
Understanding the Foreign Body Response via Single-Cell Meta-Analysis

 , , , , and
, , , , and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
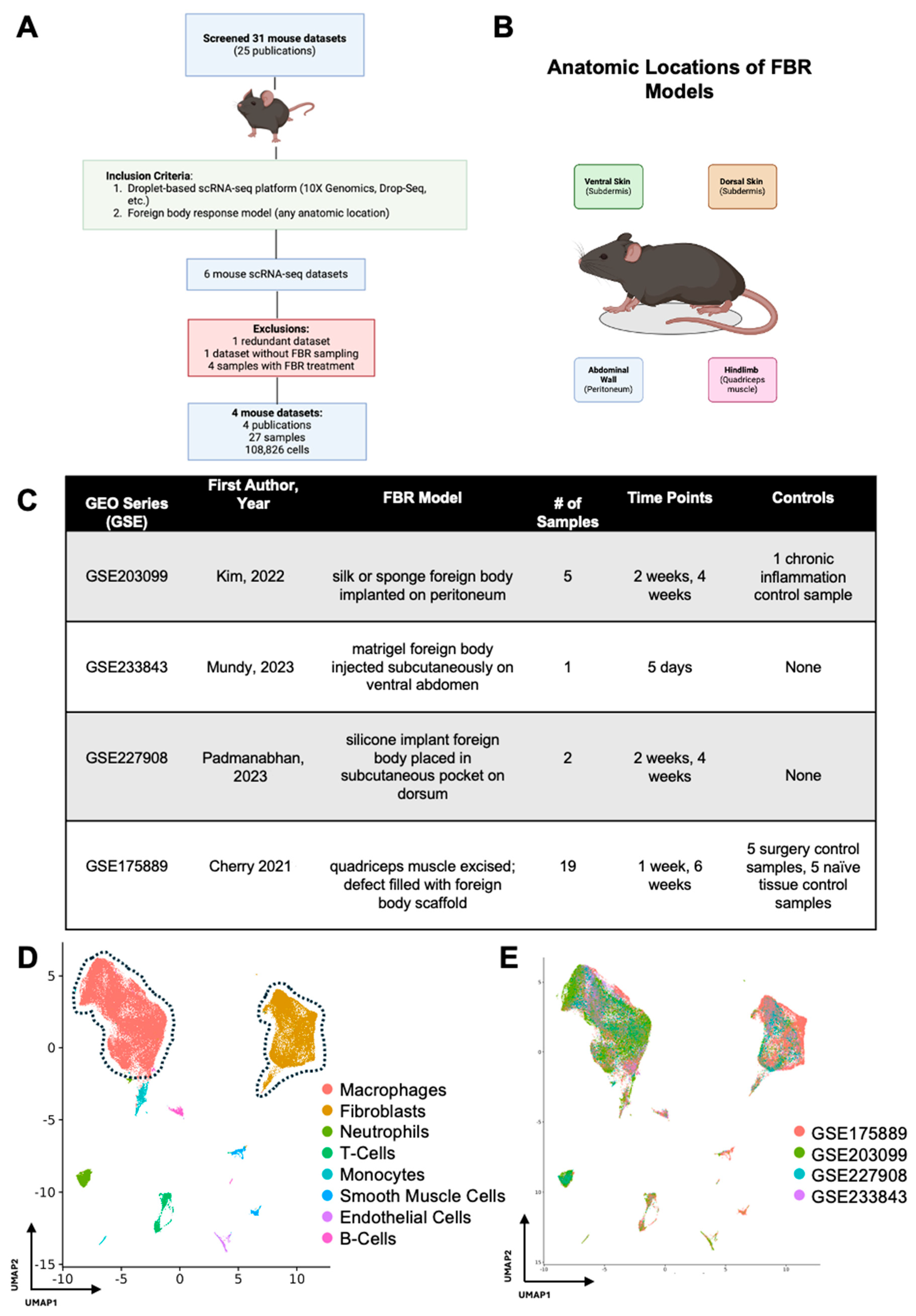
2.1. Screening of FBR-Related scRNA-seq Datasets
- GEO: (“implant” OR “Foreign Body*” OR “FBR”) AND ((“single cell*” OR “single-cell*” OR “scRNA*” OR “sc-RNA*”) OR (“single nuc*” OR “single-nuc*” OR “snRNA*” OR “sn-RNA*”))
- PubMed: (“implant” OR “Foreign Body*” OR “FBR”) AND ((“single cell*” OR “single-cell*” OR “scRNA*” OR “sc-RNA*”) OR (“single nuc*” OR “single-nuc*” OR “snRNA*” OR “sn-RNA*”))
2.2. Integration of FBR Datasets
3. Results
3.1. Identification of Major Cell Types Involved in FBR
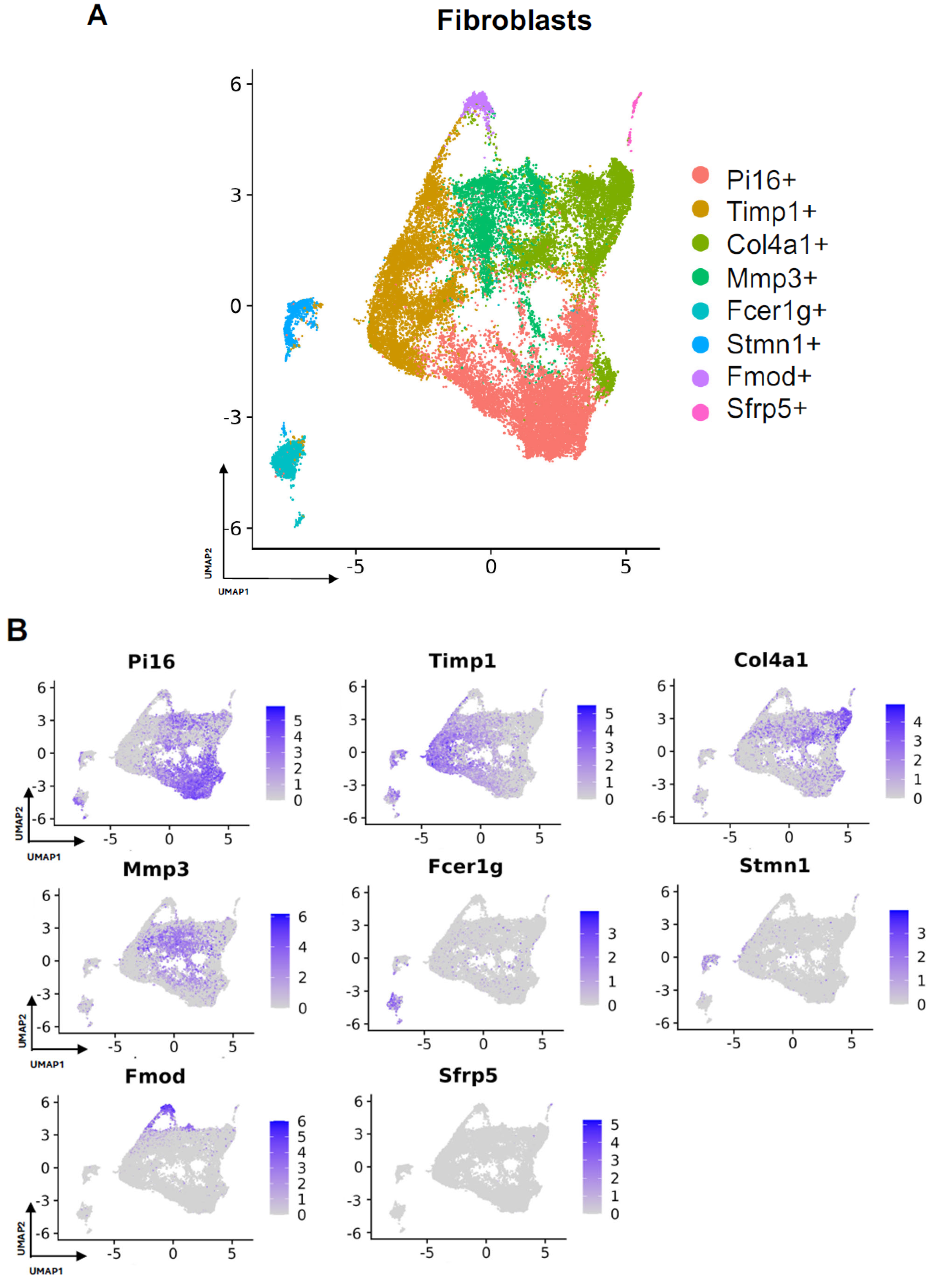
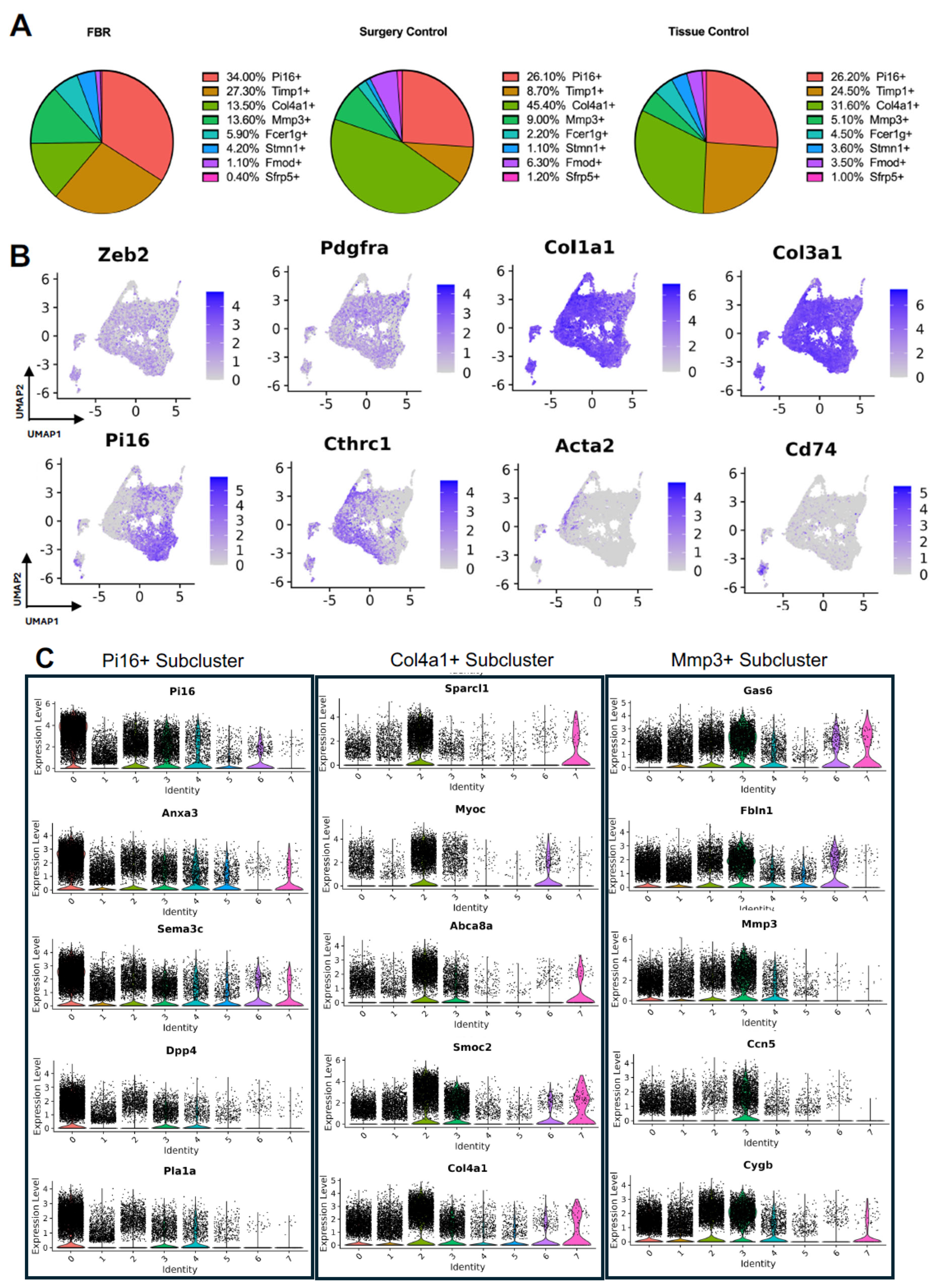
3.2. Detailing the Fibroblast Subpopulations Driving FBR
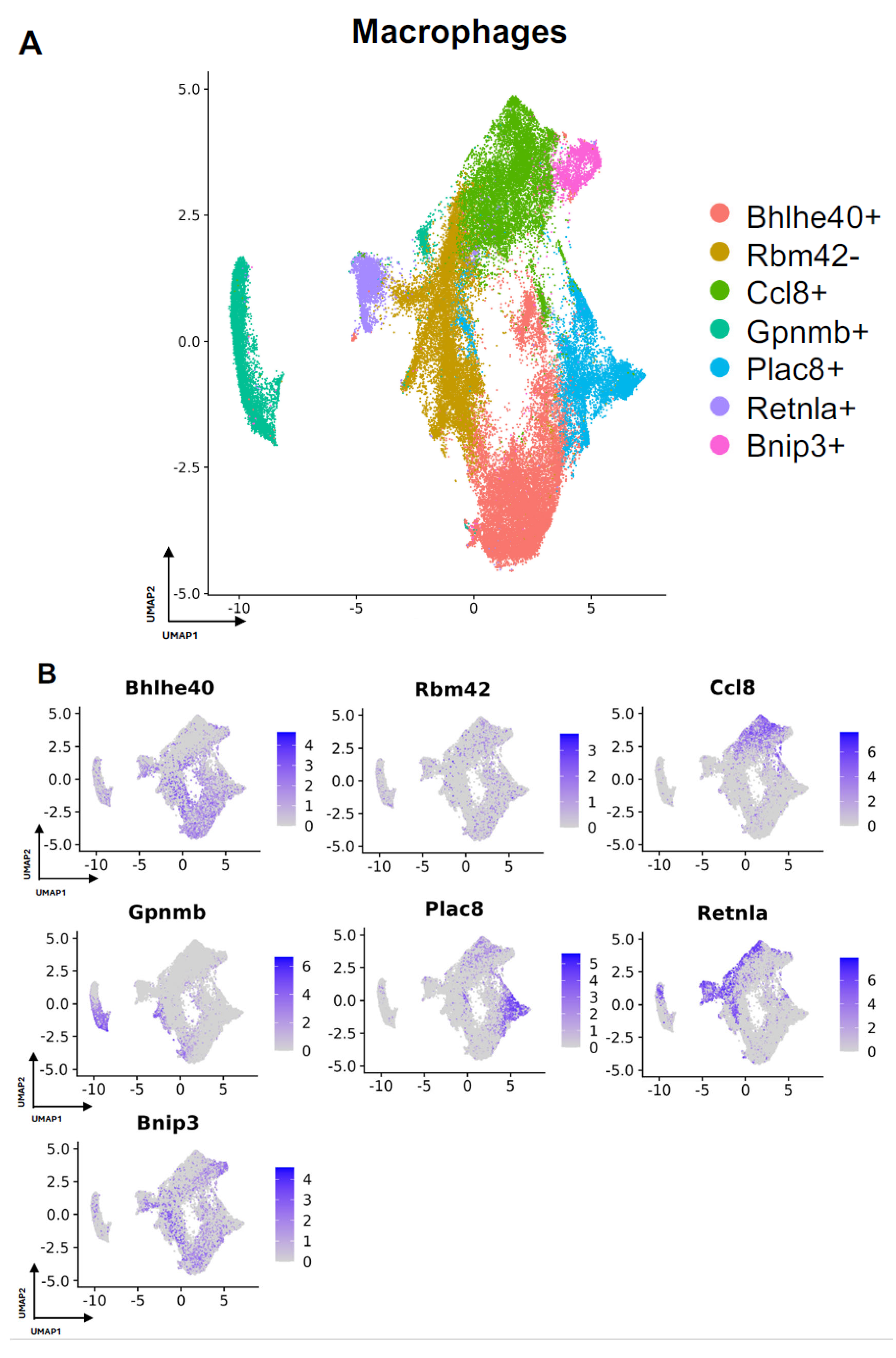
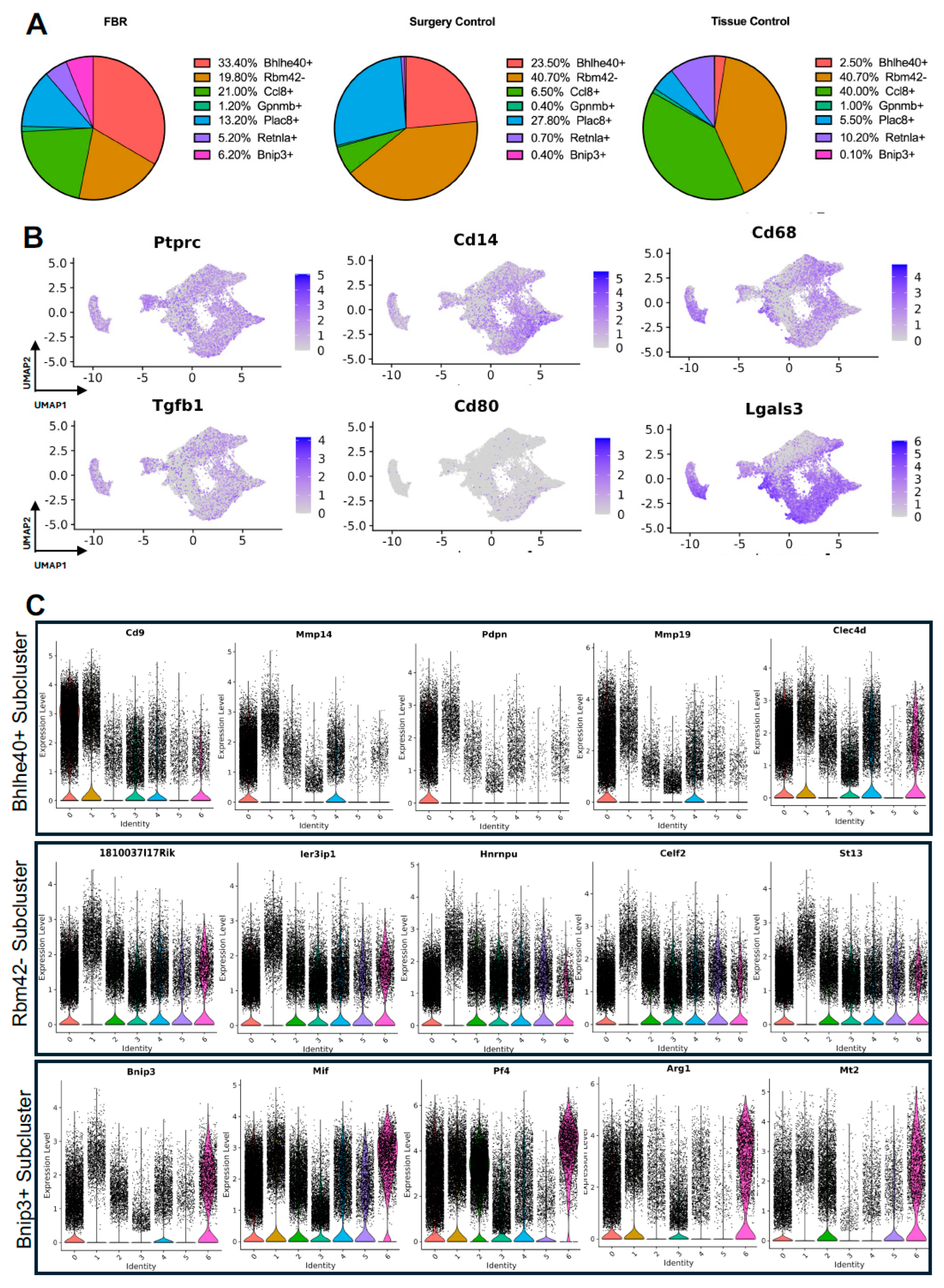
3.3. Characterizing the Key Macrophage Subpopulations in FBR
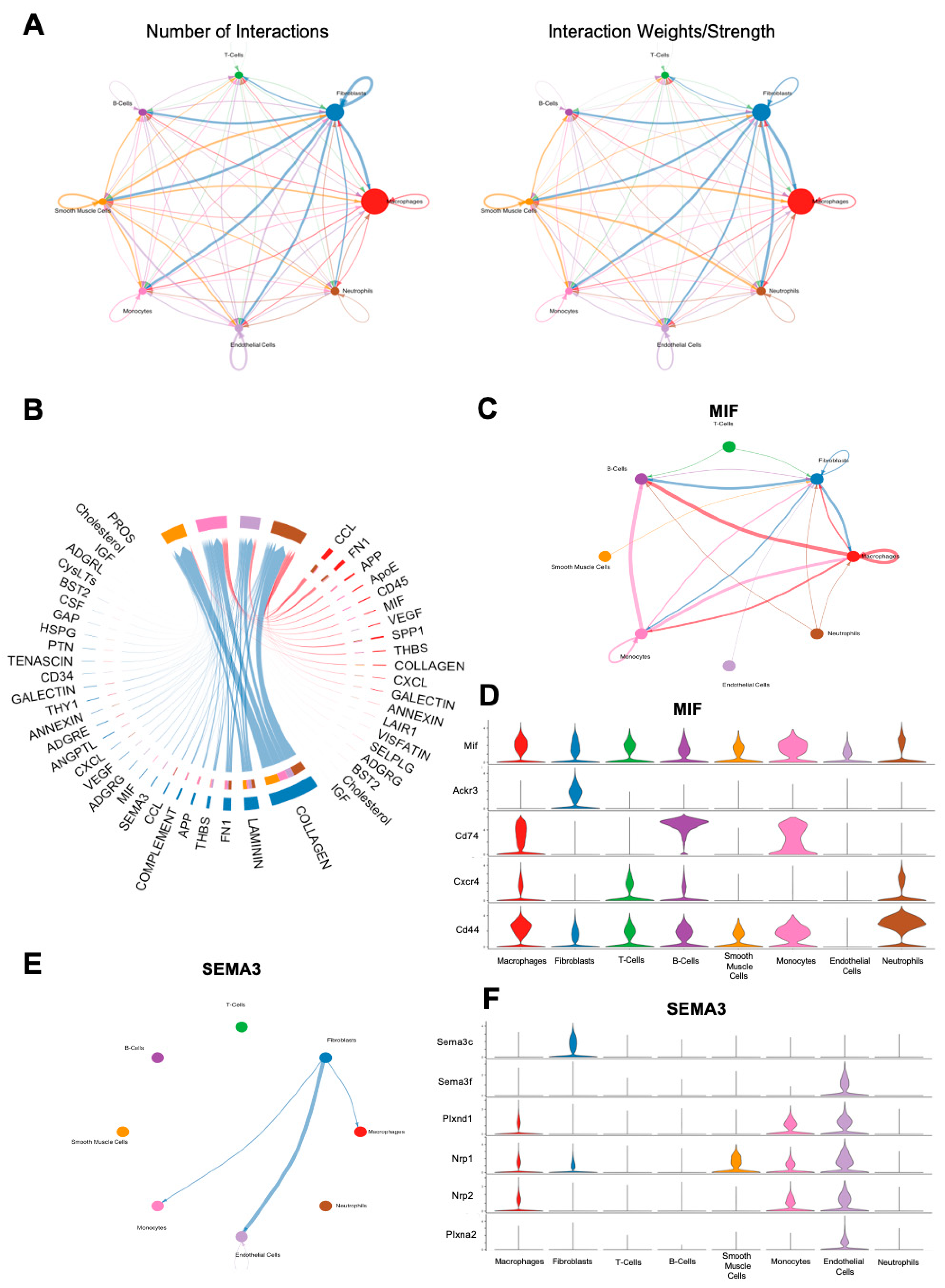
3.4. CellChat Analysis of FBR Subpopulations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Parker, J.B.; Griffin, M.F.; Spielman, A.F.; Wan, D.C.; Longaker, M.T. Exploring the Overlooked Roles and Mechanisms of Fibroblasts in the Foreign Body Response. Adv. Wound Care 2023, 12, 85–96. [Google Scholar] [CrossRef]
- Major, M.R.; Wong, V.W.; Nelson, E.R.; Longaker, M.T.; Gurtner, G.C. The foreign body response: At the interface of surgery and bioengineering. Plast. Reconstr. Surg. 2015, 135, 1489–1498. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.M.; Rodriguez, A.; Chang, D.T. Foreign body reaction to biomaterials. Semin Immunol. 2008, 20, 86–100. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Li, Y.; Jia, L.; Han, Y.; Cheng, J.; Li, H.; Qi, Y.; Du, J. Macrophage-stimulated cardiac fibroblast production of IL-6 is essential for TGF beta/Smad activation and cardiac fibrosis induced by angiotensin II. PLoS ONE 2012, 7, e35144. [Google Scholar]
- Mascharak, S.; desJardins-Park, H.E.; Davitt, M.F.; Griffin, M.; Borrelli, M.R.; Moore, A.L.; Chen, K.; Duoto, B.; Chinta, M.; Foster, D.S.; et al. Preventing Engrailed-1 activation in fibroblasts yields wound regeneration without scarring. Science 2021, 372, eaba2374. [Google Scholar] [CrossRef]
- Kim, Y.S.; Shin, S.; Choi, E.J.; Moon, S.W.; Jung, C.K.; Chung, Y.J.; Lee, S.H. Different Molecular Features of Epithelioid and Giant Cells in Foreign Body Reaction Identified by Single-Cell RNA Sequencing. J. Investig. Dermatol. 2022, 142, 3232–3242 e16. [Google Scholar] [CrossRef]
- Mundy, C.; Yao, L.; Shaughnessy, K.A.; Saunders, C.; Shore, E.M.; Koyama, E.; Pacifici, M. Palovarotene Action against Heterotopic Ossification Includes a Reduction of Local Participating Activin A-Expressing Cell Populations. JBMR Plus 2023, 7, e10821. [Google Scholar] [CrossRef]
- Padmanabhan, J.; Chen, K.; Sivaraj, D.; Henn, D.; Kuehlmann, B.A.; Kussie, H.C.; Zhao, E.T.; Kahn, A.; Bonham, C.A.; Dohi, T.; et al. Allometrically scaling tissue forces drive pathological foreign-body responses to implants via Rac2-activated myeloid cells. Nat. Biomed. Eng. 2023, 7, 1419–1436. [Google Scholar] [CrossRef] [PubMed]
- Cherry, C.; Maestas, D.R.; Han, J.; Andorko, J.I.; Cahan, P.; Fertig, E.J.; Garmire, L.X.; Elisseeff, J.H. Computational reconstruction of the signalling networks surrounding implanted biomaterials from single-cell transcriptomics. Nat. Biomed. Eng. 2021, 5, 1228–1238. [Google Scholar] [CrossRef]
- Sommerfeld, S.D.; Cherry, C.; Schwab, R.M.; Chung, L.; Maestas, D.R., Jr.; Laffont, P.; Stein, J.E.; Tam, A.; Ganguly, S.; Housseau, F.; et al. Interleukin-36γ-producing macrophages drive IL-17-mediated fibrosis. Sci. Immunol. 2019, 4, eaax4783. [Google Scholar] [CrossRef]
- Luecken, M.D.; Buttner, M.; Chaichoompu, K.; Danese, A.; Interlandi, M.; Mueller, M.F.; Strobl, D.C.; Zappia, L.; Dugas, M.; Colomé-Tatché, M.; et al. Benchmarking atlas-level data integration in single-cell genomics. Nat. Methods 2022, 19, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Guerrero-Juarez, C.F.; Zhang, L.; Chang, I.; Ramos, R.; Kuan, C.H.; Myung, P.; Plikus, M.V.; Nie, Q. Inference and analysis of cell-cell communication using CellChat. Nat. Commun. 2021, 12, 1088. [Google Scholar] [CrossRef] [PubMed]
- Socarras, T.O.; Vasconcelos, A.C.; Campos, P.P.; Pereira, N.B.; Souza, J.P.; Andrade, S.P. Foreign body response to subcutaneous implants in diabetic rats. PLoS ONE 2014, 9, e110945. [Google Scholar] [CrossRef] [PubMed]
- Rothuizen, T.C.; Damanik, F.F.; Anderson, J.M.; Lavrijsen, T.; Cox, M.A.; Rabelink, T.J.; Moroni, L.; Rotmans, J.I. Tailoring the foreign body response for in situ vascular tissue engineering. Tissue Eng. Part C Methods 2015, 21, 436–446. [Google Scholar] [CrossRef] [PubMed]
- Li, A.G.; Quinn, M.J.; Siddiqui, Y.; Wood, M.D.; Federiuk, I.F.; Duman, H.M.; Ward, W.K. Elevation of transforming growth factor beta (TGFbeta) and its downstream mediators in subcutaneous foreign body capsule tissue. J. Biomed. Mater. Res. A 2007, 82, 498–508. [Google Scholar] [CrossRef] [PubMed]
- Cassini-Vieira, P.; de Carvalho Santuchi, M.; da Silva, R.F.; Russo, R.C.; Araujo, F.A.; Dos Santos, R.A.S.; Andrade, S.P.; Teixeira, M.M.; Barcelos, L.S. Lack of interferon-gamma attenuates foreign body reaction to subcutaneous implants in mice. J. Biomed. Mater. Res. A 2018, 106, 2243–2250. [Google Scholar] [CrossRef] [PubMed]
- de Lazari, M.G.T.; Pereira, L.X.; Orellano, L.A.A.; Scheuermann, K.; Machado, C.T.; Vasconcelos, A.C.; Andrade, S.P.; Campos, P.P. Sodium Butyrate Downregulates Implant-Induced Inflammation in Mice. Inflammation 2020, 43, 1259–1268. [Google Scholar] [CrossRef]
- Sicari, B.M.; Agrawal, V.; Siu, B.F.; Medberry, C.J.; Dearth, C.L.; Turner, N.J.; Badylak, S.F. A murine model of volumetric muscle loss and a regenerative medicine approach for tissue replacement. Tissue Eng. Part A 2012, 18, 1941–1948. [Google Scholar] [CrossRef]
- Meng, X.M.; Nikolic-Paterson, D.J.; Lan, H.Y. TGF-beta: The master regulator of fibrosis. Nat. Rev. Nephrol. 2016, 12, 325–338. [Google Scholar] [CrossRef]
- Stojadinovic, O.; Brem, H.; Vouthounis, C.; Lee, B.; Fallon, J.; Stallcup, M.; Merchant, A.; Galiano, R.D.; Tomic-Canic, M. Molecular pathogenesis of chronic wounds: The role of beta-catenin and c-myc in the inhibition of epithelialization and wound healing. Am. J. Pathol. 2005, 167, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Kaji, K.; Yoshiji, H.; Ikenaka, Y.; Noguchi, R.; Aihara, Y.; Douhara, A.; Moriya, K.; Kawaratani, H.; Shirai, Y.; Yoshii, J.; et al. Dipeptidyl peptidase-4 inhibitor attenuates hepatic fibrosis via suppression of activated hepatic stellate cell in rats. J. Gastroenterol. 2014, 49, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, T. Extracellular Interactions between Fibulins and Transforming Growth Factor (TGF)-beta in Physiological and Pathological Conditions. Int. J. Mol. Sci. 2018, 19, 2787. [Google Scholar] [CrossRef]
- Liang, N.E.; Griffin, M.F.; Berry, C.E.; Parker, J.B.; Downer, M.A.; Wan, D.C.; Longaker, M.T. Attenuating Chronic Fibrosis: Decreasing Foreign Body Response with Acellular Dermal Matrix. Tissue Eng. Part B Rev. 2023, 29, 671–680. [Google Scholar] [CrossRef]
- Rinkevich, Y.; Walmsley, G.G.; Hu, M.S.; Maan, Z.N.; Newman, A.M.; Drukker, M.; Januszyk, M.; Krampitz, G.W.; Gurtner, G.C.; Lorenz, H.P.; et al. Skin fibrosis. Identification and isolation of a dermal lineage with intrinsic fibrogenic potential. Science 2015, 348, aaa2151. [Google Scholar] [CrossRef]
- Soare, A.; Gyorfi, H.A.; Matei, A.E.; Dees, C.; Rauber, S.; Wohlfahrt, T.; Chen, C.-W.; Ludolph, I.; Horch, R.E.; Bäuerle, T.; et al. Dipeptidylpeptidase 4 as a Marker of Activated Fibroblasts and a Potential Target for the Treatment of Fibrosis in Systemic Sclerosis. Arthritis Rheumatol. 2020, 72, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.C.; Sung, P.H.; Yang, Y.H.; Chiang, J.Y.; Yip, H.K.; Yang, C.C. Dipeptidyl peptidase 4 promotes peritoneal fibrosis and its inhibitions prevent failure of peritoneal dialysis. Commun. Biol. 2021, 4, 144. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Cooley, M.A.; Jarnicki, A.G.; Hsu, A.C.; Nair, P.M.; Haw, T.J.; Fricker, M.; Gellatly, S.L.; Kim, R.Y.; Inman, M.D.; et al. Fibulin-1 regulates the pathogenesis of tissue remodeling in respiratory diseases. JCI Insight 2016, 1, e86380. [Google Scholar] [CrossRef]
- Veiseh, O.; Vegas, A.J. Domesticating the foreign body response: Recent advances and applications. Adv. Drug Deliv. Rev. 2019, 144, 148–161. [Google Scholar] [CrossRef]
- Meckmongkol, T.T.; Harmon, R.; McKeown-Longo, P.; Van De Water, L. The fibronectin synergy site modulates TGF-beta-dependent fibroblast contraction. Biochem. Biophys. Res. Commun. 2007, 360, 709–714. [Google Scholar] [CrossRef] [PubMed]
- Du, R.; Liu, B.; Zhou, L.; Wang, D.; He, X.; Xu, X.; Zhang, L.; Niu, C.; Liu, S. Downregulation of annexin A3 inhibits tumor metastasis and decreases drug resistance in breast cancer. Cell Death Dis. 2018, 9, 126. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Ji, J.; Hou, J.; Tan, Y.; Han, X. Identification of Key Candidate Genes Involved in the Progression of Idiopathic Pulmonary Fibrosis. Molecules 2021, 26, 1123. [Google Scholar] [CrossRef] [PubMed]
- Fourcot, A.; Couchie, D.; Chobert, M.N.; Zafrani, E.S.; Mavier, P.; Laperche, Y.; Brouillet, A. Gas6 deficiency prevents liver inflammation, steatohepatitis, and fibrosis in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, G1043–G1053. [Google Scholar] [CrossRef] [PubMed]
- Rizzi, M.; Tonello, S.; D’Onghia, D.; Sainaghi, P.P. Gas6/TAM Axis Involvement in Modulating Inflammation and Fibrosis in COVID-19 Patients. Int. J. Mol. Sci. 2023, 24, 951. [Google Scholar] [CrossRef] [PubMed]
- Nassar, M.; Tabib, Y.; Capucha, T.; Mizraji, G.; Nir, T.; Pevsner-Fischer, M.; Zilberman-Schapira, G.; Heyman, O.; Nussbaum, G.; Bercovier, H.; et al. GAS6 is a key homeostatic immunological regulator of host-commensal interactions in the oral mucosa. Proc. Natl. Acad. Sci. USA 2017, 114, E337–E346. [Google Scholar] [CrossRef] [PubMed]
- Herro, R.; Da Silva Antunes, R.; Aguilera, A.R.; Tamada, K.; Croft, M. Tumor necrosis factor superfamily 14 (LIGHT) controls thymic stromal lymphopoietin to drive pulmonary fibrosis. J. Allergy Clin. Immunol. 2015, 136, 757–768. [Google Scholar] [CrossRef] [PubMed]
- Herro, R.; Antunes, R.D.S.; Aguilera, A.R.; Tamada, K.; Croft, M. The Tumor Necrosis Factor Superfamily Molecule LIGHT Promotes Keratinocyte Activity and Skin Fibrosis. J. Invest. Dermatol. 2015, 135, 2109–2118. [Google Scholar] [CrossRef] [PubMed]
- Steele, H.; Cheng, J.; Willicut, A.; Dell, G.; Breckenridge, J.; Culberson, E.; Ghastine, A.; Tardif, V.; Herro, R. TNF superfamily control of tissue remodeling and fibrosis. Front. Immunol. 2023, 14, 1219907. [Google Scholar] [CrossRef]
- Rabhi, N.; Desevin, K.; Belkina, A.C.; Tilston-Lunel, A.; Varelas, X.; Layne, M.D.; Farmer, S.R. Obesity-induced senescent macrophages activate a fibrotic transcriptional program in adipocyte progenitors. Life Sci. Alliance 2022, 5, e202101286. [Google Scholar] [CrossRef]
- Cimini, M.; Kishore, R. Role of Podoplanin-Positive Cells in Cardiac Fibrosis and Angiogenesis after Ischemia. Front. Physiol. 2021, 12, 667278. [Google Scholar] [CrossRef] [PubMed]
- Layton, T.B.; Williams, L.; McCann, F.; Zhang, M.; Fritzsche, M.; Colin-York, H.; Cabrita, M.; Ng, M.T.H.; Feldmann, M.; Sansom, S.N.; et al. Cellular census of human fibrosis defines functionally distinct stromal cell types and states. Nat. Commun. 2020, 11, 2768. [Google Scholar] [CrossRef] [PubMed]
- Astarita, J.L.; Acton, S.E.; Turley, S.J. Podoplanin: Emerging functions in development, the immune system, and cancer. Front. Immunol. 2012, 3, 283. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Wang, L.; Yang, J.; Chen, X.; Guo, X.; Xu, K.; Wang, N.; Zhao, W.; Xia, C.; Lian, H.; et al. Endothelial cell-derived MMP19 promotes pulmonary fibrosis by inducing E(nd)MT and monocyte infiltration. Cell Commun. Signal. 2023, 21, 56. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.I.; Song, M.K.; Yuk, J.E.; Seo, H.J.; Lee, K. Establishment of an artificial particulate matter-induced lung disease model through analyzing pathological changes and transcriptomic profiles in mice. Sci. Rep. 2023, 13, 5955. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Livingston, M.J.; Dong, G.; Wei, Q.; Zhang, M.; Mei, S.; Zhu, J.; Zhang, C.; Dong, Z. HIF-1 contributes to autophagy activation via BNIP3 to facilitate renal fibrosis in hypoxia in vitro and UUO in vivo. Am. J. Physiol. Cell Physiol. 2024, 326, C935–C947. [Google Scholar] [CrossRef] [PubMed]
- Welford, S.M.; Bedogni, B.; Gradin, K.; Poellinger, L.; Broome Powell, M.; Giaccia, A.J. HIF1alpha delays premature senescence through the activation of MIF. Genes Dev. 2006, 20, 3366–3371. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Yi, H.; Huang, X.; Lin, G.; Kuang, Y.; Guo, Y.; Xie, C. Inhibition of macrophage migration inhibitory factor (MIF) as a therapeutic target in bleomycin-induced pulmonary fibrosis rats. Am. J. Physiol. Lung Cell Mol. Physiol. 2021, 321, L6–L16. [Google Scholar] [CrossRef]
- Aumailley, M. The laminin family. Cell Adh. Migr. 2013, 7, 48–55. [Google Scholar] [CrossRef]
- Sikder, M.K.U.; Tong, W.; Pingle, H.; Kingshott, P.; Needham, K.; Shivdasani, M.N.; Fallon, J.B.; Seligman, P.; Ibbotson, M.R.; Prawer, S.; et al. Laminin coated diamond electrodes for neural stimulation. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 118, 111454. [Google Scholar] [CrossRef]
- Azemi, E.; Gobbel, G.T.; Cui, X.T. Seeding neural progenitor cells on silicon-based neural probes. J. Neurosurg. 2010, 113, 673–681. [Google Scholar] [CrossRef]
- Oyane, A.; Hyodo, K.; Uchida, M.; Sogo, Y.; Ito, A. Preliminary in vivo study of apatite and laminin-apatite composite layers on polymeric percutaneous implants. J. Biomed. Mater. Res. B Appl. Biomater. 2011, 97, 96–104. [Google Scholar] [CrossRef]
- Cao, Y.; Zhang, B.; Croll, T.; Rolfe, B.E.; Campbell, J.H.; Campbell, G.R.; Martin, D.; Cooper-White, J.J. Engineering tissue tubes using novel multilayered scaffolds in the rat peritoneal cavity. J. Biomed. Mater. Res. A 2008, 87, 719–727. [Google Scholar] [CrossRef]
- Sasaki, F.; Hata, Y.; Hamada, H.; Takahashi, H.; Uchino, J. Laminin and procollagen-III-peptide as a serum marker for hepatic fibrosis in congenital biliary atresia. J. Pediatr. Surg. 1992, 27, 700–703. [Google Scholar] [CrossRef]
- Mak, K.M.; Mei, R. Basement Membrane Type IV Collagen and Laminin: An Overview of Their Biology and Value as Fibrosis Biomarkers of Liver Disease. Anat. Rec. 2017, 300, 1371–1390. [Google Scholar] [CrossRef]
- Younesi, S.; Parsian, H. Diagnostic accuracy of glycoproteins in the assessment of liver fibrosis: A comparison between laminin, fibronectin, and hyaluronic acid. Turk. J. Gastroenterol. 2019, 30, 524–531. [Google Scholar] [CrossRef]
- Nevo, Y.; Halevy, O.; Genin, O.; Moshe, I.; Turgeman, T.; Harel, M.; Biton, E.; Reif, S.; Pines, M. Fibrosis inhibition and muscle histopathology improvement in laminin-α2-deficient mice. Muscle Nerve 2010, 42, 218–229. [Google Scholar] [CrossRef]
- Morales-Nebreda, L.I.; Rogel, M.R.; Eisenberg, J.L.; Hamill, K.J.; Soberanes, S.; Nigdelioglu, R.; Chi, M.; Cho, T.; Radigan, K.A.; Ridge, K.M.; et al. Lung-specific loss of α3 laminin worsens bleomycin-induced pulmonary fibrosis. Am. J. Respir. Cell Mol. Biol. 2015, 52, 503–512. [Google Scholar] [CrossRef]
- Patten, J.; Wang, K. Fibronectin in development and wound healing. Adv. Drug Deliv. Rev. 2021, 170, 353–368. [Google Scholar] [CrossRef]
- Kao, W.J.; Lee, D.; Schense, J.C.; Hubbell, J.A. Fibronectin modulates macrophage adhesion and FBGC formation: The role of RGD, PHSRN, and PRRARV domains. J. Biomed. Mater. Res. 2001, 55, 79–88. [Google Scholar] [CrossRef]
- Keselowsky, B.G.; Bridges, A.W.; Burns, K.L.; Tate, C.C.; Babensee, J.E.; LaPlaca, M.C.; García, A.J. Role of plasma fibronectin in the foreign body response to biomaterials. Biomaterials 2007, 28, 3626–3631. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Ma, T.; Lian, X.; Gao, J.; Wang, W.; Weng, W.; Lu, X.; Sun, W.; Cheng, Y.; Fu, Y.; et al. Clopidogrel Reduces Fibronectin Accumulation and Improves Diabetes-Induced Renal Fibrosis. Int. J. Biol. Sci. 2019, 15, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Altrock, E.; Sens, C.; Wuerfel, C.; Vasel, M.; Kawelke, N.; Dooley, S.; Sottile, J.; Nakchbandi, I.A. Inhibition of fibronectin deposition improves experimental liver fibrosis. J. Hepatol. 2015, 62, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Klingberg, F.; Chau, G.; Walraven, M.; Boo, S.; Koehler, A.; Chow, M.L.; Olsen, A.L.; Im, M.; Lodyga, M.; Wells, R.G.; et al. The fibronectin ED-A domain enhances recruitment of latent TGF-β-binding protein-1 to the fibroblast matrix. J. Cell Sci. 2018, 131, jcs201293. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GSE | GSM | Species | Genotype | FBR Model | Condition | Treatment? | Time Point | Anatomic Region | Sorted? |
|---|---|---|---|---|---|---|---|---|---|
| GSE203099 | GSM6153772 | mouse | C57BL/6 | control | Chronic Inflammation Control | N | NA | peritoneum | unsorted |
| GSE203099 | GSM6153773 | mouse | C57BL/6 | sponge | FBR | N | 2 weeks | peritoneum | unsorted |
| GSE203099 | GSM6153774 | mouse | C57BL/6 | sponge | FBR | N | 4 weeks | peritoneum | unsorted |
| GSE203099 | GSM6153775 | mouse | C57BL/6 | silk | FBR | N | 2 weeks | peritoneum | unsorted |
| GSE203099 | GSM6153776 | mouse | C57BL/6 | silk | FBR | N | 4 weeks | peritoneum | unsorted |
| GSE233843 | GSM7437854 | mouse | CD-1 | matrigel only | FBR | N | 5 days | subdermis | unsorted |
| GSE227908 | GSM7110608 | mouse | C57BL/6 | implant | FBR | N | 2 weeks | subdermis | unsorted |
| GSE227908 | GSM7110609 | mouse | C57BL/6 | implant | FBR | N | 4 weeks | subdermis | unsorted |
| GSE175889 | GSM5350798 | mouse | C57BL/6 | ECM implant | FBR | N | 1 week | quadricep | CD45-enriched |
| GSE175889 | GSM5350799 | mouse | C57BL/6 | PCL implant | FBR | N | 1 week | quadricep | CD45-enriched |
| GSE175889 | GSM5350800 | mouse | C57BL/6 | Saline | Surgery Control | N | 1 week | quadricep | CD45-enriched |
| GSE175889 | GSM5350801 | mouse | C57BL/6 | control (sham) | Tissue Control | N | 1 week | quadricep | CD45-enriched |
| GSE175889 | GSM5350802 | mouse | C57BL/6 | ECM implant | FBR | N | 1 week | quadricep | CD45-enriched |
| GSE175889 | GSM5350803 | mouse | C57BL/6 | PCL implant | FBR | N | 1 week | quadricep | CD45-enriched |
| GSE175889 | GSM5350804 | mouse | C57BL/6 | Saline | Surgery Control | N | 1 week | quadricep | CD45-enriched |
| GSE175889 | GSM5350805 | mouse | C57BL/6 | control (sham) | Tissue Control | N | 1 week | quadricep | CD45-enriched |
| GSE175889 | GSM5350806 | mouse | C57BL/6 | ECM implant | FBR | N | 1 week | quadricep | Fibroblasts;(CD45-CD19-CD31-CD29+) |
| GSE175889 | GSM5350807 | mouse | C57BL/6 | PCL implant | FBR | N | 1 week | quadricep | Fibroblasts;(CD45-CD19-CD31-CD29+) |
| GSE175889 | GSM5350808 | mouse | C57BL/6 | Saline | Surgery Control | N | 1 week | quadricep | Fibroblasts;(CD45-CD19-CD31-CD29+) |
| GSE175889 | GSM5350809 | mouse | C57BL/6 | control (sham) | Tissue Control | N | 1 week | quadricep | Fibroblasts;(CD45-CD19-CD31-CD29+) |
| GSE175889 | GSM5350810 | mouse | C57BL/6 | ECM implant | FBR | N | 6 weeks | quadricep | Fibroblasts;(CD45-CD19-CD31-CD29+) |
| GSE175889 | GSM5350811 | mouse | C57BL/6 | PCL implant | FBR | N | 6 weeks | quadricep | Fibroblasts;(CD45-CD19-CD31-CD29+) |
| GSE175889 | GSM5350812 | mouse | C57BL/6 | Saline | Surgery Control | N | 6 weeks | quadricep | Fibroblasts;(CD45-CD19-CD31-CD29+) |
| GSE175889 | GSM5350813 | mouse | C57BL/6 | control (sham) | Tissue Control | N | 6 weeks | quadricep | Fibroblasts;(CD45-CD19-CD31-CD29+) |
| GSE175889 | GSM5350814 | mouse | C57BL/6 | ECM implant | FBR | N | 1 week | quadricep | Macrophages from previously published dataset: (CD45+F4/80hi+Ly6c+CD64+) |
| GSE175889 | GSM5350815 | mouse | C57BL/6 | PCL implant | Surgery Control | N | 1 week | quadricep | Macrophages from previously published dataset: (CD45+F4/80hi+Ly6c+CD64+) |
| GSE175889 | GSM5350816 | mouse | C57BL/6 | Saline | Tissue Control | N | 1 week | quadricep | Macrophages from previously published dataset: (CD45+F4/80hi+Ly6c+CD64+) |
| Timp1+ | Col4a1+ | Fcer1g+ | Stmn1+ | Fmod+ | Sfrp5+ | |
|---|---|---|---|---|---|---|
| Wiki |
|
| X | X |
|
|
| KEGG |
|
| Complement and coagulation cascades | X |
|
|
| NCI |
|
| X | X |
|
|
| Biological |
|
| X | X |
|
|
| Cellular |
|
| X | X |
|
|
| Molecular |
|
| X | X |
| X |
| Rbm42− | Ccl8+ | Gpnmb+ | Plac8+ | Retnla+ | |
|---|---|---|---|---|---|
| Wiki | X |
| X |
|
|
| KEGG |
|
| X | X | X |
| NCI | X |
| X | X | X |
| Biological | X | X | X | X | X |
| Cellular |
|
|
| X |
|
| Molecular |
|
|
| X |
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, N.E.; Parker, J.B.; Lu, J.M.; Januszyk, M.; Wan, D.C.; Griffin, M.; Longaker, M.T. Understanding the Foreign Body Response via Single-Cell Meta-Analysis. Biology 2024, 13, 540. https://doi.org/10.3390/biology13070540
Liang NE, Parker JB, Lu JM, Januszyk M, Wan DC, Griffin M, Longaker MT. Understanding the Foreign Body Response via Single-Cell Meta-Analysis. Biology. 2024; 13(7):540. https://doi.org/10.3390/biology13070540
Chicago/Turabian StyleLiang, Norah E., Jennifer B. Parker, John M. Lu, Michael Januszyk, Derrick C. Wan, Michelle Griffin, and Michael T. Longaker. 2024. "Understanding the Foreign Body Response via Single-Cell Meta-Analysis" Biology 13, no. 7: 540. https://doi.org/10.3390/biology13070540
APA StyleLiang, N. E., Parker, J. B., Lu, J. M., Januszyk, M., Wan, D. C., Griffin, M., & Longaker, M. T. (2024). Understanding the Foreign Body Response via Single-Cell Meta-Analysis. Biology, 13(7), 540. https://doi.org/10.3390/biology13070540