
Developmental Cascades Link Maternal–Newborn Skin-to-Skin Contact with Young Adults’ Psychological Symptoms, Oxytocin, and Immunity; Charting Mechanisms of Developmental Continuity from Birth to Adulthood


Abstract
Simple Summary
Abstract
1. Introduction
1.1. Premature Birth and Its Developmental Consequences
1.2. Kangaroo Care (KC) and Mother–Infant Physical Contact
1.3. KC Effects across Development via Three Pathways; Maternal Mood, Child Attention and Regulation, and Dyadic Synchrony
1.4. Three Mechanisms of Developmental Continuity
1.5. The Current Study
2. Materials and Methods
2.1. Neonatal Period: Cohort and KC Intervention
2.2. Dyadic Synchrony
2.3. Focused Attention at 24 Months
2.4. Executive Functions Score
2.5. Saliva Samples Collection and Measuring
2.6. Anxiety/Depressive Symptoms
2.7. Statistical Analysis
2.8. Structural Equation Modeling
2.9. Preregistration
3. Results
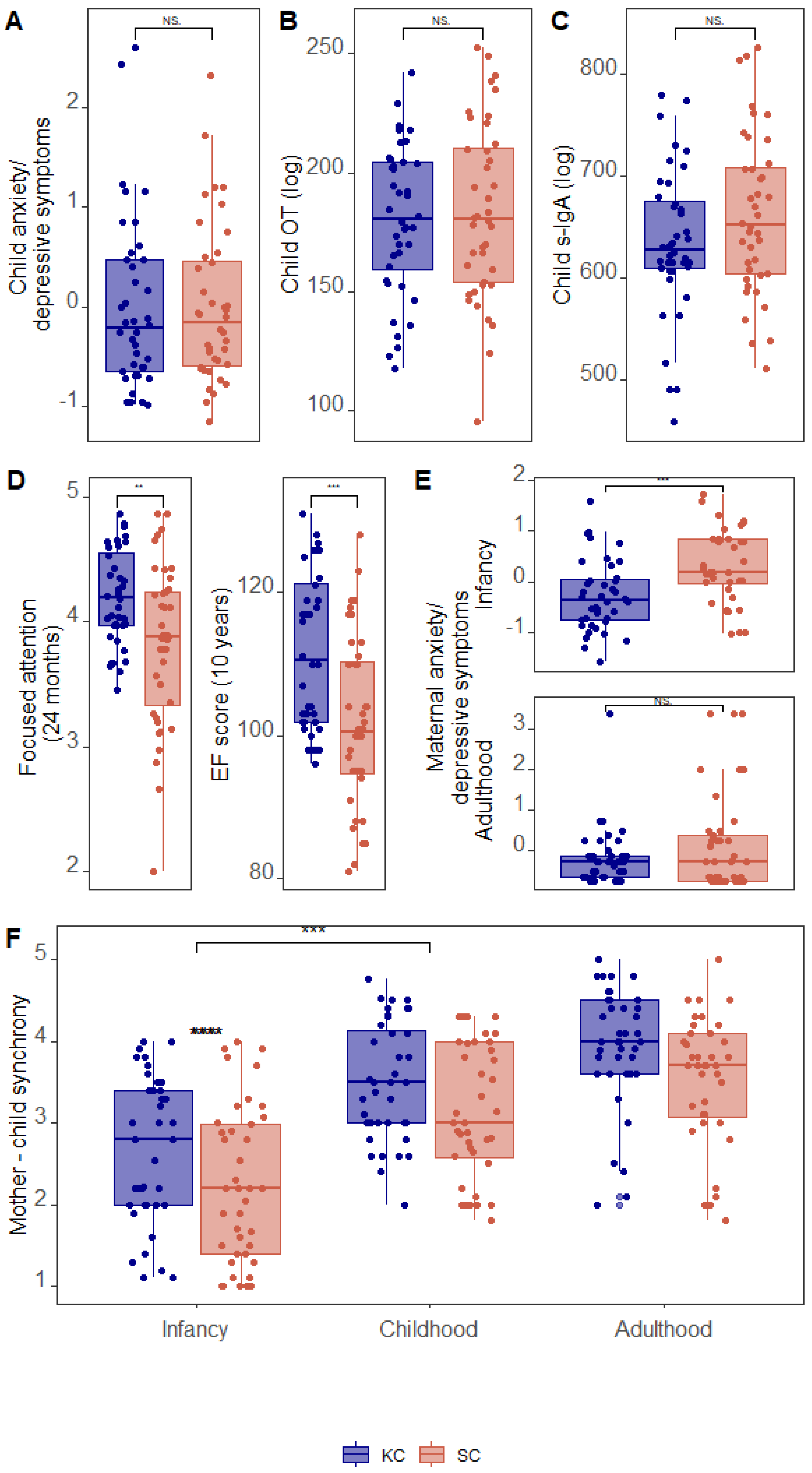
3.1. Group Differences in Study Variables
3.2. Correlations among Study Variables
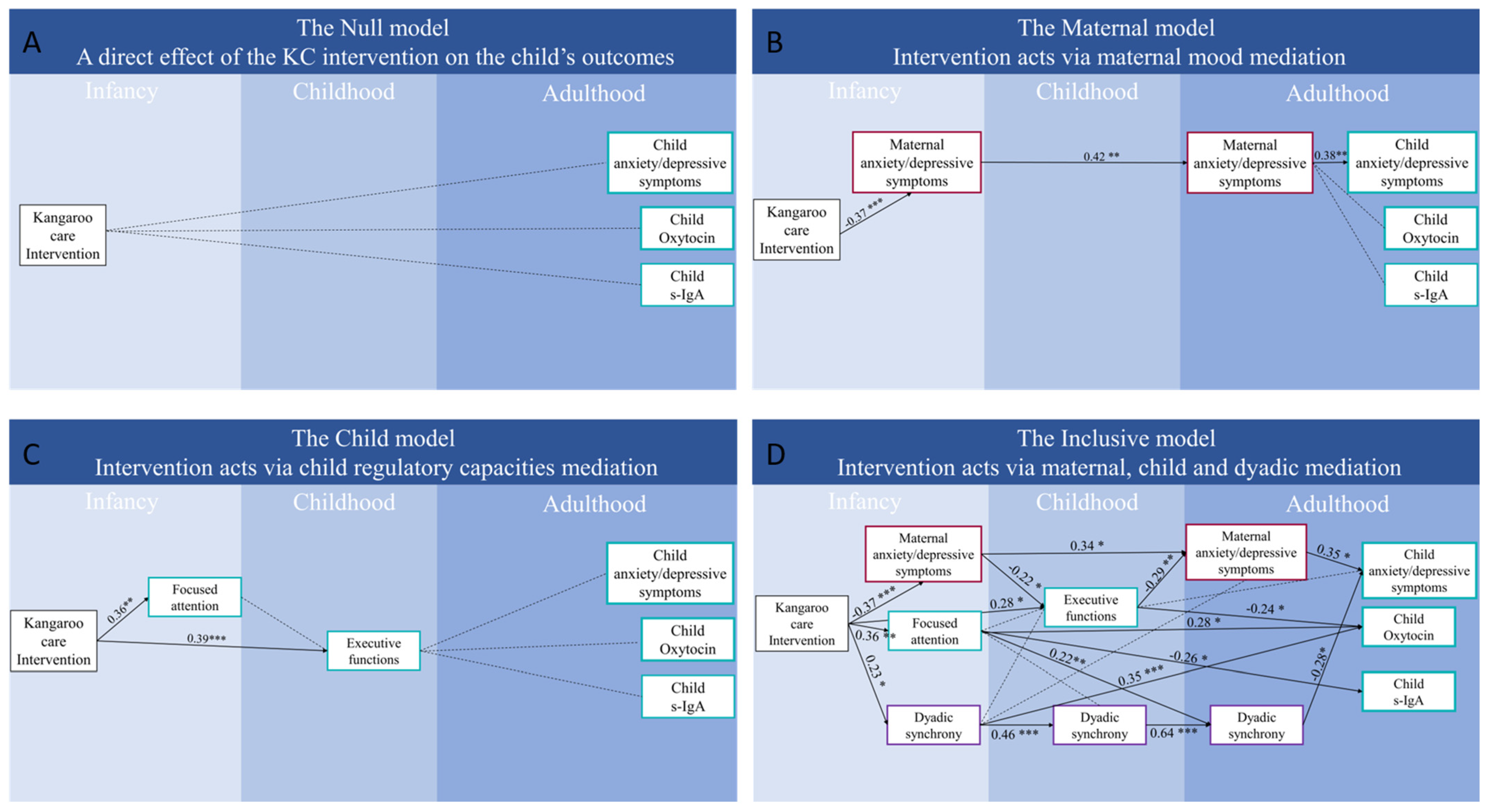
3.3. Path Models Linking Kangaroo Care with Adult Outcome via Developmental Cascades
4. Discussion
4.1. Effects of a Birth Intervention on Adult Outcome via Three Mechanisms of Continuity
4.2. KC Impact on Adult Outcome via “Maternal Effects”
4.3. KC Impact on Adult Outcome via “Child Effects”
4.4. KC Impacts on Adult Outcome Via “Dyadic Effects”
4.5. KC, Oxytocin, and S-IgA
5. Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Allen, M.C.; Cristofalo, E.; Kim, C. Preterm Birth: Transition to Adulthood. Dev. Disabil. Res. Rev. 2010, 16, 323–335. [Google Scholar] [CrossRef]
- Sarda, S.P.; Sarri, G.; Siffel, C. Global Prevalence of Long-Term Neurodevelopmental Impairment Following Extremely Preterm Birth: A Systematic Literature Review. J. Int. Med. Res. 2021, 49, 1–21. [Google Scholar] [CrossRef]
- Aarnoudse-Moens, C.S.H.; Duivenvoorden, H.J.; Weisglas-Kuperus, N.; Van Goudoever, J.B.; Oosterlaan, J. The Profile of Executive Function in Very Preterm Children at 4 to 12 Years. Dev. Med. Child Neurol. 2012, 54, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Edgin, J.O.; Inder, T.E.; Anderson, P.J.; Hood, K.M.; Clark, C.A.C.; Woodward, L.J. Executive Functioning in Preschool Children Born Very Preterm: Relationship with Early White Matter Pathology. J. Int. Neuropsychol. Soc. 2008, 14, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Witt, A.; Theurel, A.; Tolsa, C.B.; Lejeune, F.; Fernandes, L.; van Hanswijck de Jonge, L.; Monnier, M.; Graz, M.B.; Barisnikov, K.; Gentaz, E.; et al. Emotional and Effortful Control Abilities in 42-Month-Old Very Preterm and Full-Term Children. Early Hum. Dev. 2014, 90, 565–569. [Google Scholar] [CrossRef]
- Healy, E.; Reichenberg, A.; Nam, K.W.; Allin, M.P.G.; Walshe, M.; Rifkin, L.; Murray, S.R.M.; Nosarti, C. Preterm Birth and Adolescent Social Functioning-Alterations in Emotion-Processing Brain Areas. J. Pediatr. 2013, 163, 1596–1604. [Google Scholar] [CrossRef]
- Zmyj, N.; Witt, S.; Weitkämper, A.; Neumann, H.; Lücke, T. Social Cognition in Children Born Preterm: A Perspective on Future Research Directions. Front. Psychol. 2017, 8, 455. [Google Scholar] [CrossRef]
- Witt, S.; Weitkämper, A.; Neumann, H.; Lücke, T.; Zmyj, N. Delayed Theory of Mind Development in Children Born Preterm: A Longitudinal Study. Early Hum. Dev. 2018, 127, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Bhutta, A.; Cleves, M.; Casey, P. Cognitive and Behavioral Outcomes of School-Aged Children Who Were Born Preterm: A Meta-Analysis. JAMA 2002, 288, 728–737. [Google Scholar] [CrossRef] [PubMed]
- Hack, M.; Youngstrom, E.A.; Cartar, L.; Schluchter, M.; Taylor, H.G.; Flannery, D.; Klein, N.; Borawski, E.; Gerry Taylor, H.; Flannery, D.; et al. Behavioral Outcomes and Evidence of Psychopathology among Very Low Birth Weight Infants at Age 20 Years. Pediatrics 2004, 114, 932–940. [Google Scholar] [CrossRef]
- Fitzallen, G.C.; Sagar, Y.K.; Taylor, H.G.; Bora, S. Anxiety and Depressive Disorders in Children Born Preterm: A Meta-Analysis. J. Dev. Behav. Pediatr. 2021, 42, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Loret De Mola, C.; De França, G.V.A.; De Avila Quevedo, L.; Horta, B.L. Low Birth Weight, Preterm Birth and Small for Gestational Age Association with Adult Depression: Systematic Review and Meta-Analysis. Br. J. Psychiatry 2014, 205, 340–347. [Google Scholar] [CrossRef]
- Burnett, A.C.; Anderson, P.J.; Cheong, J.; Doyle, L.W.; Davey, C.G.; Wood, S.J. Prevalence of Psychiatric Diagnoses in Preterm and Full-Term Children, Adolescents and Young Adults: A Meta-Analysis. Psychol. Med. 2011, 41, 2463–2474. [Google Scholar] [CrossRef]
- Anderson, P.J.; de Miranda, D.M.; Albuquerque, M.R.; Indredavik, M.S.; Evensen, K.A.I.; Van Lieshout, R.; Saigal, S.; Taylor, H.G.; Raikkonen, K.; Kajantie, E.; et al. Psychiatric Disorders in Individuals Born Very Preterm/Very Low-Birth Weight: An Individual Participant Data (IPD) Meta-Analysis. eClinicalMedicine 2021, 42, 101216. [Google Scholar] [CrossRef]
- Finken, M.J.J.; van der Voorn, B.; Hollanders, J.J.; Ruys, C.A.; de Waard, M.; van Goudoever, J.B.; Rotteveel, J. Programming of the Hypothalamus-Pituitary-Adrenal Axis by Very Preterm Birth. Ann. Nutr. Metab. 2017, 70, 170–174. [Google Scholar] [CrossRef]
- Mörelius, E.; He, H.-G.; Shorey, S. Salivary Cortisol Reactivity in Preterm Infants in Neonatal Intensive Care: An Integrative Review. Int. J. Environ. Res. Public Health 2016, 13, 337. [Google Scholar] [CrossRef]
- Wadhwa, P.D. Psychoneuroendocrine Processes in Human Pregnancy Influence Fetal Development and Health. Psychoneuroendocrinology 2005, 30, 724–743. [Google Scholar] [CrossRef]
- Brummelte, S.; Grunau, R.E.; Zaidman-Zait, A.; Weinberg, J.; Nordstokke, D.; Cepeda, I.L. Cortisol Levels in Relation to Maternal Interaction and Child Internalizing Behavior in Preterm and Full-Term Children at 18 Months Corrected Age. Dev. Psychobiol. 2011, 53, 184–195. [Google Scholar] [CrossRef] [PubMed]
- Feldman, R.; Weller, A.; Sirota, L.; Eidelman, A.I. Skin-to-Skin Contact (Kangaroo Care) Promotes Self-Regulation in Premature Infants: Sleep-Wake Cyclicity, Arousal Modulation, and Sustained Exploration. Dev. Psychol. 2002, 38, 194–207. [Google Scholar] [CrossRef]
- Mendonça, M.; Bilgin, A.; Wolke, D. Association of Preterm Birth and Low Birth Weight With Romantic Partnership, Sexual Intercourse, and Parenthood in Adulthood: A Systematic Review and Meta-Analysis. JAMA Netw. Open 2019, 2, e196961. [Google Scholar] [CrossRef] [PubMed]
- Reyes, L.M.; Jaekel, J.; Bartmann, P.; Wolke, D. Peer Relationship Trajectories in Very Preterm and Term Individuals from Childhood to Early Adulthood. J. Dev. Behav. Pediatr. 2021, 42, 621–630. [Google Scholar] [CrossRef]
- Reyes, L.M.; Jaekel, J.; Heuser, K.M.; Wolke, D. Developmental Cascades of Social Inhibition and Friendships in Preterm and Full-Term Children. Infant Child Dev. 2019, 28, e2165. [Google Scholar] [CrossRef]
- Blencowe, H.; Cousens, S.; Chou, D.; Oestergaard, M.; Say, L.; Moller, A.-B.; Kinney, M.; Lawn, J.; The Born Too Soon Preterm Birth Action Group. Born Too Soon: The Global Epidemiology of 15 Million Preterm Births. Reprod. Health 2013, 10, S2. [Google Scholar] [CrossRef]
- Anderson, P.J.; Treyvaud, K.; Neil, J.J.; Cheong, J.L.Y.; Hunt, R.W.; Thompson, D.K.; Lee, K.J.; Doyle, L.W.; Inder, T.E. Associations of Newborn Brain Magnetic Resonance Imaging with Long-Term Neurodevelopmental Impairments in Very Preterm Children. J. Pediatr. 2017, 187, 58–65.e1. [Google Scholar] [CrossRef] [PubMed]
- Kline, J.E.; Illapani, V.S.P.; He, L.; Altaye, M.; Logan, J.W.; Parikh, N.A. Early Cortical Maturation Predicts Neurodevelopment in Very Preterm Infants. Arch. Dis. Child.-Fetal Neonatal Ed. 2020, 105, 460–465. [Google Scholar] [CrossRef]
- Morin, E.L.; Howell, B.R.; Meyer, J.S.; Sanchez, M.M. Effects of Early Maternal Care on Adolescent Attention Bias to Threat in Nonhuman Primates. Dev. Cogn. Neurosci. 2019, 38, 100643. [Google Scholar] [CrossRef]
- Addabbo, M.; Licht, V.; Turati, C. Past and Present Experiences with Maternal Touch Affect Infants’ Attention toward Emotional Faces. Infant Behav. Dev. 2021, 63, 101558. [Google Scholar] [CrossRef]
- Scatliffe, N.; Casavant, S.; Vittner, D.; Cong, X. Oxytocin and Early Parent-Infant Interactions: A Systematic Review. Int. J. Nurs. Sci. 2019, 6, 445–453. [Google Scholar] [CrossRef]
- Swain, J.E.; Kim, P.; Spicer, J.; Ho, S.S.; Dayton, C.J.; Elmadih, A.; Abel, K.M. Approaching the Biology of Human Parental Attachment: Brain Imaging, Oxytocin and Coordinated Assessments of Mothers and Fathers. Brain Res. 2014, 1580, 78–101. [Google Scholar] [CrossRef] [PubMed]
- Feldman, R. Sensitive Periods in Human Social Development: New Insights from Research on Oxytocin, Synchrony, and High-Risk Parenting. Dev. Psychopathol. 2015, 27, 369–395. [Google Scholar] [CrossRef]
- Barry, E.S. Co-Sleeping as a Proximal Context for Infant Development: The Importance of Physical Touch. Infant Behav. Dev. 2019, 57, 101385. [Google Scholar] [CrossRef]
- Wang, D.; Levine, J.L.S.; Avila-Quintero, V.; Bloch, M.; Kaffman, A. Systematic Review and Meta-Analysis: Effects of Maternal Separation on Anxiety-like Behavior in Rodents. Transl. Psychiatry 2020, 10, 174. [Google Scholar] [CrossRef]
- Nishi, M.; Horii-Hayashi, N.; Sasagawa, T. Effects of Early Life Adverse Experiences on the Brain: Implications from Maternal Separation Models in Rodents. Front. Neurosci. 2014, 8, 186. [Google Scholar] [CrossRef] [PubMed]
- Givrad, S.; Hartzell, G.; Scala, M. Promoting Infant Mental Health in the Neonatal Intensive Care Unit (NICU): A Review of Nurturing Factors and Interventions for NICU Infant-Parent Relationships. Early Hum. Dev. 2021, 154, 105281. [Google Scholar] [CrossRef]
- Cinar, N.; Zengin, H.; Hur, G.; Eroğlu, A.; Karakaya, O.; Kolukisa, T. The Effects of Kangaroo Mother Care on Physiological Parameters of Premature in NICU: Systematic Review and Meta-Analysis. J. Pediatr. Nurs. 2022, 16, 2920–2925. [Google Scholar] [CrossRef]
- Campbell-Yeo, M.; Celeste Johnston, C.; Benoit, B.; Disher, T.; Caddell, K.; Vincer, M.; Walker, C.D.; Latimer, M.; Streiner, D.L.; Inglis, D. Sustained Efficacy of Kangaroo Care for Repeated Painful Procedures over Neonatal Intensive Care Unit Hospitalization: A Single-Blind Randomized Controlled Trial. Pain 2019, 160, 2580–2588. [Google Scholar] [CrossRef]
- Scime, N.V.; Gavarkovs, A.G.; Chaput, K.H. The Effect of Skin-to-Skin Care on Postpartum Depression among Mothers of Preterm or Low Birthweight Infants: A Systematic Review and Meta-Analysis. J. Affect. Disord. 2019, 253, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Bigelow, A.; Power, M.; Maclellan-Peters, J.; Alex, M.; Mcdonald, C. Effect of Mother/Infant Skin-to-Skin Contact on Postpartum Depressive Symptoms and Maternal Physiological Stress. J. Obstet. Gynecol. Neonatal Nurs. 2012, 41, 369–382. [Google Scholar] [CrossRef]
- Feldman, R.; Eidelman, A.I.; Sirota, L.; Weller, A. Comparison of Skin-to-Skin (Kangaroo) and Traditional Care: Parenting Outcomes and Preterm Infant Development. Pediatrics 2002, 110, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Feldman, R.; Rosenthal, Z.; Eidelman, A.I. Maternal-Preterm Skin-to-Skin Contact Enhances Child Physiologic Organization and Cognitive Control across the First 10 Years of Life. Biol. Psychiatry 2014, 75, 56–64. [Google Scholar] [CrossRef]
- Feldman, R.; Weller, A.; Sirota, L.; Eidelman, A.I. Testing a Family Intervention Hypothesis: The Contribution of Mother-Infant Skin-to-Skin Contact (Kangaroo Care) to Family Interaction, Proximity, and Touch. J. Fam. Psychol. 2003, 17, 94–107. [Google Scholar] [CrossRef]
- Rogers, C.E.; Kidokoro, H.; Wallendorf, M.; Inder, T.E. Identifying Mothers of Very Preterm Infants At-Risk for Postpartum Depression and Anxiety before Discharge. J. Perinatol. 2013, 33, 171–176. [Google Scholar] [CrossRef]
- Ionio, C.; Colombo, C.; Brazzoduro, V.; Mascheroni, E.; Confalonieri, E.; Castoldi, F.; Lista, G. Mothers and Fathers in Nicu: The Impact of Preterm Birth on Parental Distress. Eur. J. Psychol. 2016, 12, 604–621. [Google Scholar] [CrossRef]
- Yates, R.; Anderson, P.J.; Lee, K.J.; Doyle, L.W.; Cheong, J.L.Y.; Pace, C.C.; Spittle, A.J.; Spencer-Smith, M.; Treyvaud, K. Maternal Mental Health Disorders Following Very Preterm Birth at 5 Years Post-Birth. J. Pediatr. Psychol. 2022, 47, 327–336. [Google Scholar] [CrossRef]
- Goodman, S.H.; Rouse, M.H.; Connell, A.M.; Broth, M.R.; Hall, C.M.; Heyward, D. Maternal Depression and Child Psychopathology: A Meta-Analytic Review. Clin. Child Fam. Psychol. Rev. 2011, 14, 589–609. [Google Scholar] [CrossRef]
- Herba, C.M.; Glover, V.; Ramchandani, P.G.; Rondon, M.B. Maternal Depression and Mental Health in Early Childhood: An Examination of Underlying Mechanisms in Low-Income and Middle-Income Countries. Lancet Psychiatry 2016, 3, 983–992. [Google Scholar] [CrossRef]
- Feldman, R.; Eidelman, A.I. Skin-to-Skin Contact (Kangaroo Care) Accelerates Autonomic and Neurobehavioural Maturation in Preterm Infants. Dev. Med. Child Neurol. 2007, 45, 274–281. [Google Scholar] [CrossRef]
- Ritter, B.C.; Nelle, M.; Perrig, W.; Steinlin, M.; Everts, R. Executive Functions of Children Born Very Preterm--Deficit or Delay? Eur. J. Pediatr. 2013, 172, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Sammallahti, S.; Heinonen, K.; Andersson, S.; Lahti, M.; Pirkola, S.; Lahti, J.; Pesonen, A.K.; Lano, A.; Wolke, D.; Eriksson, J.G.; et al. Growth after Late-Preterm Birth and Adult Cognitive, Academic, and Mental Health Outcomes. Pediatr. Res. 2017, 81, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Farroni, T.; Della Longa, L.; Valori, I. The Self-Regulatory Affective Touch: A Speculative Framework for the Development of Executive Functioning. Curr. Opin. Behav. Sci. 2022, 43, 167–173. [Google Scholar] [CrossRef]
- Kroll, J.; Karolis, V.; Brittain, P.J.; Tseng, C.E.J.; Froudist-Walsh, S.; Murray, R.M.; Nosarti, C. Real-Life Impact of Executive Function Impairments in Adults Who Were Born Very Preterm. J. Int. Neuropsychol. Soc. 2017, 23, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Rickards, A.L.; Kelly, E.A.; Doyle, L.W.; Callanan, C. Cognition, Academic Progress, Behavior and Self-Concept at 14 Years of Very Low Birth Weight Children. J. Dev. Behav. Pediatr. 2001, 22, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Green, S.L.; Phipps, W.D. Interactional Pattern Analysis of Mother-Baby Pairs: Kangaroo Mother Care versus Incubator Care. S. Afr. J. Psychol. 2015, 45, 194–206. [Google Scholar] [CrossRef]
- Feldman, R. Mutual Influences between Child Emotion Regulation and Parent-Child Reciprocity Support Development across the First 10 Years of Life: Implications for Developmental Psychopathology. Dev. Psychopathol. 2015, 27, 1007–1023. [Google Scholar] [CrossRef]
- Feldman, R.; Bamberger, E.; Kanat-Maymon, Y. Parent-Specific Reciprocity from Infancy to Adolescence Shapes Children’s Social Competence and Dialogical Skills. Attach. Hum. Dev. 2013, 15, 407–423. [Google Scholar] [CrossRef]
- Wolke, D.; Eryigit-Madzwamuse, S.; Gutbrod, T. Very Preterm/Very Low Birthweight Infants’ Attachment: Infant and Maternal Characteristics. Arch. Dis. Child. Fetal Neonatal Ed. 2014, 99, F70–F75. [Google Scholar] [CrossRef]
- Sømhovd, M.J.; Hansen, B.M.; Brok, J.; Esbjørn, B.H.; Greisen, G. Anxiety in Adolescents Born Preterm or with Very Low Birthweight: A Meta-Analysis of Case-Control Studies. Dev. Med. Child Neurol. 2012, 54, 988–994. [Google Scholar] [CrossRef]
- Baracz, S.J.; Everett, N.A.; Robinson, K.J.; Campbell, G.R.; Cornish, J.L. Maternal Separation Changes Maternal Care, Anxiety-like Behaviour and Expression of Paraventricular Oxytocin and Corticotrophin-Releasing Factor Immunoreactivity in Lactating Rats. J. Neuroendocrinol. 2020, 32, e12861. [Google Scholar] [CrossRef]
- Vittner, D.; McGrath, J.; Robinson, J.A.; Lawhon, G.; Cusson, R.; Eisenfeld, L.; Walsh, S.; Young, E.; Cong, X. Increase in Oxytocin From Skin-to-Skin Contact Enhances Development of Parent–Infant Relationship. Biol. Res. Nurs. 2018, 20, 54–62. [Google Scholar] [CrossRef]
- Mah, B.L.; Van Ijzendoorn, M.H.; Out, D.; Smith, R.; Bakermans-Kranenburg, M.J. The Effects of Intranasal Oxytocin Administration on Sensitive Caregiving in Mothers with Postnatal Depression. Child Psychiatry Hum. Dev. 2017, 48, 308–315. [Google Scholar] [CrossRef]
- Ellberg, C.C.; Sayler, K.; Hibel, L.C. Maternal Distress across the Postnatal Period Is Associated with Infant Secretory Immunoglobulin A. Dev. Psychobiol. 2020, 62, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Van den Bergh, B.R.H.; Van Calster, B.; Smits, T.; Van Huffel, S.; Lagae, L. Antenatal Maternal Anxiety Is Related to HPA-Axis Dysregulation and Self-Reported Depressive Symptoms in Adolescence: A Prospective Study on the Fetal Origins of Depressed Mood. Neuropsychopharmacology 2008, 33, 2301. [Google Scholar] [CrossRef]
- Abraham, E.; Zagoory-Sharon, O.; Feldman, R. Early Maternal and Paternal Caregiving Moderates the Links between Preschoolers’ Reactivity and Regulation and Maturation of the HPA-Immune Axis. Dev. Psychobiol. 2021, 63, 1482–1498. [Google Scholar] [CrossRef]
- Marques-Feixa, L.; Castro-Quintas, Á.; Palma-Gudiel, H.; Romero, S.; Morer, A.; Rapado-Castro, M.; Martín, M.; Zorrilla, I.; Blasco-Fontecilla, H.; Ramírez, M.; et al. Secretory Immunoglobulin A (s-IgA) Reactivity to Acute Psychosocial Stress in Children and Adolescents: The Influence of Pubertal Development and History of Maltreatment. Brain. Behav. Immun. 2022, 103, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, C.E. Continuity and Discontinuity of Attachment from Infancy through Adolescence. Child Dev. 2000, 71, 690–694. [Google Scholar] [CrossRef] [PubMed]
- Bornstein, M.H.; Putnick, D.L.; Esposito, G. Continuity and Stability in Development. Child Dev. Perspect. 2017, 11, 113–119. [Google Scholar] [CrossRef]
- Feldman, R. Parent-Infant Synchrony: Biological Foundations and Developmental Outcomes. Curr. Dir. Psychol. Sci. 2007, 48, 329–354. [Google Scholar] [CrossRef]
- Thelen, E.; Smith, L.B. A Dynamic Systems Approach to the Development of Cognition and Action; MIT Press: Cambridge, MA, USA, 1994. [Google Scholar]
- Popper, K. The Logic of Scientific Discovery; Routledge: London, UK, 2005. [Google Scholar]
- Yaniv, A.U.; Salomon, R.; Waidergoren, S.; Shimon-Raz, O.; Djalovski, A.; Feldman, R. Synchronous Caregiving from Birth to Adulthood Tunes Humans’ Social Brain. Proc. Natl. Acad. Sci. USA 2021, 118, e2012900118. [Google Scholar] [CrossRef]
- Beck, A.T.; Steer, R.A.; Brown, G.K. Manual for the Beck Depression Inventory-II; The Psychological Corporation: San Antonio, TX, USA, 1996; pp. 1–82. [Google Scholar]
- Spielberger, C.D.; Gorsuch, R.L.; Lushene, P.R.; Vagg, P.R.; Jacobs, A.G. Manual for the State-Trait Anxiety Inventory; Consulting Psychologists Press: Palo Alto, CA, USA, 1983. [Google Scholar]
- Feldman, R. Coding Interactive Behavior; Bar Ilan University: Ramat-Gan, Israel, 1998; pp. 1–54. [Google Scholar]
- Pratt, M.; Goldstein, A.; Feldman, R. Child Brain Exhibits a Multi-Rhythmic Response to Attachment Cues. Soc. Cogn. Affect. Neurosci. 2018, 13, 957–966. [Google Scholar] [CrossRef]
- Bayley, N. Manual for the Bayley Scales of Infant Development, 2nd ed.; Psychological Corporation: New York, NY, USA, 1993. [Google Scholar]
- Korkman, M.; Kirk, U.; Kemp, S. NEPSY: A Developmental Neuropsychological Assessment; The Psychological Corporation: San Antonio, TX, USA, 1998. [Google Scholar]
- Diamond, A.; Barnett, W.S.; Thomas, J.; Munro, S. The Early Years: Preschool Program Improves Cognitive Control. Science 2007, 318, 1387–1388. [Google Scholar] [CrossRef]
- Pruessner, M.; Hellhammer, D.H.; Pruessner, J.C.; Lupien, S.J. Self-Reported Depressive Symptoms and Stress Levels in Healthy Young Men: Associations With the Cortisol Response to Awakening. Psychosom. Med. 2003, 65, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Beck, A.T.; Ward, C.H.; Mendelson, M.; Mock, J.; Erbaugh, J. An Inventory for Measuring Depression. Arch. Gen. Psychiatry 1961, 4, 561–571. [Google Scholar] [CrossRef] [PubMed]
- JASP Team. JASP (Version 0.11.1) [Computer Software]. 2019. Available online: https://jasp-stats.org/ (accessed on 8 June 2023).
- van Buuren, S.; Groothuis-Oudshoorn, K. Mice: Multivariate Imputation by Chained Equations in R. J. Stat. Softw. 2011, 45, 1–67. [Google Scholar] [CrossRef]
- Schafer, J.L.; Graham, J.W. Missing Data: Our View of the State of the Art. Psychol. Methods 2002, 7, 147–177. [Google Scholar] [CrossRef]
- Rosseel, Y. Lavaan: An R Package for Structural Equation. J. Stat. Softw. 2012, 48, 1–36. [Google Scholar] [CrossRef]
- Hu, L.; Bentler, P.M. Cutoff Criteria for Fit Indexes in Covariance Structure Analysis: Conventional Criteria versus New Alternatives. Struct. Equ. Model. A Multidiscip. J. 1999, 6, 1–55. [Google Scholar] [CrossRef]
- Nyström, K.; Axelsson, K. Mothers’ Experience of Being Separated from Their Newborns. J. Obstet. Gynecol. Neonatal Nurs. 2002, 31, 275–282. [Google Scholar] [CrossRef]
- Shorey, S.; Chee, C.Y.I.; Ng, E.D.; Chan, Y.H.; Tam, W.W.S.; Chong, Y.S. Prevalence and Incidence of Postpartum Depression among Healthy Mothers: A Systematic Review and Meta-Analysis. J. Psychiatr. Res. 2018, 104, 235–248. [Google Scholar] [CrossRef]
- de Paula Eduardo, J.A.F.; de Rezende, M.G.; Menezes, P.R.; Del-Ben, C.M. Preterm Birth as a Risk Factor for Postpartum Depression: A Systematic Review and Meta-Analysis. J. Affect. Disord. 2019, 259, 392–403. [Google Scholar] [CrossRef]
- Bernier, A.; Carlson, S.M.; Whipple, N. From External Regulation to Self-Regulation: Early Parenting Precursors of Young Children’s Executive Functioning. Child Dev. 2010, 81, 326–339. [Google Scholar] [CrossRef]
- Hughes, C.; Devine, R.T. For Better or for Worse? Positive and Negative Parental Influences on Young Children’s Executive Function. Child Dev. 2019, 90, 593–609. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.A.; Goodman, M. Development of Emotion Regulation: More than Meets the Eye. In Emotion Regulation and Psychopathology: A Transdiagnostic Approach to Etiology and Treatment; The Guilford Press: New York, NY, USA, 2010; pp. 38–58. ISBN 1-60623-450-1. [Google Scholar]
- Pearson, R.M.; Bornstein, M.H.; Cordero, M.; Scerif, G.; Mahedy, L.; Evans, J.; Abioye, A.; Stein, A. Maternal Perinatal Mental Health and Offspring Academic Achievement at Age 16: The Mediating Role of Childhood Executive Function. J. Child Psychol. Psychiatry Allied Discip. 2016, 57, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Priel, A.; Zeev-Wolf, M.; Djalovski, A.; Feldman, R. Maternal Depression Impairs Child Emotion Understanding and Executive Functions: The Role of Dysregulated Maternal Care across the First Decade of Life. Emotion 2020, 20, 1042–1058. [Google Scholar] [CrossRef]
- Dias, N.M.; Trevisan, B.T.; León, C.B.R.; Prust, A.P.; Seabra, A.G. Can Executive Functions Predict Behavior in Preschool Children? Psychol. Neurosci. 2017, 10, 383–393. [Google Scholar] [CrossRef]
- Böhm, B.; Smedler, A.C.; Forssberg, H. Impulse Control, Working Memory and Other Executive Functions in Preterm Children When Starting School. Acta Paediatr. Int. J. Paediatr. 2004, 93, 1363–1371. [Google Scholar] [CrossRef]
- Trossman, R.; Spence, S.L.; Mielke, J.G.; McAuley, T. How Do Adverse Childhood Experiences Impact Health? Exploring the Mediating Role of Executive Functions. Psychol. Trauma Theory Res. Pract. Policy 2021, 13, 206–213. [Google Scholar] [CrossRef]
- Feldman, R. Social Behavior as a Transdiagnostic Marker of Resilience. Annu. Rev. Clin. Psychol. 2021, 17, 153–180. [Google Scholar] [CrossRef]
- Cong, X.; Ludington-Hoe, S.M.; Hussain, N.; Cusson, R.M.; Walsh, S.; Vazquez, V.; Briere, C.E.; Vittner, D. Parental Oxytocin Responses during Skin-to-Skin Contact in Pre-Term Infants. Early Hum. Dev. 2015, 91, 401–406. [Google Scholar] [CrossRef]
- Endevelt-Shapira, Y.; Djalovski, A.; Dumas, G.; Feldman, R. Maternal Chemosignals Enhance Infant-Adult Brain-to-Brain Synchrony. Sci. Adv. 2021, 7, 6867. [Google Scholar] [CrossRef]
- Korja, R.; Latva, R.; Lehtonen, L. The Effects of Preterm Birth on Mother-Infant Interaction and Attachment during the Infant’s First Two Years. Acta Obstet. Gynecol. Scand. 2012, 91, 164–173. [Google Scholar] [CrossRef]
- Ladd, C.O.; Owens, M.J.; Nemeroff, C.B. Persistent Changes in Corticotropin-Releasing Factor Neuronal Systems Induced by Maternal Deprivation. Endocrinology 1996, 137, 1212–1218. [Google Scholar] [CrossRef] [PubMed]
- Thibeau, S.; D’Apolito, K. Review of the Relationships Between Maternal Characteristics and Preterm Breastmilk Immune Components. Biol. Res. Nurs. 2012, 14, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Hart, S.; Boylan, L.M.; Border, B.; Carroll, S.R.; McGunegle, D.; Lampe, R.M. Breast Milk Levels of Cortisol and Secretory Immunoglobulin A (SIgA) Differ with Maternal Mood and Infant Neuro-Behavioral Functioning. Infant Behav. Dev. 2004, 27, 101–106. [Google Scholar] [CrossRef]
- Groer, M.; Davis, M.; Steele, K. Associations between Human Milk SIgA and Maternal Immune, Infectious, Endocrine, and Stress Variables. J. Hum. Lact. 2004, 20, 153–158. [Google Scholar] [CrossRef]
- Ma, D.; Serbin, L.A.; Stack, D.M. Children’s Anxiety Symptoms and Salivary Immunoglobulin A: A Mutual Regulatory System? Dev. Psychobiol. 2018, 60, 202–215. [Google Scholar] [CrossRef]
- Yirmiya, K.; Djalovski, A.; Motsan, S.; Zagoory-Sharon, O.; Feldman, R. Stress and Immune Biomarkers Interact with Parenting Behavior to Shape Anxiety Symptoms in Trauma-Exposed Youth. Psychoneuroendocrinology 2018, 98, 153–160. [Google Scholar] [CrossRef]
- Ulmer-Yaniv, A.; Djalovski, A.; Yirmiya, K.; Halevi, G.; Zagoory-Sharon, O.; Feldman, R.; Ulmer-Yaniv, A.; Feldman, R.; Zagoory-Sharon, O.; Djalovski, A.; et al. Maternal Immune and Affiliative Biomarkers and Sensitive Parenting Mediate the Effects of Chronic Early Trauma on Child Anxiety. Psychol. Med. 2017, 48, 1020–1033. [Google Scholar] [CrossRef]
- Krahel, A.; Paszynska, E.; Slopien, A.; Gawriolek, M.; Otulakowska-Skrzynska, J.; Rzatowski, S.; Hernik, A.; Hanć, T.; Bryl, E.; Szczesniewska, P.; et al. Stress/Immune Biomarkers in Saliva among Children with Adhd Status. Int. J. Environ. Res. Public Health 2021, 18, 769. [Google Scholar] [CrossRef]
- Tang, Y.Y.; Ma, Y.; Wang, J.; Fan, Y.; Feng, S.; Lu, Q.; Yu, Q.; Sui, D.; Rothbart, M.K.; Fan, M.; et al. Short-Term Meditation Training Improves Attention and Self-Regulation. Proc. Natl. Acad. Sci. USA 2007, 104, 17152–17156. [Google Scholar] [CrossRef]
- Feldman, R. Mother-Infant Synchrony and the Development of Moral Orientation in Childhood and Adolescence: Direct and Indirect Mechanisms of Developmental Continuity. Am. J. Orthopsychiatry 2007, 77, 582–597. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| KC | Control | t/χ2 | Cohen’s d | |
|---|---|---|---|---|
| Gender (% Female) | 19 (47.5%) | 16 (40%) | χ2(1) = 0.457, p = 0.499 | |
| Child age | 18.49 (SD = 0.10) | 18.45 (SD = 0.84) | t(78) = 0.218, p = 0.82 | 0.049 |
| Mother age | 48.2 (SD = 4.46) | 48.5 (SD = 5.12) | t(67) = −0.428, p = 0.67 | 0.103 |
| Mother education (above average) | 22 (62.9%) | 26 (70.3%) | χ2(4) = 0.178, p = 0.776 | |
| Household income (above average) | 12 (30.76%) | 12 (33.33%) | χ2(2) = 4.135, p = 0.13 | |
| Birth week (weeks) | 30.769 (SD = 2.76) | 31.475 (SD = 2.43) | t(78) = −1.254, p = 0.21 | −0.280 |
| Birth weight (gram) | 1296 (SD = 278) | 1460 (SD = 356) | t(78) = −2.301, p = 0.024 | −0.514 |
| KC Mean (SD) | Control Mean (SD) | t-Test | Cohen’s d | |
|---|---|---|---|---|
| Child depression/anxiety | −0.04 (0.86) | −0.01 (0.73) | t(78) = −0.155, p = 0.87 | −0.009 |
| Child OT | 180.13 (31.2) | 183.52 (36.9) | t(78) = −0.442, p = 0.66 | −0.099 |
| Child s-IgA | 634.79 (72.4) | 661.91 (79.0) | t(78) = −1.599, p = 0.11 | −0.358 |
| Child Focused attention | 4.19 (0.39) | 3.82 (0.64) | t(78) = 3.157, p = 0.002 | 0.706 |
| Child EF score | 111.87 (11.06) | 101.85 (11.9) | t(78) = 3.902, p < 0.001 | 0.873 |
| Mother depression/anxiety | −0.27 (0.51) | 0.30 (1.08) | t(78) = −1.43, p = 0.157 | −0.320 |
| Mother depression/anxiety (infancy) | −0.19 (0.71) | 0.08 (0.71) | t(78) = −3.580, p < 0.001 | −0.800 |
| Dyadic synchrony infancy | 2.66 (0.87) | 2.23 (0.95) | t(78) = 2.108, p = 0.038 | 0.471 |
| Dyadic synchrony childhood | 3.49 (0.71) | 3.13 (0.82) | t(78) = 2.073, p = 0.041 | 0.464 |
| Dyadic synchrony adulthood | 3.95 (0.74) | 3.51 (0.80) | t(78) = 2.537, p = 0.013 | 0.567 |
| Dyadic Synchrony | Child Regulatory Capacities | Maternal Anxiety/Depression | Child—Adulthood Outcomes | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Infancy | Childhood | Adulthood | Focused Attention | EF Score | Infancy | Adulthood | Emotional Distress | s-IgA | OT | |
| Dyadic synchrony (Infancy) | — | |||||||||
| Dyadic synchrony (Childhood) | 0.506 *** | — | ||||||||
| Dyadic synchrony (Adulthood) | 0.490 *** | 0.762 *** | — | |||||||
| Focused attention | 0.215 | 0.300 ** | 0.431 *** | |||||||
| Child EF score | 0.341 ** | 0.225 * | 0.161 | |||||||
| Maternal depression/anxiety (Infancy) | −0.298 ** | −0.264 * | −0.261 * | −0.172 | −0.386 *** | — | ||||
| Maternal depression/anxiety (Adulthood) | −0.095 | −0.172 | −0.073 | −0.074 | −0.387 *** | 0.451 *** | — | |||
| Child depression/anxiety | −0.182 | −0.260 * | −0.248 * | −0.169 | −0.231 ** | 0.420 *** | 0.376 *** | — | ||
| Child s-IgA | −0.115 | 0.143 | 0.004 | −0.259 * | −0.084 | 0.126 | −0.059 | −0.016 | — | |
| Child OT | 0.321 ** | 0.179 | 0.299 ** | 0.289 ** | −0.051 | 0.052 | −0.120 | −0.019 | 0.03 | — |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ulmer-Yaniv, A.; Yirmiya, K.; Peleg, I.; Zagoory-Sharon, O.; Feldman, R. Developmental Cascades Link Maternal–Newborn Skin-to-Skin Contact with Young Adults’ Psychological Symptoms, Oxytocin, and Immunity; Charting Mechanisms of Developmental Continuity from Birth to Adulthood. Biology 2023, 12, 847. https://doi.org/10.3390/biology12060847
Ulmer-Yaniv A, Yirmiya K, Peleg I, Zagoory-Sharon O, Feldman R. Developmental Cascades Link Maternal–Newborn Skin-to-Skin Contact with Young Adults’ Psychological Symptoms, Oxytocin, and Immunity; Charting Mechanisms of Developmental Continuity from Birth to Adulthood. Biology. 2023; 12(6):847. https://doi.org/10.3390/biology12060847
Chicago/Turabian StyleUlmer-Yaniv, Adi, Karen Yirmiya, Itai Peleg, Orna Zagoory-Sharon, and Ruth Feldman. 2023. "Developmental Cascades Link Maternal–Newborn Skin-to-Skin Contact with Young Adults’ Psychological Symptoms, Oxytocin, and Immunity; Charting Mechanisms of Developmental Continuity from Birth to Adulthood" Biology 12, no. 6: 847. https://doi.org/10.3390/biology12060847
APA StyleUlmer-Yaniv, A., Yirmiya, K., Peleg, I., Zagoory-Sharon, O., & Feldman, R. (2023). Developmental Cascades Link Maternal–Newborn Skin-to-Skin Contact with Young Adults’ Psychological Symptoms, Oxytocin, and Immunity; Charting Mechanisms of Developmental Continuity from Birth to Adulthood. Biology, 12(6), 847. https://doi.org/10.3390/biology12060847