
Influence of Naturally Occurring Bacteria on Embryonic and Larval Development of Common Toad Tadpoles




Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Isolation and Characterization of Bacteria
2.2. Experimental Design
2.3. Molecular Biomarkers and Energy Budget Analyses
2.4. Statistical Analysis
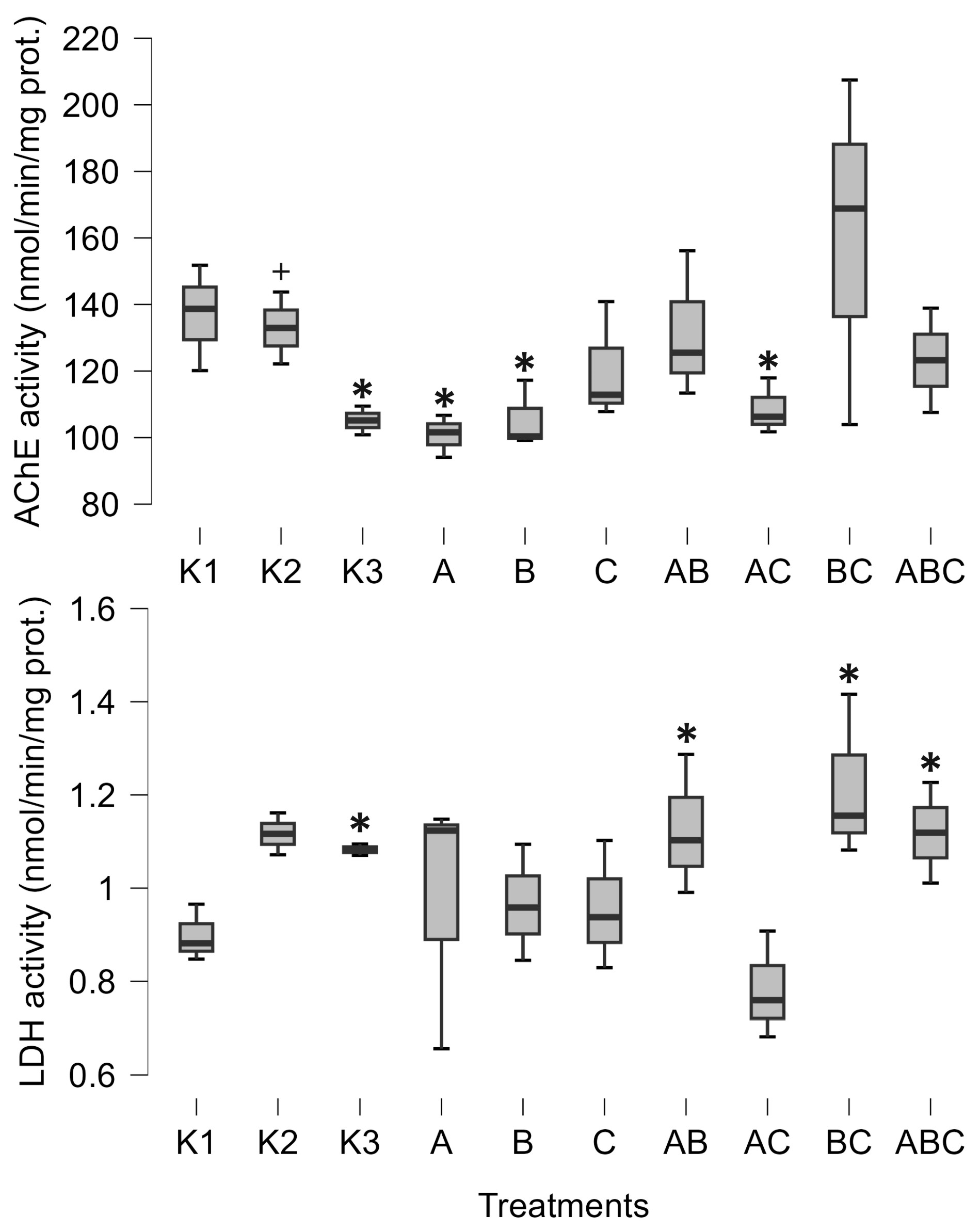
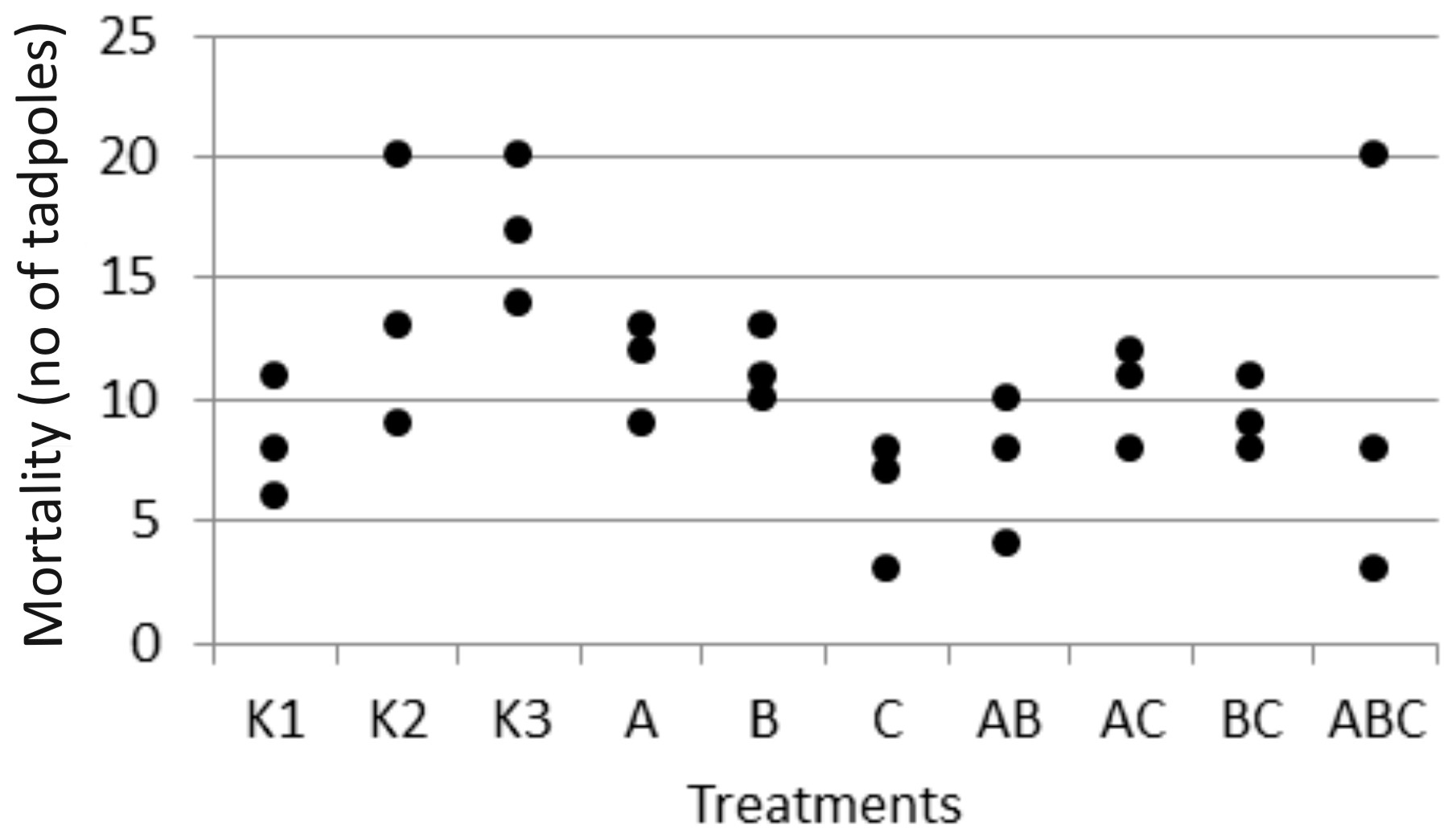
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dobrzyński, J.; Jakubowska, Z.; Dybek, B. Potential of Bacillus pumilus to Directly Promote Plant Growth. Front. Microbiol. 2022, 13, 1069053. [Google Scholar] [CrossRef] [PubMed]
- Manoj, R.R.S.; Latrofa, M.S.; Epis, S.; Otranto, D. Wolbachia: Endosymbiont of Onchocercid Nematodes and Their Vectors. Parasit. Vectors 2021, 14, 245. [Google Scholar] [CrossRef] [PubMed]
- Hoye, B.J.; Fenton, A. Animal Host–Microbe Interactions. J. Anim. Ecol. 2018, 87, 315–319. [Google Scholar] [CrossRef]
- McFall-Ngai, M.; Hadfield, M.G.; Bosch, T.C.G.; Carey, H.V.; Domazet-Lošo, T.; Douglas, A.E.; Dubilier, N.; Eberl, G.; Fukami, T.; Gilbert, S.F.; et al. Animals in a Bacterial World, a New Imperative for the Life Sciences. Proc. Natl. Acad. Sci. USA 2013, 110, 3229–3236. [Google Scholar] [CrossRef]
- Scalvenzi, T.; Clavereau, I.; Bourge, M.; Pollet, N. Gut microbial ecology of Xenopus tadpoles across life stages. Peer Community J. 2021, 1, e41. [Google Scholar] [CrossRef]
- Hou, K.; Wu, Z.-X.; Chen, X.-Y.; Wang, J.-Q.; Zhang, D.; Xiao, C.; Zhu, D.; Koya, J.B.; Wei, L.; Li, J.; et al. Microbiota in Health and Diseases. Sig Transduct. Target. Ther. 2022, 7, 1–28. [Google Scholar] [CrossRef]
- Hadfield, M.G. Biofilms and Marine Invertebrate Larvae: What Bacteria Produce That Larvae Use to Choose Settlement Sites. Annu. Rev. Mar. Sci. 2011, 3, 453–470. [Google Scholar] [CrossRef]
- Brühl, C.A.; Schmidt, T.; Pieper, S.; Alscher, A. Terrestrial Pesticide Exposure of Amphibians: An Underestimated Cause of Global Decline? Sci. Rep. 2013, 3, 1135. [Google Scholar] [CrossRef]
- Jovanović Glavaš, O.; Stjepanović, N.; Hackenberger, B.K. Influence of Nano and Bulk Copper on Agile Frog Development. Ecotoxicology 2022, 31, 357–365. [Google Scholar] [CrossRef]
- Karaoğlu, K.; Gül, S. Characterization of Microplastic Pollution in Tadpoles Living in Small Water-Bodies from Rize, the Northeast of Turkey. Chemosphere 2020, 255, 126915. [Google Scholar] [CrossRef]
- Jones, K.R.; Hughey, M.C.; Belden, L.K. Colonization Order of Bacterial Isolates on Treefrog Embryos Impacts Microbiome Structure in Tadpoles. Proc. Biol. Sci. 2023, 290, 20230308. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-K.; Park, W.-B.; Do, Y. Tadpole Growth Rates and Gut Bacterial Community: Dominance of Developmental Stages over Temperature Variations. PLoS ONE 2023, 18, e0292521. [Google Scholar] [CrossRef] [PubMed]
- Santos, B.; Martins, F.M.S.; Sabino-Pinto, J.; Licata, F.; Crottini, A. Skin and Gut Microbiomes of Tadpoles Vary Differently with Host and Water Environment: A Short-Term Experiment Using 16S Metabarcoding. Sci. Rep. 2023, 13, 16321. [Google Scholar] [CrossRef]
- Villatoro-Castañeda, M.; Forsburg, Z.R.; Ortiz, W.; Fritts, S.R.; Gabor, C.R.; Carlos-Shanley, C. Exposure to Roundup and Antibiotics Alters Gut Microbial Communities, Growth, and Behavior in Rana Berlandieri Tadpoles. Biology 2023, 12, 1171. [Google Scholar] [CrossRef]
- Bishop, T.F.; Beck, C.W. Bacterial Lipopolysaccharides Can Initiate Regeneration of the Xenopus Tadpole Tail. iScience 2021, 24, 103281. [Google Scholar] [CrossRef]
- Saxena, A.K.; Kumar, M.; Chakdar, H.; Anuroopa, N.; Bagyaraj, D.J. Bacillus Species in Soil as a Natural Resource for Plant Health and Nutrition. J. Appl. Microbiol. 2020, 128, 1583–1594. [Google Scholar] [CrossRef]
- Thomas, P. Isolation of Bacillus pumilus from in Vitro Grapes as a Long-Term Alcohol-Surviving and Rhizogenesis Inducing Covert Endophyte. J. Appl. Microbiol. 2004, 97, 114–123. [Google Scholar] [CrossRef]
- Sari, E.; Etebarian, H.R.; Aminian, H. The Effects of Bacillus pumilus, Isolated from Wheat Rhizosphere, on Resistance in Wheat Seedling Roots against the Take-All Fungus, Gaeumannomyces graminis var. tritici. J. Phytopathol. 2007, 155, 720–727. [Google Scholar] [CrossRef]
- Hill, J.E.; Baiano, J.C.F.; Barnes, A.C. Isolation of a Novel Strain of Bacillus pumilus from Penaeid Shrimp That Is Inhibitory against Marine Pathogens. J. Fish. Dis. 2009, 32, 1007–1016. [Google Scholar] [CrossRef]
- Elshaghabee, F.M.F.; Rokana, N.; Gulhane, R.D.; Sharma, C.; Panwar, H. Bacillus as Potential Probiotics: Status, Concerns, and Future Perspectives. Front. Microbiol. 2017, 8, 01490. [Google Scholar] [CrossRef]
- Bates, K.A.; Friesen, J.; Loyau, A.; Butler, H.; Vredenburg, V.T.; Laufer, J.; Chatzinotas, A.; Schmeller, D.S. Environmental and Anthropogenic Factors Shape the Skin Bacterial Communities of a Semi-Arid Amphibian Species. Microb. Ecol. 2023, 86, 1393–1404. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.N.; James, T.Y.; Lauer, A.; Simon, M.A.; Patel, A. Amphibian Pathogen Batrachochytrium dendrobatidis Is Inhibited by the Cutaneous Bacteria of Amphibian Species. EcoHealth 2006, 3, 53–56. [Google Scholar] [CrossRef]
- McGrath-Blaser, S.; Steffen, M.; Grafe, T.U.; Torres-Sánchez, M.; McLeod, D.S.; Muletz-Wolz, C.R. Early Life Skin Microbial Trajectory as a Function of Vertical and Environmental Transmission in Bornean Foam-Nesting Frogs. Anim. Microbiome 2021, 3, 83. [Google Scholar] [CrossRef]
- Luedtke, J.A.; Chanson, J.; Neam, K.; Hobin, L.; Maciel, A.O.; Catenazzi, A.; Borzée, A.; Hamidy, A.; Aowphol, A.; Jean, A.; et al. Ongoing Declines for the World’s Amphibians in the Face of Emerging Threats. Nature 2023, 622, 308–314. [Google Scholar] [CrossRef]
- AmphibiaWeb Bufo bufo: Common Toad. 2024. Available online: https://amphibiaweb.org/species/127 (accessed on 7 January 2025).
- Massoulié, J.; Sussman, J.; Bon, S.; Silman, I. Chapter 15: Structure and Functions of Acetylcholinesterase and Butyrylcholinesterase. In Progress in Brain Research; Cuello, A.C., Ed.; Cholinergic Function and Dysfunction; Elsevier: Amsterdam, The Netherlands, 1993; Volume 98, pp. 139–146. [Google Scholar]
- Johnson, C.S.; Schwarzbach, S.E.; Henderson, J.D.; Wilson, B.W.; Tjeerdema, R.S. Influence of Water Temperature on Acetylcholinesterase Activity in the Pacific Tree Frog (Hyla regilla). Environ. Toxicol. Chem. 2005, 24, 2074–2077. [Google Scholar] [CrossRef]
- Attademo, A.M.; Curi, L.M.; Boccioni, A.P.C.; Barrios, C.E.; Peltzer, P.M.; Simoniello, M.F.; Lajmanovich, R.C.; Michlig, M.P.; Repetti, M.R.; Ríos, J.M. Microplastics and Plastic Additives as Contaminants of Emerging Concern: A Multi-Biomarker Approach Using Rhinella arenarum Tadpoles. Environ. Adv. 2023, 14, 100444. [Google Scholar] [CrossRef]
- Attademo, A.M.; Lajmanovich, R.C.; Peltzer, P.M.; Junges, C.M. Acute Toxicity of Metaldehyde in the Invasive Rice Snail Pomacea Canaliculata and Sublethal Effects on Tadpoles of a Non-Target Species (Rhinella arenarum). Water Air Soil. Pollut. 2016, 227, 400. [Google Scholar] [CrossRef]
- Vanderlinde, R.E. Measurement of Total Lactate Dehydrogenase Activity. Ann. Clin. Lab. Sci. 1985, 15, 13–31. [Google Scholar]
- Adams, E.; Finnegan, C.V. An Investigation of Lactate Dehydrogenase Activity in Early Amphibian Development. J. Exp. Zool. 1965, 158, 241–251. [Google Scholar] [CrossRef]
- Bennett, A.F. Enzymatic Correlates of Activity Metabolism in Anuran Amphibians. Am. J. Physiol.-Leg. Content 1974, 226, 1149. [Google Scholar] [CrossRef]
- Mendiola, P.; De Costa, J. The Effects of pH and Temperature on the Kinetic Properties of Skeletal Muscle Lactate Dehydrogenase from Anuran Amphibians. J. Comp. Physiol. B 1990, 160, 105–111. [Google Scholar] [CrossRef]
- Senko, H.; Kajić, S.; Huđ, A.; Palijan, G.; Petek, M.; Rajnović, I.; Šamec, D.; Udiković-Kolić, N.; Mešić, A.; Brkljačić, L.; et al. Will the Beneficial Properties of Plant-Growth Promoting Bacteria Be Affected by Waterlogging Predicted in the Wake of Climate Change: A Model Study. Appl. Soil. Ecol. 2024, 198, 105379. [Google Scholar] [CrossRef]
- Gosner, K.L. A Simplified Table for Staging Anuran Embryos and Larvae with Notes on Identification. Herpetologica 1960, 16, 183–190. [Google Scholar]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Frings, C.S.; Fendley, T.W.; Dunn, R.T.; Queen, C.A. Improved Determination of Total Serum Lipids by the Sulfo-Phospho-Vanillin Reaction. Clin. Chem. 1972, 18, 673–674. [Google Scholar] [CrossRef]
- Jermyn, M.A. Increasing the Sensitivity of the Anthrone Method for Carbohydrate. Anal. Biochem. 1975, 68, 332–335. [Google Scholar] [CrossRef]
- Bergmeyer, H.-U.; Bernt, E.; Hess, B. Lactic Dehydrogenase. In Methods of Enzymatic Analysis; Bergmeyer, H.-U., Ed.; Academic Press: Cambridge, MA, USA, 1965; pp. 736–743. ISBN 978-0-12-395630-9. [Google Scholar]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A New and Rapid Colorimetric Determination of Acetylcholinesterase Activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- van Doorn, J.; van den Bergh, D.; Böhm, U.; Dablander, F.; Derks, K.; Draws, T.; Etz, A.; Evans, N.J.; Gronau, Q.F.; Haaf, J.M.; et al. The JASP Guidelines for Conducting and Reporting a Bayesian Analysis. Psychon. Bull. Rev. 2021, 28, 813–826. [Google Scholar] [CrossRef]
- Ahmed, I.; Yokota, A.; Fujiwara, T. A Novel Highly Boron Tolerant Bacterium, Bacillus boroniphilus Sp. Nov., Isolated from Soil, That Requires Boron for Its Growth. Extremophiles 2007, 11, 217–224. [Google Scholar] [CrossRef]
- Müller, N.; Scherag, F.D.; Pester, M.; Schink, B. Bacillus stamsii Sp. Nov., a Facultatively Anaerobic Sugar Degrader That Is Numerically Dominant in Freshwater Lake Sediment. Syst. Appl. Microbiol. 2015, 38, 379–389. [Google Scholar] [CrossRef]
- Zhang, G.; Yang, R.; Chen, T.; Zhang, B.; Yang, H.; Wu, X.; Gao, H.; Zhang, W.; Liu, G. Mesobacillus harenae Sp. Nov., Isolated from the Sandy Soil of a Cold Desert. Int. J. Syst. Evol. Microbiol. 2021, 71, 004594. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Mañero, F.J.; Ramos-Solano, B.; Probanza, A.; Mehouachi, J.; Tadeo, F.R.; Talon, M. The Plant-Growth-Promoting Rhizobacteria Bacillus pumilus and Bacillus licheniformis Produce High Amounts of Physiologically Active Gibberellins. Physiol. Plant. 2001, 111, 206–211. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, S.; Mukherjee, A.; Rastogi, R.P.; Verma, J.P. Salt-Tolerant Plant Growth-Promoting Bacillus pumilus Strain JPVS11 to Enhance Plant Growth Attributes of Rice and Improve Soil Health under Salinity Stress. Microbiol. Res. 2021, 242, 126616. [Google Scholar] [CrossRef]
- Murugappan, R.M.; Begum, S.B.; Roobia, R.R. Symbiotic Influence of Endophytic Bacillus pumilus on Growth Promotion and Probiotic Potential of the Medicinal Plant Ocimum Sanctum. Symbiosis 2013, 60, 91–99. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced Systemic Resistance by Beneficial Microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef]
- Dökenel, G.; Özer, S. Bacterial Agents Isolated from Cultured Marsh Frog (Pelophylax ridibundus, Pallas 1771). EgeJFAS 2019, 36, 115–124. [Google Scholar] [CrossRef]
- Burmølle, M.; Ren, D.; Bjarnsholt, T.; Sørensen, S.J. Interactions in Multispecies Biofilms: Do They Actually Matter? Trends Microbiol. 2014, 22, 84–91. [Google Scholar] [CrossRef]
- Cuzziol Boccioni, A.P.; Lener, G.; Peluso, J.; Peltzer, P.M.; Attademo, A.M.; Aronzon, C.; Simoniello, M.F.; Demonte, L.D.; Repetti, M.R.; Lajmanovich, R.C. Comparative Assessment of Individual and Mixture Chronic Toxicity of Glyphosate and Glufosinate Ammonium on Amphibian Tadpoles: A Multibiomarker Approach. Chemosphere 2022, 309, 136554. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Models | P (M) | P (M|Data) | BF10 | Error % |
|---|---|---|---|---|
| Null model | 0.5 | 0.434 | 1 | |
| AChE | 0.5 | 0.566 | 1.306 | 3.386 × 10−5 |
| Null model | 0.5 | 0.393 | 1 | |
| LDH | 0.5 | 0.607 | 1.542 | 2.694 × 10−5 |
| Null model | 0.5 | 0.344 | 1 | |
| Weight | 0.5 | 0.656 | 1.906 | 2.556 × 10−5 |
| Null model | 0.5 | 0.579 | 1 | |
| TL | 0.5 | 0.421 | 0.728 | 8.520 × 10−5 |
| Null model | 0.5 | 6.828 × 10−4 | 1 | |
| SVL | 0.5 | 0.999 | 1463.601 | 2 × 10−3 |
| Null model | 0.5 | 0.305 | 1 | |
| Lipids | 0.5 | 0.695 | 2.278 | 1.594 × 10−5 |
| Null model | 0.5 | 0.31 | 1 | |
| Carbohydrates | 0.5 | 0.69 | 2.223 | 1.968 × 10−5 |
| Null model | 0.5 | 0.624 | 1 | |
| Proteins | 0.5 | 0.376 | 0.603 | 1.079 × 10−4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jovanović Glavaš, O.; Sviličić Petrić, I.; Palijan, G. Influence of Naturally Occurring Bacteria on Embryonic and Larval Development of Common Toad Tadpoles. Biology 2025, 14, 308. https://doi.org/10.3390/biology14030308
Jovanović Glavaš O, Sviličić Petrić I, Palijan G. Influence of Naturally Occurring Bacteria on Embryonic and Larval Development of Common Toad Tadpoles. Biology. 2025; 14(3):308. https://doi.org/10.3390/biology14030308
Chicago/Turabian StyleJovanović Glavaš, Olga, Ines Sviličić Petrić, and Goran Palijan. 2025. "Influence of Naturally Occurring Bacteria on Embryonic and Larval Development of Common Toad Tadpoles" Biology 14, no. 3: 308. https://doi.org/10.3390/biology14030308
APA StyleJovanović Glavaš, O., Sviličić Petrić, I., & Palijan, G. (2025). Influence of Naturally Occurring Bacteria on Embryonic and Larval Development of Common Toad Tadpoles. Biology, 14(3), 308. https://doi.org/10.3390/biology14030308