
Nitrogen-Fixing Bacteria Promote the Growth of Fritillaria taipaiensis P. Y. Li by Regulating Physiological and Biochemical Reactions and Protecting Enzyme System-Related Gene Expression

 ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
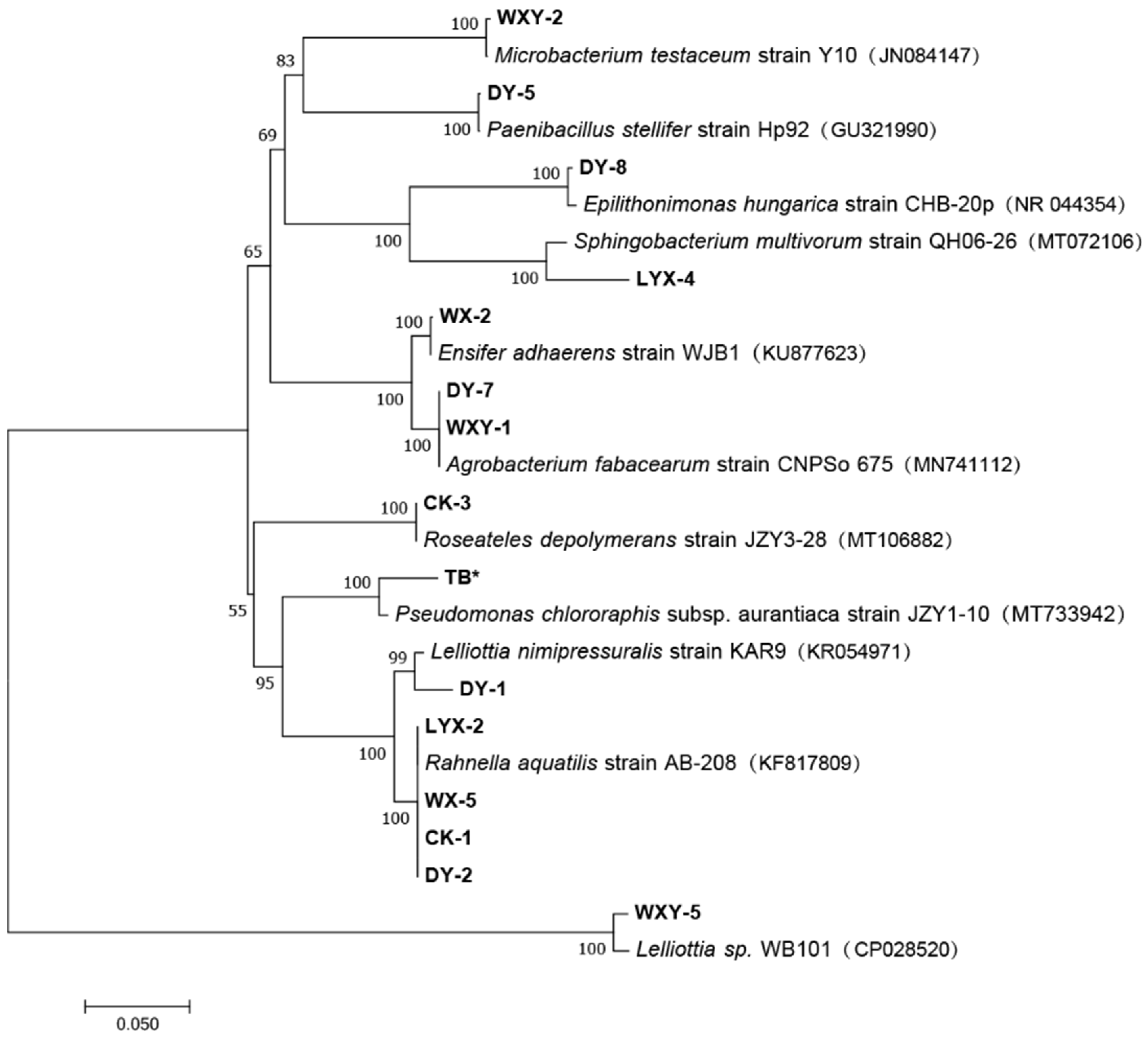
2.1. Strain Isolation and Identification
2.2. Test Site and Experimental Design
2.3. Index Measurement
2.4. Statistical Analysis of Data
3. Results
3.1. Strain Isolation and Screen
3.2. Influences of Different Treatments on the Growth Index of F. taipaiensis Leaves
3.3. Influences of Different Treatments on Photosynthetic Parameters and Photosynthetic Pigment Content of F. taipaiensis Leaves
3.4. Influences of Different Treatments on MDA, Soluble Sugar, Soluble Protein, and Proline Levels of F. taipaiensis Leaves
3.5. Influences of Different Treatments on Antioxidant Enzyme Activities in the Leaves of F. taipaiensis
3.6. Influences of Different Treatments on Genes Related to Antioxidant Enzyme Systems in the Leaves of F. taipaiensis
3.7. The Pearson Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | 16S rDNA Gene Sequences |
|---|---|
| WX-2 | AACKGGCGGCMGGCTTAACaCATGCgAGTCGAgCGCCCCGCAAGGGGAGYGGCMGACGGGTGAGTAACGCGTGGGAATCTACCSTKYYCTRCGGAATAACKCMKGGAAACKKGWRYTAATACCGYATRMGCCCTWCGGGGGAAAGATTTATCGGGRAAKGATGAGCCCGCGTTGGATTAGCTAGTTGGWGGGGTAAAGGCCTACCAAGGCGACGATCCATAGCTGGTCTGASAGGATGATCARCCACATTGGGACTGARACACGGSCCAAACTCCTACGGGAGGCAGCAGTGGGGAATATTGGACAATGGGCGCAAGCCTGATCCAGCCATGCCGCGTGAGTGATGAAGGCCYTAGGGTTGTAAAGCTCTTTCACCGGTGAAGATAATGACGGTAACCGGAGAAGAAGCCCCGGCTAACTTCGTGCCARCAGCCGCGGTAATACGAAGGGGGCTAGCGTTGTTCGGAATTACTGGGCGTAAAGCGCACGTAGGCGGACATTTAAGTCAGGGGTGAAATCCCGGGGCTCAACCCCGGAACTGCCTTTGATACTGGGTGTCTAGAGTATGGAAGAGGTGAGTGGAATTCCGAGTGTAGAGGTGAAATTCGTAGATATTCGGAGGAACACCAGTGGCGAAGGCGGCTCACTGGTCCATTACTGACGCTGAGGTGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCTGGTAGTCCACGCCGTAAACGATGAATGTTAGCCGTCGGGCAGTWTACTGTTCGGTGGCGCAGCTAACGCATTAAACATTCCGCCTGGGGAGTACGGTCGCAAGATTAAAACTCAAAGGAATTGACGGGGGCCCGCACAAGCGGTGGAGCATGTGGTTTAATTYGAAGCAACGCGCAGAACCTTACCAGCCCTTGACATCCCGATTKGGGcATTacGGAGACGTTTTTCCTTCAGTTTGGCTGGATCGGAgACAGGTGSTGCATGGCTGTTGTCAGCTYGTGTYGTGAGATGTTGGGTTAAGTCCCGCAACGAGCGCAACCCTCSCCCTTAGTTGCCAGCATTTAGTTGGGCMCTYTAAGGGGACTGCCGGTGATAAGCCGAGAGGAAGGTGGGGATGACGTCAAGTCYTCATGGCCCTTACGGGYTGGGCTACACACGTGCTACAATGGTGGTGACAGTGGGCAGCGAGACMGCGAKGTCGAGCTAATCTCCAAAASCCATCTCAGTTCGGATTGCACTCTGCAACTCGAGTGCATGAAGTTGGAATCGCTAGTAATCGCAGATCAGCATGYTGCGGTGAATACGTTCCCGGGCCTTGTACACACCGCCCGTCACACCATGGGAGTTGGTTHTMCCCGAAGGTAGTGCGCTAACCGCAAGGAGGCAGCTAACCACaGKAGGTMAMGGGGGG |
| WX-5 | CGCAGTGCGGCAGCTACACATGCAGTCGAGCGGCAGCGGAAAGTAGCTTGCTACTTTGCCGGCGAGCGGCGGACGGGTGAGTAATGTCTGGGAAACTGCCTGATGGAGGGGGATAACTACTGGAAACGGTAGCTAATACCGCATGACCTCGAAAGAGCAAAGTGGGGGATCTTCGGACCTCACGCCATCGGATGTGCCCAGATGGGATTAGCTAGTAGGTGAGGTAATGGCTCACCTAGGCGACGATCCCTAGCTGGTCTGAGAGGATGACCAGCCACACTGGAACTGAGACACGGTCCAGACTCCTACGGGAGGCAGCAGTGGGGAATATTGCACAATGGGCGCAAGCCTGATGCAGCCATGCCGCGTGTGTGAAGAAGGCCTTAGGGTTGTAAAGCACTTTCAGCGAGGAGGAAGGCATCACACTTAATACGTGTGGTGATTGACGTTACTCGCAGAAGAAGCACCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGGTGCAAGCGTTAATCGGAATTACTGGGCGTAAAGCGCACGCAGGCGGTTTGTTAAGTCAGATGTGAAATCCCCGCGCTTAACGTGGGAACTGCATTTGAAACTGGCAAGCTAGAGTCTTGTAGAGGGGGGTAGAATTCCAGGTGTAGCGGTGAAATGCGTAGAGATCTGGAGGAATACCGGTGGCGAAGGCGGCCCCCTGGACAAAGACTGACGCTCAGGTGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCTGGTAGTCCACGCTGTAAACGATGTCGACTTGGAGGTTGTGCCCTTGAGGCGTGGCTTCCGGAGCTAACGCGTTAAGTCGACCGCCTGGGGAGTACGGCCGCAAGGTTAAAACTCAAATGAATTGACGGGGGCCCGCACAAGCGGTGGAGCATGTGGTTTAATTCGATGCAACGCGAAGAACCTTACCTACTCTTGACATCCACGGAATTCGCCAGAGATGGCTTAGTGCCTTCGGGAACCGTGAGACAGGTGCTGCATGGCTGTCGTCAGCTCGTGTTGTGAAATGTTGGGTTAAGTCCCGCAACGAGCGCAACCCTTATCCTTTGTTGCCAGCACGTAATGGTGGGAACTCAAAGGAGACTGCCGGTGATAAACCGGAGGAAGGTGGGGATGACGTCAAGTCATCATGGCCCTTACGAGTAGGGCTACACACGTGCTACAATGGCATATACAAAGAGAAGCGAACTCGCGAGAGCAAGCGGACCTCATAAAGTATGTCGTAGTCCGGATTGGAGTCTGCAACTCGACTCCATGAAGTCGGAATCGCTAGTAATCGTAGATCAGAATGCTACGGTGAATACGTTCCCGGGCCTTGTACACACCGCCCGTCACACCATGGGAGTGGGTTGCAAAAGAAGTAGGTAGCTTAACCTTCGGGAGGGCGCTTACCACTTGGAATCCGGGGG |
| DY-1 | TTGCGGGCGGGCTACACATGCAGTCGAGCGGTAGCACAGAGAGCTTGCTCTCGGGTGACGAGCGGCGGACGGGTGAGTAATGTCTGGGAAACTGCCTGATGGAGGGGGATAACTACTGGAAACGGTAGCTAATACCGCATAACGTCTCAAGACCAAAGAGGGGGACCTTCGGGCCTCTTGCCATCTCATGTGCCCAGATGGGATTAGCTAGTAGGTGGGGTAATGGGTCACCTATGCGACAATCCCTATCTGGTCTGAGAGGATGACCACCCACACTGGAACTGAGACACGGTCCACACTCCTACGGGAGGCAGCAGTGGGGAATATTGCGCAATGGGCGCAAGCCTGATGCACCCATGCCGCGTGTATGAAAAAAGCCTTCTGGTTGTAAAGTACTTTCTCCGAGGAGGAAAGCATTGTGGTTAATAACCCCAGTGATTGACGTTACTCTCACAAAAAACACCGGCTAACTCCCTGCCACCAGCCGCGGTAATACAGAGGGTGCAAGCGTTAATCTGAATTACTGGGCGTAAAGCGCACGCASGCGGTCTGTCAAGTCGGATGTGAAATCCCCGGGCTCAACCTGGGAACTGCRTTCGAAACTGGCAGGCTAKAGTCTTGTAGAGGGGGGTAGAATTCCAGGTGTAGCGGTGAAATGCGTAGAGATCTGGAGGAATACCGGTGGCGAARGCGGCCCCCTGGACAAAGACTGACRCTCASGTGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCTGGTAGTCCACGCCGTAAACGATGTCKACTTGGAGGTTGTTCCCTTGAGGAGTGGCTTCCGGAGCTAACGCGTTAAGTCGACCGCCTGGGGAGTACGGCCGCAAGGTTAAAACTCAAATGAATTGACGGGGGCCCGCACAAgCGGTGGAGCATGTGGTTTAATTCGATGCaACGCGAAGAACCTtACCTACTCTTGACATCCACGGAATTTAGCAGAGATGCTTTAGTGCCTTCGGGAACCGTGAGACAGGTGCTGCATGGCTGTCGTCAGCTCGTGTTGTGAAATGTTGGGTTAAGTCCCGCAACGAGCGCAACCCTTATCCTTTGTTGCCAGCGGTTCGGCCGGGAACTCAAAGGAGACTGCCAGTGATAAACTGGAGGAAGGTGGGGATGACGTCAAGTCATCATGGCCCTTACGAGTAGGGCTACACACGTGCTACAATGGCGCATACAAAGAGAAGCGACCTCGCGAGAGCAAGCGGACCTCATAAAGTGCGTCGTAGTCCGGATTGGAGTCTGCAACTCGACTCCATGAAGTCGGAATCGCTAGTAATCGTAGATCAGAATGCTACGGTGAATACGTTCCCGGGCCTTGTACACACCGCCCGTCACACCATGGGAGTGGGTTGCAAAAGAAGTAGGTAGCTTAACCTTCGGGAGGGCGCTTACCACTTTTGATAA |
| DY-2 | CGCAAGGGGGGCAGCTACACATGCAGTCGAGCGGCAGCGGAAAGTAGCTTGCTACTTTGCCGGCGAGCGGCGGACGGGTGAGTAATGTCTGGGAAACTGCCTGATGGAGGGGGATAACTACTGGAAACGGTAGCTAATACCGCATGACCTCGAAAGAGCAAAGTGGGGGATCTTCGGACCTCACGCCATCGGATGTGCCCAGATGGGATTAGCTAGTAGGTGAGGTAATGGCTCACCTAGGCGACGATCCCTAGCTGGTCTGAGAGGATGACCAGCCACACTGGAACTGAGACACGGTCCAGACTCCTACGGGAGGCAGCAGTGGGGAATATTGCACAATGGGCGCAAGCCTGATGCAGCCATGCCGCGTGTGTGAAGAAGGCCTTAGGGTTGTAAAGCACTTTCAGCGAGGAGGAAGGCATCATACTTAATACGTGTGGTGATTGACGTTACTCGCAGAAGAAGCACCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGGTGCAAGCGTTAATCGGAATTACTGGGCGTAAAGCGCACGCAGGCGGTTTGTTAAGTCAGATGTGAAATCCCCGCGCTTAACGTGGGAACTGCATTTGAAACTGGCAAGCTAGAGTCTTGTAGAGGGGGGTAGAATTCCAGGTGTAGCGGTGAAATGCGTAGAGATCTGGAGGAATACCGGTGGCGAAGGCGGCCCCCTGGACAAAGACTGACGCTCAGGTGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCTGGTAGTCCACGCTGTAAACGATGTCGACTTGGAGGTTGTGCCCTTGAGGCGTGGCTTCCGGAGCTAACGCGTTAAGTCGACCGCCTGGGGAGTACGGCCGCAAGGTTAAAACTCAAATGAATTGACGGGGGCCCGCACAAGCGGTGGAGCATGTGGTTTAATTCGATGCAACGCGAAGAACCTTACCTACTCTTGACATCCACGGAATTCGCCAGAGATGGCTTAGTGCCTTCGGGAACCGTGAGACAGGTGCTGCATGGCTGTCGTCAGCTCGTGTTGTGAAATGTTGGGTTAAGTCCCGCAACGAGCGCAACCCTTATCCTTTGTTGCCAGCACGTAATGGTGGGAACTCAAAGGAGACTGCCGGTGATAAACCGGAGGAAGGTGGGGATGACGTCAAGTCATCATGGCCCTTACGAGTAGGGCTACACACGTGCTACAATGGCATATACAAAGAGAAGCGAACTCGCGAGAGCAAGCGGACCTCATAAAGTATGTCGTAGTCCGGATTGGAGTCTGCAACTCGACTCCATGAAGTCGGAATCGCTAGTAATCGTAGATCAGAATGCTACGGTGAATACGTTCCCGGGCCTTGTACACACCGCCCGTCACACCATGGGAGTGGGTTGCAAAAGAAGTAGGTAGCTTAACCTTCGGGAGGGCGCTACCACTTTGAATTCCAGG |
| DY-5 | GAACGGGGGCGGCGTGCTATACATGCAAGTCGAGCGGAGTTATGAAGGAGCTTGCTCCGGATTAACTTAGCGGCGGACGGGTGAGTAACACGTAGGCAACCTGCCCTTCAGACTGGGATAACTACCGGAAACGGTAGCTAATACCGGATAATTCCTTTCTTCTCCTGAGGAAAGGATGAAAGACGGAGCAATCTGTTACTGAGGGATGGGCCTGCGGCGCATTAGCTAGTTGGTGGGGTAACGGCTCACCAAGGCGACGATGCGTAGCCGACCTGAGAGGGTGAACGGCCACACTGGGACTGAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTAGGGAATCTTCCGCAATGGGCGAAAGCCTGACGGAGCAACGCCGCGTGAGTGATGAAGGTTTTCGGATCGTAAAGCTCTGTTGCCAGGGAAGAACGTCTCTTAGAGTAACTGCTAAAAGAGTGACGGTACCTGAGAAGAAAGCCCCGGCTAACTACGTGCCAGCAGCCGCGGTAATACGTAGGGGGCAAGCGTTGTCCGGAATTATTGGGCGTAAAGCGCGCGCAGGCGGCGATTTAAGTCTGGTGTTTAAACCATGGGCTCAACCTGTGGTCGCATCGGAAACTGGATGGCTTGAGTGCAGAAGAGGAAAGTGGAATTCCACGTGTAGCGGTGAAATGCGTAGAGATGTGGAGGAACACCAGTGGCGAAGGCGACTTTCTGGGCTGTAACTGACGCTGAGGCGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCTGGTAGTCCACGCCGTAAACGATGAGTGCTAGGTGTTAGGGGTTTCGATACCCTTGGTGCCGAAGTTAACACAGTAAGCACTCCGCCTGGGGAGTACGGTCGCAAGACTGAAACTCAAAGGAATTGACGGGGACCCGCACAAGCAGTGGAGTATGTGGTTTAATTCGAAGCAACGCGAAGAACCTTACCAGGTCTTGACATCCCCCTGAATACGTTAGAGATAGCGTAGGCCTTCGGGACAGGGGAGACAGGTGGTGCATGGTTGTCGTCAGCTCGTGTCGTGAGATGTTGGGTTAAGTCCCGCAACGAGCGCAACCCTTGACTTTAGTTGCCAGCAGGTAAGGCTGGGCACTCTAGAGTGACTGCCGGTGACAAACCGGAGGAAGGTGGGGATGACGTCAAATCATCATGCCCCTTATGACCTGGGCTACACACGTACTACAATGGCCGGTACAACGGGAAGCGAAACCGCGAGGTGGAGCCAATCTTATAAAGCCGGTCTCAGTTCGGATTGCAGGCTGCAACTCGCCTGCATGAAGTCGGAATTGCTAGTAATCGCGGATCAGCATGCCGCGGTGAATACGTTCCCGGGTCTTGTACACACCGCCCGTCACACCACGAGAGTTTACAACACCCGAAGTCGGTGGGGTAACCCGTAAGGGAGCCAGCCGCCGAAGGTGGGTAAGATAAGT |
| DY-7 | GAAACCTGGGCCGGGCAGGGCTTAACACATGCAAGTCGAACGCCCCGCAAGGGGAGTGGCAGACGGGTGAGTAACGCGTGGGAATCTACCGTGCCCTGCGGAATAGCTCCGGGAAACTGGAATTAATACCGCATACGCCCTACGGGGGAAAGATTTATCGGGGTATGATGAGCCCGCGTTGGATTAGCTAGTTGGTGGGGTAAAGGCCTACCAAGGCGACGATCCATAGCTGGTCTGAGAGGATGATCAGCCACATTGGGACTGAGACACGGCCCAAACTCCTACGGGAGGCAGCAGTGGGGAATATTGGACAATGGGCGCAAGCCTGATCCAGCCATGCCGCGTGAGTGATGAAGGCCTTAGGGTTGTAAAGCTCTTTCACCGGAGAAGATAATGACGGTATCCGGAGAAGAAGCCCCGGCTAACTTCGTGCCAGCAGCCGCGGTAATACGAAGGGGGCTAGCGTTGTTCGGAATTACTGGGCGTAAAGCGCACGTAGGCGGATATTTAAGTCAGGGGTGAAATCCCAGAGCTCAACTCTGGAACTGCCTTTGATACTGGGTATCTTGAGTATGGAAGAGGTAAGTGGAATTCCGAGTGTAGAGGTGAAATTCGTAGATATTCGGAGGAACACCAGTGGCGAAGGCGGCTTACTGGTCCATTACTGACGCTGAGGTGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCTGGTAGTCCACGCCGTAAACGATGAATGTTAGCCGTCGGGCAGTATACTGTTCGGTGGCGCAGCTAACGCATTAAACATTCCGCCTGGGGAGTACGGTCGCAAGATTAAAACTCAAAGGAATTGACGGGGGCCCGCACAAGCGGTGGAGCATGTGGTTTAATTCGAAGCAACGCGCAGAACCTTACCAGCTCTTGACATTCGGGGTTTGGGCAGTGGAGACATTGTCCTTCAGTTAGGCTGGCCCCAGAACAGGTGCTGCATGGCTGTCGTCAGCTCGTGTCGTGAGATGTTGGGTTAAGTCCCGCAACGAGCGCAACCCTCGCCCTTAGTTGCCAGCATTTAGTTGGGCACTCTAAGGGGACTGCCGGTGATAAGCCGAGAGGAAGGTGGGGATGACGTCAAGTCCTCATGGCCCTTACGGGCTGGGCTACACACGTGCTACAATGGTGGTGACAGTGGGCAGCGAGACAGCGATGTCGAGCTAATCTCCAAAAGCCATCTCAGTTCGGATTGCACTCTGCAACTCGAGTGCATGAAGTTGGAATCGCTAGTAATCGCAGATCAGCATGCTGCGGTGAATACGTTCCCGGGCCTTGTACACACCGCCCGTCACACCATGGGAGTTGGTTTTACCCGAAGGTAGTGCGCTAACCGCAAGGAGGCAGCTAACCACGTAGTCAGGCGGTGTT |
| DY-8 | GATGCCCCTTGCCGGGGAGGCTACACATGCAAGCCGAGCGGTAGAGGTCCTTCGGGACCTTGAGAGCGGCGCACCCGGTGCGGAACACGTGTGCAACCTGCCTTTATCTGGGGGATAGCCTTTCGAAAGGAAGATTAATACCCCATAATATATTGAATGGCATCATTTGATATTGAAAACTCCGGTGGATAGAGATGGGCACGCGCAAGATTAGATAGTTGGTAGGGTAACGGCCTACCAAGTCGATGATCTTTAGGGGGCCTGAGAGGGTGATCCCCCACACTGGTACTGAGACACGGACCAGACTCCTACGGGAGGCAGCAGTGAGGAATATTGGACAATGGGTGAGAGCCTGATCCAGCCATCCCGCGTGAAGGACGACGGCCCTATGGGTTGTAAACTTCTTTTGTACAGGGATAAACCTTTCCACGTGTGGAAAGCTGAAGGTACTGTACGAATAAGCACCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGGTGCAAGCGTTATCCGGATTTATTGGGTTTAAAGGGTCCGTAGGCGGGCCTGTAAGTCAGTGGTGAAATCTCATAGCTCAACTATGAAACTGCCATTGATACTGCAGGCCTTGAGTAGATTTGAAGTGGCTGGAATAAGTAGTGTAGCGGTGAAATGCATAGATATTACTTAGAACACCAATTGCGAAGGCAGGTCACTAAGATCTAACTGACGCTGATGGACGAAAGCGTGGGGAGCGAACAGGATTAGATACCCTGGTAGTCCACGCCGTAAACGATGCTAACTCGTTTTTGGGCTTTCGGGCTCAGAGACTAAGCGAAAGTGATAAGTTAGCCACCTGGGGAGTACGTTCGCAAGAATGAAACTCAAAGGAATTGACGGGGGCCCGCACAAGCGGTGGATTATGTGGTTTAATTCGATGATACGCGAGGAACCTTACCAAGACTTAAATGGGAAATGACAGATTTAGAAATAGATCCTTCTTCGGACATTTTCCAAGGTGCTGCATGGTTGTCGTCAGCTCGTGCCGTGAGGTGTTAGGTTAAGTCCTGCAACGAGCGCAACCCCTGTCACTAGTTGCTACCATTAAGTTGAGGACTCTAGTGAGACTGCCTACGCAAGTAGAGAGGAAGGTGGGGATGACGTCAAATCATCACGGCCCTTACGTCTTGGGCCACACACGTAATACAATGGCCGGTACAGAGGGCAGCTACACAGCGATGTGATGCAAATCTCGAAAGCCGGTCTCAGTTCGGATTGGAGTCTGCAACTCGACTCTATGAAGCTGGAATCGCTAGTAATCGCGCATCAGCCATGGCGCGGTGAATACGTTCCCGGGCCTTGTACACACCGCCCGTCAAGCCATGGAAGTCTGGGGTACCTGAAGTCGGTGACCGTAACAGGAGCTGCCTAGGTTAAACCAAGGAATCGT |
| CK-1 | CGCAGTGGGGCAGCTACACATGCAGTCGAGCGGCAGCGGAAAGTAGCTTGCTACTTTGCCGGCGAGCGGCGGACGGGTGAGTAATGTCTGGGAAACTGCCTGATGGAGGGGGATAACTACTGGAAACGGTAGCTAATACCGCATGACCTCGAAAGAGCAAAGTGGGGGATCTTCGGACCTCACGCCATCGGATGTGCCCAGATGGGATTAGCTAGTAGGTGAGGTAATGGCTCACCTAGGCGACGATCCCTAGCTGGTCTGAGAGGATGACCAGCCACACTGGAACTGAGACACGGTCCAGACTCCTACGGGAGGCAGCAGTGGGGAATATTGCACAATGGGCGCAAGCCTGATGCAGCCATGCCGCGTGTGTGAAGAAGGCCTTAGGGTTGTAAAGCACTTTCAGCGAGGAGGAAGGCATCACACTTAATACGTGTGGTGATTGACGTTACTCGCAGAAGAAGCACCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGGTGCAAGCGTTAATCGGAATTACTGGGCGTAAAGCGCACGCAGGCGGTTTGTTAAGTCAGATGTGAAATCCCCGCGCTTAACGTGGGAACTGCATTTGAAACTGGCAAGCTAGAGTCTTGTAGAGGGGGGTAGAATTCCAGGTGTAGCGGTGAAATGCGTAGAGATCTGGAGGAATACCGGTGGCGAAGGCGGCCCCCTGGACAAAGACTGACGCTCAGGTGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCTGGTAGTCCACGCTGTAAACGATGTCGACTTGGAGGTTGTGCCCTTGAGGCGTGGCTTCCGGAGCTAACGCGTTAAGTCGACCGCCTGGGGAGTACGGCCGCAAGGTTAAAACTCAAATGAATTGACGGGGGCCCGCACAAGCGGTGGAGCATGTGGTTTAATTCGATGCAACGCGAAGAACCTTACCTACTCTTGACATCCACGGAATTCGCCAGAGATGGCTTAGTGCCTTCGGGAACCGTGAGACAGGTGCTGCATGGCTGTCGTCAGCTCGTGTTGTGAAATGTTGGGTTAAGTCCCGCAACGAGCGCAACCCTTATCCTTTGTTGCCAGCACGTAATGGTGGGAACTCAAAGGAGACTGCCGGTGATAAACCGGAGGAAGGTGGGGATGACGTCAAGTCATCATGGCCCTTACGAGTAGGGCTACACACGTGCTACAATGGCATATACAAAGAGAAGCGAACTCGCGAGAGCAAGCGGACCTCATAAAGTATGTCGTAGTCCGGATTGGAGTCTGCAACTCGACTCCATGAAGTCGGAATCGCTAGTAATCGTAGATCAGAATGCTACGGTGAATACGTTCCCGGGCCTTGTACACACCGCCCGTCACACCATGGGAGTGGGTTGCAAAAGAAGTAGGTAGCTTAACCTTCGGGAGGGCGCTACCACCTTTGATTACCGG |
| CK-3 | GTAGTTGCGGCATGCTTAACATGCAGTCGAACGGTAACGCGGGGCAACCTGGCGACGAGTGGCGAACGGGTGAGTAATATATCGGAACGTGCCCAGTTGTGGGGGATAACTGCTCGAAAGAGCAGCTAATACCGCATACGACCTGAGGGTGAAAGCGGGGGATCGCAAGACCTCGCGCAATTGGAGCGGCCGATATCAGATTAGGTAGTTGGTGGGGTAAAGGCCTACCAAGCCGACGATCTGTAGCTGGTCTGAGAGGACGACCAGCCACACTGGGACTGAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATTTTGGACAATGGGGGCAACCCTGATCCAGCCATGCCGCGTGCGGGAAGAAGGCCTTCGGGTTGTAAACCGCTTTTGTCAGGGAAGAAAAGACTCCTACTAATACTGGGGGTTCATGACGGTACCTGAAGAATAAGCACCGGCTAACTACGTGCCAGCAGCCGCGGTAATACGTAGGGTGCAAGCGTTAATCGGAATTACTGGGCGTAAAGCGTGCGCAGGCGGTTATGCAAGACAGATGTGAAATCCCCGGGCTCAACCTGGGAACTGCATTTGTGACTGCATGGCTAGAGTACGGTAGAGGGGGATGGAATTCCGCGTGTAGCAGTGAAATGCGTAGATATGCGGAGGAACACCGATGGCGAAGGCAATCCCCTGGACCTGTACTGACGCTCATGCACGAAAGCGTGGGGAGCAAACAGGATTAGATACCCTGGTAGTCCACGCCCTAAACGATGTCAACTGGTTGTTGGGAGGGTTTCTTCTCAGTAACGTAGCTAACGCGTGAAGTTGACCGCCTGGGGAGTACGGCCGCAAGGTTGAAACTCAAAGGAATTGACGGGGACCCGCACAAGCGGTGGATGATGTGGTTTAATTCGATGCAACGCGAAAAACCTTACCTACCCTTGACATGCCAGGAATCCTGCAGAGATGTGGGAGTGCTCGAAAGAGAACCTGGACACAGGTGCTGCATGGCCGTCGTCAGCTCGTGTCGTGAGATGTTGGGTTAAGTCCCGCAACGAGCGCAACCCTTGTCATTAGTTGCTACGAAAGGGCACTCTAATGAGACTGCCGGTGACAAACCGGAGGAAGGTGGGGATGACGTCAGGTCATCATGGCCCTTATGGGTAGGGCTACACACGTCATACAATGGCCGGGACAGAGGGCTGCCAACCCGCGAGGGGGAGCTAATCCCAGAAACCCGGTCGTAGTCCGGATCGCAGTCTGCAACTCGACTGCGTGAAGTCGGAATCGCTAGTAATCGCGGATCAGCTTGCCGCGGTGAATACGTTCCCGGGTCTTGTACACACCGCCCGTCACACCATGGGAGCGGGTTCTGCCAGAAGTAGTTAGCCTAACCGCAAGGGGGGCGATACCACCGGCAGGTTCCGTTCCC |
| TB * | GACCTTCGGGCCTTGCGCTATCAGATGAGCCTAGGTCGGATTAGCTAATTGGTGAGGTAATGGCTCACCAAAGGAACGATCCGTAACTGGTCTGAAAGGATGATCAGTCCCTCTGGAACTGAGACACGGGCCATACTCCTACGGGAGGCCGCAGTGGGGAATATTGGACAATGGGCGAAAGCCTGATCCAGCCATGCCGCGTGTGTGAAGAAGGTCTTCGGATTGTAAAGCACTTTAAATTGGGAGGAAGGGTTGTTCATGAATACTCTGCAATTTTGACGTTACCGACTGAATAAGCTCCGAATAACTCTGTGCCAACAGCCGCGGTAATACTAAGGGTGCAAGCGTTAATCGGAATTACTGGGCGTAAAGCGCKCGTAGGTGGTTCGTTAATCTGGATGTGAAATCCCCGGGCTCAACCTGGRAACTGCATCCRAACTGGCGAGCTWKAGTATGGWACAGGGTgGTGGAATTTCCTGTGTAGCGGTGAAATGCRTAKATATAtGRAGGAACACCAGTGGCGAASGCGACCACCTGSACTGATACTGACAcTGASGTGCGAAAaGCGTGRGGAGCAAACASGATTASaTACCCTGGDVgTCCACKCCGTAWaCGATGTCAwCTAGCCKTTGGGAGCCTTGAGCTCTTAGTGKCGCAGCTAACGCATTAAGTTGACCGCCTGGGGAGTACGKCCGCAAGGWTAAAACTCAAATGAATTGACGGKGSCCCGCACAAGCGGTGGAGCWTGTGGTTtAATTMGAAGCAACgCGRAGAACCTTACCAGGCCTTCACATCCAATGAACTTTCCAGAGATGGATTGGTGCATTCGGGAACATTGAGACAGGTGCTGCATGGCTGTCGTCAGCTCGTGTCGTGAGATGTTGGGTTAAGTCCCGTAACGAGAGCACCCCTTTTCCCTAGTTACCACCACGTAATGGTGGGCACTCTAAGGAGACTGCCGGTGACAAACCGGAGGAAGGTGGGGAAGACGTCAAGTCCTCATGGCCCTTACGGCCTGGGCTACCCACGTGCTACCATGGTCGGTACAAAGGGTCACCAAGCCACGAGGTGGAGCTAATTCCATAAAACCGATCGTAGTCCGGATCGCAGTCTGCAACTCGAGTGCGTGAAGTCGGAATCGCTAGTAATCGCGAATCAGAATGTCGCGGTGAATACGTTCCCGGGCCTTGTACACACCGCCCGTCACACCATGGGAGTGGGTTGCACCAGAAGTAGTTAGTTTAACCTTAAGGAGGAACGTTTACCCCCGGTGGTTCCAGGGGGT |
| LYX-2 | CATACTTTCCGGCGGGCCTAACACATGCAAGTCGAGCGGCAGCGGAAAGTAGCTTGCTACTTTGCCGGCGAGCGGCGGACGGGTGAGTAATGTCTGGGAAACTGCCTGATGGAGGGGGATAACTACTGGAAACGGTAGCTAATACCGCATGACCTCGAAAGAGCAAAGTGGGGGATCTTCGGACCTCACGCCATCGGATGTGCCCAGATGGGATTAGCTAGTAGGTGAGGTAATGGCTCACCTAGGCGACGATCCCTAGCTGGTCTGAGAGGATGACCAGCCACACTGGAACTGAGACACGGTCCAGACTCCTACGGGAGGCAGCAGTGGGGAATATTGCACAATGGGCGCAAGCCTGATGCAGCCATGCCGCGTGTGTGAAGAAGGCCTTAGGGTTGTAAAGCACTTTCAGCGAGGAGGAAGGCATCACACTTAATACGTGTGGTGATTGACGTTACTCGCAGAAGAAGCACCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGGTGCAAGCGTTAATCGGAATTACTGGGCGTAAAGCGCACGCAGGCGGTTTGTTAAGTCAGATGTGAAATCCCCGCGCTTAACGTGGGAACTGCATTTGAAACTGGCAAGCTAGAGTCTTGTAGAGGGGGGTAGAATTCCAGGTGTAGCGGTGAAATGCGTAGAGATCTGGAGGAATACCGGTGGCGAAGGCGGCCCCCTGGACAAAGACTGACGCTCAGGTGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCTGGTAGTCCACGCTGTAAACGATGTCGACTTGGAGGTTGTGCCCTTGAGGCGTGGCTTCCGGAGCTAACGCGTTAAGTCGACCGCCTGGGGAGTACGGCCGCAAGGTTAAAACTCAAATGAATTGACGGGGGCCCGCACAAGCGGTGGAGCATGTGGTTTAATTCGATGCAACGCGAAGAACCTTACCTACTCTTGACATCCACGGAATTCGCCAGAGATGGCTTAGTGCCTTCGGGAACCGTGAGACAGGTGCTGCATGGCTGTCGTCAGCTCGTGTTGTGAAATGTTGGGTTAAGTCCCGCAACGAGCGCAACCCTTATCCTTTGTTGCCAGCACGTAATGGTGGGAACTCAAAGGAGACTGCCGGTGATAAACCGGAGGAAGGTGGGGATGACGTCAAGTCATCATGGCCCTTACGAGTAGGGCTACACACGTGCTACAATGGCATATACAAAGAGAAGCGAACTCGCGAGAGCAAGCGGACCTCATAAAGTATGTCGTAGTCCGGATTGGAGTCTGCAACTCGACTCCATGAAGTCGGAATCGCTAGTAATCGTAGATCAGAATGCTACGGTGAATACGTTCCCGGGCCTTGTACACACCGCCCGTCACACCATGGGAGTGGGTTGCAAAAGAAGTAGGTAGCTTAACCTTCGGGAGGGCGCTTACCCACTTTGGATTCACGGGGT |
| LYX-4 | GAATGCGGAGCTATACATGCAGTCGGACGGGATCCGTCGGAGAGCTTGCTCGAAGACGGTGAGAGTGGCGCACGGGTGCGTAACGCGTGAGCAACCTACCTCTATCAGGGGGATAGCCTCTCGAAAGAGAGATTAACACCGCATAACATATGTGACCGGCATCGGTTGGATATTAAATATTTATAGGATAGAGATGGGCTCGCGTGACATTAGCTAGTTGGTAGGGTAACGGCTTACCAAGGCGACGATGTCTAGGGGCTCTGAGAGGAGAATCCCCCACACTGGTACTGAGACACGGACCAGACTCCTACGGGAGGCAGCAGTAAGGAATATTGGTCAATGGGCGGAAGCCTGAACCAGCCATGCCGCGTGCAGGATGACTGCCCTATGGGTTGTAAACTGCTTTTGTCCAGGAATAAACCTTTCTACGTGTAGGAAGCTGAATGTACTGGAAGAATAAGGATCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGATCCGAGCGTTATCCGGATTTATTGGGTTTAAAGGGTGCGTAGGCGGCCTATTAAGTCAGGGGTGAAATACGGTGGCTCAACCATCGCAGTGCCTTTGATACTGATGGGCTTGAATCCATTTGAAGTGGGCGGAATAAGACAAGTAGCGGTGAAATGCATAGATATGTCTTAGAACTCSGATTGCGAAGSCAGYTCAYTAAGCTGGTATTGACGCTGATGCACGAAAGCGTGGGGATCGACCAGGATTAGATACCCTGGTAGTCCACGCCCTAAACGATGATAACTCGATGTTGGGGATAGACACCCAGCGTCCAAGCGAAAGCGTTAAGTTATCCACGTGGGGAGTACGCCCGCAAGGGTGAAATTAAAGGGAATTGCCGGGGCCCCGCACAAGGGGAGGACCATGGGTTTAAATTGAAGAaTCCGGGAGGACCTTTACCCGGGCTGAAAAGTTAGGAAAGGGTGCAGAGCCCCCTTGTTCCTTGGGACcAGGAAATTAGGGCTGTCATGACGTTGTtCAGTTGGTCCCGGGGGGTGTTGGGTTAAGTCCCGCAACGAGCCCACCCCTTTTGTTTATTGCCCAGCTGTTAAGGTGGGGACTCTAAACAGACTGCCTGTGCAAACAGAGAGGAAGGTGGGGACGACGTCAAGTCATCATGGCCCTTACGTCCGGGGCTACACACGTGCTACAATGGATGGTACAGCGGGCAGCTACATAGCAATATGATGCTAATCTCTAAAAGCCATTCACAGTTCGGATTGGGGTCTGCAACTCGACCCCATGAAGTTGGATTCGCTAGTAATCGCGTATCAGCAATGACGCGGTGAATACGTTCCCGGGCCTTGTACACACCGCCCGTCAAGCCATGAAAGTTGGGGGTACCTAAAGCATGTTACCGCAAGGAGCGTGTAGGTAAGCCCAG |
| WXY-1 | GACCATTTGGCCGGGAGGGCCTTAACACATGCAAGTCGAACGCCCCGCAAGGGGAGTGGCAGACGGGTGAGTAACGCGTGGGAATCTACCGTGCCCTGCGGAATAGCTCCGGGAAACTGGAATTAATACCGCATACGCCCTACGGGGGAAAGATTTATCGGGGTATGATGAGCCCGCGTTGGATTAGCTAGTTGGTGGGGTAAAGGCCTACCAAGGCGACGATCCATAGCTGGTCTGAGAGGATGATCAGCCACATTGGGACTGAGACACGGCCCAAACTCCTACGGGAGGCAGCAGTGGGGAATATTGGACAATGGGCGCAAGCCTGATCCAGCCATGCCGCGTGAGTGATGAAGGCCTTAGGGTTGTAAAGCTCTTTCACCGGAGAAGATAATGACGGTATCCGGAGAAGAAGCCCCGGCTAACTTCGTGCCAGCAGCCGCGGTAATACGAAGGGGGCTAGCGTTGTTCGGAATTACTGGGCGTAAAGCGCACGTAGGCGGATATTTAAGTCAGGGGTGAAATCCCAGAGCTCAACTCTGGAACTGCCTTTGATACTGGGTATCTTGAGTATGGAAGAGGTAAGTGGAATTCCGAGTGTAGAGGTGAAATTCGTAGATATTCGGAGGAACACCAGTGGCGAAGGCGGCTTACTGGTCCATTACTGACGCTGAGGTGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCTGGTAGTCCACGCCGTAAACGATGAATGTTAGCCGTCGGGCAGTATACTGTTCGGTGGCGCAGCTAACGCATTAAACATTCCGCCTGGGGAGTACGGTCGCAAGATTAAAACTCAAAGGAATTGACGGGGGCCCGCACAAGCGGTGGAGCATGTGGTTTAATTCGAAGCAACGCGCAGAACCTTACCAGCTCTTGACATTCGGGGTTTGGGCAGTGGAGACATTGTCCTTCAGTTAGGCTGGCCCCAGAACAGGTGCTGCATGGCTGTCGTCAGCTCGTGTCGTGAGATGTTGGGTTAAGTCCCGCAACGAGCGCAACCCTCGCCCTTAGTTGCCAGCATTTAGTTGGGCACTCTAAGGGGACTGCCGGTGATAAGCCGAGAGGAAGGTGGGGATGACGTCAAGTCCTCATGGCCCTTACGGGCTGGGCTACACACGTGCTACAATGGTGGTGACAGTGGGCAGCGAGACAGCGATGTCGAGCTAATCTCCAAAAGCCATCTCAGTTCGGATTGCACTCTGCAACTCGAGTGCATGAAGTTGGAATCGCTAGTAATCGCAGATCAGCATGCTGCGGTGAATACGTTCCCGGGCCTTGTACACACCGCCCGTCACACCATGGGAGTTGGTTTTACCCGAAGGTAGTGCGCTAACCGCAAGGAGGCAGCTAACCACGGTAGGTACCGGGGGGT |
| WXY-2 | AGGGACCTTGCGGGTTGCTTTAACCATGCAAGTCGAACGGTGAAGCCAAGCTTGCTTGGTGGATCAGTGCCGAACGGGTGAGTAACACGTGAGCAACCTGCCCTGGACTCTGGGATAAGCGCTGGAAACGGCGTCTAATACTGGATATGAGACGTGATCGCATGGTCGTGTTTGGAAAGATTTTTCGGTCTGGGATGGGCTCGCGGCCTATCAGCTTGTTGGTGAGGTAATGGCTCACCAAGGCGTCGACGGGTAGCCGGCCTGAGAGGGTGACCGGCCACACTGGGACTGAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATATTGCACAATGGGCGAAAGCCTGATGCAGCAACGCCGCGTGAGGGATGACGGCCTTCGGGTTGTAAACCTCTTTTAGCATGGAAGAAGCGAAAGTGACGGTACCTGCAGAAAAAGCGCCGGCTAACTACGTGCCAGCAGCCGCGGTAATACGTAGGGSGCAAGCGTTATCSGGAATTAtTGGGCGTAAAGAGCTCGTAGGCGgTTTGTCGCGTCTGcTGTGaAATCCCGAGGCTCAACCTCGGGCCTGCAGTGGgTACKGGCAGACTAGAGTGCGGTAGGGGAGATTGGAATTCCTGGTGTAKCGGTGGAATGCGCAGATATCAGGAgGGAACACCGATGGCGAAGGCAGATCTCTGGGCCGTAACTGACGCTGAGGAGCGAAAGGGTGGGGAGCAAACAGGCTTAGATACCcTGGTAGTCCACCCCGTAAACGTTGGGGAACTAGTTGTGGGGGACCATTCCACGGTTTCCGTGACGCAGCTAACGCATTAAGTTCCCCGCCTGGGGAGTACGGCCGCAAGGCTAAAACTCAAAGGAATTGACGGGGACCCGCACAAGCGGCGGAGCATGCGGATTAATTCGATGCAACGCGAAGAACCTTACCAAGGCTTGACATATACGAGAACGGGCCAGAAATGGTCAACTCTTTGGACACTCGTAAACAGGTGGTGCATGGTTGTCGTCAGCTCGTGTCGTGAGATGTTGGGTTAAGTCCCGCAACGAGCGCAACCCTCGTTCTATGTTGCCAGCACGTAATGGTGGGAACTCATGGGATACTGCCGGGGTCAACTCGGAGGAAGGTGGGGATGACGTCAAATCATCATGCCCCTTATGTCTTGGGCTTCACGCATGCTACAATGGCCGGTACAAAGGGCTGCAATACCGTGAGGTGGAGCGAATCCCAAAAAGCCGGTCCCAGTTCGGATTGAGGTCTGCAACTCGACCTCATGAAGTCGGAGTCGCTAGTAATCGCAGATCAGCAACGCTGCGGTGAATACGTTCCCGGGTCTTGTACACACCGCCCGTCAAGTCATGAAAGTCGGTAACACCTGAAGCCGGTGGCCCAACCCTTGTGGGAAGGGAAGCCCCGTTTGCT |
| WXY-5 | CGATACAAAGTGGTAGCGCCCTCCCGAAGGTTAAGCTACCTACTTCTTTTGCAACCCACTCCCATGGTGTGACGGGCGGTGTGTACAAGGCCCGGGAACGTATTCACCGTAGCATTCTGATCTACGATTACTAGCGATTCCGACTTCATGGAGTCGAGTTGCAGACTCCAATCCGGACTACGACGCACTTTATGAGGTCCGCTTGCTCTCGCGAGGTCGCTTCTCTTTGTATGCGCCATTGTAGCACGTGTGTAGCCCTACTCGTAAGGGCCATGATGACTTGACGTCATCCCCACCTTCCTCCAGTTTATCACTGGCAGTCTCCTTTGAGTTCCCGGCCGAACCGCTGGCAACAAAGGATAAGGGTTGCGCTCGTTGCGGGACTTAACCCAACATTTCACaACACGAGCTGACGACAGCCATGCAGCACcTGTCTCACGGTTCCCGAAGGCACTAAAGCATCTCTGCTAAATTCCGTGGATGTCAAGAGTAGGTAAGGTTCTTCGCGTTGCATCGAATTAAACCACATGCTCCACCGCTTGTGCGGGCCCCCGTCAATTCATTTGAGTTTTAACCTTGCGGCCGTACTCCCCAGGCGGTCGACTTAACGCGTTAGCTCCGGAAGCCACTCCTCAAGGGAACAACCTCCAAGTCGACATCGTTTACGGCGTGGACTACCAGGGTATCTAATCCTGTTTGCTCCCCACGCTTTCGCACCTGAGCGTCAGTCTTTGTCCAGGGGGCCGCCTTCGCCACCGGTATTCCTCCAGATCTCTACGCATTTCACCGCTACACCTGGAATTCTACCCCCCTCTACAAGACTCTAGCCTGCCAGTTTCGAATGCAGTTCCCAGGTTGAGCCCGGGGATTTCACATCCGACTTGACAGACCGCCTGCGTGCGCTTTACGCCCAGTAATTCCGATTAACGCTTGCACCCTCCGTATTACCGCGGCTGCTGGCACGGAGTTAGCCGGTGCTTTTTCTGCGAGTAACGTCAATCACTGTGGtTATTAACCACAATGCCTTCCTCCTCGCTGAAAGTACTTTACAaCCCGAAGgCcTtCtTCATACACGCGGCATGGCTGCATCAGgCTTGCGCCCATTGTGCAATATTCCCCACTGCTGCCTCCCGTAGGAGTCTGGACCGTGTCTCAGTTCCAGTGTGGCTGGTCATCCTCTCAGACCAGCTAGGGATCGTCGCCTAGGTGAGCCATTACCCCACCTACTAGCTAATCCCATCTGGGCACATCTGATGGCAAGAGGCCCGAAGGTCCCCCTCTTTGGTCTTGCGACGTTATGCGGTATTAGCTACCGTTTCCAGTAGTTATCCCCCTCCATCAGGCAGTTTCCCAGACATTACTCACCCGTCCGCCGCTCGTCACCCGAGAGCAAGCTCTCTGTGCTACCGCTCGACTGCATGGTAGTCTGCGC |
| Strains Code | Nitrogen Concentration mg/mL | Glucose Concentration g/mL | Nitrogen Fixation Efficiency Value mg/g |
|---|---|---|---|
| WXY-2 | 0.032 | 0.064 | 0.49 ± 0.21 gh |
| DY-5 | 0.126 | 0.049 | 2.57 ± 0.30 cd |
| DY-8 | 0.088 | 0.065 | 1.34 ± 0.18 f |
| LYX-4 | 0.047 | 0.06 | 0.78 ± 0.01 gh |
| WX-2 | 0.066 | 0.022 | 2.97 ± 0.33 c |
| DY-7 | 0.129 | 0.054 | 2.41 ± 0.21 d |
| WXY-1 | 0.030 | 0.064 | 0.47 ± 0.09 gh |
| CK-3 | 0.049 | 0.059 | 0.83 ± 0.06 g |
| TB * | 0.147 | 0.066 | 2.23 ± 0.11 d |
| DY-1 | 0.047 | 0.060 | 0.79 ± 0.07 gh |
| LYX-2 | 0.107 | 0.062 | 1.81 ± 0.14 e |
| WX-5 | 0.317 | 0.037 | 8.49 ± 0.70 a |
| CK-1 | 0.208 | 0.060 | 3.46 ± 0.09 b |
| DY-2 | 0.023 | 0.061 | 0.38 ± 0.07 h |
| WXY-5 | 0.232 | 0.067 | 3.45 ± 0.12 b |
| Treatments | Ack | AS | t (min) | CAT Activity |
|---|---|---|---|---|
| N1 | 0.573 | 0.998 | 1 | 272.448 |
| N2 | 0.576 | 1.226 | 0.5 | 268.145 |
| N3 | 0.575 | 0.947 | 1 | 363.615 |
| N4 | 0.570 | 0.886 | 1 | 281.073 |
| N5 | 0.579 | 0.890 | 1 | 386.264 |
| N6 | 0.570 | 0.967 | 1 | 295.869 |
| N7 | 0.579 | 0.979 | 0.5 | 277.243 |
| CK | 0.576 | 1.221 | 0.5 | 187.301 |
| Treatments | SOD Activity | POD Activity | CAT Activity |
|---|---|---|---|
| N1 | 240.893 ± 22.626 c | 191.156 ± 14.512 d | 272.448 ± 9.830 d |
| N2 | 165.450 ± 27.354 e | 175.136 ± 17.594 d | 268.145 ± 17.285 d |
| N3 | 258.581 ± 20.496 c | 302.837 ± 26.645 c | 363.615 ± 10.146 b |
| N4 | 213.118 ± 8.238 d | 366.403 ± 32.585 b | 281.073 ± 7.611 cd |
| N5 | 376.226 ± 25.008 c | 429.907 ± 13.612 a | 386.264 ± 26.560 a |
| N6 | 284.913 ± 10.039 b | 350.404 ± 26.323 b | 295.869 ± 22.913 c |
| N7 | 243.979 ± 11.109 c | 302.666 ± 26.383 c | 277.243 ± 6.843 cd |
| CK | 156.072 ± 21.728 e | 164.975 ± 8.209 d | 187.301 ± 10.351 e |
| Treatments | SOD Gene | POD Gene | CAT Gene |
|---|---|---|---|
| N1 | 2.911 ± 0.530 bcd | 3.349 ± 0.914 d | 4.724 ± 0.141 d |
| N2 | 1.712 ± 0.277 de | 1.963 ± 0.092 e | 4.048 ± 0.122 d |
| N3 | 3.668 ± 0.217 bc | 4.630 ± 0.065 cd | 6.781 ± 1.075 b |
| N4 | 2.690 ± 0.044 cd | 7.335 ± 0.766 b | 4.899 ± 0.360 cd |
| N5 | 5.722 ± 1.050 a | 9.212 ± 1.184 a | 8.794 ± 0.844 a |
| N6 | 4.009 ± 0.589 b | 6.765 ± 0.635 b | 5.911 ± 0.637 bc |
| N7 | 3.491 ± 1.076 bc | 5.400 ± 0.451 c | 4.643 ± 0.390 d |
| CK | 1.100 ± 0.088 e | 0.973 ± 0.156 e | 1.032 ± 0.030 e |
References
- Zhou, Q.; Lei, Q.; Zhao, J.; Fang, Q.; Fang, S. Identification and Quality Research on Fritillariae Cirrhosae Bulbus (Cultivation) of Sichuan Dao-di Herbs. World Chin. Med. 2020, 15, 225–230. [Google Scholar] [CrossRef]
- Luo, M.; Deng, C.; Li, P.; Tan, Q.; Luo, S.; Xu, G.; Zhang, W. Research progress in Medicinal Plant Fritillaria taipaiensis P. Y. Li. Chin. Wild Pl. Resour. 2021, 40, 42–45+56. [Google Scholar] [CrossRef]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China; China Medical Science and Technology Press: Beijing, China, 2020; pp. 38–39.
- Zou, M.; Jiang, R.; Li, J.; Duan, Q.; Liao, H.; Zhou, J. Prediction of potential distribution of Fritillaria taibaiensis based on MaxEnt model. Chin. J. Inf. Tradit. Chin. Med. 2021, 28, 1–5. [Google Scholar]
- Liu, X. Comparative introduction experiment of Fritillaria cirrhosa and Fritillaria taibaiensis. China J. Chin. Mater. Med. 1994, 81–82. [Google Scholar]
- Tian, Q.; Qiang, Y.; Chen, K.; Wang, J. Advance of Valuable Endangered Medicinal Plant Fritillaria taibaiensis. Shaanxi J. Agri. Sci. 2018, 64, 96–98. [Google Scholar]
- Duan, B.; Chen, X.; Huang, L.; Lu, Q.; Li, X.; Chen, S. Overview of Fritillaria taibaiensis resource study. Mod. Chin. Med. 2010, 12, 12–14. [Google Scholar]
- Ministry of Agriculture and Rural Affairs. Action Plan for Chemical Fertilizer Reduction by 2025; Ministry of Agriculture and Rural Affairs: Beijing, China, 16 November 2022.
- Zheng, J.; Li, M.; Shi, F.; Dong, F. Effects of Reducing Chemical Fertilizer and Applying Organic and Microbial Fertilizers on Tomato Photosynthetic Characteristics and Fertilizer Use Efficiency. China Cucurbits Veg. 2024, 37, 74–79. [Google Scholar] [CrossRef]
- Luo, Y.; Zhou, M.; Man, B.; Chai, S.; He, Y. Status quo of microbial fertilizer in China and application in Chinese medicinal herb cultivation. Chin. J. Soil Fertil. Sci. 2024, 11, 222–230. [Google Scholar]
- Meng, C.; Zhao, Y.; Chen, J.; Zhang, Y.; Wang, Y.; Feng, L.; Sun, Y.; Guo, C. Screening and identification of two strains of nitrogen-fixing bacteria from the silage maize rhizosphere and their roles in plant growth promotion. Acta Prataculturae Sin. 2024, 33, 174–185. [Google Scholar]
- Wang, X.; Cao, Y.; Tang, X.; Ma, X.; Gao, J.; Zhang, X. Rice endogenous nitrogen fixing and growth promoting bacterium Herbaspirillum seropedicae DX35. Acta Microbiol. Sin. 2014, 54, 292–298. [Google Scholar] [CrossRef]
- Hu, C.; Lin, L.; Shi, G.; Wang, Q.; Wang, Q.; Li, Y. Screening and identification of associative nitrogen fixation bacteria in rhizosphere of sugarcane in Guangxi. Acta Ecol. Sin. 2012, 32, 4745–4752. [Google Scholar]
- Wei, Z.; Pan, X.; Huang, X.; Li, H.; Guo, D.; Zhou, N. Effects of inoculation of Fritillaria Taipaiensis P.Y. Li with growth-promoting bacteria on inorganic elements in rhizosphere soil. Environ. Chem. 2021, 40, 1254–1262. [Google Scholar] [CrossRef]
- Quan, Z.; Zhang, X.; Davidson, E.A.; Zhu, F.; Li, S.; Zhao, X.; Chen, X.; Zhang, L.; He, J.; Wei, W.; et al. Fates and Use Efficiency of Nitrogen Fertilizer in Maize Cropping Systems and Their Responses to Technologies and Management Practices: A Global Analysis on Field 15N Tracer Studies. Earth’s Future 2021, 9, e2020EF001514. [Google Scholar] [CrossRef]
- Herridge, D.; Peoples, M.; Boddey, R. Global inputs of biological nitrogen fixation in agricultural systems. Plant Soil 2008, 311, 1–18. [Google Scholar] [CrossRef]
- Lu, Y.; Zhao, Q.; Peng, X. Isolation and identification of 2 strains of nitrogen-fixing bacteria. Jiangsu Agric. Sci. 2020, 48, 298–302. [Google Scholar]
- Li, S.; Sun, Y.; Zhao, J.; Liu, X.; Zhao, J.; Ma, C.; Zhang, Q. Effects of Nitrogen Addition on Microbial Quantity, Enzyme Activity in Rhizosphere Soil and Alfalfa Hay Yield. Chin. J. Grassl. 2018, 24, 38–44. [Google Scholar]
- Tang, X.; Jiang, J.; Jin, H.; Zhou, C.; Liu, G.; Yang, H. Effects of shading on chlorophyll content and photosynthetic characteristics in leaves of Phoebe bournei. Chin. J. Appl. Ecol. 2019, 30, 2941–2948. [Google Scholar]
- Negi, S.; Barry, A.N.; Friedland, N.; Sudasinghe, N.; Subramanian, S.; Pieris, S.; Holguin, F.O.; Dungan, B.; Schaub, T.; Sayre, R. Impact of nitrogen limitation on biomass, photosynthesis, and lipid accumulation in Chlorella sorokiniana. J. Appl. Phycol. 2016, 28, 803–812. [Google Scholar] [CrossRef]
- Xu, T.; Lv, T.; Zhang, Y.; Liu, H.; Liu, Y.; Cai, W.; Zhang, R.; Song, W.; Xing, J.; Zhao, J.; et al. Effects of high-temperature stress on photosynthetic characteristics, protective enzyme activity, and yield of maize hybrids along with their parental inbred lines. Chin. J. Eco-Agric. 2024, 32, 1470–1480. [Google Scholar]
- Kumari, K.; Samantaray, S.; Sahoo, D.; Tripathy, B.C. Nitrogen, phosphorus and high CO2 modulate photosynthesis, biomass and lipid production in the green alga Chlorella vulgaris. Photosynth. Res. 2021, 148, 17–32. [Google Scholar] [CrossRef]
- Domonkos, I.; Kis, M.; Gombos, Z.; Ughy, B. Carotenoids, versatile components of oxygenic photosynthesis. Prog. Lipid Res. 2013, 52, 539–561. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Yang, Y.; Li, X.; Lu, J.; Wang, L.; Liu, D. Effect of Various Abiotic Stresses on Growth and Physiological Characteristics of Alfalfa (Medicago sativa L.) WL712 Seedlings. Chin. J. Grassland. 2024, 46, 78–86. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, H.; Jiang, S.; Sun, H.; Yang, P.; Du, J.; Sun, H.; Zhou, Y. Effect of Different Nitrogen Concentrations on Growth and Antioxidant Enzymes Activity in Notopterygium incisum Seedling in Cultivation Matrix. Chin. J. Exp. Tradit. Med. Formulae 2018, 24, 38–44. [Google Scholar]
- Xie, J.; Li, M.; Shi, M.; Kang, C.; Zhang, R.; Liu, Y.; Qing, S.; Zhang, W. Effects of Potassium Fertilizer on Physiological Characteristics, Yield and Quality of Potato under Drought Stress. Acta Agric. Boreali-Occident. Sin. 2024, 33, 2059–2069. [Google Scholar]
- Zhan, Z.; Gou, X.; Wang, Z.; Zhang, Y.; Wang, X.; Guo, J. Effects of Seed Soaking with Beauveria bassiana and Metarhizium rileyi on Defense Enzyme Activities and Insect Resistance in Maize Leaves. Chin. J. Biol. Control 2025, 41, 54–62. [Google Scholar] [CrossRef]
- Zhou, X.; Yang, Y.; Hu, P.; Xia, Y. Investigation on the Resources of Fritillaria taipaiensis. J. Anhui Agric. Sci. 2015, 17, 84–85. [Google Scholar] [CrossRef]
- Fu, S.; Chen, H.; Yuan, D.; Jia, H.; Pu, C.; Lu, Q. Investigation of Chongqing Fritillaria taipaiensis Resources. Chin. J. Inf. Tradit. Chin. Med. 2016, 9, 1–4. [Google Scholar]
- Jiang, Y.; Wu, Y.; Wang, G.; Xu, W.; Zhang, Z.; Xu, L.; Hu, F.; Li, H. Plant Growth-promoting Bacterium Variovorax sp. JX14 from Calcareous Alluvial Soil: Characterization and Growth Promotion on Peanuts. Soil 2015, 47, 698–703. [Google Scholar] [CrossRef]
- Wang, B. Screening of Functional Bacteria from Rhizosphere Soil of Allium Plants in Inner Mongolia Grassland and Their Role in the Groeth of Oats. Master’s Thesis, Inner Mongolia University, Hohhot, China, 2021. [Google Scholar] [CrossRef]
- Cai, M. Isolation and Characterization of Associative Nitrogen-Fixing Bacteria in the Rhizosphere of Camellia oleifera. Master’s Thesis, Nanjing Forestry University, Nanjing, China, 2011. [Google Scholar]
- Ye, M. Isolation and Screening of Nitrogen-Fixing Bacteria from Fritillaria taibaiensis and Its Nitrogen-Fixing Effect. Master’s Thesis, Chongqing Three Gorges University, Chongqing, China, 2024. [Google Scholar] [CrossRef]
- Ma, N.; Liu, Y.; Li, X.; Chen, X.; Yang, X. Comprehensive Evaluation of Growth Promotion Effect of PGPR Strains on Tobacco Based on Entropy Weight Method. J. Agric. Univ. 2020, 43, 887–895. [Google Scholar]
- Kandiannan, K.; Parthasarathy, U.; Krishnamurthy, K.S.; Thankamani, C.K.; Srinivasan, V. Modeling individual leaf area of ginger (Zingiber officinale Roscoe) using leaf length and width. Sci. Hortic. 2009, 120, 532–537. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Sharkey, T.D. Stomatal Conductance and Photosynthesis. Annu. Rev. Plant Physiol. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Zhang, Z.; Qu, W.; Li, X. Experimental Guide to Plant Physiology; Higher Education Press: Beijing, China, 2009; pp. 78–88. [Google Scholar]
- Bai, B.; Tang, X. Testing Techniques in Plant Physiology; China Science and Technology Press: Beijing, China, 1993. [Google Scholar]
- Zou, Q. Guidance on Plant Physiology; China Agriculture Press: Beijing, China, 2003. [Google Scholar]
- Zhang, Z. Experimental Guidance on Plant Physiology Techniques, 2nd ed.; Higher Education Press: Beijing, China, 1990. [Google Scholar]
- Ge, K.; Wang, P.J.; Shao, H.L.; Chen, D.L.; Du, B. Physiological characteristics of heavy metal accumulation and resistance in leaves of typical urban greening tree species. J. Shanxi Agric. Univ. Nat. Sci. Ed. 2022, 42, 96–104. [Google Scholar]
- Wang, Y.H.; Yuan, L.; Wang, Y.H.; Lang, J.Q.; Ye, M.Y.; Liu, Q.Q.; Ma, Q.; Zhou, N. Phosphorus-solubilizing fungi promote the growth of Fritillaria taipaiensis P. Y. Li by regulating physiological and biochemical reactions and protecting enzyme system–related gene expression. Front. Genet. 2025, 15, 1459191. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chen, J.Q.; Zhao, G.Y.; Wei, Y.H.; Dong, Y.H.; Hou, L.Y.; Jiao, R.Z. Isolation and screening of multifunctional phosphate solubilizing bacteria and its growth-promoting effect on Chinese fir seedlings. Sci. Rep. 2021, 11, 9081. [Google Scholar] [CrossRef]
- Parmar, P.; Sindhu, S.S. The novel and efficient method for isolating potassium solubilizing bacteria from rhizosphere soil. Geomicrobiol. J. 2019, 36, 130–136. [Google Scholar] [CrossRef]
- Ke, X.; Feng, S.; Wang, J.; Lu, W.; Zhang, W.; Chen, M.; Lin, M. Effect of inoculation with nitrogen-fixing bacterium Pseudomonas stutzeri A1501 on maize plant growth and the microbiome indigenous to the rhizosphere. Syst. Appl. Microbiol. 2019, 42, 248–260. [Google Scholar] [CrossRef]
- Li, Y.; Li, Y.; Zhang, H.; Wang, M.; Chen, S. Diazotrophic Paenibacillus beijingensis BJ-18 Provides Nitrogen for Plant and Promotes Plant Growth, Nitrogen Uptake and Metabolism. Front. Microbiol. 2019, 10, 1119. [Google Scholar] [CrossRef]
- Ai, C.; Zhao, Y.; Zhang, L.; Zhang, M.; Huang, S.; Wang, S.; Zhou, W. Research progress on associative nitrogen fixation of gramineous crops. J. Plant Nutr. Fert. 2024, 34, 1307–1321. [Google Scholar]
- Luo, J.; Mu, M.; Wang, Q.; Wen, M.; Zhu, F.; Wu, X.; Zhou, N. Effect of Different Nitrogen Forms and Concentrations on Yield and Quality of Fritillaria thunbergii. Chin. J. Exp. Tradit. Med. Formulae 2020, 26, 168–174. [Google Scholar]
- Li, Z.; Luo, L.; Lang, J.; Ye, M.; Yin, F.; Zhao, J.; Zhou, N. Effect of Organophosphate-Solubilizing Bacteria on Photosynthesis, Physiology, and Biochemistry of Paris polyphylla var. yunnanensis. Chin. J. Exp. Tradit. Med. Formulae 2022, 28, 165–171. [Google Scholar]
- Zhang, S.; Ding, D.; He, J.; Wang, X.; Ma, D.; Deng, Y.; Zhang, X. Effects of Two Biofertilizer Mixtures on Growth and Photosynthetic Characteristics of replanted Zanthoxylum bungeanum. Acta Agric. Boreali-Occident. Sin. 2021, 30, 1355–1364. [Google Scholar]
- Zhao, X.; Zhang, X.; Liu, Z.; Lv, Y.; Song, T.; Cui, J.; Chen, T.; Li, J.; Zeng, F.; Zhan, Y. Comparing the Effects of N and P Deficiency on Physiology and Growth for Fast- and Slow-Growing Provenances of Fraxinus mandshurica. Forests 2021, 12, 1760. [Google Scholar] [CrossRef]
- Cheng, B.; Ma, H.; Yang, T.; Cheng, Z.; Niu, X.; Ma, X. Effect of Different Microbial Fertilizer Dosage on the Elements of Fritillaria pallidiflora Schvek. Xinjiang Agric. Sci. 2017, 54, 871–877. [Google Scholar]
- Zhao, J.; Liang, S.; Yang, T.; Wu, X.; Qi, P.; Wang, S.; Wang, Z. Isolation of Growth-Promoting Bacteria and Effect of Compound Bacteria on Yield of Fritillaria przewalskii. Chin. J. Exp. Tradit. 2021, 27, 163–170. [Google Scholar]
- Zhang, J.; Wang, X.; Feng, B.; Zhou, X.; Chen, H.; Pan, S.; Wu, C. Effects of Staggered Planting and Increasing Bacterial Fertilizer on Photosynthetic Characteristics, Yield and Quality of Fritillaria taipaiensis P. Y. Li. Acta Agric. Boreali-Occident. Sin. 2022, 31, 628–639. [Google Scholar]
- Mu, H.; Lin, L.; Yu, J.; Li, S.; Jiang, X.; Xia, F. Photosynthesis of Autotetraploid Betula platyphylla Seedling Responding to NaHCO3 Stress. J. Beihua Univ. Nat. Sci. Ed. 2017, 18, 25–29. [Google Scholar]
- Pei, Z.; Zhou, J.; Xu, X.; Lan, H.; Wang, J.; Lang, S.; Zhang, W. Effects of Drought Treatment on Photosynthesis Rate, Antioxidant Properties of Leaves and Yield of Different Maize Varieties. Crops 2021, 37, 95–100. [Google Scholar]
- Wu, K.; Liang, J.; Wei, L.; Luo, T.; Xing, Y.; Li, Y.; Yang, L. Effects of nitrogen fixing bacteria on photosynthesis and chlorophyll fluorescence in sugarcane at elongating stage. Guihaia 2011, 31, 668–673. [Google Scholar]
- Zheng, S.; Guo, S.; Zhang, Y.; Song, X.; Fang, C.; Zhang, J.; Sun, J. Effects of Arbuscular Mycorrhizal Fungi on Characteristics of Photosynthesis, Microbial Diversity and Enzymes Activity in Rhizosphere of Pepper Plants Cultivated in Organic Substrate. Acta Bot. Boreali-Occident. Sin. 2014, 34, 800–809. [Google Scholar] [CrossRef]
- Tian, F.; Chen, X.; Wang, P.; Zhong, L.; Ou, E. Effect of microbial agents on resistant enzyme system and biochemical indexes of drought resistance in Sophora david. Plant Physiol. J. 2022, 58, 435–446. [Google Scholar]
- Zhang, H.; Zhao, G.; Li, M.; Xia, C.; Wang, C. Physiological responses of Pennisetum longissimum var. intermediumseedlings to PEG, low temperature and salt stress treatments. Acta Prataculturae Sin. 2014, 23, 180–188. [Google Scholar] [CrossRef]
- Yan, M.; Yao, Y.; Mu, K.; Dan, Y.; Li, W.; Liao, W. Involvement of abscisic acid in hydrogen gas-enhanced drought resistance by improving antioxidant enzyme activity and gene expression in tomato seedlings. Acta Agric. Zhejiangensis 2022, 34, 1901–1910. [Google Scholar] [CrossRef]
- Zhao, H.; Ren, Q.; Wang, X.; Li, Z.; Tang, X.; Jiang, L.; Liu, P.; Xing, C. Effects of arbuscular mycorrhizal fungi on antioxidant enzymes activities and photosynthetic characteristics of Solanum lycopersicum L.under salt stress. Acta Agric. Zhejiangensis 2021, 33, 2075–2084. [Google Scholar] [CrossRef]



| Treatments | Inoculated Strain | Inoculation Measurement | Pots |
|---|---|---|---|
| N1 | Rahnella aquatilis | 120 mL/strain | 10 |
| N2 | Pseudomonas chlororaphis | 120 mL/strain | 10 |
| N3 | Paenibacillus stellifer | 120 mL/strain | 10 |
| N4 | R. Aquatilis, P. chlororaphis | 60 mL/strain | 10 |
| N5 | R. Aquatilis, P. stellifer | 60 mL/strain | 10 |
| N6 | P. Chlororaphis, P. stellifer | 60 mL/strain | 10 |
| N7 | R. Aquatilis, P. chlororaphis, P. stellifer | 40 mL/strain | 10 |
| CK | Replace with sterile water | 120 mL sterile water | 10 |
| Genes | Forward /Reverse | Sequences | Product Size (bp) | Tm/°C |
|---|---|---|---|---|
| SOD | F: | TTCAGTTTCTTAGTGACAATAGGCG | 195 | 58.80 |
| R: | GGTCTTAGTCTGGATACGGCAA | 60.30 | ||
| POD | F: | TTTCCTTTCCATTCACCCG | 175 | 58.20 |
| R: | AAGACCCTTCCCTTTGTTCG | 58.40 | ||
| CAT | F: | TATTCCACAACAACGAAAGCAC | 183 | 58.10 |
| R: | GGACCCGAATCCGTTAGTATG | 58.10 | ||
| rpl16 | F: | TTCGTGCTACATTCGTAGGGTC | 190 | 59.60 |
| R: | GTTCCATTGCGGAGTTCGG | 61.00 |
| Strains Code | Nitrogen Fixation Efficiency Value (g/kg) | Increased Indole-3-Acetic Acid Content (µg/mL) | Comprehensive Score |
|---|---|---|---|
| WXY-2 | 0.49 ± 0.21 gh | 5.85 ± 0.14 g | 0.113 |
| DY-5 | 2.57 ± 0.30 cd | 10.12 ± 0.18 c | 0.494 |
| DY-8 | 1.34 ± 0.18 f | 8.74 ± 0.23 d | 0.559 |
| LYX-4 | 0.78 ± 0.01 gh | 6.26 ± 0.18 f | 0.228 |
| WX-2 | 2.97 ± 0.33 c | 3.70 ± 0.06 i | 0.298 |
| DY-7 | 2.41 ± 0.21 d | 2.13 ± 0.14 kl | 0.101 |
| WXY-1 | 0.47 ± 0.09 gh | 6.38 ± 0.14 f | 0.493 |
| CK-3 | 0.83 ± 0.06 g | 1.27 ± 0.18 m | 0.026 |
| TB * | 2.23 ± 0.11 d | 2.29 ± 0.16 j | 0.470 |
| DY-1 | 0.79 ± 0.07 gh | 20.67 ± 0.20 a | 0.265 |
| LYX-2 | 1.81 ± 0.14 e | 0.69 ± 0.16 n | 0.059 |
| WX-5 | 8.49 ± 0.70 a | 7.42 ± 0.21 e | 0.604 |
| CK-1 | 3.46 ± 0.09 b | 1.94 ± 0.14 l | 0.142 |
| DY-2 | 0.38 ± 0.07 h | 5.11 ± 0.16 h | 0.055 |
| WXY-5 | 3.45 ± 0.12 b | 14.93 ± 0.08 b | 0.457 |
| Treatments | Leaf Area (cm2) | Leaf Thickness (mm) | Stem Diameter (mm) | Plant Height (cm) |
|---|---|---|---|---|
| N1 | 1.175 ± 0.188 c | 0.462 ± 0.081 b | 1.167 ± 0.114 ab | 5.488 ± 0.406 b |
| N2 | 1.172 ± 0.235 c | 0.512 ± 0.044 ab | 1.040 ± 0.093 bc | 5.432 ± 0.389 b |
| N3 | 1.163 ± 0.235 c | 0.527 ± 0.058 ab | 1.077 ± 0.147 abc | 5.425 ± 0.698 b |
| N4 | 1.576 ± 0.274 b | 0.510 ± 0.042 ab | 1.203 ± 0.131 ab | 6.457 ± 0.410 a |
| N5 | 1.867 ± 0.252 a | 0.523 ± 0.048 ab | 1.223 ± 0.152 a | 7.082 ± 0.744 a |
| N6 | 1.404 ± 0.198 bc | 0.575 ± 0.039 a | 1.242 ± 0.104 a | 6.587 ± 0.772 a |
| N7 | 1.225 ± 0.196 c | 0.547 ± 0.052 a | 1.252 ± 0.145 a | 4.923 ± 0.403 b |
| CK | 0.726 ± 0.068 d | 0.293 ± 0.062 c | 0.967 ± 0.164 c | 4.083 ± 0.485 c |
| Treatments | Net Photosynthetic Rate (Pn) Mmol/(m2·s) | Stomatal Conductance (Cond) μmol/(m2·s) | Intercellular CO2 Concentration (Ci) μmol/(m2·s) | Transpiration Rate (Tr) Mmol/(m2·s) | Stomatal Limitation (LS) mmol/mol |
|---|---|---|---|---|---|
| N1 | 10.108 ± 0.069 g | 0.073 ± 0.003 e | 148.167 ± 3.710 e | 1.514 ± 0.008 f | 0.630 ± 0.009 b |
| N2 | 11.211 ± 0.008 d | 0.069 ± 0.006 ef | 146.000 ± 2.098 e | 1.459 ± 0.006 g | 0.661 ± 0.005 a |
| N3 | 10.647 ± 0.233 f | 0.079 ± 0.004 d | 165.333 ± 2.805 c | 1.608 ± 0.074 d | 0.587 ± 0.007 d |
| N4 | 12.459 ± 0.071 c | 0.084 ± 0.004 c | 169.667 ± 3.724 b | 1.775 ± 0.006 c | 0.576 ± 0.009 e |
| N5 | 12.108 ± 0.007 a | 0.110 ± 0.005 a | 199.500 ± 3.507 a | 1.970 ± 0.006 a | 0.502 ± 0.009 f |
| N6 | 11.703 ± 0.007 b | 0.095 ± 0.002 b | 157.167 ± 2.639 d | 1.840 ± 0.004 b | 0.607 ± 0.006 c |
| N7 | 10.915 ± 0.016 e | 0.079 ± 0.003 d | 159.667 ± 3.386 d | 1.561 ± 0.003 e | 0.601 ± 0.008 c |
| CK | 9.326 ± 0.019 h | 0.068 ± 0.005 f | 135.833 ± 2.229 f | 1.388 ± 0.005 h | 0.635 ± 0.005 b |
| Treatments | Chlorophyll a (mg/g) | Chlorophyll b (mg/g) | Carotenoid (mg/g) | Chlorophyll a/b | Total Chlorophyll (mg/g) |
|---|---|---|---|---|---|
| N1 | 0.996 ± 0.034 a | 0.344 ± 0.008 a | 0.304 ± 0.012 a | 2.900 ± 0.084 cd | 1.339 ± 0.039 ab |
| N2 | 0.965 ± 0.041 b | 0.339 ± 0.020 ab | 0.278 ± 0.037 b | 2.847 ± 0.095 cd | 1.304 ± 0.059 b |
| N3 | 0.931 ± 0.011 c | 0.305 ± 0.006 c | 0.262 ± 0.038 bc | 3.049 ± 0.052 bc | 1.236 ± 0.014 c |
| N4 | 0.819 ± 0.017 e | 0.304 ± 0.016 c | 0.241 ± 0.013 cd | 2.702 ± 0.155 d | 1.124 ± 0.023 d |
| N5 | 1.021 ± 0.006 a | 0.351 ± 0.004 a | 0.307 ± 0.004 a | 2.912 ± 0.031 cd | 1.372 ± 0.008 a |
| N6 | 1.017 ± 0.032 a | 0.325 ± 0.021 b | 0.278 ± 0.006 d | 3.146 ± 0.279 b | 1.342 ± 0.032 ab |
| N7 | 0.864 ± 0.005 d | 0.286 ± 0.007 d | 0.236 ± 0.003 b | 3.019 ± 0.063 bc | 1.150 ± 0.010 d |
| CK | 0.923 ± 0.031 c | 0.235 ± 0.019 e | 0.305 ± 0.004 a | 3.951 ± 0.386 a | 1.158 ± 0.030 d |
| Treatments | MDA mmol/g | Soluble Sugar mmol/g | Soluble Protein mg/g | Proline mg/g |
|---|---|---|---|---|
| N1 | 0.029 ± 0.001 b | 0.596 ± 0.009 c | 14.475 ± 0.385 g | 0.784 ± 0.005 b |
| N2 | 0.023 ± 0.001 de | 0.492 ± 0.004 g | 18.115 ± 0.458 d | 0.594 ± 0.005 f |
| N3 | 0.022 ± 0.001 de | 0.689 ± 0.008 a | 21.263 ± 0.608 c | 0.767 ± 0.058 c |
| N4 | 0.029 ± 0.002 b | 0.430 ± 0.008 h | 22.603 ± 0.388 b | 0.750 ± 0.006 c |
| N5 | 0.021 ± 0.002 e | 0.557 ± 0.010 d | 28.148 ± 0.357 a | 0.694 ± 0.008 d |
| N6 | 0.024 ± 0.001 d | 0.514 ± 0.008 e | 17.072 ± 0.330 e | 0.657 ± 0.004 e |
| N7 | 0.027 ± 0.001 c | 0.654 ± 0.011 b | 15.003 ± 0.374 f | 0.944 ± 0.007 a |
| CK | 0.034 ± 0.001 a | 0.503 ± 0.008 f | 13.187 ± 0.245 h | 0.550 ± 0.005 g |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, M.; Lang, J.; Kong, X.; Shi, Z.; Duan, F.; Qin, G.; Zhang, H.; Guo, D.; Zhou, N. Nitrogen-Fixing Bacteria Promote the Growth of Fritillaria taipaiensis P. Y. Li by Regulating Physiological and Biochemical Reactions and Protecting Enzyme System-Related Gene Expression. Biology 2025, 14, 325. https://doi.org/10.3390/biology14040325
Ye M, Lang J, Kong X, Shi Z, Duan F, Qin G, Zhang H, Guo D, Zhou N. Nitrogen-Fixing Bacteria Promote the Growth of Fritillaria taipaiensis P. Y. Li by Regulating Physiological and Biochemical Reactions and Protecting Enzyme System-Related Gene Expression. Biology. 2025; 14(4):325. https://doi.org/10.3390/biology14040325
Chicago/Turabian StyleYe, Mingyan, Jiaqi Lang, Xiaotian Kong, Zhifen Shi, Fengjie Duan, Guiyong Qin, Hua Zhang, Dongqin Guo, and Nong Zhou. 2025. "Nitrogen-Fixing Bacteria Promote the Growth of Fritillaria taipaiensis P. Y. Li by Regulating Physiological and Biochemical Reactions and Protecting Enzyme System-Related Gene Expression" Biology 14, no. 4: 325. https://doi.org/10.3390/biology14040325
APA StyleYe, M., Lang, J., Kong, X., Shi, Z., Duan, F., Qin, G., Zhang, H., Guo, D., & Zhou, N. (2025). Nitrogen-Fixing Bacteria Promote the Growth of Fritillaria taipaiensis P. Y. Li by Regulating Physiological and Biochemical Reactions and Protecting Enzyme System-Related Gene Expression. Biology, 14(4), 325. https://doi.org/10.3390/biology14040325