Bi-Pinnate Compound Serianthes nelsonii Leaf-Level Plasticity Magnifies Leaflet-Level Plasticity



Simple Summary
Abstract
1. Introduction
2. Materials and Methods
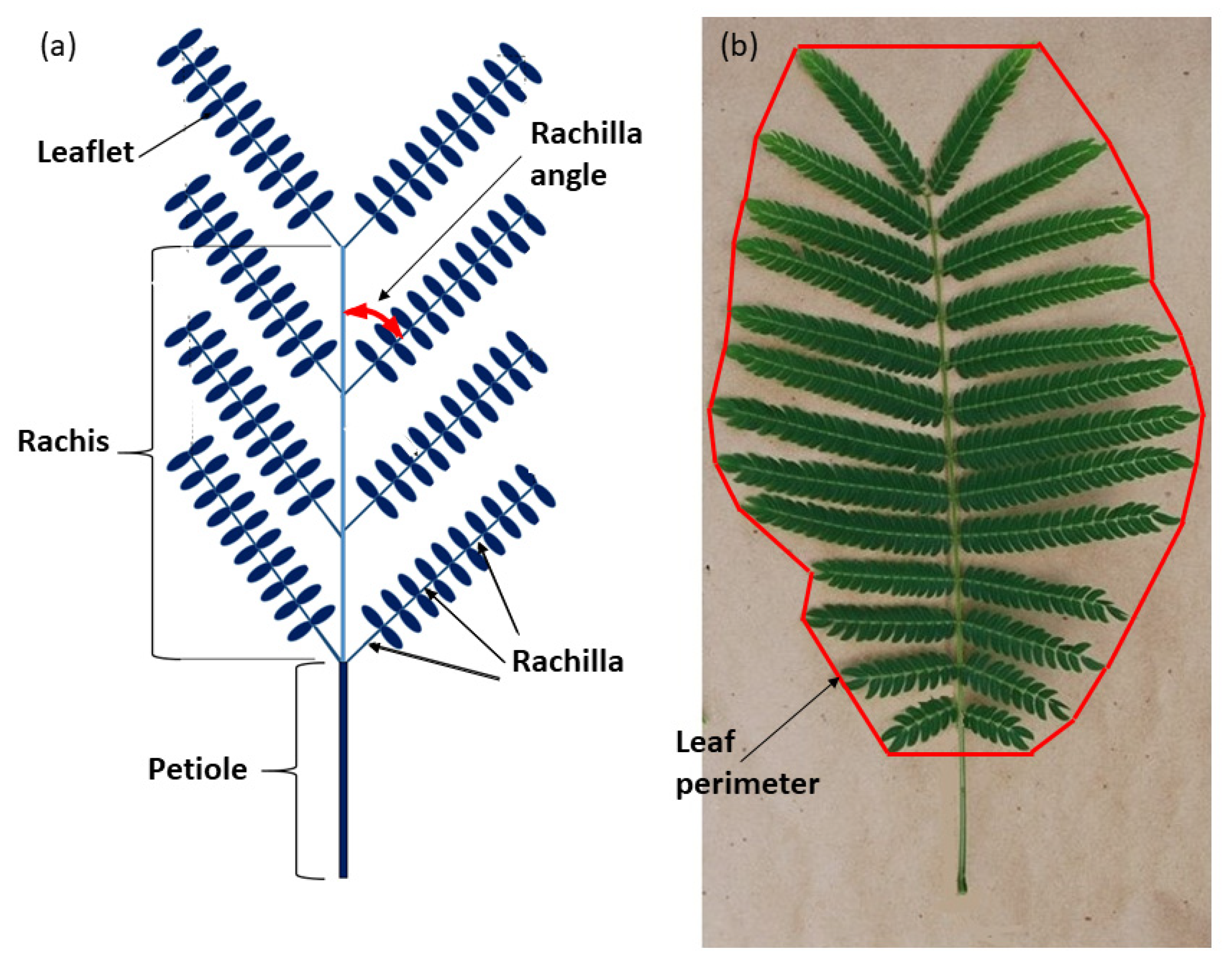
2.1. Leaf Measurements
2.2. Photographic and Derived Analyses
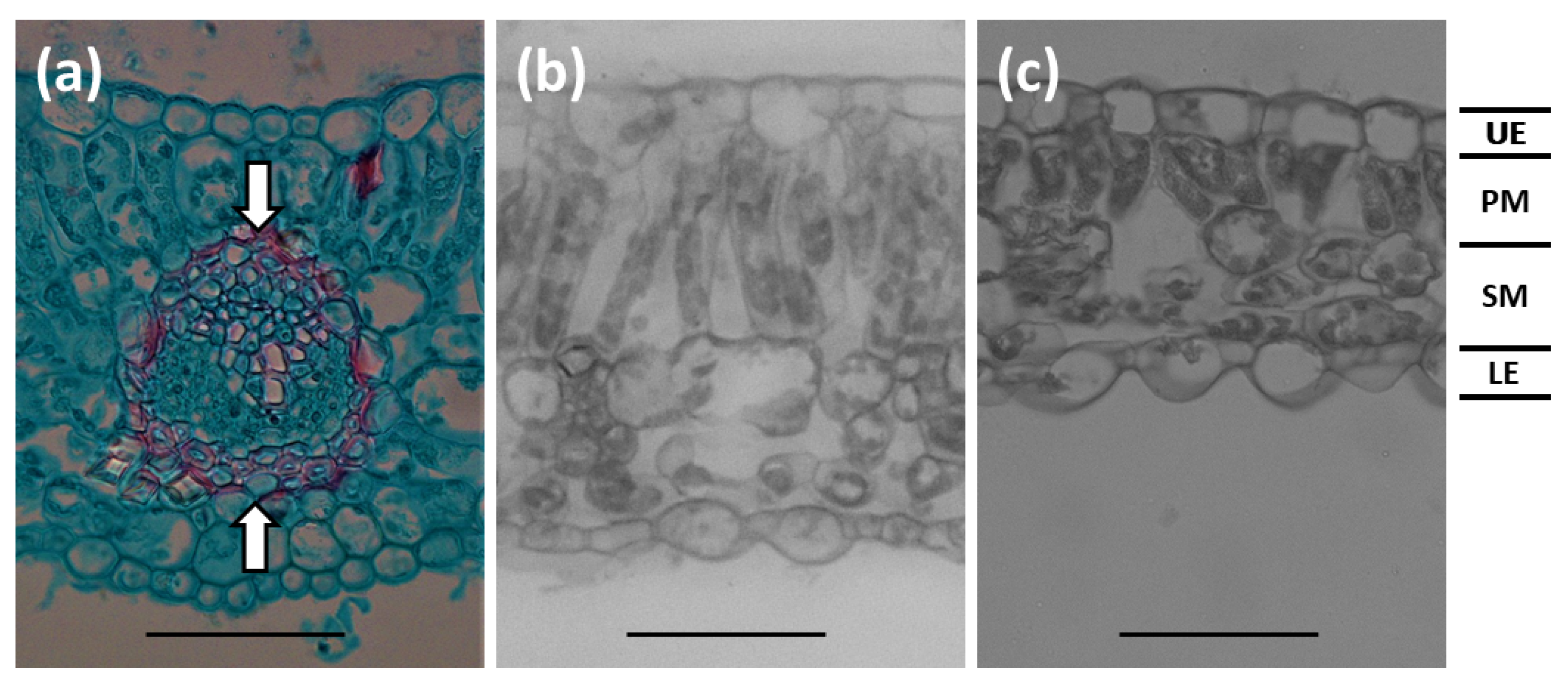
2.3. Microscopy
2.4. Statistical Analyses
3. Results
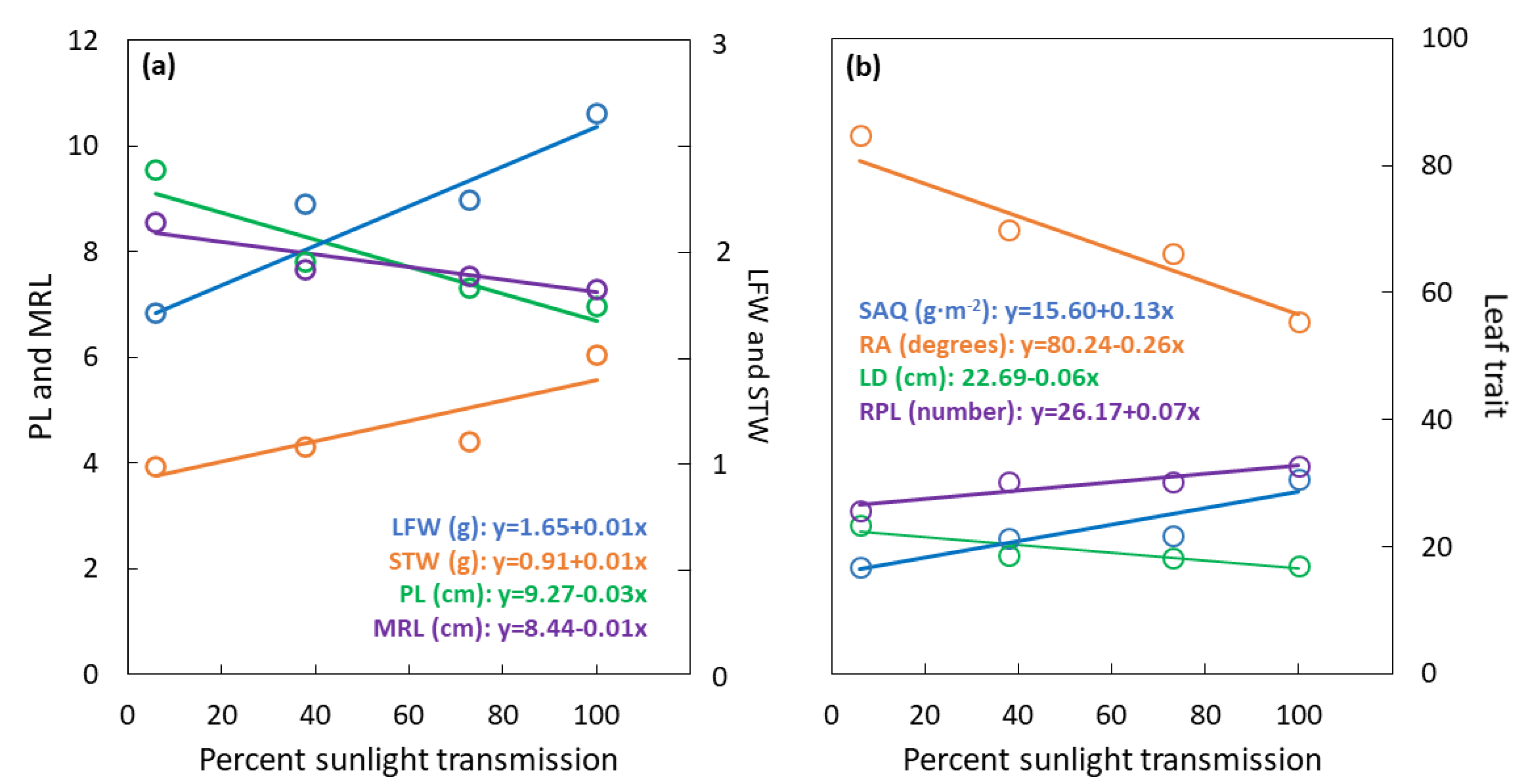
3.1. Whole Leaf Traits
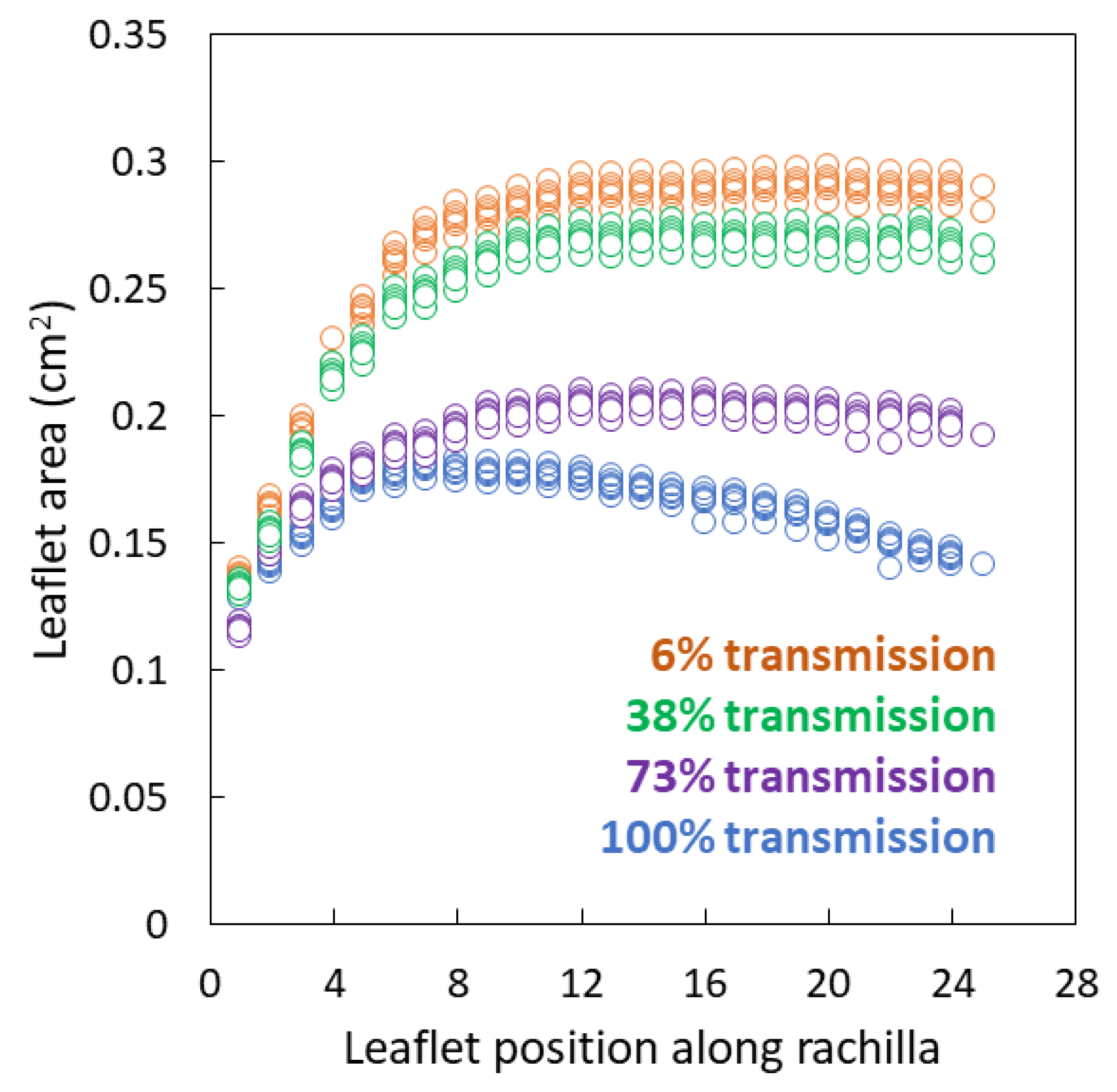
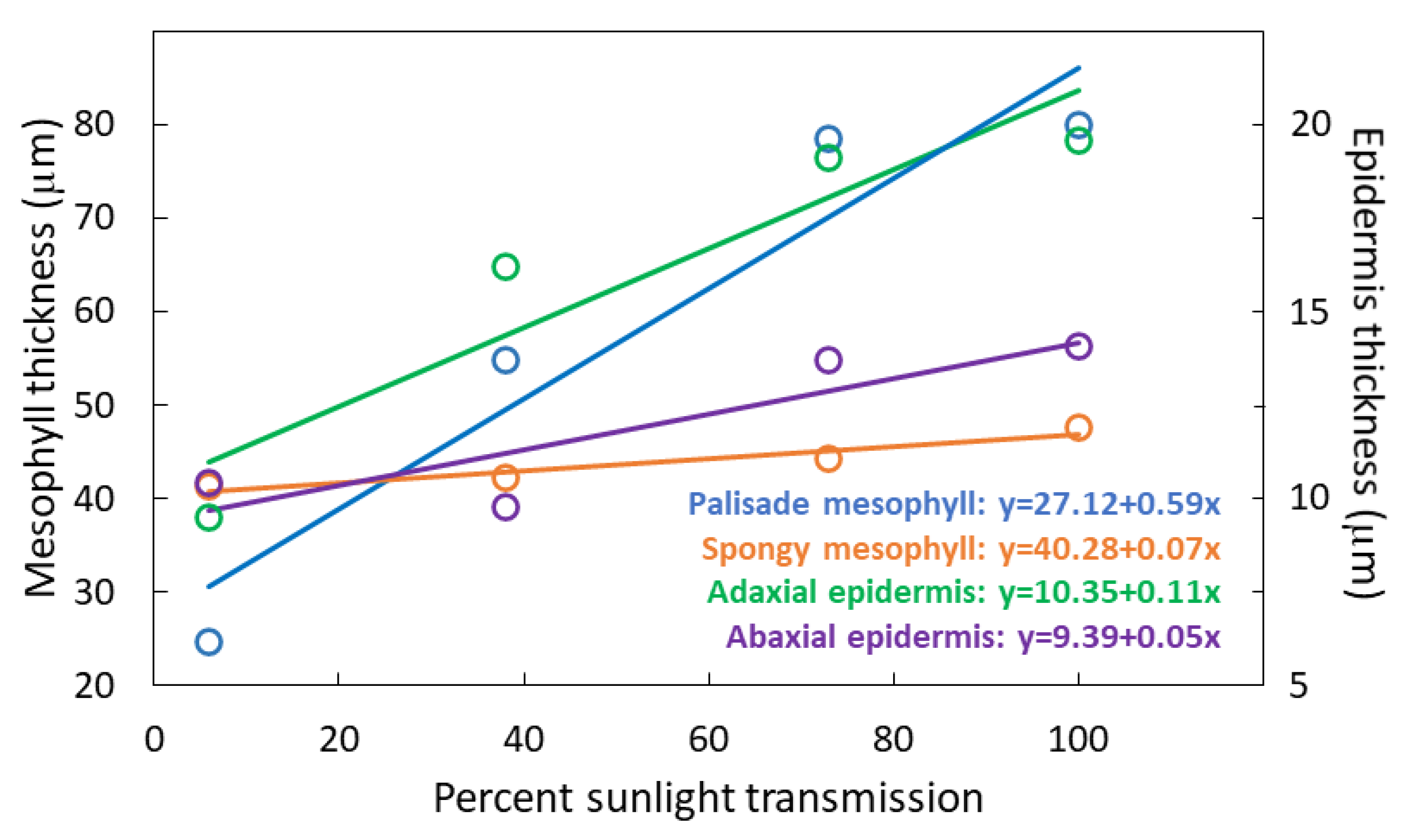
3.2. Leaflet Traits
3.3. Plasticity
4. Discussion
4.1. Leaflet Versus Whole Leaf
4.2. Benefits of Acclimation
4.3. Conservation Biology
4.4. Future Directions
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Boardman, N.K. Comparative photosynthesis of sun and shade plants. Annu. Rev. Plant Physiol. 1977, 28, 355–377. [Google Scholar] [CrossRef]
- Björkman, O. Responses to different quantum flux densities. In Physiological Plant Ecology. I. Responses to the Physical Environment, Encycl. Plant Physiol. New Ser.; Lange, O.L., Nobel, P.S., Osmond, C.B., Ziegler, H., Eds.; Springer: New York, NY, USA, 1981; Volume 12A, pp. 57–107. [Google Scholar]
- Givnish, T.J. Adaptation to sun and shade: A whole-plant perspective. Aust. J. Plant Physiol. 1988, 15, 63–92. [Google Scholar] [CrossRef]
- Sims, D.A.; Pearcy, R.W. Scaling sun and shade photosynthetic acclimation of Alocasia macrorrhiza to whole plant performance. I. Carbon balance and allocation at different daily photon flux densities. Plant Cell Environ. 1994, 17, 881–887. [Google Scholar] [CrossRef]
- Evans, J.R.; Poorter, H. Photosynthetic acclimation of plants to growth irradiance: The relative importance of specific leaf area and nitrogen partitioning in maximizing carbon gain. Plant Cell Environ. 2001, 24, 755–767. [Google Scholar] [CrossRef]
- Quero, J.L.; Villar, R.; Maranon, T.; Zamora, R. Interactions of drought and shade effects on seedlings of four Quercus species: Physiological and structural leaf responses. New Phytol. 2006, 170, 819–834. [Google Scholar] [CrossRef]
- Walters, R.G. Towards an understanding of photosynthetic acclimation. J. Expt. Bot. 2005, 56, 435–447. [Google Scholar] [CrossRef]
- Gates, D.M. Biophysical Ecology; Springer: New York, NY, USA, 1980. [Google Scholar]
- Bongers, F.; Popma, J. Is exposure-related variation in leaf characteristics of tropical rain forest species adaptive? In Plant Form and Vegetation Structure; Werger, M.J.A., van der Aart, P.J.M., During, H.J., Verhoeven, J.T.A., Eds.; Academic Publishing: The Hague, The Netherlands, 1988; pp. 191–200. [Google Scholar]
- Parkhurst, D.F.; Loucks, O.L. Optimal leaf size in relation to environment. J. Ecol. 1972, 60, 505–537. [Google Scholar] [CrossRef]
- Zimmermann, M.H. Xylem Structure and the Ascent of Sap; Springer: Berlin, Germany, 1983. [Google Scholar]
- Ort, D.R. When there is too much light. Plant Physiol. 2001, 125, 29–32. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams, W.W., III. Photoprotection in an ecological context: The remarkable complexity of thermal energy dissipation. New Phytol. 2006, 172, 11–21. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Kull, K. Leaf weight per area and leaf size of 85 Estonian woody species in relation to shade tolerance and light availability. For. Ecol. Manag. 1994, 70, 1–10. [Google Scholar] [CrossRef]
- Rozendaal, D.; Hurtado, V.; Poorter, L. Plasticity in leaf traits of 38 tropical tree species in response to light; relationships with light demand and adult stature. Funct. Ecol. 2006, 20, 207–216. [Google Scholar] [CrossRef]
- Markesteijn, L.; Poorter, L.; Bongers, F. Light-dependent leaf trait variation in 43 tropical dry forest tree species. Amer. J. Bot. 2007, 94, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Fisher, J.B. Sun and shade effects on the leaf of Guarea (Meliaceae): Plasticity of a branch analogue. Bot. Baz. 1986, 147, 84–89. [Google Scholar] [CrossRef]
- Niinemets, Ü. Are compound-leaved woody species inherently shade-intolerant? An analysis of species ecological requirements and foliar support costs. Plant Ecol. 1998, 134, 1–11. [Google Scholar] [CrossRef]
- Xu, F.; Guo, W.; Xu, W.; Wei, Y.; Wang, R. Leaf morphology correlates with water and light availability: What consequences for simple and compound leaves? Prog. Nat. Sci. 2009, 19, 1789–1798. [Google Scholar] [CrossRef]
- Anderson, J.M.; Osmond, C.B. Sun-shade responses: Compromises between acclimation and photoinhibition. In Photoinhibition; Kyle, D.J., Osmond, C.B., Arntzen, C.J., Eds.; Elsevier: Amsterdam, The Netherlands, 1987; pp. 1–38. [Google Scholar]
- Bazzaz, F.A. The physiological ecology of plant succession. Annu. Rev. Ecol. Syst. 1979, 10, 351–371. [Google Scholar] [CrossRef]
- Strauss-Debenedetti, S.; Bazzaz, F.A. Plasticity and acclimation to light in tropical Moraceae of different successional positions. Oecologia 1991, 87, 377–387. [Google Scholar] [CrossRef]
- Valladares, F.; Wright, S.J.; Lasso, E.; Kitajima, K.; Pearcy, R.W. Plastic phenotypic response to light of 16 congeneric shrubs from Panamanian rainforest. Ecology 2000, 8, 1925–1936. [Google Scholar] [CrossRef]
- Murchie, E.H.; Horton, P. Acclimation of photosynthesis to irradiance and spectral quality in British plant species: Chlorophyll content, photosynthetic capacity and habitat preference. Plant Cell Environ. 1997, 20, 438–448. [Google Scholar] [CrossRef]
- Popma, J.; Bongers, F.; Werger, M.J.A. Gap-dependence and leaf characteristics of trees in a tropical lowland rain forest in Mexico. Oikos 1992, 63, 207–214. [Google Scholar] [CrossRef]
- Thomas, S.C.; Bazzaz, F.A. Asymptotic height as a predictor of photosynthetic characteristics in Malaysian rain forest trees. Ecology 1999, 80, 1607–1622. [Google Scholar] [CrossRef]
- Poorter, L.; Bongers, F.; Sterck, F.J.; Wöll, H. Beyond the regeneration phase: Differentiation of height–light trajectories among tropical tree species. J. Ecol. 2005, 93, 256–267. [Google Scholar] [CrossRef]
- Marler, T.E.; Cascasan, A.N. Number of emerged seedlings and seedling longevity of the non-recruiting, critically endangered Håyun lågu tree Serianthes nelsonii Merr. (Fabales: Leguminosae) are influenced by month of emergence. J. Threat. Taxa 2015, 7, 8221–8225. [Google Scholar] [CrossRef]
- Marler, T.E.; Cruz, G.N. Extreme wind events influence seed rain and seedling dynamics of Guam’s Serianthes nelsonii Merr. Trop. Conserv. Sci. 2019, 12, 1–6. [Google Scholar] [CrossRef]
- Marler, T.; Musser, C. Potential stressors leading to seedling mortality in the endemic Håyun lågu tree (Serianthes nelsonii Merr.) in the island of Guam. Trop. Conserv. Sci 2015, 8, 738–744. [Google Scholar] [CrossRef]
- Ng, M.; Smith, S.Y. Evaluating stasis in Metasequoia (Cupressaceae): Testing the relationship between leaf traits and climate. Int. J. Plant Sci. 2020, 181, 157–174. [Google Scholar] [CrossRef]
- Wiles, G.; Williams, E. Serianthes nelsonii. The IUCN Red List of Threatened Species 2017. Available online: https://www.iucnredlist.org/species/30437/98715973 (accessed on 15 September 2020).
- Marler, T.E.; Cascasan, A.N.; Lawrence, J.H. Threatened native trees in Guam: Short-term seed storage and shade conditions influence emergence and growth of seedlings. HortScience 2015, 50, 1049–1054. [Google Scholar] [CrossRef]
- Marler, T.E. Thigmomorphogenesis and biomechanical responses of shade-grown Serianthes nelsonii plants to stem flexure. Plant Signal. Behav. 2019, 14, e1601953. [Google Scholar] [CrossRef]
- Marler, T.E. Repetitive pruning of Serianthes nursery plants improves transplant quality and post-transplant survival. Plant Signal. Behav. 2019, 14, e1621246. [Google Scholar] [CrossRef]
- National Institutes of Health. ImageJ Image Processing and Analysis in Java. Available online: https://imagej.nih.gov (accessed on 15 September 2020).
- Rice, S.A.; Bazzaz, F.A. Growth consequences of plasticity of plant traits in response to light conditions. Oecologia 1989, 78, 508–512. [Google Scholar] [CrossRef]
- Poorter, H.; Niinemets, Ü.; Poorter, L.; Wright, I.J.; Villar, R. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis. New Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef] [PubMed]
- Asner, G.P.; Martin, R.E. Convergent elevation trends in canopy chemical traits of tropical forests. Glob. Chang. Biol. 2016, 22, 2216–2227. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.E.; Asner, G.P.; Bentley, L.P.; Shenkin, A.; Salinas, N.; Huaypar, K.Q.; Pillco, M.M.; Ccori Álvarez, F.D.; Enquist, B.J.; Diaz, S.; et al. Covariance of sun and shade leaf traits along a tropical forest elevation gradient. Front. Plant Sci. 2020, 10, 1810. [Google Scholar] [CrossRef] [PubMed]
- Hanson, H.C. Leaf-structure as related to environment. Amer. J. Bot. 1917, 4, 533–560. [Google Scholar] [CrossRef]
- Niinemets, Ü. Components of leaf dry mass per area–thickness and density–alter leaf photosynthetic capacity in reverse directions in woody plants. New Phytol. 1999, 144, 35–47. [Google Scholar] [CrossRef]
- Marler, T.E. Diurnal Serianthes nelsonii Merr. leaflet paraheliotropism reduces leaflet temperature, relieves photoinhibition, and alters nyctinastic behavior. J. Threat. Taxa 2019, 11, 14112–14118. [Google Scholar] [CrossRef]
- Bugmann, H. Functional types of trees in temperate and boreal forests: Classification and testing. J. Veg. Sci. 1996, 7, 359–370. [Google Scholar] [CrossRef]
- United States Fish and Wildlife Service. Recovery Plan for Serianthes Nelsonii; USFWS: Portland, OR, USA, 1994.
- Janzen, D.H. Herbivores and the number of tree species in tropical forests. Am. Nat. 1970, 104, 501–528. [Google Scholar] [CrossRef]
- Connell, J.H. On the role of natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees. In Dynamics of Populations; den Boer, P.J., Gradwell, G.R., Eds.; Centre for Agricultural Publishing and Documentation: Wageningen, The Netherlands, 1971; pp. 298–312. [Google Scholar]
- Ekroos, J.; Leventon, J.; Fischer, J.; Newig, J.; Smith, H.G. Embedding evidence on conservation interventions within a context of multilevel governance. Conserv. Lett. 2017, 10, 139–145. [Google Scholar] [CrossRef]
- Machinski, J.; Albrecht, M.A. Center for Plant Conservation’s best practice guidelines for the reintroduction of rare plants. Plant Divers. 2017, 39, 390–395. [Google Scholar] [CrossRef]
- Djenontin, I.N.S.; Meadow, A.M. The art of co-production of knowledge in environmental sciences and management: Lessons from international practice. Environ. Manag. 2018, 61, 885–903. [Google Scholar] [CrossRef]
- Efroymson, R.; Jager, H.; Dale, V.; Westervelt, J. A framework for developing management goas for species at risk with examples from military installations in the United States. Environ. Manag. 2009, 44, 1163–1179. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.C.; Ickes, K. Ontogenetic changes in leaf size in Malaysian rain forest trees. Biotropica 1995, 27, 427–434. [Google Scholar] [CrossRef]
- Martinez-Garza, C.; Howe, H.F. Developmental strategy or immediate responses in leaf traits of tropical tree species? Int. J. Plant Sci. 2005, 166, 41–48. [Google Scholar] [CrossRef]
- Teixeira, R.T. Distinct responses to light in plants. Plants 2020, 9, 894. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Trait | Units |
|---|---|---|
| LET | Abaxial epidermis thickness | µm |
| LFD | Leaflet diameter | mm |
| LFL | Leaflets per leaf | number |
| LFQ | WLA/PA | cm2·cm−2 |
| LFT | Leaflet thickness | µm |
| LFW | Total leaflet dry weight | g |
| LL | Leaflet length | mm |
| LMA | Leaflet mass per area | g·m−2 |
| LR | Leaflets per rachilla | Number |
| LTD | Leaflet tissue density | mg·cm−3 |
| MLA | Individual leaflet area | cm2 |
| MRL | Maximum rachilla length | cm |
| PA | Area within leaf perimeter | cm2 |
| PL | Petiole length | cm |
| PMT | Palisade mesophyll thickness | µm |
| PSQ | PMT/SMT | µm·µm−1 |
| RA | Rachilla insertion angle | ° |
| RL | Rachis length | cm |
| RPL | Rachilla per leaf | Number |
| SAQ | STW/WLA | g·m−2 |
| SMT | Spongy mesophyll thickness | µm |
| STW | Support tissue dry weight | g |
| SWQ | STW/TLW | g·g−1 |
| TLW | Total leaf weight | g |
| UET | Adaxial epidermis thickness | µm |
| VAQ | VT/MLA | µm·cm−2 |
| VT | Midvein thickness | µm |
| WLA | Total area of leaflets | cm2 |
| Leaf Trait 1 | 6% Transmission | 38% Transmission | 73% Transmission | 100% Transmission | Significance 2 |
|---|---|---|---|---|---|
| Total leaf wt (g) | 2.70 ± 0.18 | 3.14 ± 0.17 | 3.33 ± 0.18 | 4.16 ± 0.20 | <0.001; L *** |
| Leaflets/leaf | 1217 ± 36 | 1426 ± 35 | 1444 ± 39 | 1504 ± 46 | 0.003; L *** |
| Rachis length (cm) | 45.4 ± 1.7 | 46.2 ± 1.2 | 44.1 ± 1.7 | 45.0 ± 1.3 | 0.538 |
| LR | 47 ± 1 | 47 ± 1 | 45 ± 1 | 45 ± 1 | 0.610 |
| PA (cm2) | 768.0 ± 19.6 | 670.7 ± 15.2 | 649.3 ± 14.9 | 602.0 ± 14.6 | <0.001; L *** |
| WLA (cm2) | 293.5 ± 17.4 | 270.4 ± 19.0 | 254.6 ± 16.9 | 251.8 ± 14.1 | 0.086 |
| LFQ (cm2·cm−2) | 0.38 ± 0.03 | 0.40 ± 0.02 | 0.40 ± 0.03 | 0.42 ± 0.03 | 0.660 |
| SWQ (g·g−1) | 0.37 ± 0.01 | 0.35 ± 0.01 | 0.33 ± 0.01 | 0.36 ± 0.01 | 0.002; Q ** |
| Leaflet Trait 1 | 6% Transmission | 38% Transmission | 73% Transmission | 100% Transmission | p Value 2 |
|---|---|---|---|---|---|
| Area (cm2) | 0.24 ± 0.01 | 0.19 ± 0.01 | 0.18 ± 0.01 | 0.17 ± 0.01 | <0.001; L ***; Q * |
| Thickness (µm) | 85.94 ± 1.39 | 122.97 ± 1.48 | 155.50 ± 1.38 | 159.59 ± 1.63 | <0.001; L ***; Q *** |
| Diameter (mm) | 4.33 ± 0.21 | 3.83 ± 0.18 | 3.63 ± 0.14 | 3.13 ± 0.11 | <0.001; L *** |
| Length (mm) | 9.57 ± 0.30 | 8.23 ± 0.28 | 7.97 ± 0.18 | 7.52 ± 0.10 | <0.001; L ***; Q * |
| Vein thickness (µm) | 44.53 ± 1.85 | 63.33 ± 1.68 | 64.33 ± 1.82 | 69.58 ± 1.55 | <0.001; L ***; Q *** |
| LMA (g·m−2) | 29.12 ± 2.54 | 39.80 ± 3.42 | 44.65 ± 3.77 | 53.83 ± 3.33 | <0.001; L *** |
| LTD (mg·cm−3) | 338.88 ± 29.63 | 323.69 ± 27.85 | 287.17 ± 24.58 | 337.33 ± 20.85 | 0.411 |
| VAQ (µm·cm−2) | 187.76 ± 6.06 | 333.19 ± 5.49 | 368.65 ± 5.07 | 421.84 ± 23.07 | <0.001; L ***; Q *** |
| PSQ (µm·µm−1) | 0.60 ± 0.03 | 1.31 ± 0.05 | 1.80 ± 0.09 | 1.66 ± 0.08 | <0.001; L ***; Q *** |
| Leaf Traits | PI | Leaflet Traits | PI |
|---|---|---|---|
| Perimeter area | 0.22 | Individual Area | 0.29 |
| Total leaf diameter | 0.28 | Length | 0.21 |
| Petiole length | 0.26 | Diameter | 0.28 |
| Rachilla length | 0.15 | Lamina thickness | 0.46 |
| Rachilla number | 0.22 | Palisade thickness | 0.68 |
| Leaflets/leaf | 0.19 | Spongy thickness | 0.13 |
| Rachilla angle | 0.35 | Adaxial epidermis thickness | 0.52 |
| STW 1 | 0.35 | Abaxial epidermis thickness | 0.46 |
| LFW | 0.35 | Midvein thickness | 0.36 |
| TLW | 0.35 | LMA | 0.46 |
| SWQ | 0.10 | VAQ | 0.55 |
| SAQ | 0.45 | PSQ | 0.64 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deloso, B.E.; Marler, T.E. Bi-Pinnate Compound Serianthes nelsonii Leaf-Level Plasticity Magnifies Leaflet-Level Plasticity. Biology 2020, 9, 333. https://doi.org/10.3390/biology9100333
Deloso BE, Marler TE. Bi-Pinnate Compound Serianthes nelsonii Leaf-Level Plasticity Magnifies Leaflet-Level Plasticity. Biology. 2020; 9(10):333. https://doi.org/10.3390/biology9100333
Chicago/Turabian StyleDeloso, Benjamin E., and Thomas E. Marler. 2020. "Bi-Pinnate Compound Serianthes nelsonii Leaf-Level Plasticity Magnifies Leaflet-Level Plasticity" Biology 9, no. 10: 333. https://doi.org/10.3390/biology9100333
APA StyleDeloso, B. E., & Marler, T. E. (2020). Bi-Pinnate Compound Serianthes nelsonii Leaf-Level Plasticity Magnifies Leaflet-Level Plasticity. Biology, 9(10), 333. https://doi.org/10.3390/biology9100333