
Competitive Advantage of Broussonetia papyrifera Growing in a Native Area as Suggested by Structural Diversity
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Sampling
2.3. Structural Indices
2.4. Data Analyses
3. Results
3.1. Number, Density, and Sex Ratio of Paper Mulberry
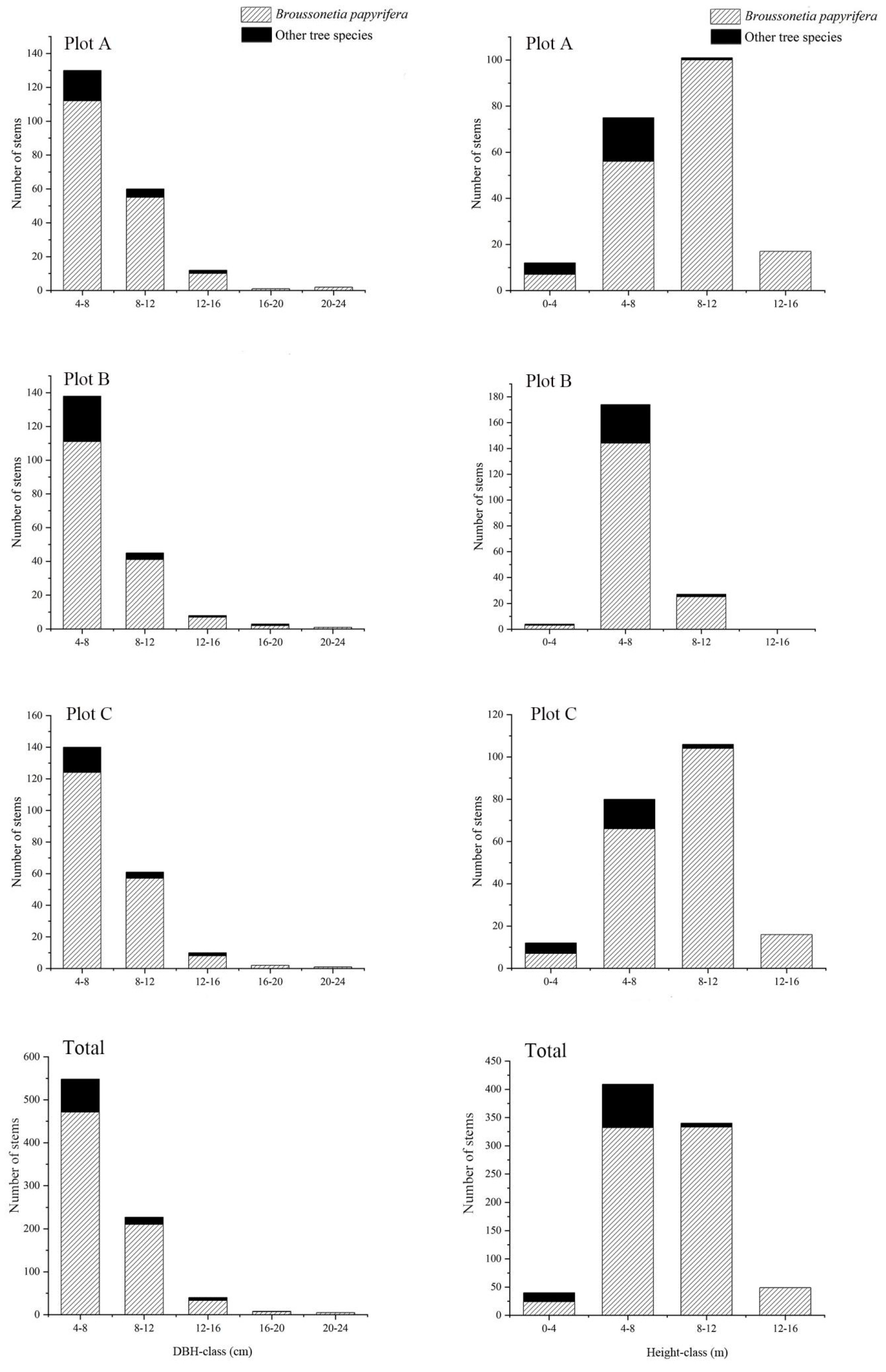
3.2. Diameter and Height Distributions of Paper Mulberry and Other Tree Species
3.3. Structural Indices of the Paper Mulberry Communities
3.3.1. The Mingling Index and the Tree–Tree Interval Index
3.3.2. The Diameter Differentiation Index and the Height Differentiation Index
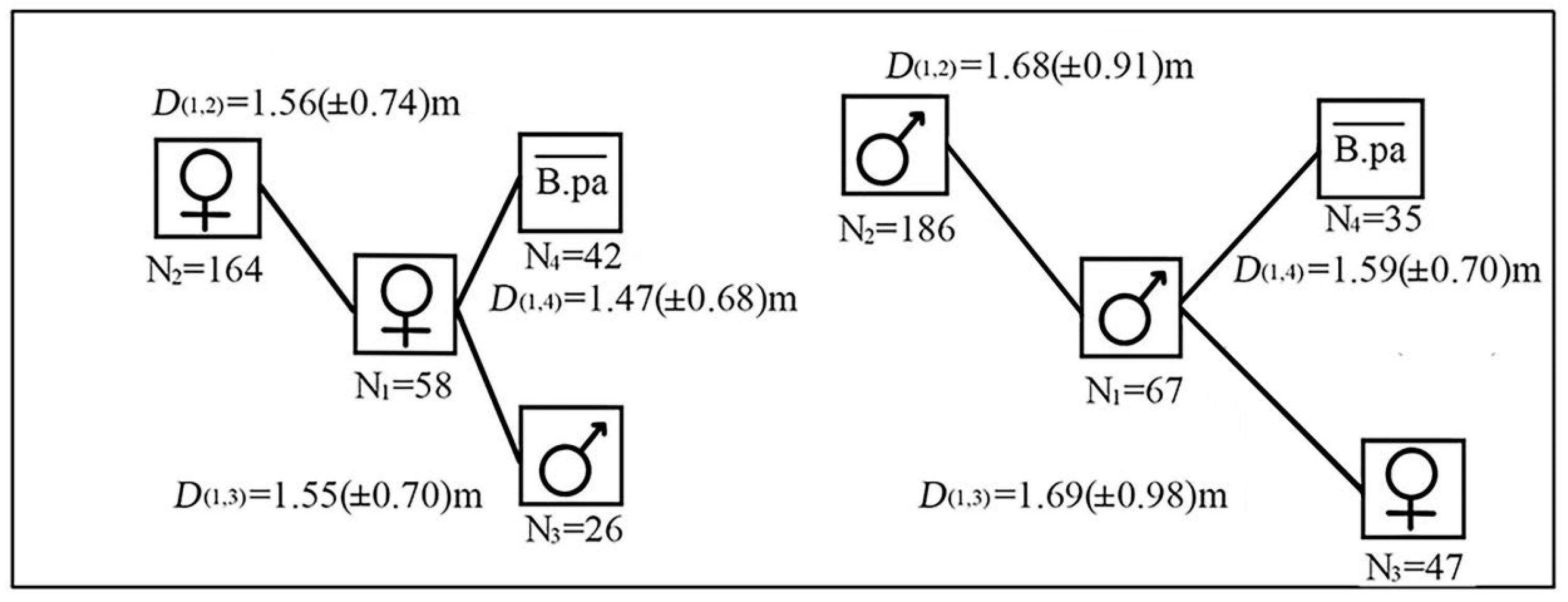
3.4. Spatial Distributions of Male and Female Paper Mulberry
4. Discussion
4.1. Competition and Structural Indices of Paper Mulberry
4.2. Spatial Configuration of Female and Male Paper Mulberry
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fu, L.G.; Xin, Y.Q.; Whittemore, A. Moraceae. In Flora of China; Wu, Z.Y., Raven, P.H., Eds.; Science Press: Beijing, China, 2003; Volume 5, pp. 1–19. [Google Scholar]
- Chen, Z.H.; Sun, M.J. Color Illustrations of Common Xylophyta in Zhejiang Province; Zhejiang University Press: Hangzhou, China, 2015. [Google Scholar]
- Li, M.; Li, Y.; Li, H.Q.; Wu, G.J. Overexpression of AtNHX5 improves tolerance to both salt and drought stress in Broussonetia papyrifera (L.) Vent. Tree Physiol. 2011, 31, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Seelenfreund, D.; Clarke, A.C.; Oyanedel, N.; Piña, R.; Lobos, S.; Smith, M.; A-Seelenfreund, E.A. Paper mulberry (Broussonetia papyrifera) as a commensal model for human mobility in Oceania: Anthropological, botanical and genetic considerations. N. Z. J. Bot. 2011, 48, 231–247. [Google Scholar] [CrossRef]
- Matthews, P.J. Ethnobotany and the origins of Broussonetia papyrifera in Polynesia: An essay on tapa prehistory. In Oceanic Culture History: Essays in Honour of Roger Green; Davidson, J., Irwin, G., Leach, F., Pawley, A., Brown, D., Eds.; Archaeol. N. Z.: Dunedin, New Zealand, 1996; pp. 117–132. [Google Scholar]
- Wang, M.Q.; Guan, Q.W. Prediction of potential suitable areas for Broussonetia papyrifera in China using the MaxEnt model and CIMP6 data. J. Plant. Ecol. 2023, 16, 4. [Google Scholar] [CrossRef]
- Huang, H.M.; Zhao, Y.L.; Xu, Z.G.; Zhang, W.; Jiang, K.K. Physiological responses of Broussonetia papyrifera to manganese stress, a candidate plant for phytoremediation. Ecotoxicol. Environ. Saf. 2019, 181, 18–25. [Google Scholar] [CrossRef]
- Payacan, C.; Moncada, X.; Rojas, G.; Clarke, A.; Chung, K.F.; Allaby, R.; Seelenfreund, D.; Seelenfreund, A. Phylogeography of herbarium specimens of asexually propagated paper mulberry [Broussonetia papyrifera (L.) L’Hér. ex Vent. (Moraceae)] reveals genetic diversity across the Pacific. Ann. Bot. 2017, 120, 387–404. [Google Scholar] [CrossRef] [PubMed]
- Maan, I.; Kaur, A.; Sharma, A.; Singh, H.P.; Batish, D.R.; Kohli, R.K.; Arora, N.K. Variations in leaf litter decomposition explain invasion success of Broussonetia papyrifera over confamilial non-invasive Morus alba in urban habitats. Urban For. Urban Green. 2022, 67, 127408. [Google Scholar] [CrossRef]
- Malik, R.N.; Husain, S.Z. Broussonetia papyrifera (L.) L’hér. ex Vent.: An environmental constraint on the Himalayan foothills vegetation. Pak. J. Bot. 2007, 39, 1045–1053. [Google Scholar]
- Florece, L.M.; Coladilla, J.O. Spatial distribution and dominance of paper mulberry (Broussonetia papyrifera) in the vicinities of Mt. Makiling, Philippines. Environ. Sci. Manag. 2006, 9, 54–65. [Google Scholar]
- Ghersa, C.M.; de la Fuente, E.; Suarez, S.; Leon, R.J.C. Woody species in the Rolling Pampa grasslands, Argentina. Agr. Ecosyst. Environ. 2002, 88, 271–278. [Google Scholar] [CrossRef]
- Morgan, E.C.; Overholt, W.A.; Sellers, B. Wildland Weeds: Paper Mulberry, Broussonetia papyrifera; Publication #ENY-702; Series of the Entomology and Nematology Department, Florida Co-operative Extension Service, Institute of Food and Agricultural Sciences, University of Florida: Gainesville, FL, USA, 2019; Available online: https://edis.ifas.ufl.edu/in498 (accessed on 22 August 2023).
- Bosu, P.P.; Apetorgbor, M.M.; Nkrumah, E.E.; Bandoh, K.P. The impact of Broussonetia papyrifera (L.) Vent. on community characteristics in the forest and forest–savannah transition ecosystems of Ghana. Afr. J. Ecol. 2013, 51, 528–535. [Google Scholar] [CrossRef]
- Khatoon, S.; Ali, S.I. Alien Invasive Species in Pakistan; University of Karachi, Pers. Comm.: Karachi, Pakistan, 1999. [Google Scholar]
- Kyereh, B.; Agyeman, V.K.; Abebrese, I.K. Ecological characteristics that enhance Broussonetia papyrifera’s invasion in a semideciduous forest in Ghana. J. Ecosyst. 2014, 2014, 270196. [Google Scholar] [CrossRef]
- Liao, S.X.; Deng, Z.H.; Cui, K.; Cui, Y.Z.; Zhang, C.H. Genetic diversity of Broussonetia papyrifera populations in southwest China. Genet. Mol. Res. 2014, 13, 7553–7563. [Google Scholar] [CrossRef] [PubMed]
- Peñailillo, J.; Olivares, G.; Moncada, X.; Payacán, C.; Chang, C.S.; Chung, K.F.; Matthews, P.J.; Seelenfreund, A.; Seelenfreund, D. Sex distribution of paper Mulberry (Broussonetia papyrifera) in the Pacific. PLoS ONE 2016, 11, e0161148. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.S.; Liu, H.L.; Moncada, X.; Seelenfreund, A.; Seelenfreund, D.; Chung, K.F. A holistic picture of Austronesian migrations revealed by phylogeography of Pacific paper mulberry. Proc. Natl. Acad. Sci. USA 2015, 112, 13537–13542. [Google Scholar] [CrossRef] [PubMed]
- Zenni, R.D.; Dickie, I.A.; Wingfield, M.J.; Hirsch, H.; Crous, C.J.; Meyerson, L.A.; Burgess, T.I.; Zimmermann, T.J.; Klock, M.M.; Siemann, E.; et al. Evolutionary dynamics of tree invasions: Complementing the unified framework for biological invasions. AoB Plants 2017, 9, plw085. [Google Scholar] [CrossRef]
- Ssemanda, D.M.; Ssekuubwa, E. Broussonetia papyrifera invasion reduces the abundance and basal area of selected native timber species in a tropical moist forest in Uganda. Biol. Invasions 2023, 25, 165–178. [Google Scholar] [CrossRef]
- Hierro, J.L.; Callaway, R.M. Allelopathy and exotic plant invasion. Plant Soil. 2003, 256, 29–39. [Google Scholar] [CrossRef]
- Levine, J.M.; Vilà, M.; Antonio, C.M.D.; Dukes, J.S.; Grigulis, K.; Lavorel, S. Mechanisms underlying the impacts of exotic plant invasions. Proc. R. Soc. B-Biol. Sci. 2003, 270, 775–781. [Google Scholar] [CrossRef]
- Guo, Q. Intercontinental biotic invasions: What can we learn from native populations and habitats? Biol. Invasions 2006, 8, 1451–1459. [Google Scholar] [CrossRef][Green Version]
- Bristow, M.; Vanclay, J.K.; Brooks, L.; Hunt, M. Growth and species interactions of Eucalyptus pellita in a mixed and monoculture plantation in the humid tropics of north Queensland. For. Ecol. Manag. 2006, 233, 285–294. [Google Scholar] [CrossRef]
- Jiang, J.; Sun, J.; Zhang, G.; Wang, J. Intraspecific and interspecific competition in the endemic and endangered Yulania zenii. Chin. J. Ecol. 2010, 29, 201–206. [Google Scholar]
- Zhang, G.F.; Yao, R.; Jiang, R.X.; Chen, F.C.; Zhang, W.Y. Intraspecific and interspecific competitive intensity of Shaniodendron subaequale in different habitats of Wanfoshan, Anhui Province, China. Chin. J. Ecol. 2016, 35, 1744–1750. [Google Scholar]
- Song, Y.F.; Yan, G.; Zhang, G.F. Light competition contributes to the death of Masson pines of coniferous and broad-leaf mixed forests in subtropical China. Forests 2022, 13, 85. [Google Scholar] [CrossRef]
- Zhang, G.F.; Li, Q.; Hou, X.R. Structural diversity of naturally regenerating Chinese yew (Taxus wallichiana var. mairei) populations in ex situ conservation. Nord. J. Bot. 2018, 36, 1–10. [Google Scholar]
- Gadow, K.V.; Zhang, C.Y.; Wehenkel, C.; Pommerening, A.; Corral-Rivas, J.; Korol, M.; Myklush, S.; Hui, G.Y.; Kiviste, A.; Zhao, X.H. Forest structure and diversity. In Continuous Cover Forestry. Managing Forest Ecosystems; Pukkala, T., Gadow, K.v., Eds.; Springer: Dordrecht, The Netherlands, 2012; Volume 23, pp. 29–83. [Google Scholar]
- Pommerening, A. Approaches to quantifying forest structures. Forestry 2002, 75, 305–324. [Google Scholar] [CrossRef]
- Hui, G.Y.; Gadow, K.V. Quantitative Analysis of Forest Spatial Structure; Science and Technology Press: Beijing, China, 2003. [Google Scholar]
- Wang, H.X.; Zhang, G.Q.; Hui, G.Y.; Li, Y.F.; Hu, Y.B.; Zhao, Z.H. The influence of sampling unit size and spatial arrangement patterns on neighborhood-based spatial structure analyses of forest stands. For. Ecosyst. 2016, 25, e056. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, G.F.; Li, X. Structural diversity and conservation implications of Parrotia subaequalis (Hamamelidaceae), a rare and endangered tree species in China. Nat. Conserv. 2021, 44, 99–115. [Google Scholar] [CrossRef]
- González-Lorca, J.; Rivera-Hutinel, A.; Moncada, X.; Lobos, S.; Seelenfreund, D.; Seelenfreund, A.N. Ancient and modern introduction of Broussonetia papyrifera ([L.] Vent.; Moraceae) into the Pacific: Genetic, geographical and historical evidence. N. Z. J. Bot. 2015, 53, 75–89. [Google Scholar] [CrossRef]
- Li, Y.M.; Ye, Q. Climate classification over China based on Köppen climate classification in the context of ENSO. Clim. Chang. Res. 2019, 15, 352–362. [Google Scholar]
- Xie, C.P. Analysis of the composition and structure of peri-urban coniferous tree communities. J. Sichuan Agric. Univ. 2015, 33, 358–363. [Google Scholar]
- Li, K.D.; Zhang, G.F. Species diversity and distribution pattern of heritage trees in the rapidly-urbanizing province of Jiangsu, China. Forests 2021, 12, 1543. [Google Scholar] [CrossRef]
- Wang, R.X. Leaf Population Dynamic of Main Plant Species from Deciduous Broad-Leaved Forest in Xianlin, Nanjing. 2015. Available online: https://kns.cnki.net/kcms2/article/abstract?v=-YY6Aedvp4bLysqFVxOu_NlcZWEr520bNsY5HsFojmkGQuXFjPLxHB3680iyeZ3z89bS7GgfaTFgwsUyBYor8NStQpQQvjjTQUPDY6J8ghcdRC4HrIlKYt6oaCqtSlTLJ0BNADXWxrWdhfFee5yhVA==&uniplatform=NZKPT&language=CHS (accessed on 20 August 2023).
- Li, G.; Rogers, P.C.; Huang, J. Black locust (Robinia pseudoacacia L.) range shifts in China: Application of a global model in climate change futures. Clim. Chang. Ecol. 2021, 2, 100036. [Google Scholar] [CrossRef]
- Wang, R.J.; Wang, G.F.; Zeng, X.F. Journal of Invasive Alien Plants of China; Shanghai Jiao Tong University Press: Shanghai, China, 2020; Volume 2, pp. 131–132. [Google Scholar]
- Ruprecht, H.; Dhar, A.; Aigner, B.; Oitzinger, G.; Klumpp, R.; Vacik, H. Structural diversity of English yew (Taxus baccata L.) populations. Eur. J. For. Res. 2010, 129, 189–198. [Google Scholar] [CrossRef]
- Pommerening, A. Evaluating structural indices by reversing forest structural analysis. For. Ecol. Manag. 2006, 224, 266–277. [Google Scholar] [CrossRef]
- Sefidi, K.; Copenheaver, C.A.; Kakavand, M.; Behjou, F.K. Structural diversity within mature forests in northern Iran: A case study from a relic population of Persian ironwood (Parrotia persica C.A. Meyer). For. Sci. 2015, 61, 258–265. [Google Scholar] [CrossRef]
- Wu, W. SPSS and Statistical Thinking; Tsinghua University Press: Beijing, China, 2019. [Google Scholar]
- Kumar, M.; Garkoti, S.C. Functional traits, growth patterns, and litter dynamics of invasive alien and co-occurring native shrub species of chir pine forest in the central Himalaya, India. Plant Ecol. 2021, 222, 723–735. [Google Scholar] [CrossRef]
- Li, K.D.; Zhang, G.F.; Zhang, Y.; Griffith, M.P. A noteworthy case of rewilding Chinese yew from a garden population in eastern China. PeerJ 2021, 9, e12341. [Google Scholar] [CrossRef]
- Qureshi, H.; Anwar, T.; Fatima, S.; Akhtar, S.; Khan, S.; Waseem, M.; Mohibullah, M.; Shirani, M.; Riaz, S.; Azeem, M. Invasion Impact Analysis of Broussonetia papyrifera in Pakistan. Pol. J. Environ. Stud. 2020, 29, 2825–2831. [Google Scholar] [CrossRef]
- Wu, Y.Y.; Liu, C.Q.; Li, P.P.; Wang, J.Z.; Xing, D.; Wang, B.L. Photosynthetic characteristics involved in adaptability to Karst soil and alien invasion of paper mulberry (Broussonetia papyrifera (L.) Vent.) in comparison with mulberry (Morus alba L.). Photosynthetica 2009, 47, 155–160. [Google Scholar] [CrossRef]
- Liu, M.; Wang, J.S. Effect of soil nitrogen addition on the photosynthesis of hybrid Broussonetia papyrifera in the subtropical red soil zone. J. Univ. Chin. Acad. Sci. 2022, 39, 732–741. [Google Scholar]
- Li, L.; Xia, Y.F.; Wang, H. Studies on the growth characteristics of Broussonetia papyrifera. Guizhou Sci. 2010, 28, 62–66. [Google Scholar]
- Sorte, C.J.B.; Williams, S.L.; Carlton, J.T. Marine range shifts and species introductions: Comparative spread rates and community impacts. Glob. Ecol. Biogeogr. 2010, 19, 303–316. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P.; Rejmanek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and invasion of alien plants: Concepts and definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Zhou, J.K.; Liu, Y.; Wu, L.; Zhao, Y.L.; Zhang, W.; Yang, G.Y.; Xu, Z.G. Effects of plant growth regulators on the rapid propagation system of Broussonetia papyrifera L. Vent explants. Forests 2021, 12, 874. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Structural Indices | Measures | Forest Structural Characteristics | Implication for Competition |
|---|---|---|---|
| Mixing index (Mi) | Species interspersion | A value of 0 indicates a pure forest stand. A value near 1 indicates that species are highly interspersed. | For a reference tree, a smaller value indicates stronger competitiveness, and vice versa. |
| Tree–tree interval index (Di) | Stand density | Higher values indicate a lower stand density. | For a reference tree, a smaller value indicates stronger competitiveness, and vice versa. |
| Diameter differentiation index (TDi) | Tree diameter variation | A value of 0 indicates uniformity in tree diameters. A value near 1 indicates a high variation in tree diameters. | Positive value indicates a reference tree has advantage over its neighbors in competition horizontally. Higher value indicates greater competitiveness, and vice versa. |
| Height differentiation index (HDi) | Tree height variation | A value of 0 indicates uniformity in tree height. A value near 1 indicates high variation in tree height. | Positive value indicates a reference tree has advantage over its neighbors in competition vertically. Higher value indicates greater competitiveness, and vice versa. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Zhang, G. Competitive Advantage of Broussonetia papyrifera Growing in a Native Area as Suggested by Structural Diversity. Biology 2023, 12, 1410. https://doi.org/10.3390/biology12111410
Zhou Y, Zhang G. Competitive Advantage of Broussonetia papyrifera Growing in a Native Area as Suggested by Structural Diversity. Biology. 2023; 12(11):1410. https://doi.org/10.3390/biology12111410
Chicago/Turabian StyleZhou, Yanrong, and Guangfu Zhang. 2023. "Competitive Advantage of Broussonetia papyrifera Growing in a Native Area as Suggested by Structural Diversity" Biology 12, no. 11: 1410. https://doi.org/10.3390/biology12111410
APA StyleZhou, Y., & Zhang, G. (2023). Competitive Advantage of Broussonetia papyrifera Growing in a Native Area as Suggested by Structural Diversity. Biology, 12(11), 1410. https://doi.org/10.3390/biology12111410