
C3H10T1/2 Mesenchymal Stem Cell Line as a New In Vitro Tool for Studying Adipocyte Dedifferentiation

 and
and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Adipogenesis
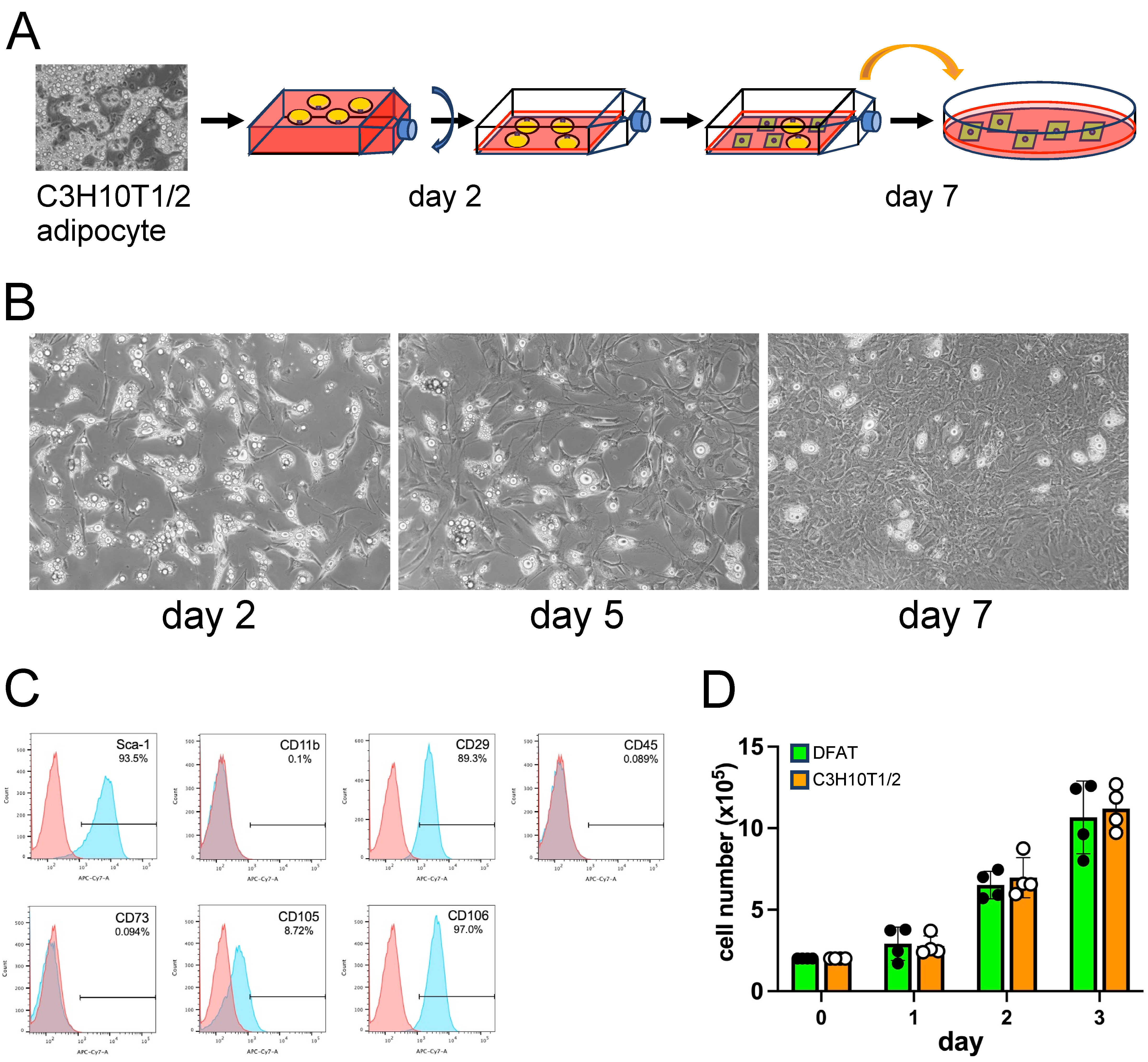
2.3. Ceiling Culture
2.4. Flow Cytometry
2.5. Oil Red O Staining
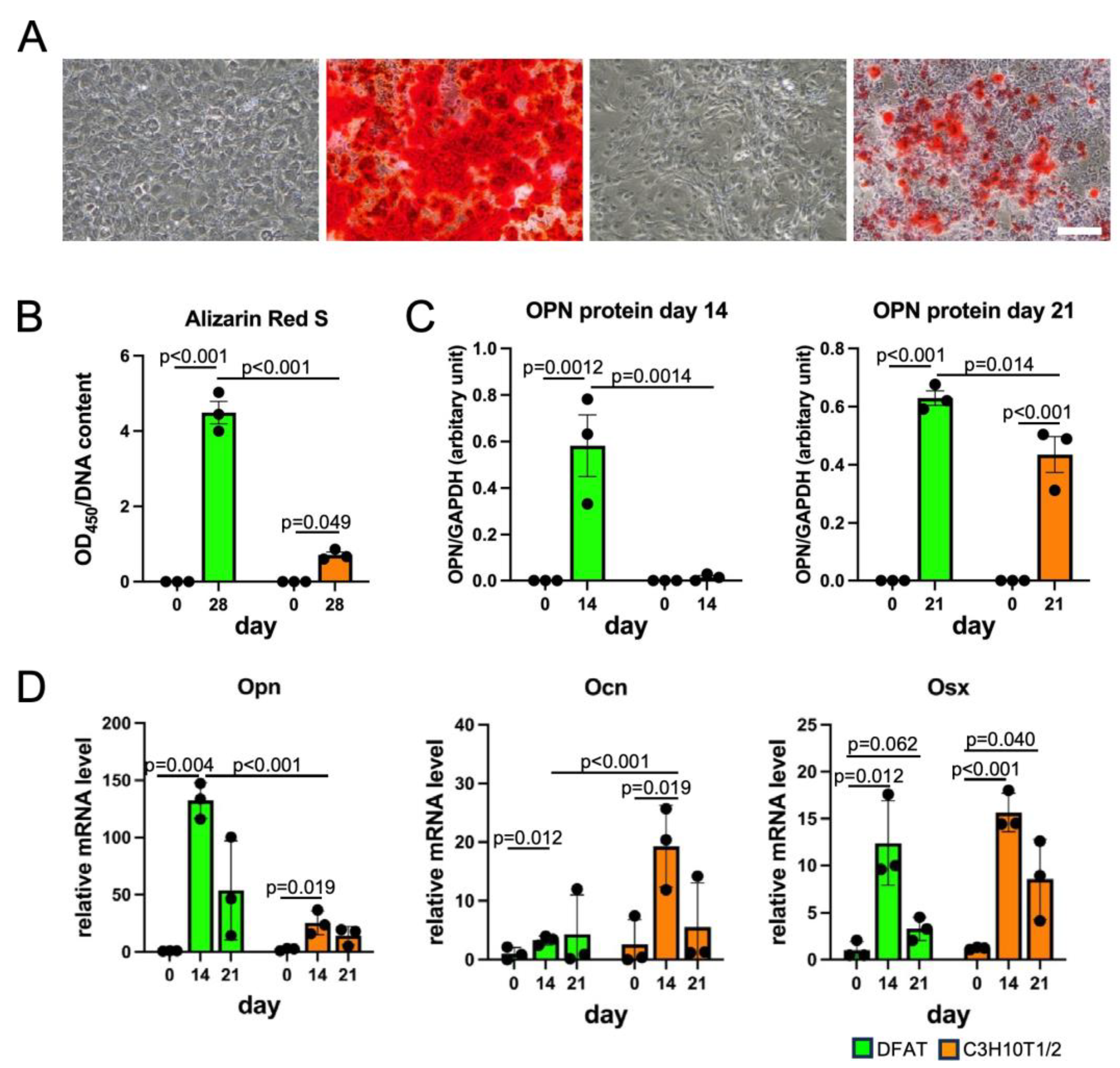
2.6. Osteogenesis and Alizarin Red S Staining
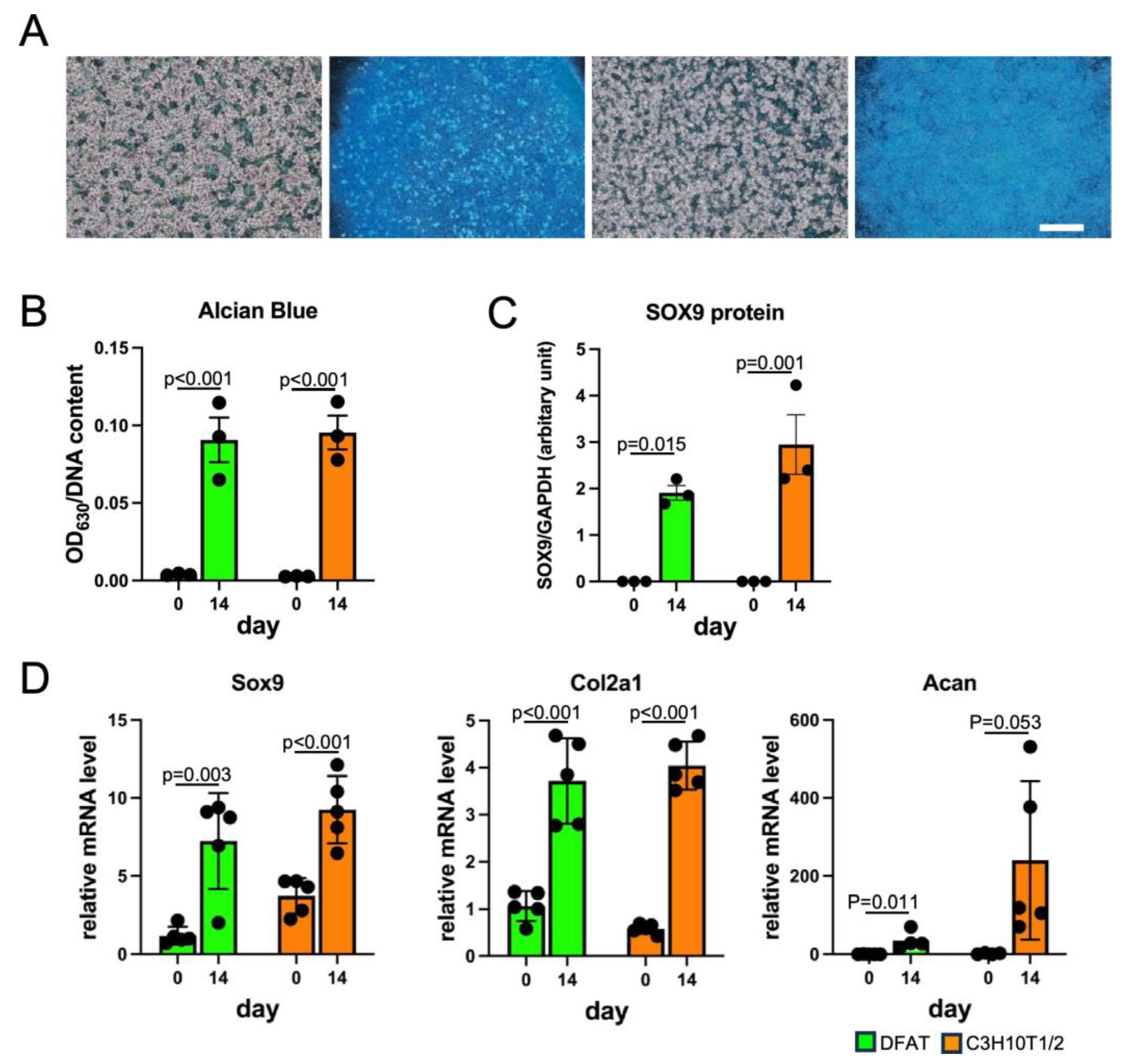
2.7. Chondrogenesis and Alcian Blue Staining
2.8. Real Time Polymerase Chain Reaction
2.9. Western Blotting
2.10. Statistical Analysis
3. Results
3.1. C3H10T1/2 Adipocytes Underwent Dedifferentiation
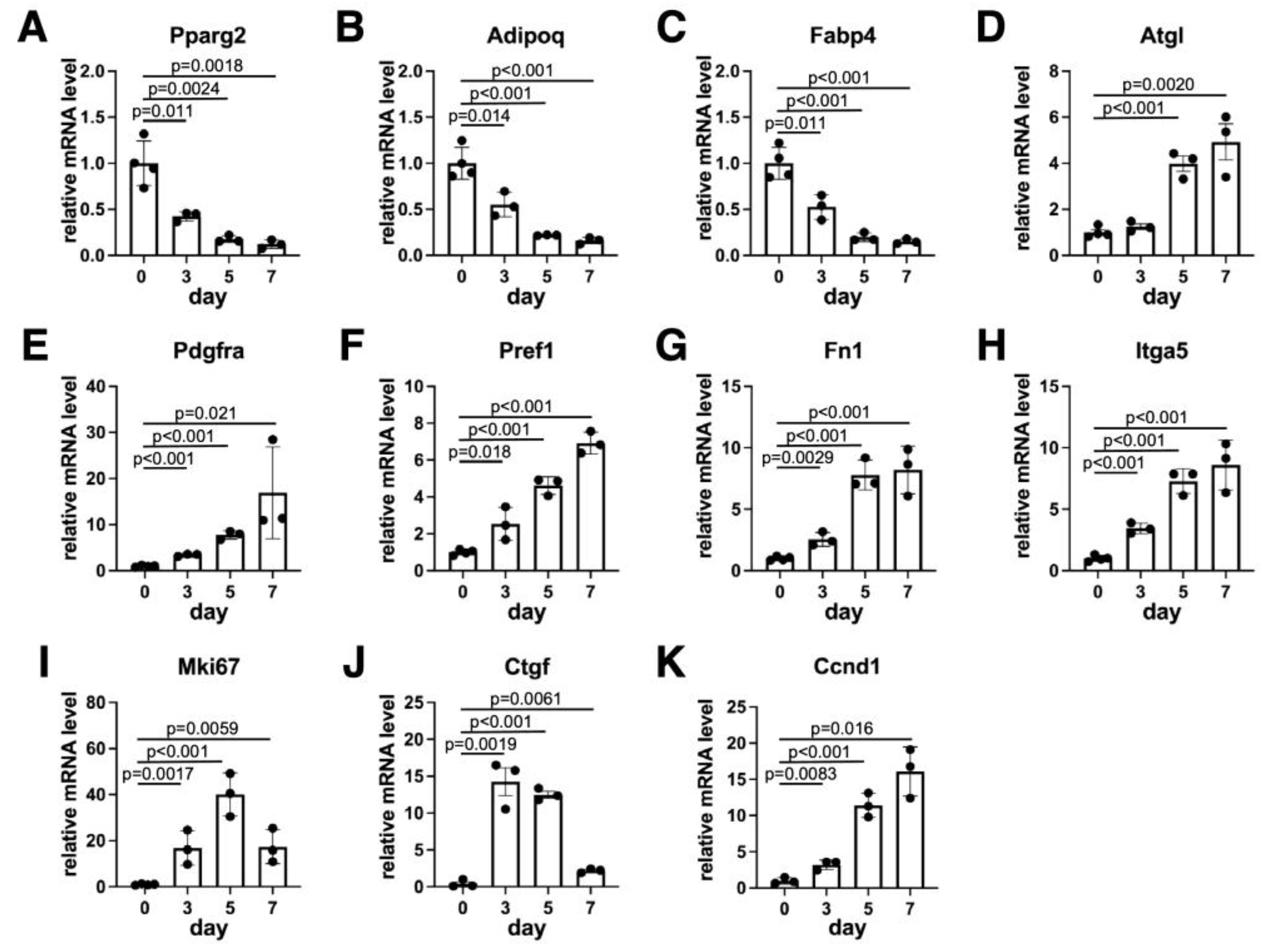
3.2. Gene Expression Levels During Dedifferentiation
3.3. DFAT-like Cells Were Able to Differentiate into Multiple Cell Lineages
3.4. Individually Cultured Single Adipocytes Underwent Dedifferentiation to Proliferate DFAT-like Cells
3.5. Single-Adipocyte-Derived DFAT-like Cells Were Also Able to Differentiate into Multiple Cell Lineages
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scherer, P.E. The many secret lives of adipocytes: Implications for diabetes. Diabetologia 2019, 62, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Uccelli, A.; Moretta, L.; Pistoia, V. Mesenchymal stem cells in health and disease. Nat. Rev. Immunol. 2008, 8, 726–736. [Google Scholar] [CrossRef] [PubMed]
- Yagi, K.; Kondo, D.; Okazaki, Y.; Kano, K. A novel preadipocyte cell line established from mouse adult mature adipocytes. Biochem. Biophys. Res. Commun. 2004, 321, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Sawada, H.; Kazama, T.; Nagaoka, Y.; Arai, Y.; Kano, K.; Uei, H.; Tokuhashi, Y.; Nakanishi, K.; Matsumoto, T. Bone marrow-derived dedifferentiated fat cells exhibit similar phenotype as bone marrow mesenchymal stem cells with high osteogenic differentiation and bone regeneration ability. J. Orthop. Surg. Res. 2023, 18, 191. [Google Scholar] [CrossRef]
- Matsumoto, T.; Kano, K.; Kondo, D.; Fukuda, N.; Iribe, Y.; Tanaka, N.; Matsubara, Y.; Sakuma, T.; Satomi, A.; Otaki, M.; et al. Mature adipocyte-derived dedifferentiated fat cells exhibit multilineage potential. J. Cell Physiol. 2008, 215, 210–222. [Google Scholar] [CrossRef]
- Viswanathan, S.; Shi, Y.; Galipeau, J.; Krampera, M.; Leblanc, K.; Martin, I.; Nolta, J.; Phinney, D.G.; Sensebe, L. Mesenchymal stem versus stromal cells: International Society for Cell & Gene Therapy (ISCT®) Mesenchymal Stromal Cell committee position statement on nomenclature. Cytotherapy 2019, 21, 1019–1024. [Google Scholar] [CrossRef]
- Watanabe, H.; Goto, S.; Kato, R.; Komiyama, S.; Nagaoka, Y.; Kazama, T.; Yamamoto, C.; Li, Y.; Konuma, N.; Hagikura, K.; et al. The neovascularization effect of dedifferentiated fat cells. Sci. Rep. 2020, 10, 9211. [Google Scholar] [CrossRef]
- Fujimaki, H.; Matsumine, H.; Osaki, H.; Ueta, Y.; Kamei, W.; Shimizu, M.; Hashimoto, K.; Fujii, K.; Kazama, T.; Matsumoto, T.; et al. Dedifferentiated fat cells in polyglycolic acid-collagen nerve conduits promote rat facial nerve regeneration. Regen. Ther. 2019, 11, 240–248. [Google Scholar] [CrossRef]
- Jumabay, M.; Zhang, L.; Yao, J.; Bostrom, K.I. Progenitor cells from brown adipose tissue undergo neurogenic differentiation. Sci. Rep. 2022, 12, 5614. [Google Scholar] [CrossRef]
- Utsunomiya, K.; Maruyama, T.; Shimizu, S.; Matsumoto, T.; Endo, M.; Kobayashi, H.; Kano, K.; Abe, M.; Fukuda, N. Implantation of dedifferentiated fat cells ameliorated antineutrophil cytoplasmic antibody glomerulonephritis by immunosuppression and increases in tumor necrosis factor-stimulated gene-6. Stem Cell Res. Ther. 2022, 13, 319. [Google Scholar] [CrossRef]
- Maruyama, T.; Fukuda, N.; Matsumoto, T.; Kano, K.; Endo, M.; Kazama, M.; Kazama, T.; Ikeda, J.; Matsuda, H.; Ueno, T.; et al. Systematic implantation of dedifferentiated fat cells ameliorated monoclonal antibody 1-22-3-induced glomerulonephritis by immunosuppression with increases in TNF-stimulated gene 6. Stem Cell Res. Ther. 2015, 6, 80. [Google Scholar] [CrossRef] [PubMed]
- Ishioka, S.; Hosokawa, T.; Ikeda, T.; Konuma, N.; Kaneda, H.; Ohashi, K.; Furuya, T.; Masuko, T.; Taniguchi, H.; Kano, K.; et al. Therapeutic potential of mature adipocyte-derived dedifferentiated fat cells for inflammatory bowel disease. Pediatr. Surg. Int. 2020, 36, 799–807. [Google Scholar] [CrossRef]
- Kakudo, T.; Kishimoto, N.; Matsuyama, T.; Momota, Y. Functional recovery by application of human dedifferentiated fat cells on cerebral infarction mice model. Cytotechnology 2018, 70, 949–959. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef]
- Kaufmann, K.B.; Buning, H.; Galy, A.; Schambach, A.; Grez, M. Gene therapy on the move. EMBO Mol. Med. 2013, 5, 1642–1661. [Google Scholar] [CrossRef] [PubMed]
- Reznikoff, C.A.; Brankow, D.W.; Heidelberger, C. Establishment and characterization of a cloned line of C3H mouse embryo cells sensitive to postconfluence inhibition of division. Cancer Res. 1973, 33, 3231–3238. [Google Scholar]
- Zehentner, B.K.; Leser, U.; Burtscher, H. BMP-2 and sonic hedgehog have contrary effects on adipocyte-like differentiation of C3H10T1/2 cells. DNA Cell Biol. 2000, 19, 275–281. [Google Scholar] [CrossRef]
- Denker, A.E.; Haas, A.R.; Nicoll, S.B.; Tuan, R.S. Chondrogenic differentiation of murine C3H10T1/2 multipotential mesenchymal cells: I. Stimulation by bone morphogenetic protein-2 in high-density micromass cultures. Differentiation 1999, 64, 67–76. [Google Scholar] [CrossRef]
- Mie, M.; Ohgushi, H.; Yanagida, Y.; Haruyama, T.; Kobatake, E.; Aizawa, M. Osteogenesis coordinated in C3H10T1/2 cells by adipogenesis-dependent BMP-2 expression system. Tissue Eng. 2000, 6, 9–18. [Google Scholar] [CrossRef]
- Lehmann, J.M.; Lenhard, J.M.; Oliver, B.B.; Ringold, G.M.; Kliewer, S.A. Peroxisome proliferator-activated receptors alpha and gamma are activated by indomethacin and other non-steroidal anti-inflammatory drugs. J. Biol. Chem. 1997, 272, 3406–3410. [Google Scholar] [CrossRef]
- Alonso-Perez, A.; Guillan-Fresco, M.; Franco-Trepat, E.; Jorge-Mora, A.; Lopez-Fagundez, M.; Pazos-Perez, A.; Crespo-Golmar, A.; Caeiro-Rey, J.R.; Gomez, R. Improved Protocol to Study Osteoblast and Adipocyte Differentiation Balance. Biomedicines 2022, 11, 31. [Google Scholar] [CrossRef] [PubMed]
- Macrae, V.E.; Davey, M.G.; McTeir, L.; Narisawa, S.; Yadav, M.C.; Millan, J.L.; Farquharson, C. Inhibition of PHOSPHO1 activity results in impaired skeletal mineralization during limb development of the chick. Bone 2010, 46, 1146–1155. [Google Scholar] [CrossRef]
- Shook, B.A.; Wasko, R.R.; Mano, O.; Rutenberg-Schoenberg, M.; Rudolph, M.C.; Zirak, B.; Rivera-Gonzalez, G.C.; Lopez-Giraldez, F.; Zarini, S.; Rezza, A.; et al. Dermal Adipocyte Lipolysis and Myofibroblast Conversion Are Required for Efficient Skin Repair. Cell Stem Cell 2020, 26, 880–895 e886. [Google Scholar] [CrossRef]
- Hirakawa, H.; Gao, L.; Tavakol, D.N.; Vunjak-Novakovic, G.; Ding, L. Cellular plasticity of the bone marrow niche promotes hematopoietic stem cell regeneration. Nat. Genet. 2023, 55, 1941–1952. [Google Scholar] [CrossRef] [PubMed]
- Cave, E.; Crowther, N.J. The Use of 3T3-L1 Murine Preadipocytes as a Model of Adipogenesis. Methods Mol. Biol. 2019, 1916, 263–272. [Google Scholar] [CrossRef] [PubMed]
- MacDougald, O.A.; Lane, M.D. Transcriptional regulation of gene expression during adipocyte differentiation. Annu. Rev. Biochem. 1995, 64, 345–373. [Google Scholar] [CrossRef]
- Siersbaek, R.; Nielsen, R.; John, S.; Sung, M.H.; Baek, S.; Loft, A.; Hager, G.L.; Mandrup, S. Extensive chromatin remodelling and establishment of transcription factor ‘hotspots’ during early adipogenesis. EMBO J. 2011, 30, 1459–1472. [Google Scholar] [CrossRef]
- Fu, M.; Sun, T.; Bookout, A.L.; Downes, M.; Yu, R.T.; Evans, R.M.; Mangelsdorf, D.J. A Nuclear Receptor Atlas: 3T3-L1 adipogenesis. Mol. Endocrinol. 2005, 19, 2437–2450. [Google Scholar] [CrossRef]
- Kishimoto, N.; Momota, Y.; Hashimoto, Y.; Tatsumi, S.; Ando, K.; Omasa, T.; Kotani, J. The osteoblastic differentiation ability of human dedifferentiated fat cells is higher than that of adipose stem cells from the buccal fat pad. Clin. Oral. Investig. 2014, 18, 1893–1901. [Google Scholar] [CrossRef]
- Wang, Q.A.; Song, A.; Chen, W.; Schwalie, P.C.; Zhang, F.; Vishvanath, L.; Jiang, L.; Ye, R.; Shao, M.; Tao, C.; et al. Reversible De-differentiation of Mature White Adipocytes into Preadipocyte-like Precursors during Lactation. Cell Metab. 2018, 28, 282–288.e283. [Google Scholar] [CrossRef]
- Zwick, R.K.; Rudolph, M.C.; Shook, B.A.; Holtrup, B.; Roth, E.; Lei, V.; Van Keymeulen, A.; Seewaldt, V.; Kwei, S.; Wysolmerski, J.; et al. Adipocyte hypertrophy and lipid dynamics underlie mammary gland remodeling after lactation. Nat. Commun. 2018, 9, 3592. [Google Scholar] [CrossRef]
- Zhang, Z.; Shao, M.; Hepler, C.; Zi, Z.; Zhao, S.; An, Y.A.; Zhu, Y.; Ghaben, A.L.; Wang, M.Y.; Li, N.; et al. Dermal adipose tissue has high plasticity and undergoes reversible dedifferentiation in mice. J. Clin. Investig. 2019, 129, 5327–5342. [Google Scholar] [CrossRef] [PubMed]
- Zoico, E.; Darra, E.; Rizzatti, V.; Budui, S.; Franceschetti, G.; Mazzali, G.; Rossi, A.P.; Fantin, F.; Menegazzi, M.; Cinti, S.; et al. Adipocytes WNT5a mediated dedifferentiation: A possible target in pancreatic cancer microenvironment. Oncotarget 2016, 7, 20223–20235. [Google Scholar] [CrossRef]
- Li, Y.; Mao, A.S.; Seo, B.R.; Zhao, X.; Gupta, S.K.; Chen, M.; Han, Y.L.; Shih, T.Y.; Mooney, D.J.; Guo, M. Compression-induced dedifferentiation of adipocytes promotes tumor progression. Sci. Adv. 2020, 6, eaax5611. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Wang, Y.; Pan, Y.; Tian, L.; Choi, M.H.; Wang, L.; Kim, J.Y.; Zhang, J.; Cheng, S.H.; Zhang, L. Hypertonicity induces mitochondrial extracellular vesicles (MEVs) that activate TNF-alpha and beta-catenin signaling to promote adipocyte dedifferentiation. Stem Cell Res. Ther. 2023, 14, 333. [Google Scholar] [CrossRef]
- Sun, L.; Zhang, X.; Wu, S.; Liu, Y.; Guerrero-Juarez, C.F.; Liu, W.; Huang, J.; Yao, Q.; Yin, M.; Li, J.; et al. Dynamic interplay between IL-1 and WNT pathways in regulating dermal adipocyte lineage cells during skin development and wound regeneration. Cell Rep. 2023, 42, 112647. [Google Scholar] [CrossRef] [PubMed]
- Park, H.W.; Kim, Y.C.; Yu, B.; Moroishi, T.; Mo, J.S.; Plouffe, S.W.; Meng, Z.; Lin, K.C.; Yu, F.X.; Alexander, C.M.; et al. Alternative Wnt Signaling Activates YAP/TAZ. Cell 2015, 162, 780–794. [Google Scholar] [CrossRef]
- Kim, J.; Park, K.Y.; Choi, S.; Ko, U.H.; Lim, D.S.; Suh, J.M.; Shin, J.H. Ceiling culture chip reveals dynamic lipid droplet transport during adipocyte dedifferentiation via actin remodeling. Lab. Chip 2022, 22, 3920–3932. [Google Scholar] [CrossRef]
- Tang, Q.Q.; Otto, T.C.; Lane, M.D. Commitment of C3H10T1/2 pluripotent stem cells to the adipocyte lineage. Proc. Natl. Acad. Sci. USA 2004, 101, 9607–9611. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (Accession #) | Forward | Reverse | |
|---|---|---|---|
| Reference | U36b4 (NM_007475) | 5′-cgtcctcgttggagtgaca-3′ | 5′-cggtgegtcagggattg-3′ |
| Adipogenesis | Pparg2 (NM_011146) | 5′-caccagtgtgaattacagcaaatc-3′ | 5′-acaggagaatcteccagagtite-3′ |
| Adipoq (NM_009605) | 5′-tgtcagtggatctgacgaca-3′ | 5′-aacaggagagcttgcaacagt-3′ | |
| Fabp4 (NM_024406) | 5′-gtgaaaacttegatgattacatgaa-3′ | 5′-gcctgccactttecttgtg-3′ | |
| Atgl (NM_025802) | 5′-caacgccactcacatctacgg-3′ | 5′-ggacacctcaataatgttggcac-3′ | |
| Osteoblast | Ocn (NM_001305448) | 5′-ccaagcaggagggcaata-3′ | 5′-tegtcacaagcagggtca-3′ |
| Osx (NM_130458) | 5′-gggagcagagtgccaaga-3′ | 5′-tactectggcgcataggg-3′ | |
| Opn (NM_001204201) | 5′-ttgcttttgcctgtttggca-3′ | 5′-gatctgggtgcaggctgtaa-3′ | |
| Sox9 (NM_011448) | 5′-gagcccgatctgaagaagga-3′ | 5′-gcttgacgtgeggcttgttc-3′ | |
| Chondrocyte | Col2a1 (NM_031163) | 5′-caccaaattcctgtteagcc-3′ | 5′-tgcacgaaacacactggtaag-3′ |
| Acan (NM_001361500) | 5′-aacaactgcaggctgcctat-3′ | 5′-ccagggaactegtccttgtc-3′ | |
| Preadipocyte | Pdgfra (NM_001083316) | 5′-ctcacagacttcggaagaga-3′ | 5′-aagtcgctctcacacactta-3′ |
| Pref1 (NM_010052) | 5′-gacctggagaaaggccagta-3′ | 5′-agggagaaccattgatcacg-3′ | |
| Fn1 (NM_010233) | 5′-gccacacctacaaccagtat-3′ | 5′-gggctggaaagattactctc-3′ | |
| Itga5 (NM_010577) | 5′-cctagccattctttttggcc-3′ | 5′-ggcttgagctgagctttttc-3′ | |
| Proliferation | Mki67 (NM_001081117) | 5′-tcattgaccgctcctttaggt-3′ | 5′-ttgaccttccccatcagggt-3′ |
| WNT/β-catenin | Ctgf (NM_010217) | 5′-ccgccaaccgcaagat-3′ | 5′-cgacccaccgaagacaca-3′ |
| Ccnd1 (NM_001379248) | 5′-caacttcctctcctgctaccg-3′ | 5′-ccttgtttagccagaggccg-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuuki, Y.; Katafuchi, T.; Kazama, T.; Matsumoto, T.; Makishima, M. C3H10T1/2 Mesenchymal Stem Cell Line as a New In Vitro Tool for Studying Adipocyte Dedifferentiation. Biology 2025, 14, 444. https://doi.org/10.3390/biology14040444
Yuuki Y, Katafuchi T, Kazama T, Matsumoto T, Makishima M. C3H10T1/2 Mesenchymal Stem Cell Line as a New In Vitro Tool for Studying Adipocyte Dedifferentiation. Biology. 2025; 14(4):444. https://doi.org/10.3390/biology14040444
Chicago/Turabian StyleYuuki, Yuriko, Takeshi Katafuchi, Tomohiko Kazama, Taro Matsumoto, and Makoto Makishima. 2025. "C3H10T1/2 Mesenchymal Stem Cell Line as a New In Vitro Tool for Studying Adipocyte Dedifferentiation" Biology 14, no. 4: 444. https://doi.org/10.3390/biology14040444
APA StyleYuuki, Y., Katafuchi, T., Kazama, T., Matsumoto, T., & Makishima, M. (2025). C3H10T1/2 Mesenchymal Stem Cell Line as a New In Vitro Tool for Studying Adipocyte Dedifferentiation. Biology, 14(4), 444. https://doi.org/10.3390/biology14040444