2012, 41, e1. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S sequencing. In Nucleic Acid Technologies in Bacterial Systematic; Stackebrant, E., Goodfellow, M., Eds.; Wiley: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Medlin, L.; Elwood, H.J.; Stickel, S.; Sogin, M.L. The characterization of enzymatically amplified eukaryotic 16S-like rRNA-coding regions. Gene 1988, 71, 491–499. [Google Scholar] [CrossRef]
- Amaral-Zettler, L.A.; McCliment, E.A.; Ducklow, H.W.; Huse, S.M. A method for studying protistan diversity using massively parallel sequencing of V9 hypervariable regions of small-subunit ribosomal RNA genes. PLoS ONE 2009, 4, e6372. [Google Scholar] [CrossRef]
- Ong, S.H.; Kukkillaya, V.U.; Wilm, A.; Lay, C.; Ho, E.X.P.; Low, L.; Hibberd, M.L.; Nagarajan, N. Species identification and profiling of complex microbial communities using shotgun Illumina sequencing of 16S rRNA amplicon sequences. PLoS ONE 2013, 8, e60811. [Google Scholar] [CrossRef] [PubMed]
- Watts, M.P.; Spurr, L.P.; Gan, H.M.; Moreau, J.W. Characterization of an autotrophic bioreactor microbial consortium degrading thiocyanate. Appl. Microbiol. Biotechnol. 2017, 101, 5889–5901. [Google Scholar] [CrossRef] [PubMed]
- Md Zoqratt, M.Z.H.; Eng, W.W.H.; Thai, B.T.; Austin, C.M.; Gan, H.M. Microbiome analysis of Pacific white shrimp gut and rearing water from Malaysia and Vietnam: Implications for aquaculture research and management. Peer J. 2018, 6, e5826. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.-i.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2′s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine learning in python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar] [CrossRef]
- Pruesse, E.; Peplies, J.; Glöckner, F.O. SINA: Accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 2012, 28, 1823–1829. [Google Scholar] [CrossRef] [PubMed]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [PubMed]
- Faith, D.P. Conservation evaluation and phylogenetic diversity. Biol. Conserv. 1992, 61, 1–10. [Google Scholar] [CrossRef]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and qualitative beta diversity measures lead to different insights into factors that structure microbial communities. Appl. Environ. Microbiol. 2007, 73, 1576–1585. [Google Scholar] [CrossRef]
- Wickham, H.; Wickham, M.H. The Ggplot Package. Available online: https://cran.r-project.org/web/packages/ggplot2/index.html (accessed on 16 March 2020).
- RStudio Team. RStudio: Integrated Development for R. Available online: http://www.rstudio.com/ (accessed on 16 March 2020).
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Mandal, S.; Van Treuren, W.; White, R.A.; Eggesbø, M.; Knight, R.; Peddada, S.D. Analysis of composition of microbiomes: A novel method for studying microbial composition. Microb. Ecol. Health Dis. 2015, 26, 27663. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Liang, W.; Shi, Y.; Lin, X.; Zhang, H.; Wu, X.; Xie, G.; Chain, P.; Grogan, P.; Chu, H. Contrasting elevational diversity patterns between eukaryotic soil microbes and plants. Ecology 2014, 95, 3190–3202. [Google Scholar] [CrossRef]
- Zhalnina, K.; Dias, R.; de Quadros, P.D.; Davis-Richardson, A.; Camargo, F.A.O.; Clark, I.M.; McGrath, S.P.; Hirsch, P.R.; Triplett, E.W. Soil pH determines microbial diversity and composition in the park grass experiment. Microb. Ecol. 2015, 69, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zhang, H.; Sun, L.; Qi, G.; Chen, S.; Zhao, X. Microbial community composition is related to soil biological and chemical properties and bacterial wilt outbreak. Sci. Rep. 2017, 7, 343. [Google Scholar] [CrossRef] [PubMed]
- Rousk, J.; Brookes, P.C.; Bååth, E. Contrasting soil pH effects on fungal and bacterial growth suggest functional redundancy in carbon mineralization. Appl. Environ. Microbiol. 2009, 75, 1589. [Google Scholar] [CrossRef]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef]
- Shamshuddin, J.; Paramananthan, S. Acidity and charge characteristics of marine alluvial soils from Carey Islands, Selangor. Pertanika 1988, 11, 15–23. [Google Scholar]
- Lynch, J.M. Microbial activity in acid soils. In Plant-Soil Interactions at Low pH: Principles and Management: Proceedings of the Third International Symposium on Plant-Soil Interactions at Low pH, Brisbane, Queensland, Australia, 12–16 September 1993; Date, R.A., Grundon, N.J., Rayment, G.E., Probert, M.E., Eds.; Springer: Dordrecht, The Netherlands, 1995; pp. 167–172. [Google Scholar] [CrossRef]
- Ownley, B.H.; Duffy, B.K.; Weller, D.M. Identification and manipulation of soil properties to improve the biological control performance of phenazine-producing Pseudomonas fluorescens. Appl. Environ. Microbiol. 2003, 69, 3333. [Google Scholar] [CrossRef]
- Li, S.; Liu, Y.; Wang, J.; Yang, L.; Zhang, S.; Xu, C.; Ding, W. Soil acidification aggravates the occurrence of bacterial wilt in South China. Front. Microbiol. 2017, 8, 703. [Google Scholar] [CrossRef]
- Duffy, B.K.; Ownley, B.H.; Weller, D.M. Soil chemical and physical properties associated with suppression of Take-all of wheat by Trichoderma koningii. Phytopathology 1997, 87, 1118–1124. [Google Scholar] [CrossRef]
- Kredics, L.; Antal, Z.; Manczinger, L.; Szekeres, A.; Kevei, F.; Nagy, E. Influence of environmental parameters on Trichoderma strains with biocontrol potential. Food Technol. Biotechnol. 2003, 41, 37–42. [Google Scholar]
- Rahman, K.A.; Othman, R. Influence of pH levels on disease development in oil palm seedling roots infected with Ganoderma boninense. Rhizosphere 2020, 13, 100181. [Google Scholar] [CrossRef]
- Nawawi, A.; Ho, Y.W. Effect of temperature and pH on growth pattern of Ganoderma boninense from oil palm in Peninsular Malaysia. Pertanika 1990, 13, 303–307. [Google Scholar]
- Marshall, I.P.G.; Starnawski, P.; Cupit, C.; Fernández Cáceres, E.; Ettema, T.J.G.; Schramm, A.; Kjeldsen, K.U. The novel bacterial phylum Calditrichaeota is diverse, widespread and abundant in marine sediments and has the capacity to degrade detrital proteins. Environ. Microbiol. Rep. 2017, 9, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Castelle, C.J.; Hug, L.A.; Wrighton, K.C.; Thomas, B.C.; Williams, K.H.; Wu, D.; Tringe, S.G.; Singer, S.W.; Eisen, J.A.; Banfield, J.F. Extraordinary phylogenetic diversity and metabolic versatility in aquifer sediment. Nat. Commun. 2013, 4, 2120. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Kennedy, D.; Fredrickson, J.; Bjornstad, B.; Konopka, A. Vertical stratification of subsurface microbial community composition across geological formations at the Hanford Site. Environ. Microbiol. 2012, 14, 414–425. [Google Scholar] [CrossRef]
- Goh, K.M.; Shahar, S.; Chan, K.-G.; Chong, C.S.; Amran, S.I.; Sani, M.H.; Zakaria, I.I.; Kahar, U.M. Current status and potential applications of underexplored prokaryotes. Microorganisms 2019, 7, 468. [Google Scholar] [CrossRef]
- Becraft, E.D.; Woyke, T.; Jarett, J.; Ivanova, N.; Godoy-Vitorino, F.; Poulton, N.; Brown, J.M.; Brown, J.; Lau, M.C.Y.; Onstott, T.; et al. Rokubacteria: Genomic giants among the uncultured bacterial phyla. Front. Microbiol. 2017, 8, 2264. [Google Scholar] [CrossRef]
- Crits-Christoph, A.; Diamond, S.; Butterfield, C.N.; Thomas, B.C.; Banfield, J.F. Novel soil bacteria possess diverse genes for secondary metabolite biosynthesis. Nature 2018, 558, 440–444. [Google Scholar] [CrossRef]
- Feng, H.; Guo, J.; Wang, W.; Song, X.; Yu, S. Soil depth determines the composition and diversity of bacterial and archaeal communities in a poplar plantation. Forests 2019, 10, 550. [Google Scholar] [CrossRef]
- Steger, K.; Kim, A.T.; Ganzert, L.; Grossart, H.-P.; Smart, D.R. Floodplain soil and its bacterial composition are strongly affected by depth. FEMS Microbiol. Ecol. 2019, 95, fiz014. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, C.J.; Schönher, C.; Proksch, P.; Kerschbaumer, D.J.; Mayr, E.; Zunabovic-Pichler, M.; Domig, K.J.; Perfler, R. Assessment of microbial community dynamics in river bank filtrate using high-throughput sequencing and flow cytometry. Front. Microbiol. 2018, 9, 2887. [Google Scholar] [CrossRef] [PubMed]
- Gayner, N.J. River Bank Inducement Influence on a Shallow Groundwater Microbial Community and Its Effects on Aquifer Reactivity. Master’s Thesis, University of Wisconsin-Milwaukee, Milwaukee, WI, USA, 2018. [Google Scholar]
- Jungbluth, S.P.; Amend, J.P.; Rappé, M.S. Metagenome sequencing and 98 microbial genomes from Juan de Fuca Ridge flank subsurface fluids. Sci. Data 2017, 4, 170037. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Zhang, H.; Li, X.; Zhang, Y.; Su, Z.; Zhang, C. Soil fungistasis and its relations to soil microbial composition and diversity: A case study of a series of soils with different fungistasis. J. Environ. Sci. 2008, 20, 871–877. [Google Scholar] [CrossRef]
- Lee, C.G.; Iida, T.; Inoue, Y.; Muramoto, Y.; Watanabe, H.; Nakaho, K.; Ohkuma, M. Prokaryotic communities at different depths between soils with and without tomato bacterial wilt but pathogen-present in a single greenhouse. Microbes Environ. 2017, 32, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.G.; Iida, T.; Uwagaki, Y.; Otani, Y.; Nakaho, K.; Ohkuma, M. Comparison of prokaryotic and eukaryotic communities in soil samples with and without tomato bacterial wilt collected from different fields. Microbes Environ. 2017, 32, 376–385. [Google Scholar] [CrossRef]
- He, Y.; Wu, Z.; Wang, W.; Liu, X.; Ye, B.-C. Bacterial community and phosphorus species changes in pepper rhizosphere soils after Pseudomonas putida Rs-198 inoculation. Rhizosphere 2019, 11, 100164. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, M.; Yang, W.; Di, H.J.; Ma, L.; Liu, W.; Li, B. Effects of microbial inoculants on phosphorus and potassium availability, bacterial community composition, and chili pepper growth in a calcareous soil: A greenhouse study. J. Soils Sediments 2019, 19, 3597–3607. [Google Scholar] [CrossRef]
- Niu, J.; Rang, Z.; Zhang, C.; Chen, W.; Tian, F.; Yin, H.; Dai, L. The succession pattern of soil microbial communities and its relationship with tobacco bacterial wilt. BMC Microbiol. 2016, 16, 233. [Google Scholar] [CrossRef]
- Gao, L.; Wang, R.; Gao, J.; Li, F.; Huang, G.; Huo, G.; Liu, Z.; Tang, W.; Shen, G. Analysis of the structure of bacterial and fungal communities in disease suppressive and disease conducive tobacco-planting soils in China. Soil Res. 2019, 58, 35–40. [Google Scholar] [CrossRef]
- Kopecky, J.; Samkova, Z.; Sarikhani, E.; Kyselková, M.; Omelka, M.; Kristufek, V.; Divis, J.; Grundmann, G.G.; Moënne-Loccoz, Y.; Sagova-Mareckova, M. Bacterial, archaeal and micro-eukaryotic communities characterize a disease-suppressive or conducive soil and a cultivar resistant or susceptible to common scab. Sci. Rep. 2019, 9, 14883. [Google Scholar] [CrossRef] [PubMed]
- Svenningsen, N.B.; Watts-Williams, S.J.; Joner, E.J.; Battini, F.; Efthymiou, A.; Cruz-Paredes, C.; Nybroe, O.; Jakobsen, I. Suppression of the activity of arbuscular mycorrhizal fungi by the soil microbiota. ISME J. 2018, 12, 1296–1307. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, A.L.; Nybroe, O.; Winding, A.; Ekelund, F.; Bjørnlund, L. Bacterial feeders, the nematode Caenorhabditis elegans and the flagellate Cercomonas longicauda, have different effects on outcome of competition among the Pseudomonas biocontrol strains CHA0 and DSS73. Microb. Ecol. 2009, 57, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Rønn, R.; McCaig, A.E.; Griffiths, B.S.; Prosser, J.I. Impact of protozoan grazing on bacterial community structure in soil microcosms. Appl. Environ. Microbiol. 2002, 68, 6094. [Google Scholar] [CrossRef]
- Pedersen, A.L.; Ekelund, F.; Johansen, A.; Winding, A. Interaction of bacteria-feeding soil flagellates and Pseudomonas spp. Biol. Fert. Soils 2010, 46, 151–158. [Google Scholar] [CrossRef]
- Pedersen, A.L.; Winding, A.; Altenburger, A.; Ekelund, F. Protozoan growth rates on secondary-metabolite-producing Pseudomonas spp. correlate with high-level protozoan taxonomy. FEMS Microbiol. Lett. 2011, 316, 16–22. [Google Scholar] [CrossRef]
- Müller, M.S.; Scheu, S.; Jousset, A. Protozoa drive the dynamics of culturable biocontrol bacterial communities. PLoS ONE 2013, 8, e66200. [Google Scholar] [CrossRef]
- Buyer, J.S.; Roberts, D.P.; Russek-Cohen, E. Microbial community structure and function in the spermosphere as affected by soil and seed type. Can. J. Microbiol. 1999, 45, 138–144. [Google Scholar] [CrossRef]
- Hirose, D.; Degawa, Y.; Inaba, S.; Tokumasu, S. The anamorphic genus Calcarisporiella is a new member of the Mucoromycotina. Mycoscience 2012, 53, 256–260. [Google Scholar] [CrossRef]
- Du, G.; Qu, L.; Shang, K.; Sun, C.; Wang, C.; Gao, P. Ciliate Uronema marinum is the causative agent of scuticociliatosis in farm raised turbot Scophthalmus maximus. J. Oceanol. Limnol. 2019, 37, 1726–1735. [Google Scholar] [CrossRef]
- Ayo, B.; Latatu, A.; Artolozaga, I.; JÜRgens, K.; Iriberri, J. Factors affecting preference responses of the freshwater ciliate Uronema nigricans to bacterial prey. J. Eukaryot. Microbiol. 2009, 56, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Ekelund, F. Tolerance of soil flagellates to increased NaCl levels. J. Eukaryot. Microbiol. 2002, 49, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, R.D.; Preslan, J.E. Cultural characteristics of a pelagic marine hymenostome ciliate, Uronema sp. J. Exp. Mar. Biol. Ecol. 1969, 4, 90–99. [Google Scholar] [CrossRef]
- Hood, A.S.C.; Aryawan, A.A.K.; Advento, A.D.; Purnomo, D.; Wahyuningsih, R.; Luke, S.H.; Ps, S.; Snaddon, J.L.; Foster, W.A.; Caliman, J.-P.; et al. Understory vegetation in oil palm plantations promotes leopard cat activity, but does not affect rats or rat damage. Front. For. Glob. Chang. 2019, 2, 51. [Google Scholar] [CrossRef]
- Azhar, B.; Lindenmayer, D.B.; Wood, J.; Fischer, J.; Zakaria, M. Ecological impacts of oil palm agriculture on forest mammals in plantation estates and smallholdings. Biodivers. Conserv. 2014, 23, 1175–1191. [Google Scholar] [CrossRef]
- Lavelle, P.; Chauvel, A.; Fragoso, C. Faunal activity in acid soils. In Plant-Soil Interactions at Low pH: Principles and Management: Proceedings of the Third International Symposium on Plant-Soil Interactions at Low pH, Brisbane, Queensland, Australia, 12–16 September 1993; Date, R.A., Grundon, N.J., Rayment, G.E., Probert, M.E., Eds.; Springer: Dordrecht, The Netherlands, 1995; pp. 201–211. [Google Scholar]
- Ashton-Butt, A.; Aryawan, A.A.K.; Hood, A.S.C.; Naim, M.; Purnomo, D.; Suhardi; Wahyuningsih, R.; Willcock, S.; Poppy, G.M.; Caliman, J.-P.; et al. Understory vegetation in oil palm plantations benefits soil biodiversity and decomposition rates. Front. For. Glob. Change 2018, 1, 10. [Google Scholar] [CrossRef]
- Gorea, E.A.; Godwin, I.D.; Mudge, A.M. Ganoderma infection of oil palm—A persistent problem in Papua New Guinea and Solomon Islands. Australas. Plant Pathol. 2019, 49, 1–9. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease Class § | Scoring | Blenheim (Typic Quartzipsamment) | Bernam (Typic Endoaquept) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 2018 | 2019 | 2018 | 2019 | ||||||
| May | Dec | May | Dec | May | Dec | May | Dec | ||
| Healthy (%) | 0 | 98.7 | 98.7 | 98.7 | 98.1 | 75.2 | 73.5 | 72.4 | 66.9 |
| FB (%) | 1 | 0 | 0 | 0 | 0 | 10.5 | 11.6 | 8.8 | 10.5 |
| Rot (%) | 2 | 0 | 0 | 0 | 0.6 | 1.1 | 1.1 | 1.7 | 1.1 |
| Rot + FB (%) | 3 | 0 | 0 | 0 | 0 | 5.5 | 5.5 | 7.7 | 11 |
| Vacant + Sanitation (%) | 4 | 1.3 | 1.3 | 1.3 | 1.3 | 7.7 | 8.3 | 9.4 | 10.5 |
| Total Palm Censused | 158 | 158 | 158 | 158 | 181 | 181 | 181 | 181 | |
| Disease Incidence (%) | 1.3 | 1.3 | 1.3 | 1.9 | 24.9 | 26.5 | 27.6 | 33.1 | |
| Disease Severity Index | 1.27 | 1.27 | 1.27 | 1.58 | 15.06 | 15.88 | 18.23 | 21.96 | |
| Year | Census Time Points | Soil Types | Status of the Palms | p-Value after Chi-Square Test | ||
|---|---|---|---|---|---|---|
| No. of Infected Palms | No. of Healthy Palms | Total Assessed Palms | ||||
| 2018 | May | Blenheim | 2 | 156 | 158 | <0.0001 |
| Bernam | 45 | 136 | 181 | |||
| December | Blenheim | 2 | 156 | 158 | <0.0001 | |
| Bernam | 48 | 134 | 181 | |||
| 2019 | May | Blenheim | 2 | 156 | 158 | <0.0001 |
| Bernam | 50 | 132 | 181 | |||
| December | Blenheim | 3 | 155 | 158 | <0.0001 | |
| Bernam | 60 | 122 | 181 | |||
| Chemical Parameters § | Blenheim (Typic Quartzipsamment) | Bernam (Typic Endoaquepts) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| May 2018 | Dec 2018 | May 2018 | Dec 2018 | |||||||||||||
| PCT * | PCS * | IPT * | IPS * | PCT | PCS | IPT | IPS | PCT | PCS | IPT | IPS | PCT | PCS | IPT | IPS | |
| pH | 7.65 (0.05) | 7.67 (0.06) | 7.73 (0.07) | 7.66 (0.06) | 7.90 (0.08) | 8.05 (0.10) | 7.90 (0.08) | 7.96 (0.08) | 5.66 (0.42) | 5.83 (0.41) | 6.23 (0.23) | 5.99 (0.35) | 5.53 (0.48) | 5.73 (0.37) | 5.71 (0.43) | 5.44 (0.50) |
| OM (%) | 5.36 (0.57) | 5.53 (0.38) | 5.08 (0.66) | 5.29 (0.52) | 5.99 (0.85) | 4.70 (0.47) | 5.07 (0.70) | 4.36 (0.48) | 11.01 (0.26) | 11.44 (0.10) | 10.79 (0.28) | 11.17 (0.16) | 11.83 (0.28) | 11.61 (0.52 | 10.80 (0.33) | 10.69 (0.19) |
| N (%) | 0.58 (0.05) | 0.61 (0.07) | 0.49 (0.04) | 0.53 (0.07) | 0.44 (0.03) | 0.35 (0.04) | 0.50 (0.03) | 0.41 (0.02) | 0.17 (0.01) | 0.18 (0.01) | 0.15 (0.01) | 0.14 (0.01) | 0.22 (0.01) | 0.17 (0.02) | 0.16 (0.01) | 0.20 (0.01) |
| Fe (%) | 0.41 (0.08) | 0.41 (0.09) | 0.36 (0.09) | 0.44 (0.12) | 0.27 (0.07) | 0.24 (0.07) | 0.30 (0.07) | 0.29 (0.08) | 1.26 (0.04) | 1.27 (0.04) | 1.15 (0.08) | 1.24 (0.05) | 1.35 (0.03) | 1.23 (0.05) | 1.23 (0.05) | 1.25 (0.05) |
| Si (%) | 0.75 (0.03) | 0.69 (0.02) | 0.75 (0.05) | 0.81 (0.05) | 0.72 (0.24) | 0.60 (0.17) | 0.93 (0.31) | 0.89 (0.39) | 4.61 (0.11) | 4.88 (0.25) | 4.65 (0.10) | 4.57 (0.18) | 4.53 (0.09) | 4.43 (0.10) | 4.47 (0.07) | 4.63 (0.08) |
| Ca (%) | 40.24 (2.76) | 38.69 (2.60) | 41.04 (2.83) | 37.46 (1.74) | 1.07 (0.31) | 0.65 (0.13) | 0.68 (0.15) | 0.59 (0.09) | ||||||||
| TP (mg/kg) | 1080.70 (180.49) | 933.56 (159.13) | 553.44 (65.13) | 583.44 (153.84) | 741.56 (144.85) | 457.56 (64.89) | 698.33 (115.76) | 540.56 (86.01) | 340.56 (25.78) | 414.36 (29.74) | 329.22 (34.53) | 275.44 (30.82) | 960 (232.79) | 428.33 (40.26) | 381.67 (41.53) | 443.11 (40.21) |
| AP (mg/kg) | 12.4 (6.20) | 7.12 (3.12) | 10.1 (4.79) | 5.42 (2.65) | 20.13 (3.66) | 10.42 (2.58) | 19.84 (5.13) | 15.56 (5.01) | 62.98 (11.15) | 81.54 (12.33) | 75.52 (21.82) | 43.01 (9.56) | 158.22 (15.59) | 85.87 (16.78) | 54.94 (9.80) | 67.93 (10.65) |
| Mn (mg/kg) | 140.44 (9.02) | 148.89 (10.84) | 143.44 (9.84) | 144.22 (11.72) | 145.33 (11.74) | 119.13 (9.48) | 161.22 (13.04) | 154.78 (16.80) | 237.11 (20.54) | 276.64 (16.56) | 231.11 (24.81) | 203.44 (17.94) | 199.67 (12.09) | 181.78 (11.77) | 225.11 (32.37) | 231.22 (14.54) |
| Zn (mg/kg) | 19.46 (2.47) | 21.70 (3.34) | 17.32 (1.90) | 19.49 (3.50) | 21.18 (2.03) | 18.16 (1.64) | 24.58 (2.88) | 23.19 (3.68) | 43.48 (0.93) | 47.35 (2.10) | 43.36 (2.48) | 42.72 (0.88) | 54.32 (2.01) | 55.32 (6.39) | 47.33 (1.67) | 48.08 (0.85) |
| Cu (mg/kg) | 12.52 (0.65) | 12.52 (0.64) | 12.52 (0.65) | 12.23 (0.64) | 11.80 (0.52) | 10.55 (0.56) | 11.84 (0.79) | 10.88 (0.55) | 7.83 (0.32) | 8.88 (0.44) | 8.19 (0.41) | 7.20 (0.58) | 9.73 (0.36) | 9.32 (0.67) | 10.29 (0.34) | 10.12 (0.13) |
| Exchangeable K (cmol(+)/kg) | 0.16 (0.05) | 0.12 (0.04) | 0.17 (0.05) | 0.13 (0.04) | 0.20 (0.05) | 0.15 (0.02) | 0.22 (0.06) | 0.18 (0.05) | 1.68 (0.15) | 1.71 (0.20) | 1.04 (0.11) | 1.33 (0.18) | 2.55 (0.28) | 2.17 (0.32) | 1.17 (0.15) | 1.70 (0.20) |
| Exchangeable Mg (cmol(+)/kg) | 0.23 (0.04) | 0.18 (0.03) | 0.21 (0.03) | 0.19 (0.04) | 0.44 (0.08) | 0.33 (0.05) | 0.19 (0.02) | 0.17 (0.02) | 8.62 (0.73) | 7.91 (0.99) | 9.40 (0.90) | 10.27 (0.72) | 7.79 (0.67) | 9.87 (0.62) | 8.23 (1.04) | 6.63 (1.07) |
| Exchangeable Na (cmol(+)/kg) | 0.15 (0.01) | 0.13 (0.01) | 0.144 (0.01) | 0.16 (0.02) | 0.15 (0.01) | 0.15 (0.01) | 0.13 (0.01) | 0.13 (0.01) | 1.45 (0.19) | 2.23 (0.26) | 0.86 (0.12) | 1.26 (0.15) | 1.16 (0.17) | 2.24 (0.32) | 0.68 (0.12) | 1.14 (0.21) |
| CEC (cmol(+)/kg) | 6.94 (1.25) | 7.29 (0.99) | 7.57 (1.25) | 7.58 (1.33) | 8.12 (1.53) | 6.62 (1.14) | 8.28 (1.26) | 7.32 (1.31) | 29.64 (0.85) | 28.75 (1.05) | 29.49 (1.04) | 30.48 (0.94) | 26.40 (0.89) | 24.61 (0.98) | 25.03 (0.69) | 26.88 (0.82) |
| EC (μS/cm) | 128.67 (11.13) | 146.11 (10.02) | 129.00 (10.70) | 136.11 (10.64) | 157.44 (26.63) | 145.02 (23.83) | 137.88 (12.62) | 142.94 (21.15) | 728.22 (77.26) | 985.82 (126.43) | 502.89 (62.49) | 613.33 (97.83) | 596.78 (61.07) | 861.00 (116.47) | 486.56 (40.65) | 533.67 (61.37) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goh, Y.K.; Zoqratt, M.Z.H.M.; Goh, Y.K.; Ayub, Q.; Ting, A.S.Y. Determining Soil Microbial Communities and Their Influence on Ganoderma Disease Incidences in Oil Palm (Elaeis guineensis) via High-Throughput Sequencing. Biology 2020, 9, 424. https://doi.org/10.3390/biology9120424
Goh YK, Zoqratt MZHM, Goh YK, Ayub Q, Ting ASY. Determining Soil Microbial Communities and Their Influence on Ganoderma Disease Incidences in Oil Palm (Elaeis guineensis) via High-Throughput Sequencing. Biology. 2020; 9(12):424. https://doi.org/10.3390/biology9120424
Chicago/Turabian StyleGoh, Yit Kheng, Muhammad Zarul Hanifah Md Zoqratt, You Keng Goh, Qasim Ayub, and Adeline Su Yien Ting. 2020. "Determining Soil Microbial Communities and Their Influence on Ganoderma Disease Incidences in Oil Palm (Elaeis guineensis) via High-Throughput Sequencing" Biology 9, no. 12: 424. https://doi.org/10.3390/biology9120424
APA StyleGoh, Y. K., Zoqratt, M. Z. H. M., Goh, Y. K., Ayub, Q., & Ting, A. S. Y. (2020). Determining Soil Microbial Communities and Their Influence on Ganoderma Disease Incidences in Oil Palm (Elaeis guineensis) via High-Throughput Sequencing. Biology, 9(12), 424. https://doi.org/10.3390/biology9120424