
Expression of the ripAA Gene in the Soilborne Pseudomonas mosselii Can Promote the Control Efficacy against Tobacco Bacterial Wilt

Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial and Plant Materials
2.2. Integration of ripAA into the P. mosselii Chromosome
2.3. Bacterial Confrontation Assay
2.4. β-Galactosidase Activity Driven by the ripAA Promoter in P. mosselii A1
2.5. Semi-Quantitative RT-PCR
2.6. Controlling Tobacco Bacterial Wilt Using P. mosselii AA1
2.7. High-Throughput RNA Sequencing
2.8. Quantitative RT-PCR
2.9. Accession Numbers
3. Results
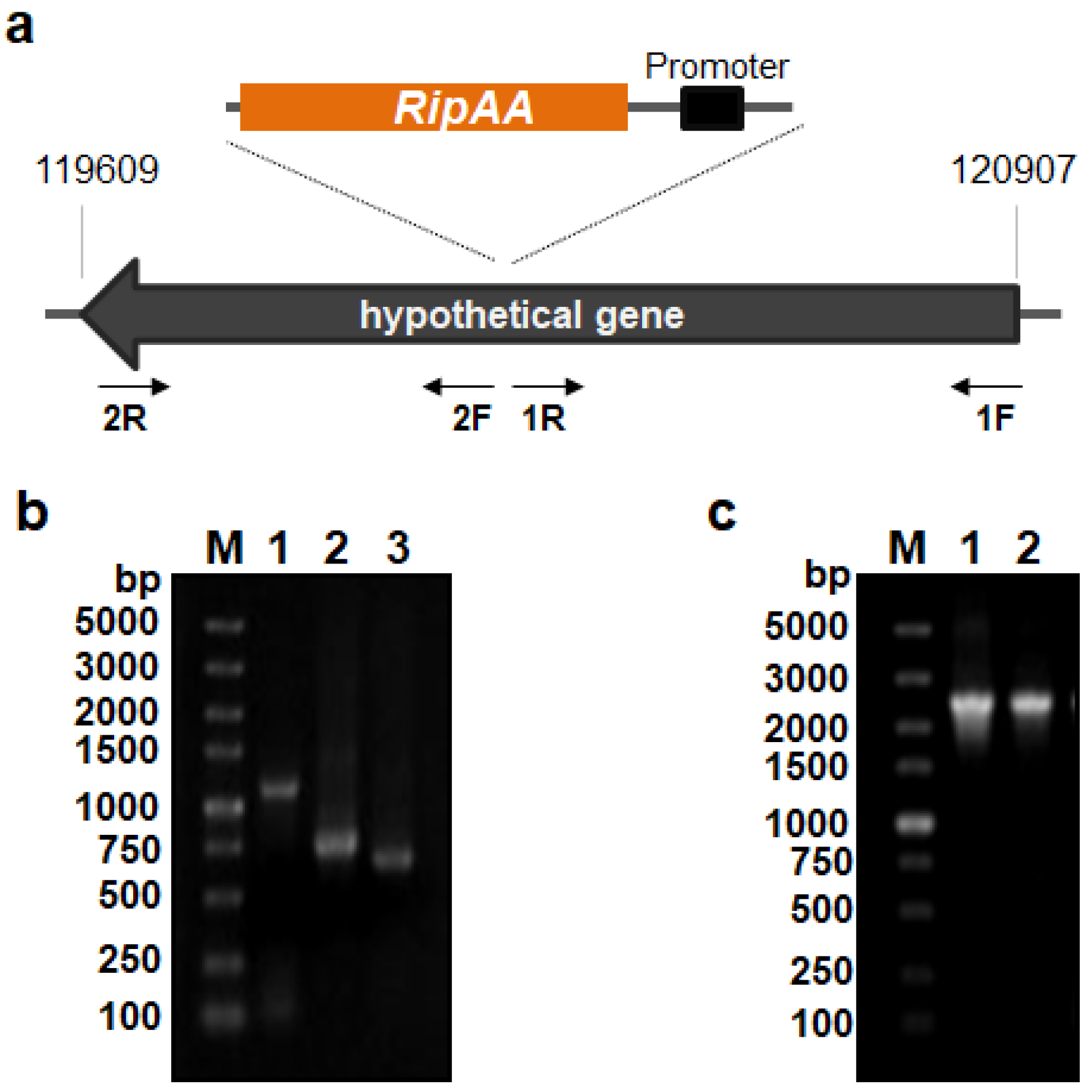
3.1. Construction of an Engineered P. mosselii A1 Carrying the ripAA Gene
3.2. The ripAA Gene Is Transcribed in P. mosselii AA1
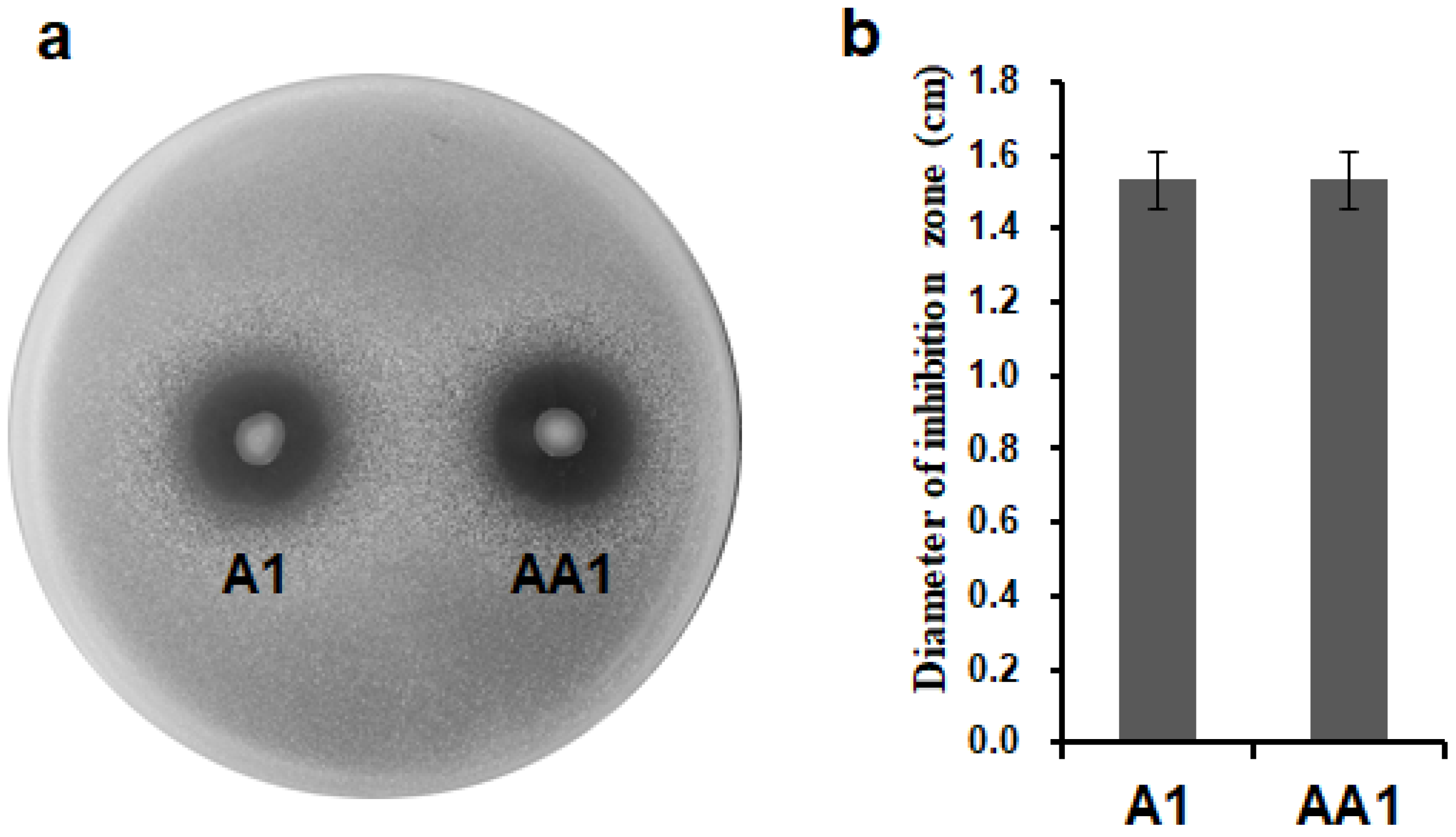
3.3. P. mosselii AA1 Retains Antimicrobial Activity on R. solanacearum
3.4. P. mosselii AA1 Increases Control Efficacy on Tobacco Bacterial Wilt
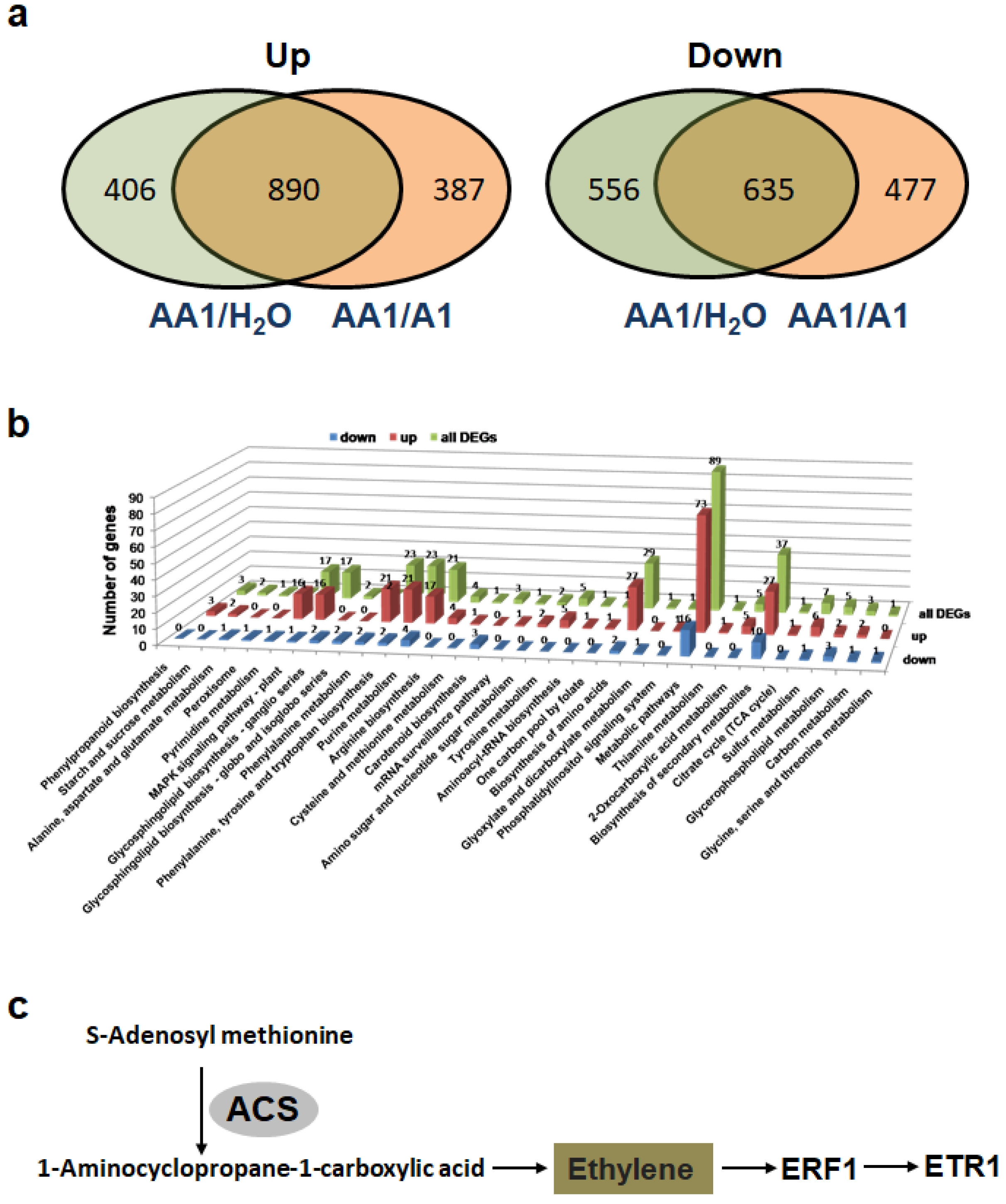
3.5. Transcriptome Analysis of Tobacco Plants Treated with P. mosselii AA1
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Flor, H.H. Current status of the gene-for-gene concept. Annu. Rev. Phytopathol. 1971, 9, 275–296. [Google Scholar] [CrossRef]
- Gassmann, W.; Bhattacharjee, S. Effector-triggered immunity signaling: From gene-for-gene pathways to protein-protein interaction networks. Mol. Plant Microbe Interact. 2012, 25, 862–868. [Google Scholar] [CrossRef] [PubMed]
- Heath, M.C. Hypersensitive response-related death. Plant Mol. Biol. 2000, 44, 321–334. [Google Scholar] [CrossRef]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Bradley, D.J.; Kjellbom, P.; Lamb, C.J. Elicitor- and wound-induced oxidative cross-linking of a proline-rich plant cell wall protein: A novel, rapid defense response. Cell 1992, 70, 21–30. [Google Scholar] [CrossRef]
- Shirasu, K.; Schulze-Lefert, P. Regulators of cell death in disease resistance. Plant Mol. Biol. 2000, 44, 371–385. [Google Scholar] [CrossRef]
- Tripathi, J.N.; Lorenzen, J.; Bahar, O.; Ronald, P.; Tripathi, L. Transgenic expression of the rice Xa21 pattern-recognition receptor in banana (Musa sp.) confers resistance to Xanthomonas campestris pv. musacearum. Plant Biotechnol. J. 2014, 12, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Hayward, A.C. Biology and epidemiology of bacterial wilt caused by Pseudomonas solanacearum. Annu. Rev. Phytopathol. 1991, 29, 65–87. [Google Scholar] [CrossRef]
- Denny, T. Plant pathogenic Ralstonia species. In Plant-Associated Bacterial; Gnanamanickam, S.S., Ed.; Spinger: Dordrecht, The Netherlands, 2006; Volume 16, pp. 573–644. [Google Scholar]
- Chen, R.; Zhu, X.; Wang, Z.; Guo, Z.; Dong, H.; Wang, L.; Liu, Y.; Shi, J. A report of investigating and studying tobacco infectious diseases of 16 main tobacco producing provinces (regions) in China. Chin. Tob. Sci. 1997, 4, 3–9. [Google Scholar]
- Poueymiro, M.; Genin, S. Secreted proteins from Ralstonia solanacearum: A hundred tricks to kill a plant. Curr. Opin. Microbiol. 2009, 12, 44–52. [Google Scholar] [CrossRef]
- Landry, D.; González-Fuente, M.; Deslandes, L.; Peeters, N. The large, diverse, and robust arsenal of Ralstonia solanacearum type III effectors and their in planta functions. Mol. Plant Pathol. 2020, 21, 1377–1388. [Google Scholar] [CrossRef]
- Coll, N.S.; Valls, M. Current knowledge on the Ralstonia solanacearum type III secretion system. Microb. Biotechnol. 2013, 6, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Wang, D.; Yan, J.; Zhou, J.; Deng, Y.; Jiang, Z.; Cao, B.; He, Z.; Zhang, L. Genomic analysis of phylotype I strain EP1 reveals substantial divergence from other strains in the Ralstonia solanacearum species complex. Front. Microbiol. 2016, 7, 1719. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tang, Y.; Qin, X.; Yang, L.; Jiang, G.; Li, S.; Ding, W. Genome sequencing of Ralstonia solanacearum CQPS-1, a phylotype I strain collected from a highland area with continuous cropping of tobacco. Front. Microbiol. 2017, 8, 974. [Google Scholar] [CrossRef] [PubMed]
- Poueymiro, M.; Cunnac, S.; Barberis, P.; Deslandes, L.; Peeters, N.; Cazale-Noel, A.C.; Boucher, C.; Genin, S. Two type III secretion system effectors from Ralstonia solanacearum GMI1000 determine host-range specificity on tobacco. Mol. Plant Microbe Interact. 2009, 22, 538–550. [Google Scholar] [CrossRef] [PubMed]
- Robertson, A.E.; Wechter, W.P.; Denny, T.P.; Fortnum, B.A.; Kluepfel, D.A. Relationship between avirulence gene (avrA) diversity in Ralstonia solanacearum and bacterial wilt incidence. Mol. Plant Microbe Interact. 2004, 17, 1376–1384. [Google Scholar] [CrossRef]
- Carney, B.F.; Denny, T.P. A cloned avirulence gene from Pseudomonas solanacearum determines incompatibility on Nicotiana tabacum at the host species level. J. Bacteriol. 1990, 172, 4836–4843. [Google Scholar] [CrossRef] [PubMed]
- Yuliar; Nion, Y.A.; Toyota, K. Recent trends in control methods for bacterial wilt diseases caused by Ralstonia solanacearum. Microbes Environ. 2015, 30, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Zhuo, T.; Hu, X.; Fan, X.; Zou, H. Identification of a Pseudomonas putida as biocontrol agent for tomato bacterial wilt disease. Biol. Control. 2017, 114, 45–50. [Google Scholar] [CrossRef]
- Preston, G.M.; Bertrand, N.; Rainey, P.B. Type III secretion in plant growth-promoting Pseudomonas fluorescens SBW25. Mol. Microbiol. 2001, 41, 999–1014. [Google Scholar] [CrossRef] [PubMed]
- King, E.O.; Ward, M.K.; Raney, D.E. Two simple media for the demonstration of pyocyanin and fluorescin. J. Lab. Clin. Med. 1954, 44, 301–307. [Google Scholar] [PubMed]
- Fan, X.; Guo, J.; Zhou, Y.; Zhuo, T.; Zou, H. The ColRS-regulated membrane protein gene XAC1347 is involved in copper homeostasis and hrp gene expression in Xanthomonas citri subsp. citri. Front. Microbiol. 2018, 9, 1171. [Google Scholar] [CrossRef] [PubMed]
- Kovach, M.E.; Phillips, R.W.; Elzer, P.H.; Roop, R.M.; Peterson, K.M. pBBR1MCS: A broad-host-range cloning vector. Biotechniques 1994, 16, 800–802. [Google Scholar]
- Schäfer, A.; Tauch, A.; Jäger, W.; Kalinowski, J.; Thierbach, G.; Pühler, A. Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli plasmids pK18 and pK19: Selection of defined deletions in the chromosome of Corynebacterium glutamicum. Gene 1994, 145, 69–73. [Google Scholar] [CrossRef]
- Miller, J.H. Experiments in Molecular Genetics; CSH Laboratory Press: Cold Spring Harbor, NY, USA, 1972. [Google Scholar]
- Cooke, B.M. Disease assessment and yield loss. In The Epidemiology of Plant Diseases; Cooke, B.M., Jone, D.G., Kaye, B., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 43–80. [Google Scholar]
- Álvarez, B.; Biosca, E.G. Bacteriophage-based bacterial wilt biocontrol for an environmentally sustainable agriculture. Front. Plant Sci. 2017, 8, 1218. [Google Scholar] [CrossRef] [PubMed]
- Elhalag, K.; Nasr-Eldin, M.; Hussien, A.; Ahmad, A. Potential use of soilborne lytic Podoviridae phage as a biocontrol agent against Ralstonia solanacearum. J. Basic Microbiol. 2018, 58, 658–669. [Google Scholar] [CrossRef] [PubMed]
- Malafaia, C.B.; Jardelino, A.C.S.; Silva, A.G.; Souza, E.B.D.; Macedo, A.J.; Correia, M.T.D.S.; Silva, M.V. Effects of Caatinga plant extracts in planktonic growth and biofilm formation in Ralstonia solanacearum. Microb. Ecol. 2018, 75, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Li, S.; Qin, X.; Jiang, G.; Chen, J.; Li, B.; Yao, X.; Liang, P.; Zhang, Y.; Ding, W. Exposure to umbelliferone reduces Ralstonia solanacearum biofilm formation, transcription of type III secretion system regulators and effectors and virulence on tobacco. Front. Microbiol. 2017, 8, 1234. [Google Scholar] [CrossRef] [PubMed]
- Frey, P.; Prior, P.; Marie, C.; Kotoujansky, A.; Trigalet-Demery, D.; Trigalet, A. Hrp- mutants of Pseudomonas solanacearum as potential biocontrol agents of tomato bacterial wilt. Appl. Environ. Microbiol. 1994, 60, 3175–3181. [Google Scholar] [CrossRef] [PubMed]
- Weller, D.M. Pseudomonas biocontrol agents of soilborne pathogens: Looking back over 30 years. Phytopathology 2007, 97, 250–256. [Google Scholar] [CrossRef]
- Dabboussi, F.; Hamze, M.E.; Geoffroy, V.; Meyer, J.M.; Izard, D. Pseudomonas mosselii sp nov., a novel species isolated from clinical specimens. Int. J. Syst. Evol. Microbiol. 2002, 52, 363–376. [Google Scholar] [CrossRef] [PubMed]
- Park, G.S.; Chu, J.H.; Hong, S.J.; Kwak, Y.; Khan, A.R.; Jung, B.K.; Ullah, I.; Shin, J.H. Complete genome sequence of the caprolactam-degrading bacterium Pseudomonas mosselii SJ10 isolated from waste water of a nylon 6 production plant. J. Biotechnol. 2014, 192, 263–264. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Xiao, W.; Chen, G.; Song, D.; Khaskheli, M.A.; Li, P.; Zhang, S.; Feng, G. Identification of Pseudomonas mosselii BS011 gene clusters required for suppression of rice blast fungus Magnaporthe oryzae. J. Biotechnol. 2018, 282, 1–9. [Google Scholar] [CrossRef]
- Zhang, D.; Xu, D.H.; Qiu, J.; Rasmussen-Ivey, C.R.; Liles, M.R.; Beck, B.H. Draft genome sequence of Pseudomonas mosselii Gil3, isolated from catfish and antagonistic against hypervirulent Aeromonas hydrophila. Genome Announc. 2016, 4, e01305–e01316. [Google Scholar] [CrossRef] [PubMed]
- Matilla, M.A.; Ramos, J.L.; Bakker, P.A.; Doornbos, R.; Badri, D.V.; Vivanco, J.M.; Ramos-González, M.I. Pseudomonas putida KT2440 causes induced systemic resistance and changes in Arabidopsis root exudation. Environ. Microbiol. Rep. 2010, 2, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Cunnac, S.; Boucher, C.; Genin, S. Characterization of the cis-acting regulatory element controlling HrpB-mediated activation of the type III secretion system and effector genes in Ralstonia solanacearum. J. Bacteriol. 2004, 186, 2309–2318. [Google Scholar] [CrossRef] [PubMed]
- Marenda, M.; Brito, B.; Callard, D.; Genin, S.; Barberis, P.; Boucher, C.; Arlat, M. PrhA controls a novel regulatory pathway required for the specific induction of Ralstonia solanacearum hrp genes in the presence of plant cells. Mol. Microbiol. 2010, 27, 437–453. [Google Scholar] [CrossRef] [PubMed]
- Nakano, M.; Mukaihara, T. The type III effector RipB from Ralstonia solanacearum RS1000 acts as a major avirulence factor in Nicotiana benthamiana and other Nicotiana species. Mol. Plant Pathol. 2019, 20, 1237–1251. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, G.W.; Delaney, S.K. Stable internal reference genes for normalization of real-time RT-PCR in tobacco (Nicotiana tabacum) during devel-opment and abiotic stress. Mol. Genet Genom. 2010, 283, 233–241. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains or Plasmids | Relevant Characteristics | Resources |
|---|---|---|
| Strains | ||
| Pseudomonas mosselii | ||
| A1 | Wild-type Pseudomonas mosselii isolated from soil | [20] |
| AA1 | Pseudomonas mosselii expressing ripAA gene of Ralstonia solanacearum | This study |
| A1/pBB:lacZ | Gmr, Pseudomonas mosselii A1 harboring pBB:lacZ | This study |
| A1/pBB:PlacZ | Gmr, Pseudomonas mosselii A1 harboring pBB:PlacZ | This study |
| Escherichia coli | ||
| DH5α | F’Φ80dlacZDM15D(lacZYA-argF)U169 endA1 deoR recA1 hsdR17(rK2 mK+) phoA supE44 l2 thi-l gyrA96 relA1 | Clontech |
| Ralstonia solanacearum | ||
| RsT1 | PBr, a Ralstonia solanacearum strain isolated from tobacco plants in Sanming, Fujian, China | Lab collection |
| Plasmids | ||
| pK18mobsacB | Kmr, suicide vector, sacB+ | [24] |
| pK18:RipAA | Kmr, a 2.3 kb fusion containing 5′ and 3′ terminal sequences of a hypothetical gene in Pseudomonas mosselii A1, which were interspaced by 1.1 kb DNA fragment of ripAA gene with native promoter | This study |
| pBBR1MCS-5 | Gmr, 4.7 kb broad-host range plasmid, lacZ | [25] |
| pLacZ-Basic | Apr, 7.5 kb pUC replication origin plasmid carrying a β-galactosidase gene | Clontech |
| pBB:lacZ | Gmr, pBBR1MCS-5 harboring a 4.6 kb KpnI/SalI fragment that contains a β-galactosidase gene from pLacZ-Basic | This study |
| pBB:PlacZ | Gmr, pBBR1MCS-5 harboring a 276-bp ripAA promoter fused with the 4.6 kb KpnI/SalI fragment from pLacZ-Basic | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhuo, T.; Chen, S.; Wang, D.; Fan, X.; Zhang, X.; Zou, H. Expression of the ripAA Gene in the Soilborne Pseudomonas mosselii Can Promote the Control Efficacy against Tobacco Bacterial Wilt. Biology 2022, 11, 1170. https://doi.org/10.3390/biology11081170
Zhuo T, Chen S, Wang D, Fan X, Zhang X, Zou H. Expression of the ripAA Gene in the Soilborne Pseudomonas mosselii Can Promote the Control Efficacy against Tobacco Bacterial Wilt. Biology. 2022; 11(8):1170. https://doi.org/10.3390/biology11081170
Chicago/Turabian StyleZhuo, Tao, Shiting Chen, Dandan Wang, Xiaojing Fan, Xiaofeng Zhang, and Huasong Zou. 2022. "Expression of the ripAA Gene in the Soilborne Pseudomonas mosselii Can Promote the Control Efficacy against Tobacco Bacterial Wilt" Biology 11, no. 8: 1170. https://doi.org/10.3390/biology11081170
APA StyleZhuo, T., Chen, S., Wang, D., Fan, X., Zhang, X., & Zou, H. (2022). Expression of the ripAA Gene in the Soilborne Pseudomonas mosselii Can Promote the Control Efficacy against Tobacco Bacterial Wilt. Biology, 11(8), 1170. https://doi.org/10.3390/biology11081170