
Complex Network Model Reveals the Impact of Inspiratory Muscle Pre-Activation on Interactions among Physiological Responses and Muscle Oxygenation during Running and Passive Recovery

 ,
,  , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Subjects
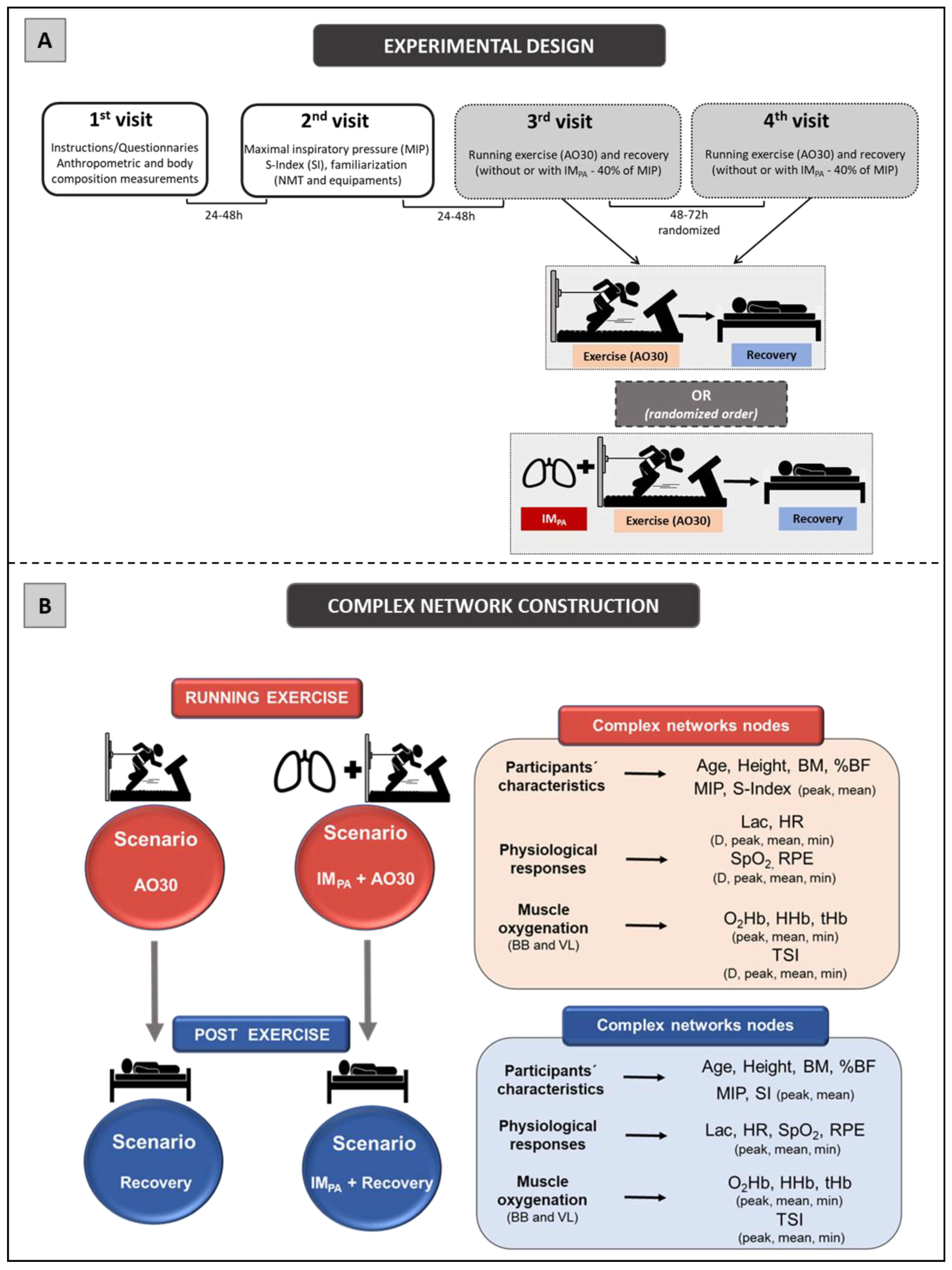
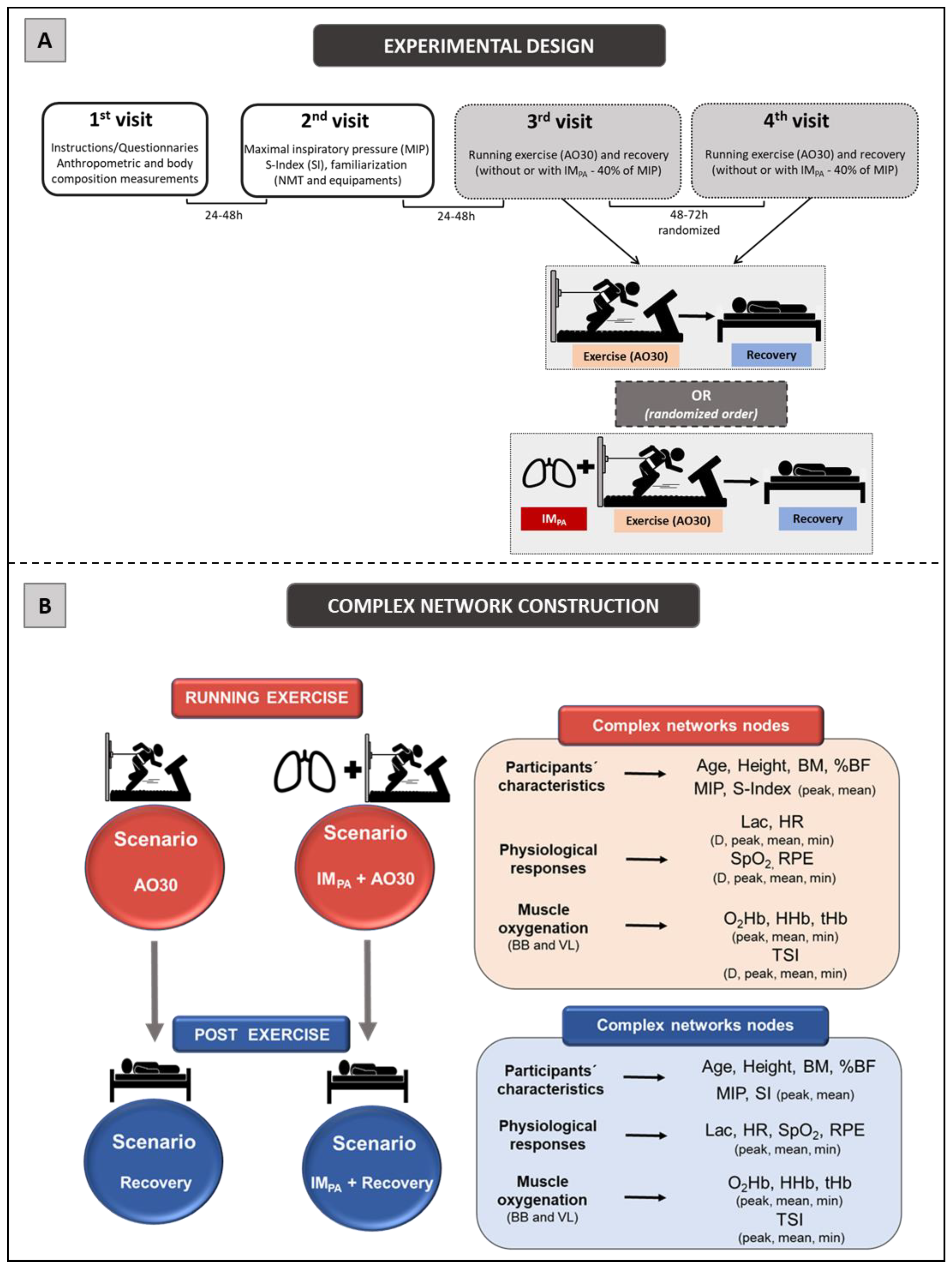
2.2. Experimental Design
2.3. Maximal Inspiratory Pressure and S-Index Determination
2.4. Inspiratory Muscle Pre-Activation
2.5. High-Intensity Running Effort and Post-Exercise
2.6. Power in High-Intensity Tethered Exercise
2.7. Muscle Oxygenation Measurements and Analyses
2.8. Arterial Oxygen Saturation, Heart Rate and Blood Lactate Concentration
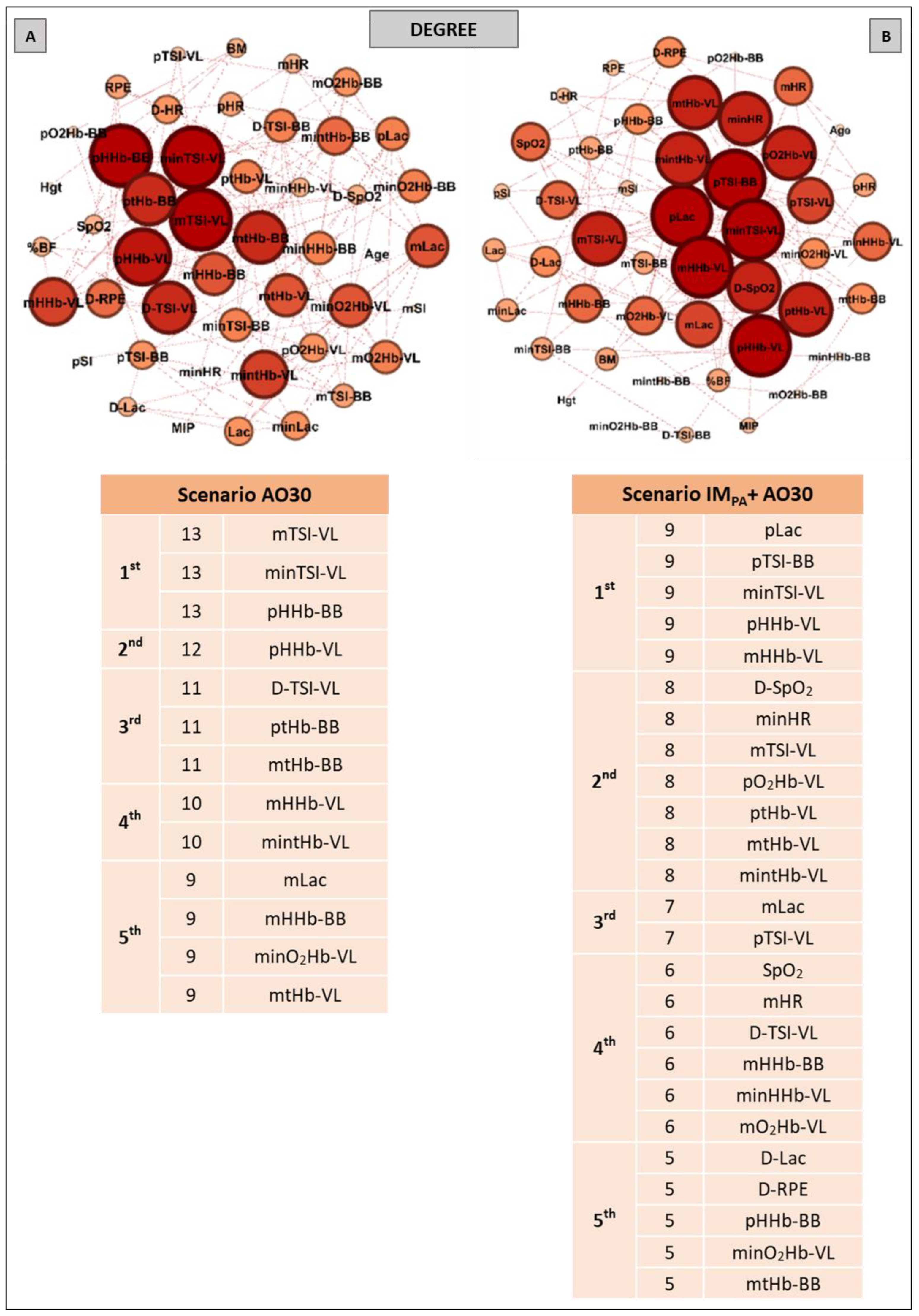
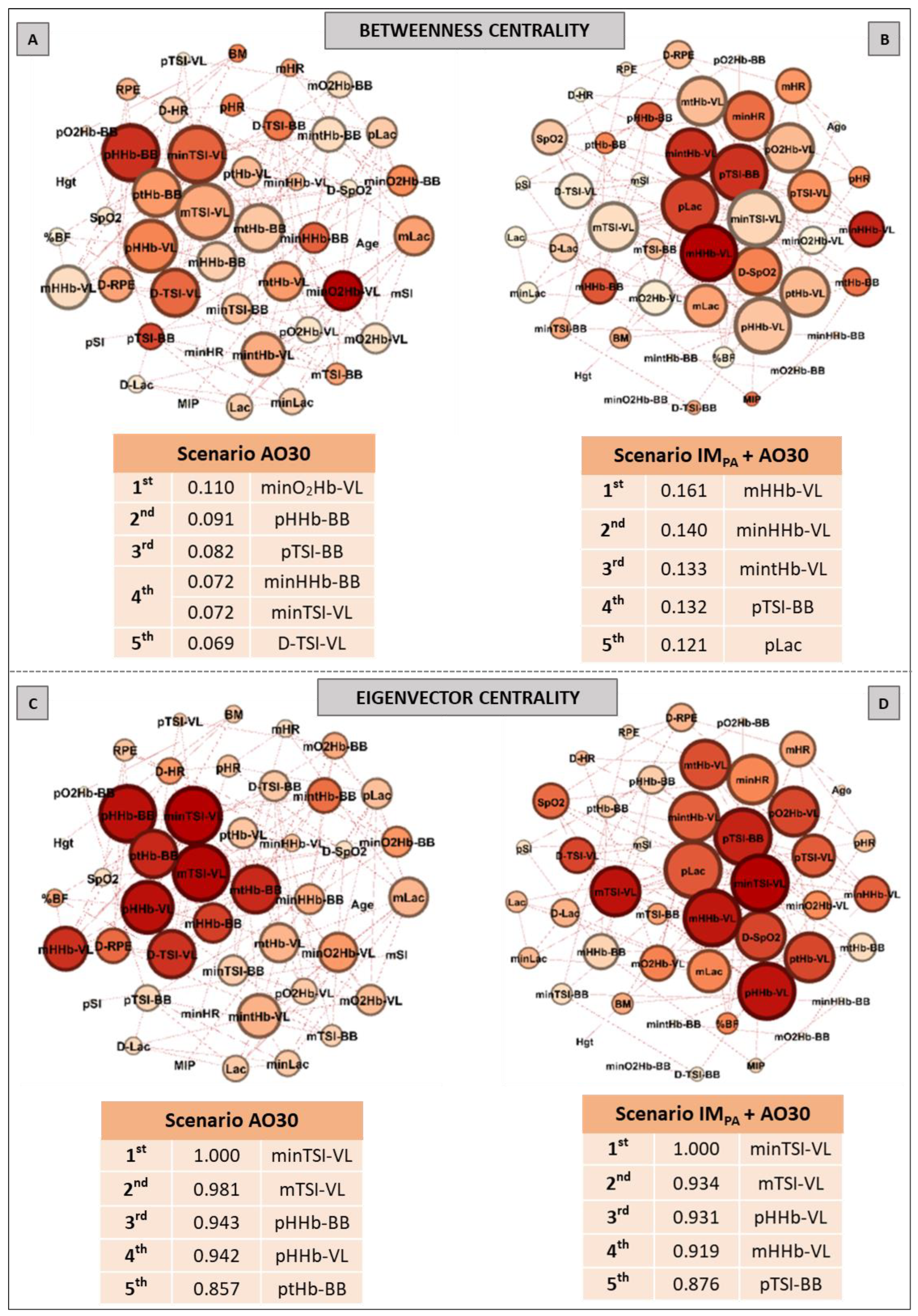
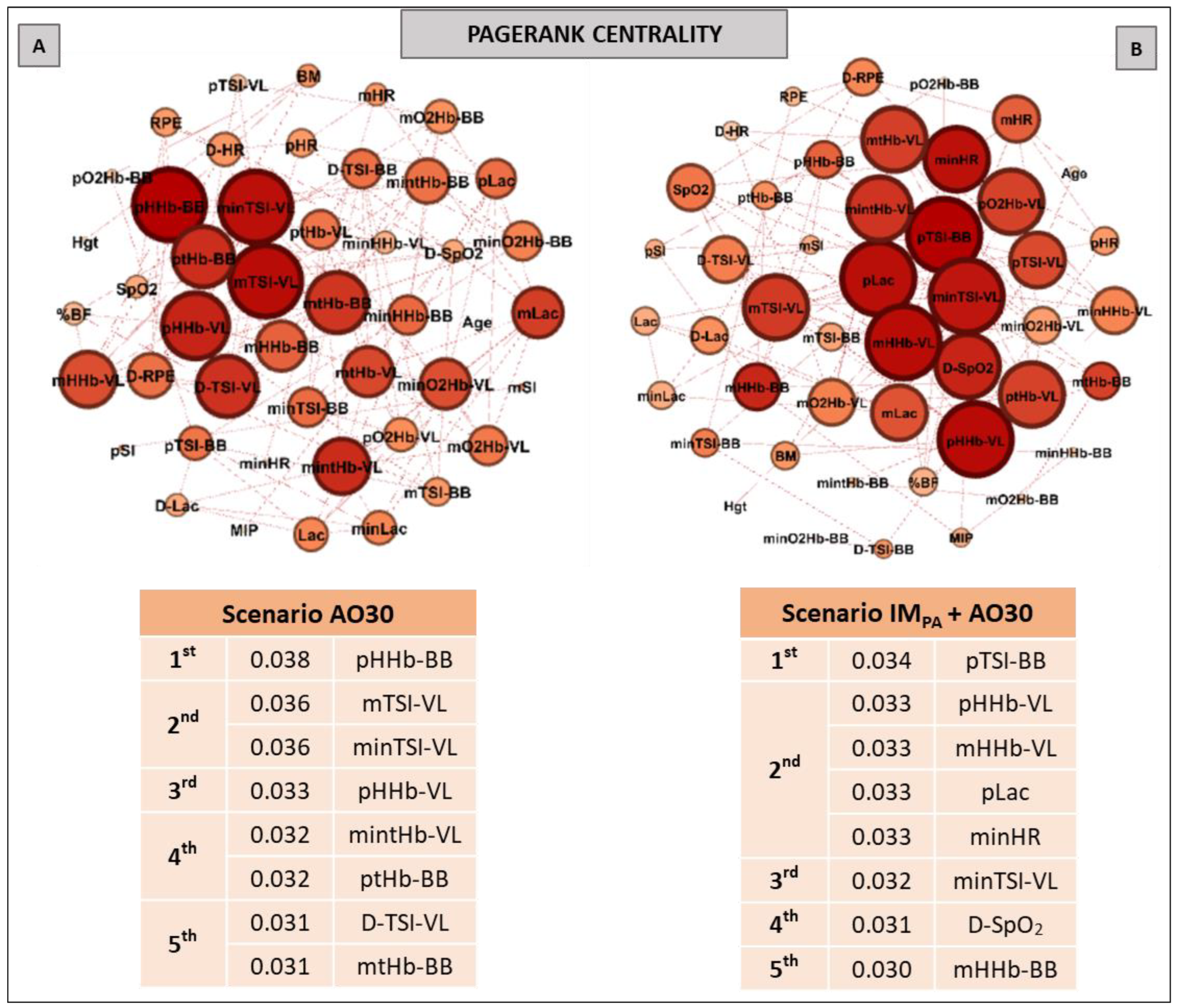
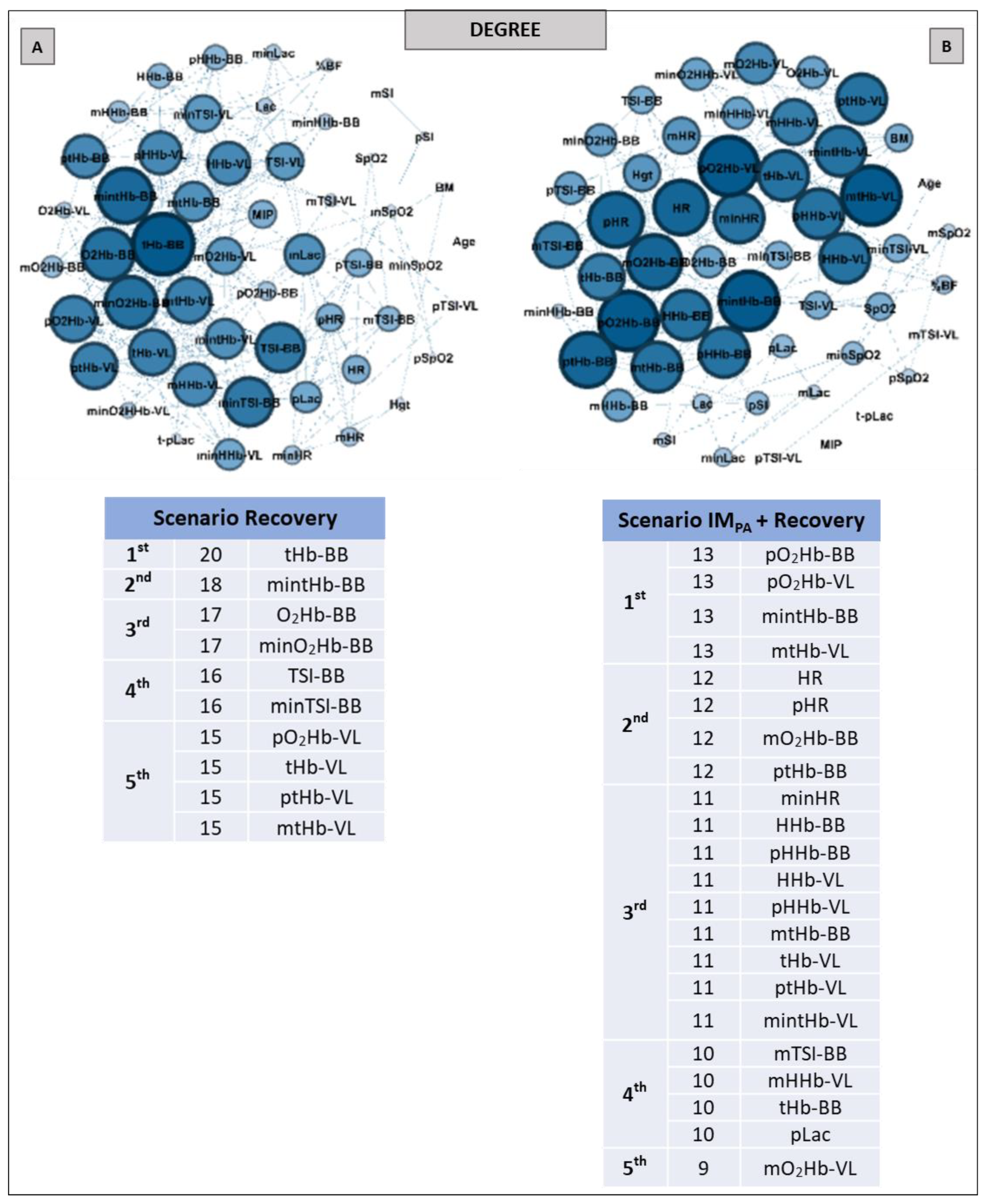
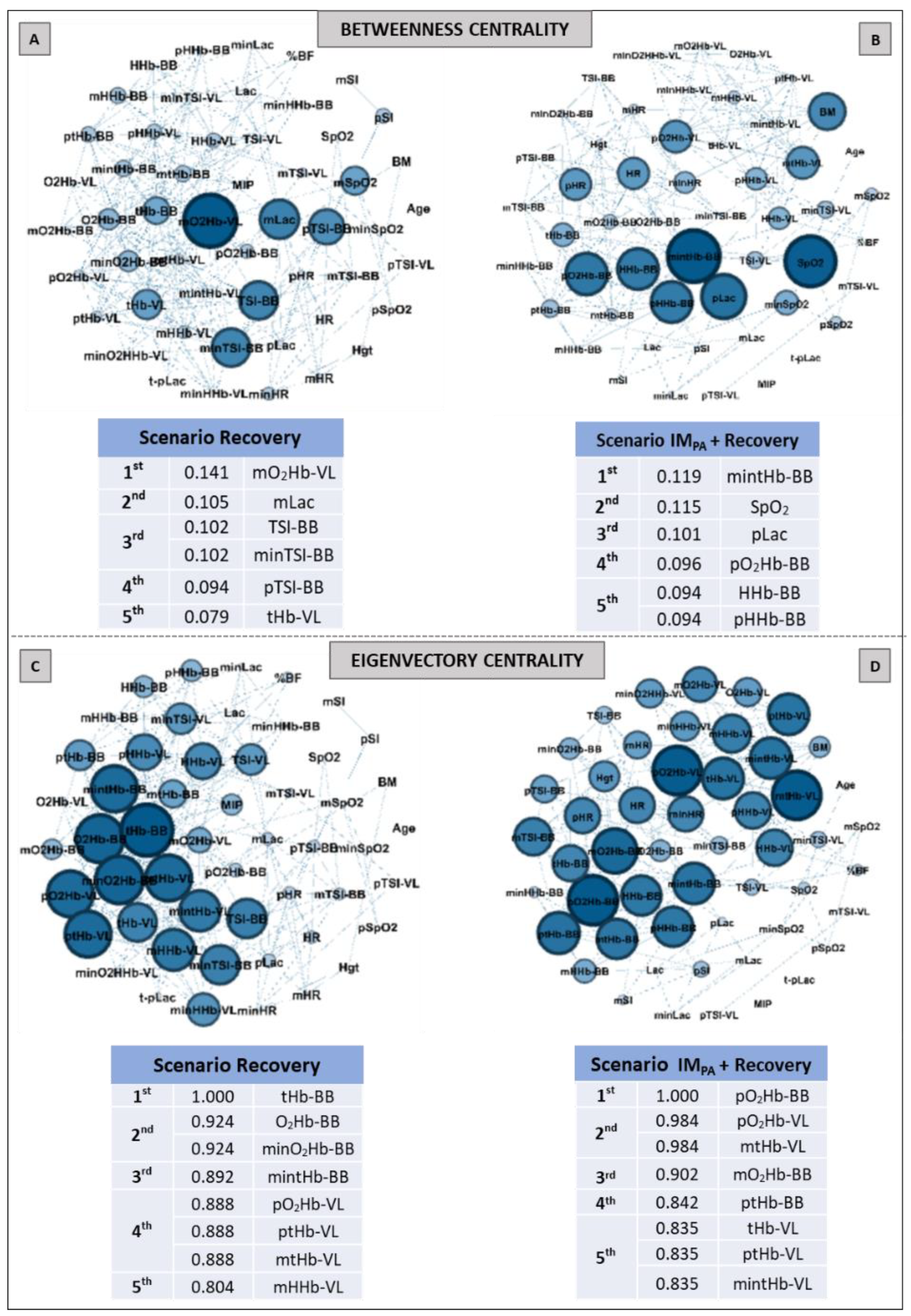
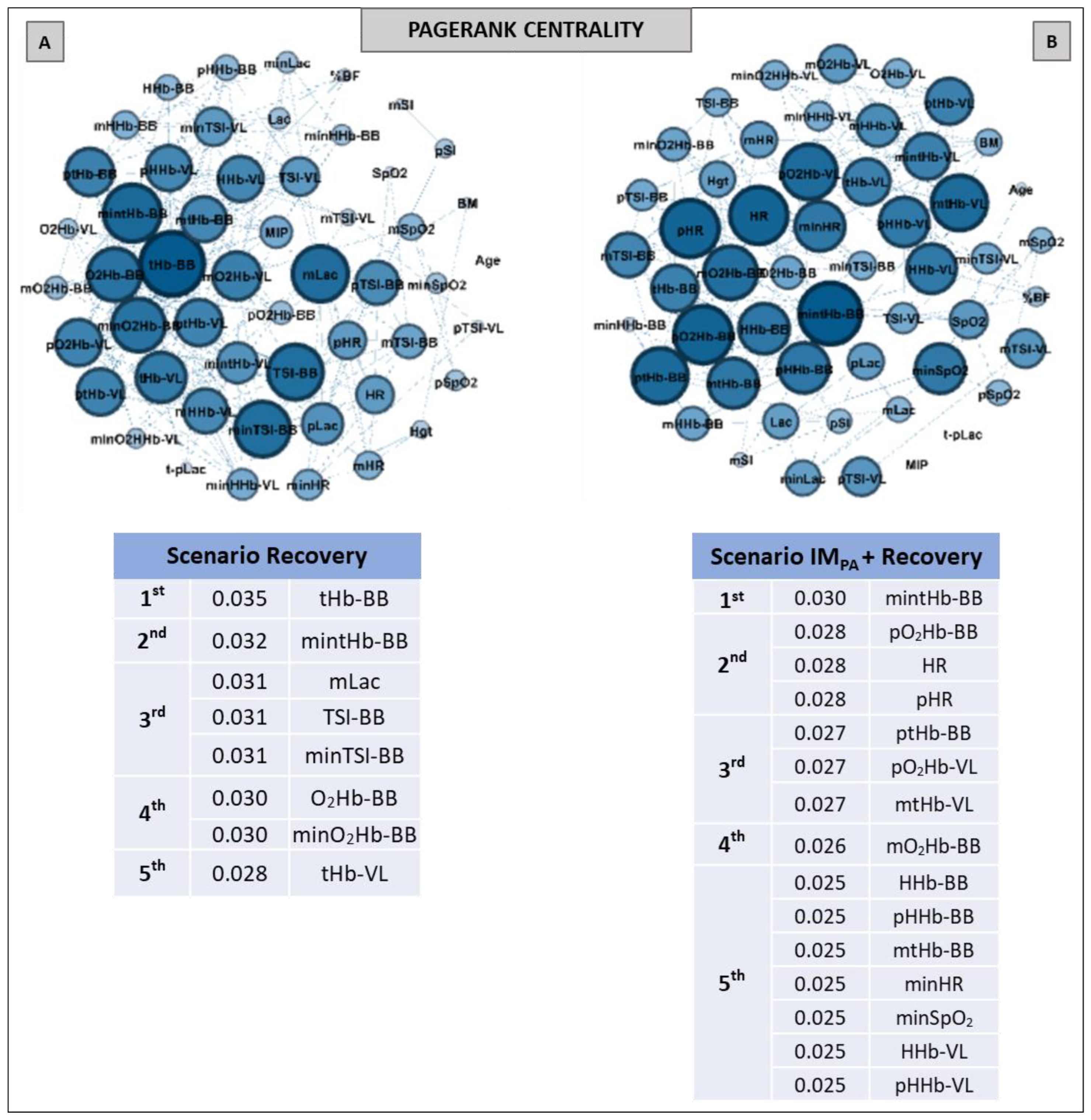
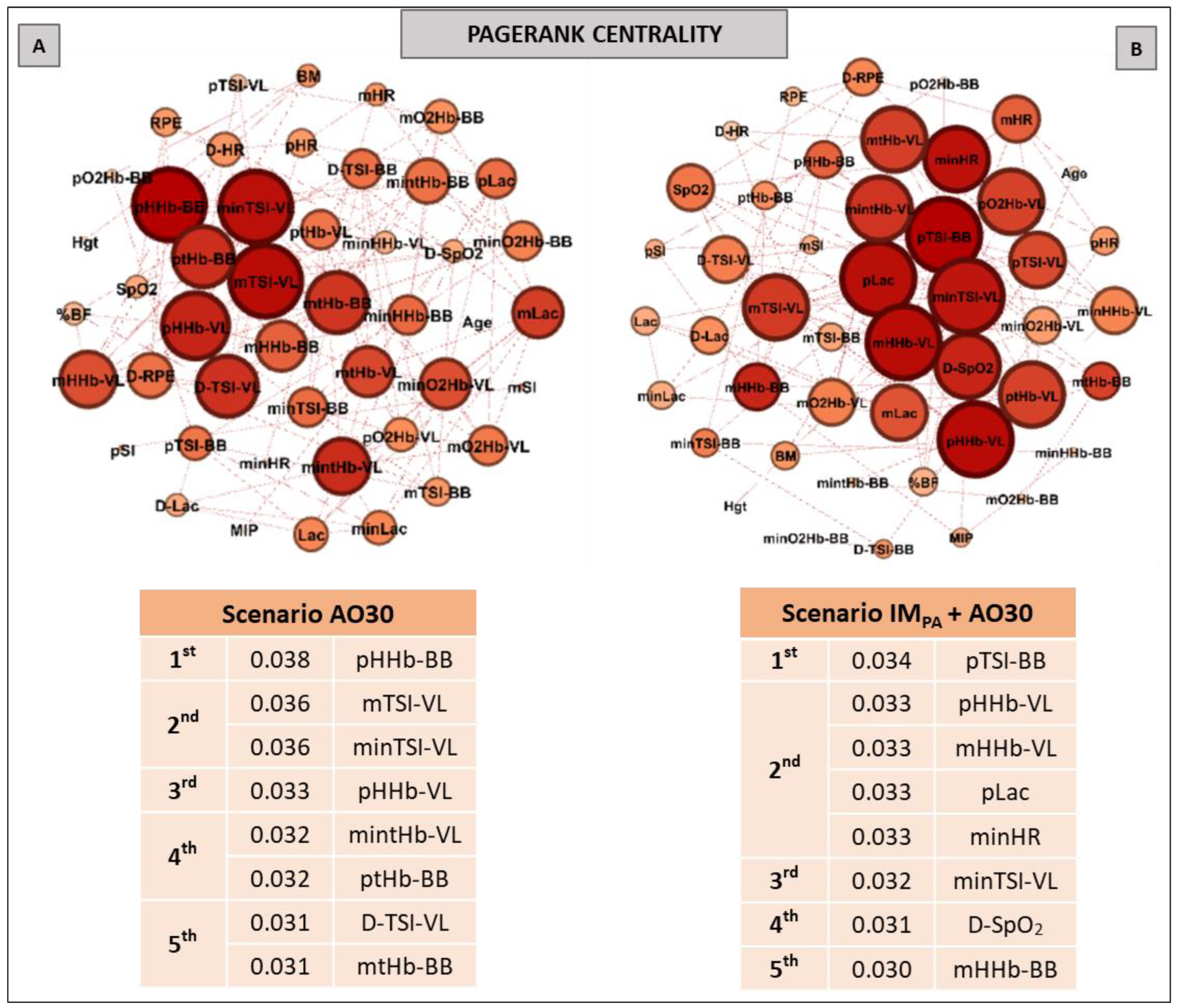
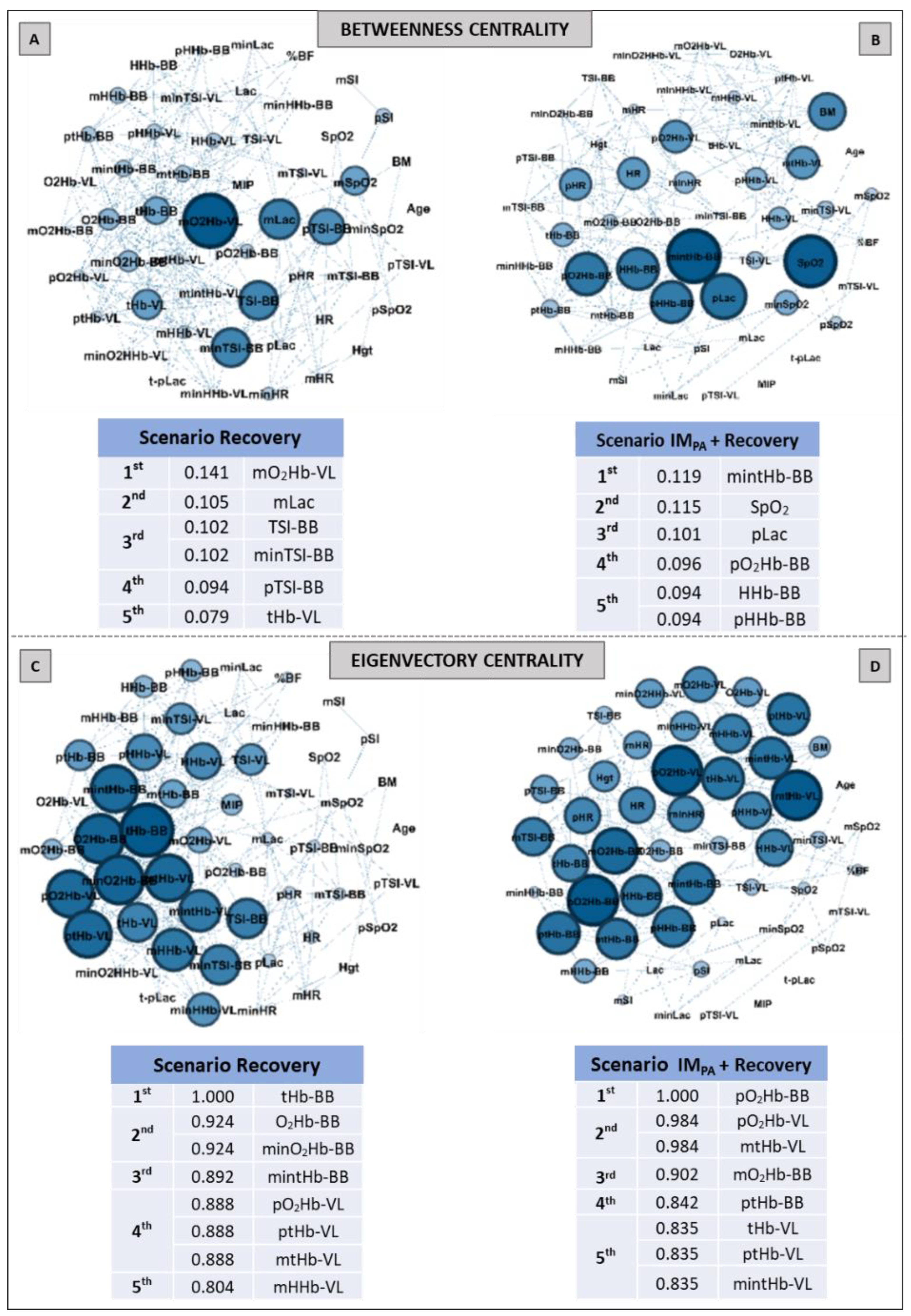
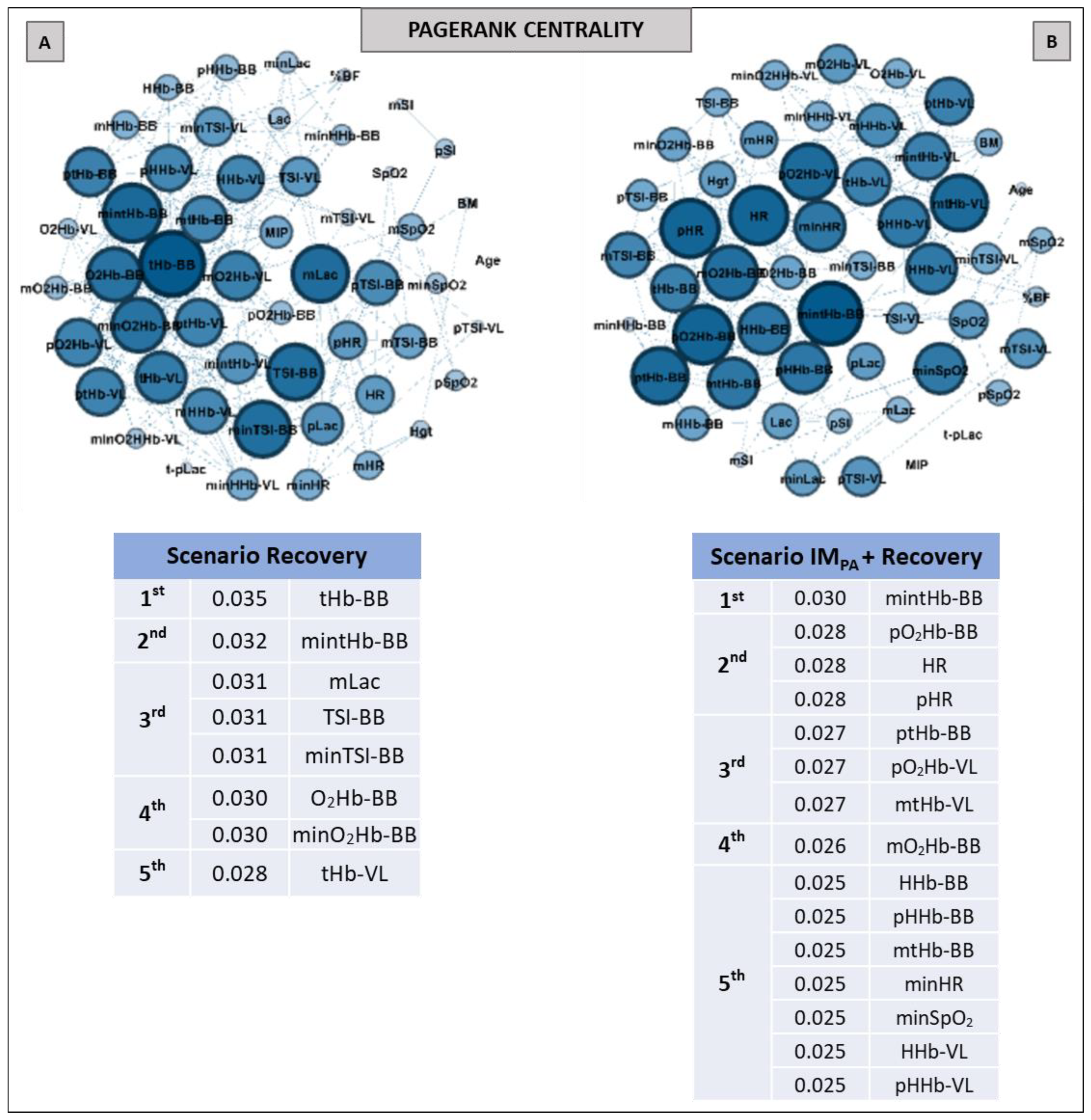
2.9. Complex Network Construction: Scenarios and Centrality Metrics
2.10. Statistical Analysis
3. Results
4. Discussion
5. Centrality Metrics in Exercise Scenarios
6. Centrality Metric in Recovery Scenarios
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sheel, A.W. Respiratory muscle training in healthy individuals: Physiological rationale and implications for exercise performance. Sports Med. 2002, 32, 567–581. [Google Scholar] [CrossRef]
- Brown, P.I.; Sharpe, G.R.; Johnson, M.A. Loading of trained inspiratory muscles speeds lactate recovery kinetics. Med. Sci. Sports Exerc. 2010, 42, 1103–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, T.K.; Baker, J.S.; Zhang, H.; Kong, Z.; Nie, J. Effects of Specific Core Re-Warm-Ups on Core Function, Leg Perfusion and Second-Half Team Sport-Specific Sprint Performance: A Randomized Crossover Study. J. Sports Sci. Med. 2019, 18, 479–489. [Google Scholar] [PubMed]
- Lorca-Santiago, J.; Jiménez, S.L.; Pareja-Galeano, H.; Lorenzo, A. Inspiratory muscle training in intermittent sports modalities: A systematic review. Int. J. Environ. Res. Public Health 2020, 17, 4448. Available online: https://www.mdpi.com/1660-4601/17/12/44484448 (accessed on 4 August 2020). [CrossRef] [PubMed]
- Antonelli, C.B.B.; Hartz, C.S.; Santos, S.D.S.; Moreno, M.A. Effects of inspiratory muscle training with progressive loading on respiratory muscle function and sports performance in high-performance wheelchair basketball athletes: A randomized clinical trial. Int. J. Sports Physiol. Perform. 2019, 15, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Hartz, C.S.; Sindorf, M.A.G.; Lopes, C.R.; Batista, J.; Moreno, M.A. Effect of inspiratory muscle training on performance of handball athletes. J. Hum. Kinet. 2018, 63, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Karsten, M.; Ribeiro, G.S.; Esquivel, M.S.; Matte, D.L. The effects of inspiratory muscle training with linear workload devices on the sports performance and cardiopulmonary function of athletes: A systematic review and meta-analysis. Phys. Ther. Sport 2018, 34, 92–104. [Google Scholar] [CrossRef] [PubMed]
- Tong, T.K.; McConnell, A.K.; Lin, H.; Nie, J.; Zhang, H.; Wang, J. “Functional” Inspiratory and Core Muscle Training Enhances Running Performance and Economy. J. Strength Cond. Res. 2016, 30, 2942–2951. [Google Scholar] [CrossRef] [Green Version]
- DeLucia, C.M.; De Asis, R.M.; Bailey, E.F. Daily inspiratory muscle training lowers blood pressure and vascular resistance in healthy men and women. Exp. Physiol. 2018, 103, 201–211. [Google Scholar] [CrossRef] [Green Version]
- Illi, S.K.; Held, U.; Frank, I.; Spengler, C.M. Effect of respiratory muscle training on exercise performance in healthy individuals: A systematic review and meta-analysis. Sports Med. 2012, 42, 707–724. [Google Scholar] [CrossRef] [PubMed]
- Basso-Vanelli, R.P.; Di Lorenzo, V.A.P.; Labadessa, I.G.; Regueiro, E.M.; Jamami, M.; Gomes, E.L.; Costa, D. Effects of inspiratory muscle training and calisthenics-and-breathing exercises in CPPD with and without respiratory muscle weakness. Respir. Care 2016, 61, 50–60. [Google Scholar] [CrossRef] [Green Version]
- Cahalin, L.P.; Arena, R.A. Breathing exercises and inspiratory muscle training in heart failure. Heart Fail. Clin. 2015, 11, 149–172. [Google Scholar] [CrossRef] [PubMed]
- Dall’Ago, P.; Chiappa, G.R.; Guths, H.; Stein, R.; Ribeiro, J.P. Inspiratory muscle training in patients with heart failure and inspiratory muscle weakness: A randomized trial. J. Am. Coll. Cardiol. 2006, 47, 757–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- HajGhanbari, B.; Yamabayashi, C.; Buna, T.R.; Coelho, J.D.; Freedman, K.D.; Morton, T.A.; Palmer, S.A.; Toy, M.A.; Walsh, C.; Sheel, A.W.; et al. Effects of respiratory muscle training on performance in athletes: A systematic review with 29 meta-analyses. J. Strength Cond. Res. 2013, 27, 1643–1663. [Google Scholar] [CrossRef] [PubMed]
- Guenette, J.A.; Sheel, A.W. Physiological consequences of a high work of breathing during heavy exercise in humans. J. Sci. Med. Sport 2007, 10, 341–350. [Google Scholar] [CrossRef]
- Özdal, M. Acute effects of inspiratory muscle warm-up on pulmonary function in healthy subjects. Respir. Physiol. Neurobiol. 2016, 227, 23–26. [Google Scholar] [CrossRef]
- Volianitis, S.; McConnell, A.K.; Jones, D.A. Assessment of maximum inspiratory pressure. Prior submaximal respiratory muscle activity (‘warm-up’) enhances maximum inspiratory activity and attenuates the learning effect of repeated measurement. Respiration 2001, 68, 22–27. [Google Scholar] [CrossRef] [Green Version]
- Barnes, K.R.; Ludge, A.R. Inspiratory muscle warm-up improves 3200-m running performance in distance runners. J. Strength Cond. Res. 2019, 35, 1739–1747. [Google Scholar] [CrossRef]
- Wilson, E.E.; McKeever, T.M.; Lobb, C.; Sherriff, T.; Gupta, L.; Hearson, G.; Martin, N.; Lindley, M.R.; Shaw, D.E. Respiratory muscle specific warm-up and elite swimming performance. Br. J. Sports Med. 2014, 48, 789–791. [Google Scholar] [CrossRef]
- Lin, H.; Tong, T.K.; Huang, C.; Nie, J.; Lu, K.; Quach, B. Specific inspiratory muscle warm-up enhances badminton footwork performance. Appl. Physiol. Nutr. Metab. 2007, 32, 1082–1088. [Google Scholar] [CrossRef]
- Cirino, C.; Gobatto, C.A.; Pinto, A.S.; Torres, R.S.; Hartz, C.S.; Azevedo, P.H.; Moreno, M.A.; Manchado-Gobatto, F.B. Complex network model indicates a positive effect of inspiratory muscles pre-activation on performance parameters in a Judo match. Sci. Rep. 2021, 11, 11148. [Google Scholar] [CrossRef] [PubMed]
- Tong, T.K.; Fu, F.H. Effect of specific inspiratory muscle warm-up on intense intermittent run to exhaustion. Eur. J. Appl. Physiol. 2006, 97, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Hill, K.; Eastwood, P. Effects of loading on upper airway and respiratory pump muscle motoneurons. Respir. Physiol. Neurobiol. 2011, 179, 64–70. [Google Scholar] [CrossRef]
- Raux, M.; Straus, C.; Redolfi, S.; Morelot-Panzini, C.; Couturier, A.; Hug, F.; Similowski, T. Electroencephalographic evidence for pre-motor cortex activation during inspiratory loading in humans. J. Physiol. 2007, 578, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Ross, E.Z.; Nowicky, A.V.; McConnell, A.K. Influence of acute inspiratory loading upon diaphragm motor-evoked potentials in healthy humans. J. Appl. Physiol. 2007, 102, 1883–1890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooks, G.A. Lactate as a fulcrum of metabolism. Redox Biol. 2020, 35, 101454. Available online: https://www.sciencedirect.com/science/article/pii/S2213231720300422 (accessed on 20 August 2020). [CrossRef] [PubMed]
- Faude, O.; Kindermann, W.; Meyer, T. Lactate threshold concepts: How valid are they? Sports Med. 2009, 39, 469–490. [Google Scholar] [CrossRef]
- Billat, V.L.; Sirvent, P.; Py, G.; Kolarlsztein, J.P.; Mercier, J. The concept of maximal lactate steady state: A bridge between biochemistry, physiology and sport science. Sports Med. 2003, 20, 407–426. [Google Scholar] [CrossRef]
- Perrey, S.; Ferrari, M. Muscle oximetry in sports science: A systematic review. Sports Med. 2018, 48, 597–616. [Google Scholar] [CrossRef]
- Ferrari, M.; Muthalib, M.; Quaresima, V. The use of near-infrared spectroscopy in understanding skeletal muscle physiology: Recent developments. Philos. Trans. A Math. Phys. Eng. Sci. 2011, 369, 4577–4590. [Google Scholar] [CrossRef]
- Lagerwaard, B.; Keijer, J.; McCully, K.K.; de Boer, V.C.J.; Nieuwenhuizen, A.G. In vivo assessment of muscle mitochondrial function in healthy, young males in relation to parameters of aerobic fitness. Eur. J. Appl. Physiol. 2019, 119, 1799–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monroe, J.C.; Song, Q.; Emery, M.S.; Hirai, D.M.; Motaganahalli, R.L.; Roseguini, B.T. Acute effects of leg heat therapy on walking performance and cardiovascular and inflammatory responses to exercise in patients with peripheral artery disease. Physiol. Rep. 2021, 8, e14650. [Google Scholar] [CrossRef] [PubMed]
- Willis, S.J.; Borrani, F.; Millet, G.P. Leg-vs arm-cycling repeated sprints with blood flow restriction and systemic hypoxia. Eur. J. Appl. Physiol. 2019, 119, 1819–1828. [Google Scholar] [CrossRef] [PubMed]
- Manchado-Gobatto, F.B.; Marostegan, A.B.; Rasteiro, F.M.; Cirino, C.; Cruz, J.P.; Moreno, M.A.; Gobatto, C.A. New insights into mechanical, metabolic and muscle oxygenation signals during and after high-intensity tethered running. Sci. Rep. 2020, 10, 6336. Available online: https://www.nature.com/articles/s41598-020-63297-w6336 (accessed on 14 April 2020). [CrossRef] [Green Version]
- Osawa, T.; Shiose, K.; Takahashi, H. Delayed onset of reoxygenation in inactive muscles after high-intensity exercise. Adv. Exp. Med. Biol. 2017, 977, 255–260. [Google Scholar] [CrossRef]
- Hesford, C.M.; Laing, S.; Cooper, C.E. Using portable NIRS to compare arm and leg muscle oxygenation during roller skiing in biathletes: A case study. Adv. Exp. Med. Biol. 2013, 789, 179–184. [Google Scholar] [CrossRef]
- Rissanen, A.P.E.; Tikkanen, H.O.; Koponen, A.S.; Aho, J.M.; Hägglund, H.; Lindholm, H.; Peltonen, J.E. Alveolar gas exchange and tissue oxygenation during incremental treadmill exercise, and their associations with blood O2 carrying capacity. Front. Physiol. 2012, 3, 265. [Google Scholar] [CrossRef] [Green Version]
- Ohya, T.; Hagiwara, M.; Suzuki, Y. Inspiratory muscle warm-up has no impact on performance or locomotor muscle oxygenation during high-intensity intermittent sprint cycling exercise. SpringerPlus 2005, 4, 556. [Google Scholar] [CrossRef] [Green Version]
- Richard, P.; Billaut, F. Effects of inspiratory muscle warm-up on locomotor muscle oxygenation in elite speed skaters during 3000 m time trials. Eur. J. Appl. Physiol. 2019, 119, 191–200. [Google Scholar] [CrossRef]
- Pereira, V.H.; Gobatto, C.A.; Lewis, T.G.; Ribeiro, L.F.; Beck, W.R.; Dos Reis, I.G.; Sousa, F.A.; Manchado-Gobatto, F.B. Computational and complex network modeling for analysis of sprinter athletes’ performance in track field tests. Front. Physiol. 2018, 9, 843. [Google Scholar] [CrossRef] [Green Version]
- Pereira, V.H.; Gama, M.C.; Sousa, F.A.; Lewis, T.G.; Gobatto, C.A.; Manchado-Gobatto, F.B. Complex network models reveal correlations among network metrics, exercise intensity and role of body changes in the fatigue process. Sci. Rep. 2015, 5, 10489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gobatto, C.A.; Torres, R.S.; Moura, F.A.; Cunha, S.A.; Giometti, C.B.; Araujo, G.G.; Sousa, F.A.B.; Manchado-Gobatto, F.B. Corresponding Assessment Scenarios in Laboratory and on-Court Tests: Centrality Measurements by complex networks analysis in young basketball players. Sci. Rep. 2020, 10, 8620. Available online: https://www.nature.com/articles/s41598-020-65420-3 (accessed on 25 May 2020). [CrossRef] [PubMed]
- Breda, F.L.; Manchado-Gobatto, F.B.; de Barros Sousa, F.A.; Beck, W.R.; Pinto, A.; Papoti, M.; Scariot, P.P.M.; Gobatto, C.A. Complex networks analysis reinforces centrality hematological role on aerobic-anaerobic performances of the Brazilian Paralympic endurance team after altitude training. Sci. Rep. 2022, 12, 1148. [Google Scholar] [CrossRef] [PubMed]
- Lewis, T.G. Network Science: Theory and Applications; Wiley: Hoboken, NJ, USA, 2009; 524p. [Google Scholar]
- Newman, M.E. The structure and function of complex networks. SIAM Rev. 2003, 45, 167–256. [Google Scholar] [CrossRef] [Green Version]
- Jeong, H.; Mason, S.P.; Barabási, A.L.; Oltvai, Z.N. Lethality and centrality in protein networks. Nature 2001, 411, 41–42. [Google Scholar] [CrossRef] [Green Version]
- Long, X.; Fonseca, P.; Aarts, R.A.; Haakma, R.; Foussier, J. Modeling cardiorespiratory interaction during human sleep with complex networks. Appl. Phys. Lett. 2014, 105, 203701. [Google Scholar] [CrossRef] [Green Version]
- Bashan, A.; Bartsch, R.P.; Kantelhardt, J.W.; Havlin, S.; Ivanov, P.C. Network physiology reveals relations between network topology and physiological function. Nat. Commun. 2012, 3, 702. Available online: https://www.nature.com/articles/ncomms1705 (accessed on 28 February 2012). [CrossRef] [Green Version]
- Ma’ayan, A. Introduction to network analysis in systems biology. Sci. Signal. 2011, 4, tr5. Available online: https://stke.sciencemag.org/content/4/190/tr5.long (accessed on 13 September 2011). [CrossRef] [Green Version]
- Vinayagam, A.; Stelzl, U.; Foulle, R.; Plassmann, S.; Zenkner, M.; Timm, J.; Assmus, H.E.; Andrade-Navarro, M.A.; Wanker, E.E. A directed protein interaction network for investigating intracellular signal transduction. Sci. Signal. 2011, 4, rs8. Available online: https://stke.sciencemag.org/content/4/189/rs8.long (accessed on 6 September 2011). [CrossRef]
- Lambert, E.V.; St Clair Gibson, A.; Noakes, T.D. Complex systems model of fatigue: Integrative homoeostatic control of peripheral physiological systems during exercise in humans. Br. J. Sports Med. 2005, 39, 52–62. [Google Scholar] [CrossRef] [Green Version]
- Oldham, S.; Fulcher, B.; Parkes, L.; Arnatkevičiūtė, A.; Suo, C.; Fornito, A. Consistency and differences between centrality measures across distinct classes of networks. PLoS ONE 2019, 14, e0220061. Available online: https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0220061 (accessed on 26 July 2019). [CrossRef] [PubMed] [Green Version]
- Brandes, U. A faster algorithm for betweenness centrality. J. Math. Sociol. 2001, 25, 163–177. [Google Scholar] [CrossRef]
- Freeman, L.C. Centrality in social networks conceptual clarification. Soc. Netw. 1978, 1, 215–239. [Google Scholar] [CrossRef] [Green Version]
- Hua, J.; Huang, M.; Huang, C. Centrality metrics’ performance comparisons on stock market datasets. Symmetry 2019, 11, 916. [Google Scholar] [CrossRef] [Green Version]
- Bonacich, P. Factoring and weighting approaches to status scores and clique identification. J. Math. Sociol. 1972, 2, 113–120. [Google Scholar] [CrossRef]
- Ainsworth, B.E.; Haskell, W.L.; Whitt, M.C.; Irwin, M.L.; Swartz, A.M.; Strath, S.J.; OBrien, W.L.; Bassett, D.R.; Schmitz, K.H.; Emplaincourt, P.O.; et al. Compendium of physical activities: An update of activity codes and MET intensities. Med. Sci. Sports Exerc. 2000, 32, S498–S504. [Google Scholar] [CrossRef] [Green Version]
- Jackson, A.S.; Pollock, M.L. Generalized equations for predicting body density of men. Br. J. Nutr. 1978, 40, 497–504. [Google Scholar] [CrossRef] [Green Version]
- Siri, W.E. Techniques for Measuring Body Composition—Session 4; The National Academies Press: Washington, DC, USA, 1961. [Google Scholar]
- Borg, G.A. Psychophysical bases of perceived exertion. Med. Sci. Sports Exerc. 1982, 14, 377–381. [Google Scholar] [CrossRef]
- Neder, J.A.; Andreoni, S.; Lerario, M.C.; Nery, L.E. References values for lung function tests. II. Maximal respiratory pressures and voluntary ventilation. Braz. J. Med. Biol. Res. 1999, 32, 719–727. [Google Scholar] [CrossRef]
- Minahan, C.; Sheehan, B.; Doutreband, R.; Kirkwood, T.; Reeves, D.; Cross, T. Repeated-sprint cycling does not induce respiratory muscle fatigue in active adults: Measurements from the Powerbreathe® inspiratory muscle trainer. J. Sports Sci. Med. 2015, 14, 233–238. [Google Scholar]
- Zhang, Z.; Wang, B.; Gong, H.; Xu, G.; Nioka, S.; Chance, B. Comparisons of muscle oxygenation changes between arm and leg muscles during incremental rowing exercise with near-infrared spectroscopy. J. Biomed. Opt. 2010, 15, 017007. Available online: https://www.spiedigitallibrary.org/journals/journal-of-biomedical-optics/volume-15/issue-01/017007/Comparisons-of-muscle-oxygenation-changes-between-arm-and-leg-muscles/10.1117/1.3309741.full (accessed on 1 January 2010). [CrossRef] [PubMed]
- Kitada, T.; Machida, S.; Naito, H. Influence of muscle fibre composition on muscle oxygenation during maximal running. BMJ Open Sport Exerc. Med. 2015, 1, e000062. Available online: https://bmjopensem.bmj.com/content/1/1/e000062 (accessed on 25 November 2015). [CrossRef] [PubMed]
- Fruchterman, T.M.J.; Reingold, E.M. Graph drawing by force-directed placement. Softw. Pract. Exp. 1991, 21, 1129–1164. [Google Scholar] [CrossRef]
- Hartz, C.S.; Razera, C.; Moreno, M.A. Effects of the application of an inspiratory muscular warm-up protocol in the physical performance of handball athletes. J. Exerc. Physiol. 2017, 20, 90–97. [Google Scholar]
- Verschueren, J.; Tassignon, B.; De Pauw, K.; Proost, M.; Teugels, A.; Van Cutsem, J.; Roelands, B.; Verhagen, E.; Meeusen, R. Does acute fatigue negatively affect intrinsic risk factors of the lower extremity injury risk profile? A systematic and critical review. Sports Med. 2020, 50, 767–784. [Google Scholar] [CrossRef]
- Balagué, N.; Hristovski, R.; Almarcha, M.; Garcia-Retortillo, S.; Ivanov, P.C. Network physiology of exercise: Vision and perspectives. Front. Physiol. 2020, 11, 611550. [Google Scholar] [CrossRef]
- Perrey, S. Muscle oxygenation unlocks the secrets of physiological responses to exercise: Time to exploit it in the training monitoring. Front. Sports Act. Living 2022, 4, 864825. [Google Scholar] [CrossRef]
- Jalili, M.; Salehzadeh-Yazdi, A.; Asgari, Y.; Arab, S.S.; Yaghmaie, M.; Ghavamzadeh, A.; Alimoghaddam, K. CentiServer: A comprehensive resource, web-based application and R package for centrality analysis. PLoS ONE 2015, 10, e0143111. Available online: https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0143111 (accessed on 16 November 2015). [CrossRef]
- Albert, R.; Barabási, A.L. Statistical mechanics of complex networks. Rev. Mod. Phys. 2002, 74, 47–97. [Google Scholar] [CrossRef] [Green Version]
- Riondato, M.; Kornaropoulos, E.M. Fast approximation of betweenness centrality through sampling. Data Min. Knowl. Discov. 2016, 30, 438–475. [Google Scholar] [CrossRef]
- Page, L.; Brin, S.; Motwani, R.; Winograd, T. The PageRank citation ranking: Bringing order to the web. World Wide Web Internet Web Inf. Syst. 1998, 54, 1–17. [Google Scholar]
- Brin, S.; Page, L. The anatomy of a large-scale hypertextual Web search engine. Comput. Netw. ISDN Syst. 1998, 30, 107–117. [Google Scholar] [CrossRef]
- Girard, O.; Brocherie, F.; Morin, J.B.; Millet, G.P.; Hansen, C. Running mechanics and leg muscle activity patterns during early and late acceleration phases of repeated treadmill sprints in male recreational athletes. Eur. J. Appl. Physiol. 2020, 120, 2785–2796. [Google Scholar] [CrossRef]
- Brooks, G.A. The science and translation of lactate shuttle theory. Cell Metab. 2018, 27, 757–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fennell, C.R.J.; Hopker, J.G. The acute physiological and perceptual effects of individualizing the recovery interval duration based upon the resolution of muscle oxygen consumption during cycling exercise. Int. J. Sports Physiol. Perform. 2021, 12, 1580–1588. [Google Scholar] [CrossRef] [PubMed]
- Kriel, Y.; Askew, C.D.; Solomon, C. Sprint interval exercise versus continuous moderate intensity exercise: Acute effects on tissue oxygenation, blood pressure and enjoyment in 18–30 year old inactive men. PeerJ 2019, 7, e7077. Available online: https://peerj.com/articles/7077/ (accessed on 7 June 2019). [CrossRef] [PubMed]
- Kriel, Y.; Kerherve, H.A.; Askew, C.D.; Solomon, C. The effect of active versus passive recovery periods during high intensity intermittent exercise on local tissue oxygenation in 18–30 year old sedentary men. PLoS ONE 2016, 11, e0163733. Available online: https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0163733 (accessed on 27 September 2016). [CrossRef] [PubMed]
- Yogev, A.; Arnold, J.; Clarke, D.; Guenette, J.A.; Sporer, B.C.; Koehle, M.S. Comparing the respiratory compensation point with muscle oxygen saturation in locomotor and non-locomotor muscles using wearable NIRS spectroscopy during whole-body exercise. Front. Physiol. 2022, 13, 818733. [Google Scholar] [CrossRef]
- Børsheim, E.; Bahr, R. Effect of exercise intensity, duration and mode on post-exercise oxygen consumption. Sports Med. 2003, 33, 1037–1060. [Google Scholar] [CrossRef]
- Bastian, M.; Heymann, S.; Mathieu Jacomy, M. Gephi: An open source software for exploring and manipulating networks. In Proceedings of the Third International Conference on Weblogs and Social Media, ICWSM 2009, San Jose, CA, USA, 17–20 May 2009. [Google Scholar]
- Newman, M.E.J. Networks: An Introduction; Oxford University Press: Oxford, UK, 2020. [Google Scholar]
- Perra, N.; Fortunato, S. Spectral centrality measures in complex networks. Phys. Rev. 2008, 78, 036107. [Google Scholar] [CrossRef] [Green Version]
- Strang, G. Introduction to Linear Algebra, 4th ed.; Wellesley-Cambridge Press: Wellesley, MA, USA, 2009. [Google Scholar]
- Van Steen, M. Graph Theory and Complex Networks: An Introduction; Maarten van Steen: Enschede, The Netherlands, 2010. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AO30 | IMPA + AO30 | p | |
|---|---|---|---|
| Mechanical responses | |||
| pP (W) | 2296.7 ± 126.3 | 2501.1 ± 115.6 | 0.05 |
| mP (W) | 1706.9 ± 104.7 | 1894.0 ± 93.8 | 0.02 |
| minP (W) | 1210.3 ± 89.2 | 1372.3 ± 81.8 | 0.06 |
| pP (W·kg−1) | 31.4 ± 1.5 | 34.0 ± 1.4 | 0.04 |
| mP (W·kg−1) | 23.3 ± 1.1 | 25.8 ± 1.0 | 0.02 |
| minP (W·kg−1) | 16.5 ± 1.0 | 18.6 ± 07 | 0.06 |
| FI (%) | 47.2 ± 2.4 | 44.7 ± 2.6 | 0.32 |
| Physiological responses | |||
| at rest | |||
| LAC (mM) | 0.8 ± 0.1 | 0.8 ± 0.1 | 0.88 |
| HR (bpm) | 65 ± 3 | 66 ± 3 | 0.75 |
| SPO2 (%) | 97.4 ± 0.2 | 97.3 ± 0.2 | 0.50 |
| TSI_BB (%) | 66.1 ± 0.9 | 68.3 ± 0.7 | 0.07 |
| TSI_VL (%) | 73.7 ± 0.8 | 72.8 ± 0.8 | 0.27 |
| Physiological responses | |||
| after effort | |||
| LAC peak (mM) | 17.2 ± 0.6 | 16.2 ± 0.6 | 0.07 |
| time to pLAC | 8.1 ± 0.6 | 8.4 ± 0.6 | 0.76 |
| HR (bpm) | 176 ± 2 | 179 ± 2 | 0.11 |
| SPO2 (%) | 96.9 ± 0.3 | 96.9 ± 0.3 | 0.87 |
| TSI_BB (%) | 32.2 ± 1.8 | 34.1 ± 1.2 | 0.38 |
| TSI_VL (%) | 50.1 ± 1.7 | 49.8 ± 1.6 | 0.78 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manchado-Gobatto, F.B.; Torres, R.S.; Marostegan, A.B.; Rasteiro, F.M.; Hartz, C.S.; Moreno, M.A.; Pinto, A.S.; Gobatto, C.A. Complex Network Model Reveals the Impact of Inspiratory Muscle Pre-Activation on Interactions among Physiological Responses and Muscle Oxygenation during Running and Passive Recovery. Biology 2022, 11, 963. https://doi.org/10.3390/biology11070963
Manchado-Gobatto FB, Torres RS, Marostegan AB, Rasteiro FM, Hartz CS, Moreno MA, Pinto AS, Gobatto CA. Complex Network Model Reveals the Impact of Inspiratory Muscle Pre-Activation on Interactions among Physiological Responses and Muscle Oxygenation during Running and Passive Recovery. Biology. 2022; 11(7):963. https://doi.org/10.3390/biology11070963
Chicago/Turabian StyleManchado-Gobatto, Fúlvia Barros, Ricardo Silva Torres, Anita Brum Marostegan, Felipe Marroni Rasteiro, Charlini Simoni Hartz, Marlene Aparecida Moreno, Allan Silva Pinto, and Claudio Alexandre Gobatto. 2022. "Complex Network Model Reveals the Impact of Inspiratory Muscle Pre-Activation on Interactions among Physiological Responses and Muscle Oxygenation during Running and Passive Recovery" Biology 11, no. 7: 963. https://doi.org/10.3390/biology11070963
APA StyleManchado-Gobatto, F. B., Torres, R. S., Marostegan, A. B., Rasteiro, F. M., Hartz, C. S., Moreno, M. A., Pinto, A. S., & Gobatto, C. A. (2022). Complex Network Model Reveals the Impact of Inspiratory Muscle Pre-Activation on Interactions among Physiological Responses and Muscle Oxygenation during Running and Passive Recovery. Biology, 11(7), 963. https://doi.org/10.3390/biology11070963