
How to Define Spacing Among Forest Trees to Mitigate Competition: A Technical Note

 ,
,  ,
,  ,
, 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
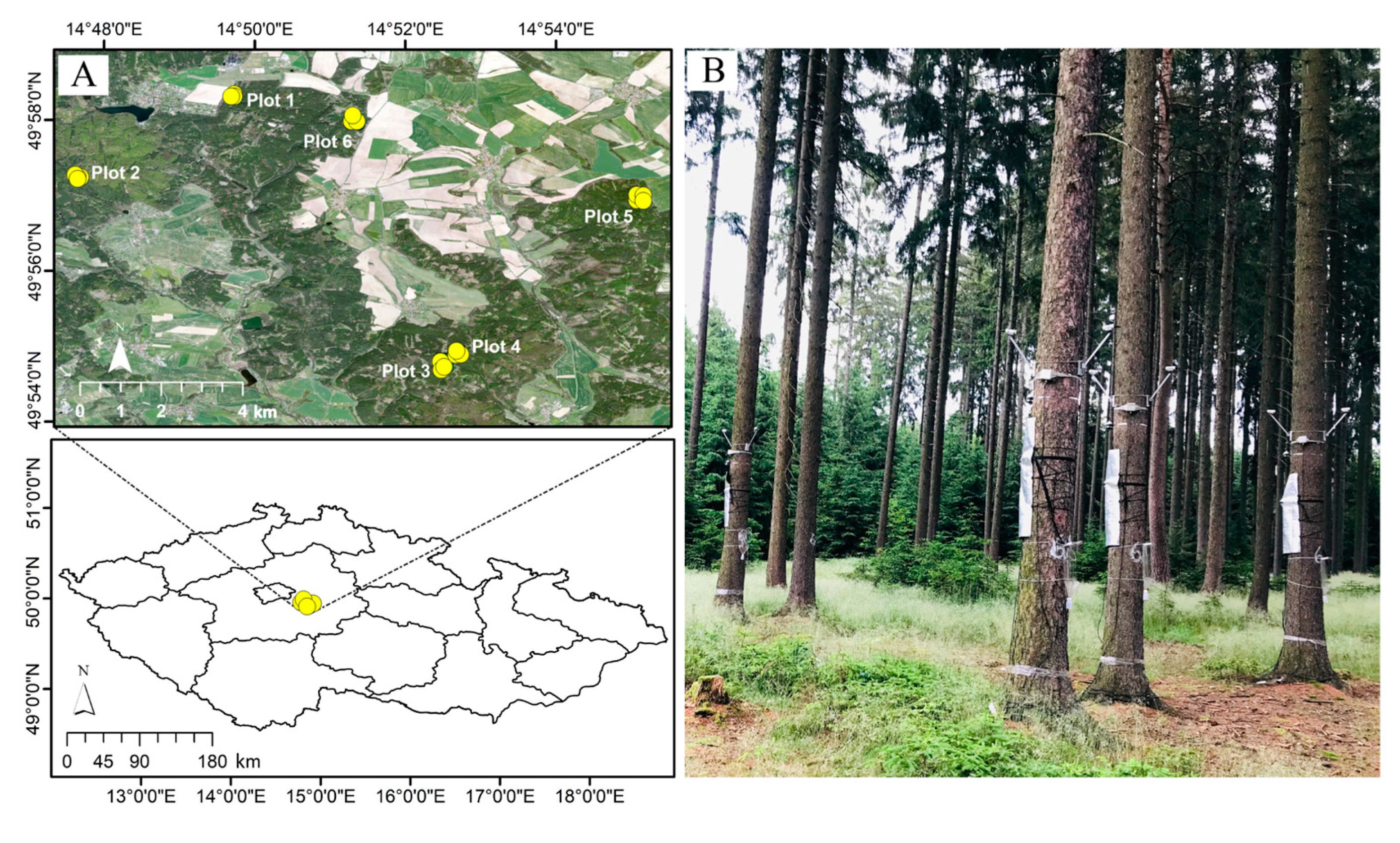
2.1. Study Area and Field Measurements
2.2. Estimated Spacing Among Trees Using Tree Density Within the Buffer
2.3. Controlling the Effects of Nuisance Variables on Sap Flow Before Developing the Models
2.4. Regression Diagnostics
2.5. Filtering out the Effects of Elevation and DBH on Sap Flow
2.6. Experimental Variogram
3. Results
3.1. Proposed Regression Models
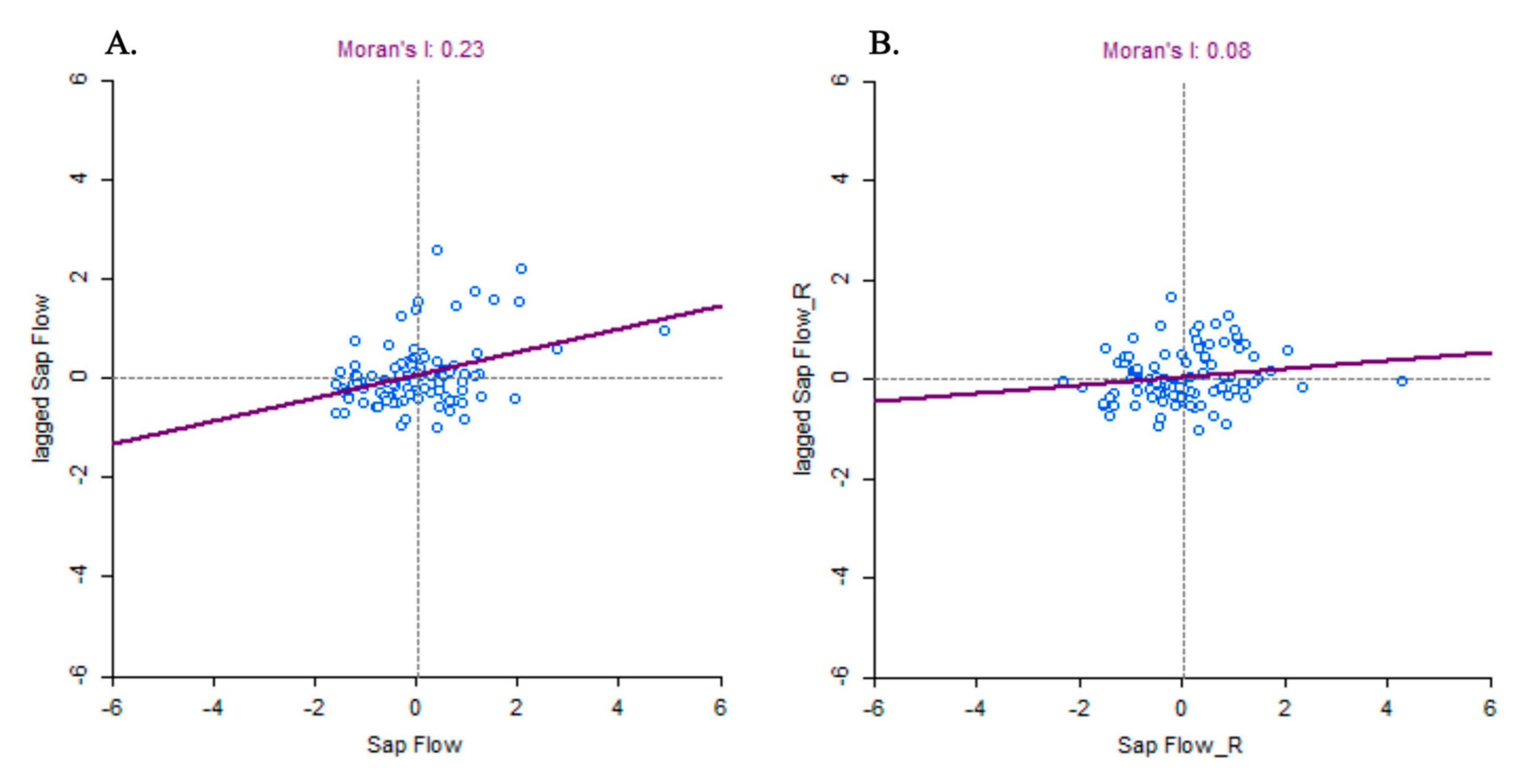
3.2. Spatial Autocorrelations in Sap Flow Drivers, Sap Flow, and Sap Flow Residuals
3.3. Experimental Variogram
4. Discussion
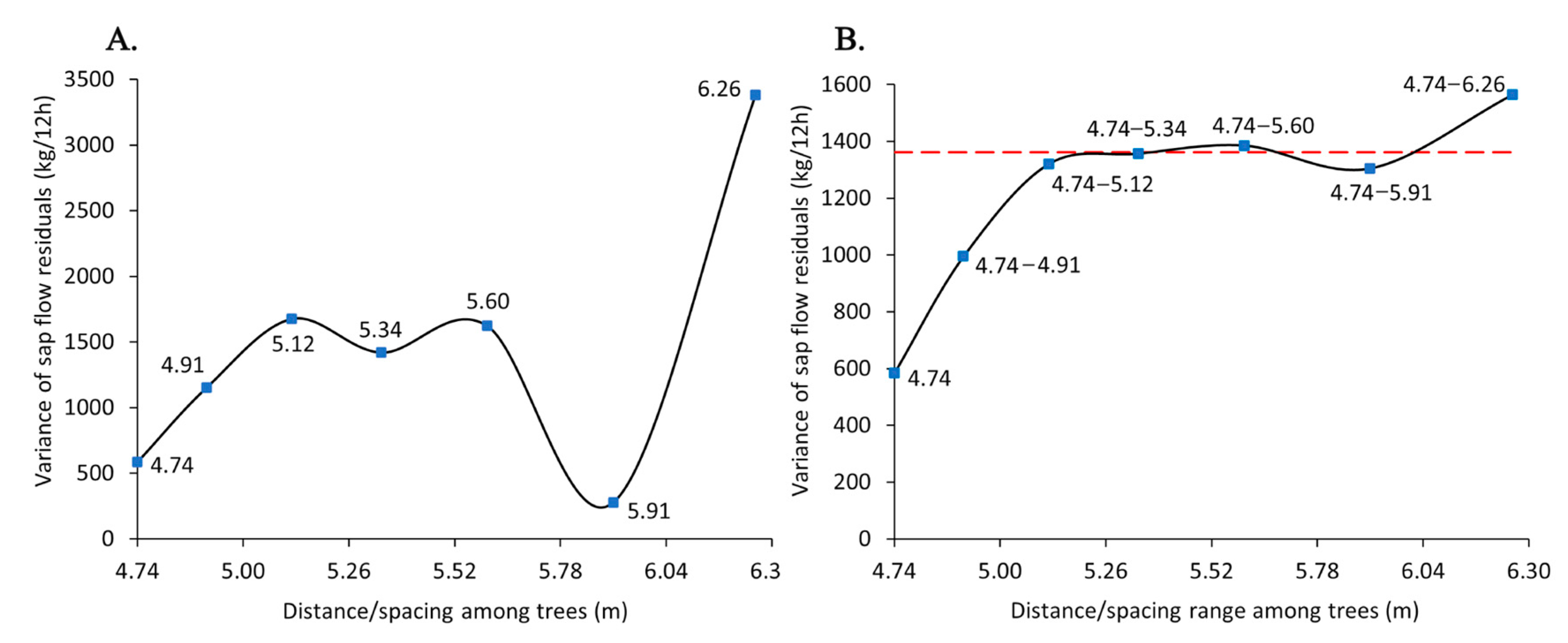
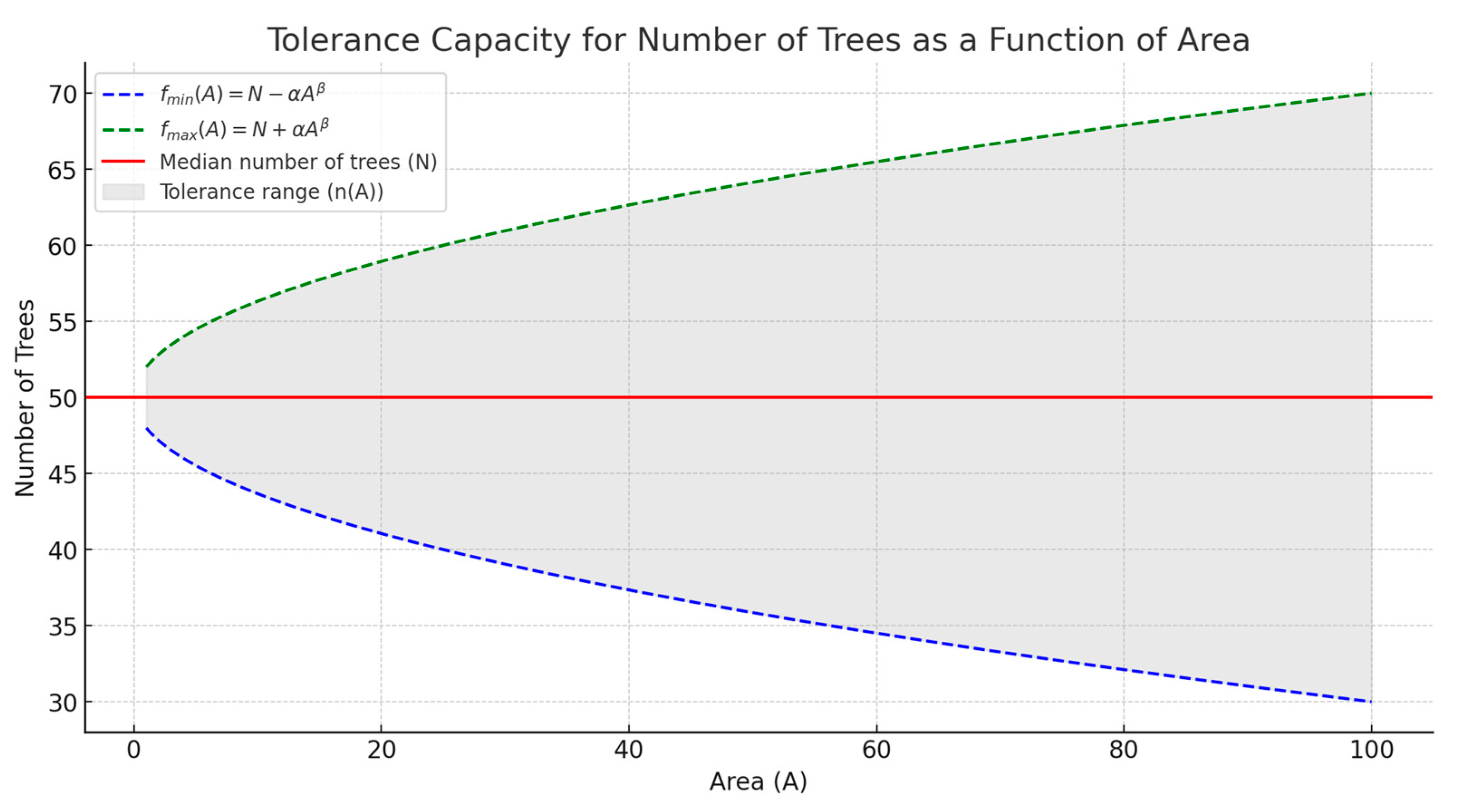
4.1. Spacing Range to Mitigate Competition Pressure
4.2. Comparison to National Forest Inventory (NFI) Data on Tree Density
4.3. The Introduced Technique
4.4. Future Silviculture Practices
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Öberg, L.; Kullman, L. Ancient Subalpine Clonal Spruces (Picea abies): Sources of Postglacial Vegetation History in the Swedish Scandes. Arctic 2011, 64, 183–196. [Google Scholar] [CrossRef]
- Singh, V.V.; Naseer, A.; Mogilicherla, K.; Trubin, A.; Zabihi, K.; Roy, A.; Jakuš, R.; Erbilgin, N. Understanding Bark Beetle Outbreaks: Exploring the Impact of Changing Temperature Regimes, Droughts, Forest Structure, and Prospects for Future Forest Pest Management. Rev. Environ. Sci. Bio Technol. 2024, 23, 257–290. [Google Scholar] [CrossRef]
- Johnson, O.; More, D. Collins Tree Guide; Collins: London, UK, 2004; pp. 42–43. [Google Scholar]
- OECD. Safety Assessment of Transgenic Organisms; OECD Consensus Documents; Harmonisation of Regulatory Oversight in Biotechnology; OECD Publishing: Paris, France, 2006; Volume 2. [Google Scholar]
- Eckenwalder, J.E. Conifers of the World: The Complete Reference; Timber Press: Portland, OR, USA, 2009. [Google Scholar]
- Caudullo, G.; Tinner, W.; de Rigo, D. Picea Abies in Europe: Distribution, Habitat, Usage and Threats. In European Atlas of Forest Tree Species; Publications Office of the European Union: Luxembourg, 2016. [Google Scholar]
- Horgan, T.; Keane, M.; McCarthy, R.; Lally, M.; Thompson, D.; O’Carroll, J. A Guide to Forest Tree Species Selection and Silviculture in Ireland; COFORD: Dublin, Ireland, 2003. [Google Scholar]
- Singh, V.V.; Zabihi, K.; Trubin, A.; Cudlín, P.; Korolyova, N.; Jakuš, R.; Blaženec, M. Effect of Diurnal Solar Radiation Regime and Tree Density on Sap Flow of Norway Spruce (Picea abies [L.] Karst.) in Fragmented Stands. Cent. Eur. For. J. 2025, 71. [Google Scholar] [CrossRef]
- Thorpe, H.C.; Astrup, R.; Trowbridge, A.; Coates, K.D. Competition and Tree Crowns: A Neighborhood Analysis of Three Boreal Tree Species. For. Ecol. Manag. 2010, 259, 1586–1596. [Google Scholar] [CrossRef]
- Kenk, G. Management of Norway Spruce Without Precommercial Thinnings? The Example of a 115 Year Old Stand in the Growth Region of the Suabian Alb. Allg. Forstztg. 1988, 30, 837–839. [Google Scholar]
- Kenk, G. Wide Spacing in Norway Spruce Stands. Development and Consequences (In German with English Summary). Forstwiss. Cent. 1990, 109, 86–100. [Google Scholar]
- Mäkinen, H.; Hein, S. Effect of Wide Spacing on Increment and Branch Properties of Young Norway Spruce. Eur. J. For. Res. 2006, 125, 239–248. [Google Scholar] [CrossRef]
- Emmel, C.; Paul-Limoges, E.; Black, T.A.; Christen, A. Vertical Distribution of Radiation and Energy Balance Partitioning Within and Above a Lodgepole Pine Stand Recovering from a Recent Insect Attack. Bound.-Layer Meteorol. 2013, 149, 133–163. [Google Scholar] [CrossRef]
- Vanderhoof, M.; Williams, C.A.; Shuai, Y.; Jarvis, D.; Kulakowski, D.; Masek, J. Albedo-Induced Radiative Forcing from Mountain Pine Beetle Outbreaks in Forests, South-Central Rocky Mountains: Magnitude, Persistence, and Relation to Outbreak Severity. Biogeosciences 2014, 11, 563–575. [Google Scholar] [CrossRef]
- Zabihi Afratakhti, K.; Singh, V.V.; Trubin, A.; Tomášková, I.; Blaženec, M.; Surový, P.; Jakuš, R. Sap Flow as a Function of Variables Within Nested Scales: Ordinary Least Squares vs. Spatial Regression Models. Environ. Res. Ecol. 2023, 2, 025002. [Google Scholar] [CrossRef]
- Hébert, F.; Krause, C.; Plourde, P.-Y.; Achim, A.; Prégent, G.; Ménétrier, J. Effect of Tree Spacing on Tree Level Volume Growth, Morphology, and Wood Properties in a 25-Year-Old Pinus banksiana Plantation in the Boreal Forest of Quebec. Forests 2016, 7, 276. [Google Scholar] [CrossRef]
- Alarcón, J.; Domingo, R.; Green, S.R.; Sánchez-Blanco, M.J.; Rodríguez, P.; Torrecillas, A. Sap Flow as an Indicator of Transpiration and the Water Status of Young Apricot Trees. Plant Soil 2000, 227, 77–85. [Google Scholar] [CrossRef]
- Özçelik, M.S.; Tomášková, I.; Surový, P.; Modlinger, R. Effect of Forest Edge Cutting on Transpiration Rate in Picea abies (L.) H. Karst. Forests 2022, 13, 1238. [Google Scholar] [CrossRef]
- West, R.R.; Lada, R.R.; MacDonald, M.T. Nutrition and Related Factors Affecting Maple Tree Health and Sap Yield. Am. J. Plant Sci. 2023, 14, 125–149. [Google Scholar] [CrossRef]
- Ježík, M.; Blaženec, M.; Letts, M.G.; Ditmarová, Ľ.; Sitková, Z.; Střelcová, K. Assessing Seasonal Drought Stress Response in Norway Spruce (Picea abies (L.) Karst.) by Monitoring Stem Circumference and Sap Flow. Ecohydrology 2015, 8, 378–386. [Google Scholar] [CrossRef]
- Korakaki, E.; Fotelli, M.N. Sap Flow in Aleppo Pine in Greece in Relation to Sapwood Radial Gradient, Temporal and Climatic Variability. Forests 2021, 12, 2. [Google Scholar] [CrossRef]
- Cao, Q.; Li, J.; Xiao, H.; Cao, Y.; Xin, Z.; Yang, B.; Liu, T.; Yuan, M. Sap Flow of Amorpha fruticosa: Implications of Water Use Strategy in a Semiarid System with Secondary Salinization. Sci. Rep. 2020, 10, 13504. [Google Scholar] [CrossRef]
- Suárez, J.C.; Casanoves, F.; Bieng, M.A.N.; Melgarejo, L.M.; Di Rienzo, J.A.; Armas, C. Prediction Model for Sap Flow in Cacao Trees Under Different Radiation Intensities in the Western Colombian Amazon. Sci. Rep. 2021, 11, 10512. [Google Scholar] [CrossRef]
- Kume, T.; Tsuruta, K.; Komatsu, H.; Shinohara, Y.; Katayama, A.; Ide, J.I.; Otsuki, K. Differences in Sap Flux-Based Stand Transpiration Between Upper and Lower Slope Positions in a Japanese Cypress Plantation Watershed. Ecohydrology 2016, 9, 1105–1116. [Google Scholar] [CrossRef]
- Schoppach, R.; Chun, K.P.; He, Q.; Fabiani, G.; Klaus, J. Species-Specific Control of DBH and Landscape Characteristics on Tree-to-Tree Variability of Sap Velocity. Agric. For. Meteorol. 2021, 307, 108533. [Google Scholar] [CrossRef]
- Hassler, S.K.; Weiler, M.; Blume, T. Tree-, Stand- and Site-Specific Controls on Landscape-Scale Patterns of Transpiration. Hydrol. Earth Syst. Sci. 2018, 22, 13–30. [Google Scholar] [CrossRef]
- Safargalieva, A.; Kochetkova, I.; Makeeva, E.; Shorgin, S. CEUR Workshop Proceedings. In Proceedings of the Workshop on Information Technology and Scientific Computing in the Framework of the XI International Conference Information and Telecommunication Technologies and Mathematical Modeling of High-Tech Systems (ITTMM-2021), Moscow, Russia, 19–23 April 2021. [Google Scholar]
- Lagergren, F.; Lindroth, A. Variation in sapflow and stem growth in relation to tree size, competition and thinning in a mixed forest of pine and spruce in Sweden. For. Ecol. Man. 2004, 188, 51–63. [Google Scholar] [CrossRef]
- Tolasz, R.; Míková, T.; Valeriánová, A.; Voženílek, V. Climate Atlas of Czechia; Czech Hydrometeorological Institute: Prague, Czech Republic, 2007; p. 256. [Google Scholar]
- Trubin, A.; Kozhoridze, G.; Zabihi, K.; Modlinger, R.; Singh, V.V.; Surový, P.; Jakuš, R. Detection of Green Attack and Bark Beetle Susceptibility in Norway Spruce: Utilizing PlanetScope Multispectral Imagery for Tri-Stage Spectral Separability Analysis. For. Ecol. Manag. 2024, 560, 121838. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2006; World Soil Resources Reports No. 103; FAO: Rome, Italy, 2006. [Google Scholar]
- Čermák, J.; Kučera, J.; Nadezhdina, N. Sap Flow Measurements with Some Thermodynamic Methods, Flow Integration Within Trees and Scaling Up from Sample Trees to Entire Forest Stands. Trees 2004, 18, 529–546. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Desktop: Release 10.8.1; Environmental Systems Research Institute: Redlands, CA, USA, 2020. [Google Scholar]
- Anselin, L.; Syabri, I.; Kho, Y. GeoDa: An Introduction to Spatial Data Analysis. Geogr. Anal. 2006, 38, 5–22. [Google Scholar] [CrossRef]
- Akaike, H. Information Theory and an Extension of the Maximum Likelihood Principle. In Proceedings of the 2nd International Symposium on Information Theory, Tsahkadsor, Armenia, 2–8 September 1971; Petrov, B.N., Csaki, F., Eds.; Akadémiai Kiadó: Budapest, Hungary, 1973; pp. 267–281. [Google Scholar]
- Ortiz, S.M.; Breidenbach, J.; Kändeler, G. Early Detection of Bark Beetle Green Attack Using TerraSAR-X and RapidEye Data. Remote Sens. 2013, 5, 1912–1931. [Google Scholar] [CrossRef]
- Zabihi, K.; Surový, P.; Trubin, A.; Singh, V.V.; Jakuš, R. A Review of Major Factors Influencing the Accuracy of Mapping Green-Attack Stage of Bark Beetle Infestations Using Satellite Imagery: Prospects to Avoid Data Redundancy. Remote Sens. Appl. Soc. Environ. 2021, 24, 100638. [Google Scholar] [CrossRef]
- Krajicek, J.E.; Brinkman, K.A.; Gingrich, S.F. Crown Competition—A Measure of Density. For. Sci. 1961, 7, 35–42. [Google Scholar]
- Katrevičs, J.; Džeriņa, B.; Neimane, U.; Desaine, I.; Bigača, Z.; Jansons, Ā. Production and Profitability of Low Density Norway Spruce (Picea abies (L.) Karst.) Plantation at 50 Years of Age: Case Study from Eastern Latvia. Agronomy 2018, 16, 113–121. [Google Scholar]
- Guidotti, A. On the Processes and Factors Shaping the Norway Spruce’s (Picea abies) Forests in the Southern Swiss Alps. Master’s Thesis, Swiss Federal Institute of Technology, Zurich, Switzerland, 2020. [Google Scholar]
- McClain, K.M.; Morris, D.M.; Hills, S.C.; Buss, L.J. The Effect of Initial Spacing on Growth and Crown Development for Planted Northern Conifers. For. Chron. 1994, 70, 174–182. [Google Scholar] [CrossRef]
- Gardiner, B.A.; Quine, C.P. Management of Forests to Reduce the Risk of Abiotic Damage: A Review with Particular Reference to the Effect of Strong Winds. For. Ecol. Manag. 2000, 135, 261–277. [Google Scholar] [CrossRef]
- Peltola, H.; Kellomäki, S.; Hassinen, A.; Granander, M. Mechanical Stability of Scots Pine, Norway Spruce, and Birch: An Analysis of Tree Pulling Experiment in Finland. For. Ecol. Manag. 2000, 135, 143–153. [Google Scholar] [CrossRef]
- Slodicak, M.; Novak, J. Silvicultural Measures to Increase the Mechanical Stability of Pure Secondary Norway Spruce Stands Before Conversion. For. Ecol. Manag. 2006, 224, 252–257. [Google Scholar] [CrossRef]
- Akers, M.K.; Kane, M.; Zhao, D.; Teskey, R.O.; Daniels, R.F. Effects of Planting Density and Cultural Intensity on Stand and Crown Attributes of Mid-Rotation Loblolly Pine Plantations. For. Ecol. Manag. 2013, 310, 468–475. [Google Scholar] [CrossRef]
- Gleason, K.E.; Bradford, J.B.; Bottero, A.; D’Amato, A.W.; Fraver, S.; Palik, B.J.; Battaglia, M.A.; Iverson, L.; Kenefic, L.; Kern, C.C. Competition Amplifies Drought Stress in Forests Across Broad Climatic and Compositional Gradients. Ecosphere 2017, 8, e01849. [Google Scholar] [CrossRef]
- Bernal, A.A.; Kane, J.M.; Knapp, E.E.; Zald, H.S.J. Tree Resistance to Drought and Bark Beetle-Associated Mortality Following Thinning and Prescribed Fire Treatments. For. Ecol. Manag. 2023, 530, 120758. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. of Trees Within a 10 m Radius Buffer (Tree Density) | Distance Among Trees Assuming Equal-Spaced Trees (m) | No. of Observed Buffers with a Particular Tree Density |
|---|---|---|
| 14 | 4.74 | 9 |
| 13 | 4.91 | 13 |
| 12 | 5.12 | 24 |
| 11 | 5.34 | 25 |
| 10 | 5.60 | 13 |
| 9 | 5.91 | 7 |
| 8 | 6.26 | 10 |
| Dates, Time | Dependent/Response Variable | Independent/Explanatory Variables (Covariates) |
|---|---|---|
| Elevation (m) | ||
| 15–26 April 2019, 6 a.m.–6 p.m. | Sap flow (sum on average; kg/12 h) | Tree density |
| DBH (cm) |
| Variable | Elevation | Sap Flow | DBH | Tree Density |
|---|---|---|---|---|
| Elevation | 1 | |||
| Sap flow | 0.49 (0.24), <0.01 | 1 | ||
| DBH | 0.31 (0.1), <0.01 | 0.47 (0.22), <0.01 | 1 | |
| Tree density | −0.50 (0.25), <0.01 | −0.43 (0.18), <0.01 | −0.21 (0.04), 0.03 | 1 |
| Proposed Model | Model no. | OLS Adjusted R2 | OLS AIC | OLS Moran’s I p-Value | Spatial Lag R2 | Spatial Lag AIC | Spatial Error R2 | Spatial Error AIC |
|---|---|---|---|---|---|---|---|---|
| TreeDensity | 1 | 0.18 | 1055.70 | 0.046 | 0.22 | 1053.37 | 0.22 | 1053.26 |
| Elevation + DBH | 2 | 0.33 | 1034.43 | 0.079 | - | - | - | - |
| TreeDensity + Elevation + DBH | 3 (Full Model) | 0.37 | 1030.83 | 0.046 | 0.40 | 1032.19 | 0.41 | 1028.51 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zabihi, K.; Singh, V.V.; Trubin, A.; Korolyova, N.; Jakuš, R. How to Define Spacing Among Forest Trees to Mitigate Competition: A Technical Note. Biology 2025, 14, 296. https://doi.org/10.3390/biology14030296
Zabihi K, Singh VV, Trubin A, Korolyova N, Jakuš R. How to Define Spacing Among Forest Trees to Mitigate Competition: A Technical Note. Biology. 2025; 14(3):296. https://doi.org/10.3390/biology14030296
Chicago/Turabian StyleZabihi, Khodabakhsh, Vivek Vikram Singh, Aleksei Trubin, Nataliya Korolyova, and Rastislav Jakuš. 2025. "How to Define Spacing Among Forest Trees to Mitigate Competition: A Technical Note" Biology 14, no. 3: 296. https://doi.org/10.3390/biology14030296
APA StyleZabihi, K., Singh, V. V., Trubin, A., Korolyova, N., & Jakuš, R. (2025). How to Define Spacing Among Forest Trees to Mitigate Competition: A Technical Note. Biology, 14(3), 296. https://doi.org/10.3390/biology14030296