
Methionine Supplementation during Pregnancy of Goats Improves Kids’ Birth Weight, Body Mass Index, and Postnatal Growth Pattern

 ,
,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
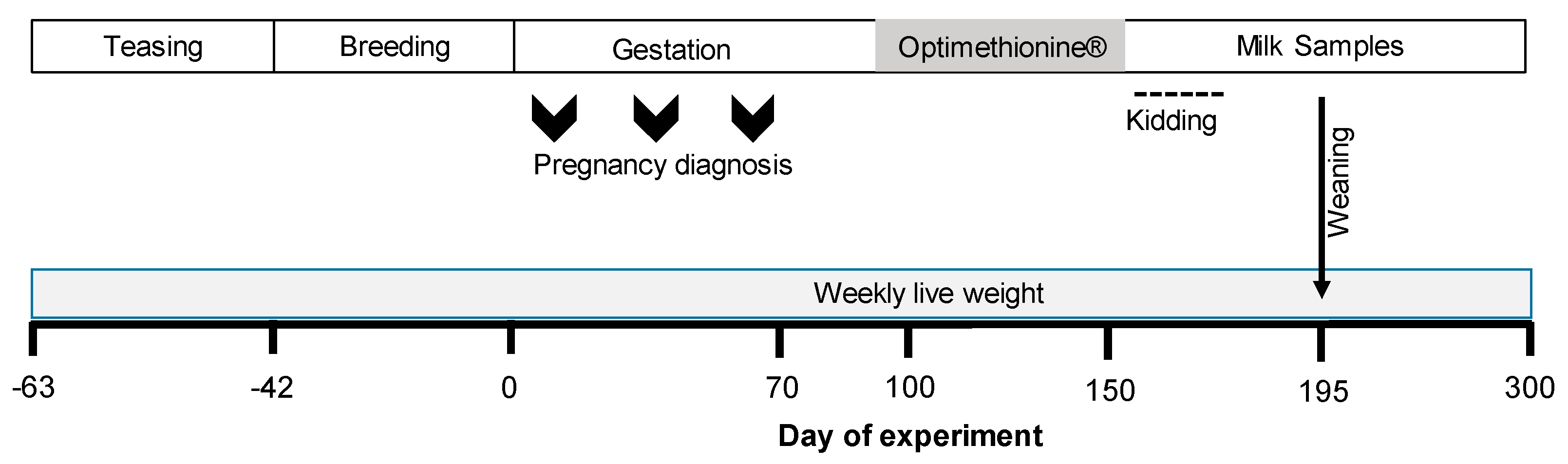
2.1. Study Site
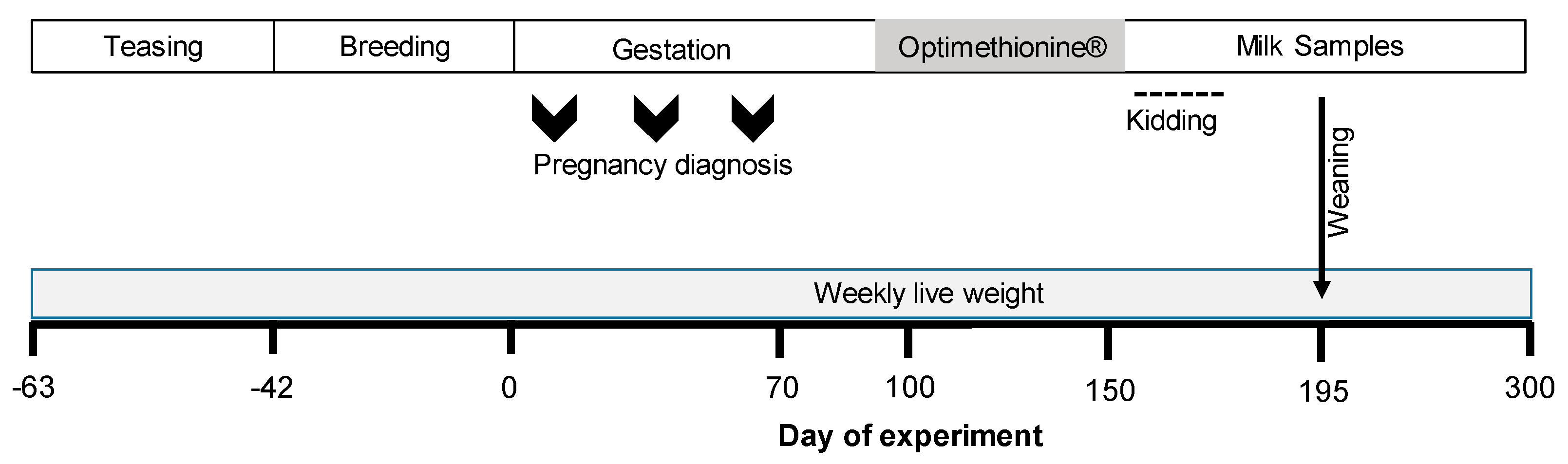
2.2. Experimental Design
2.3. Nutritional Diet and Herbal Methionine (Optimethionine®) Supplementation
2.4. Maternal Outcomes
2.5. Milk Yield and Composition
2.6. Newborn Outcomes
2.7. Data Analyses
3. Results
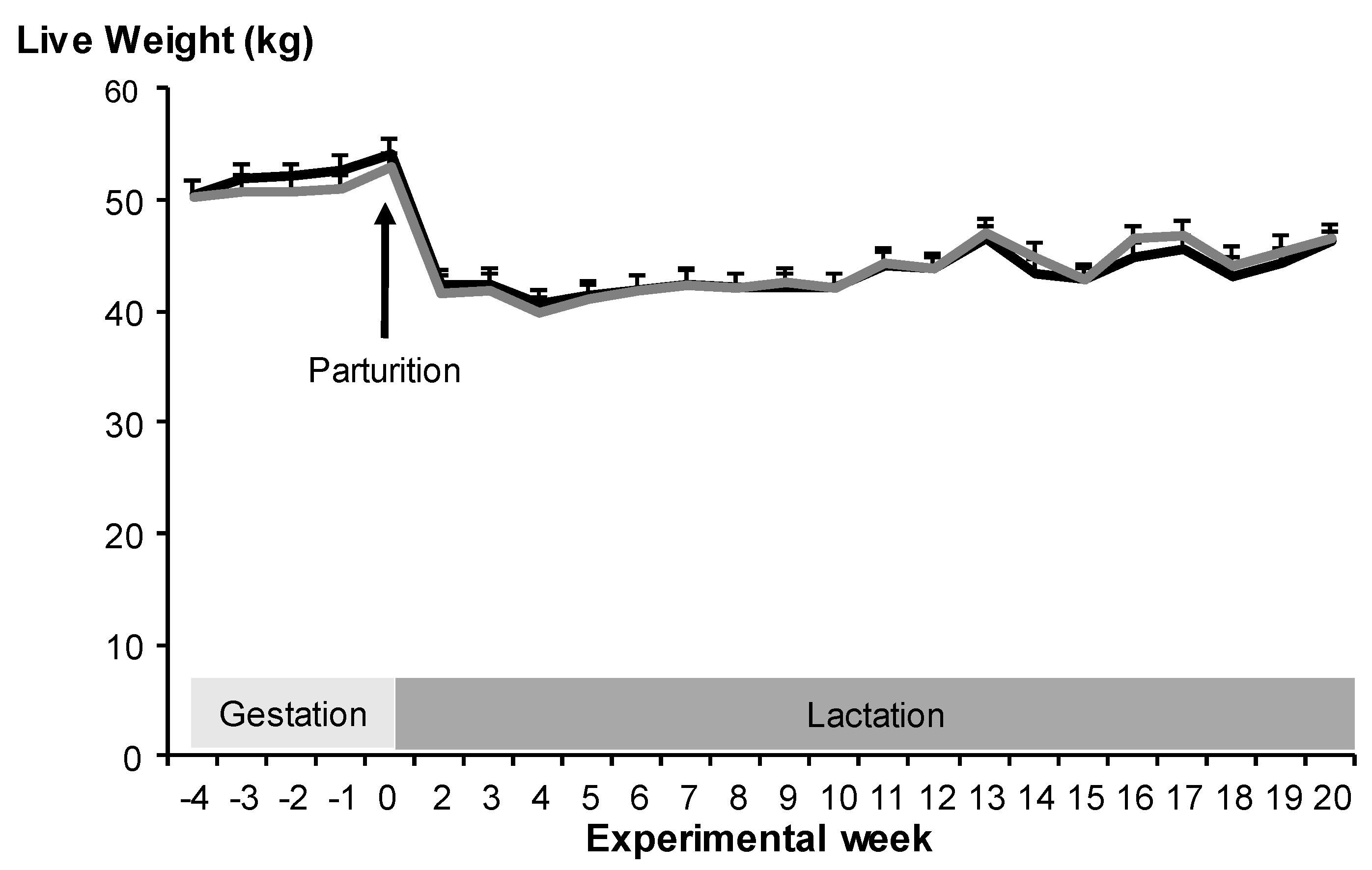
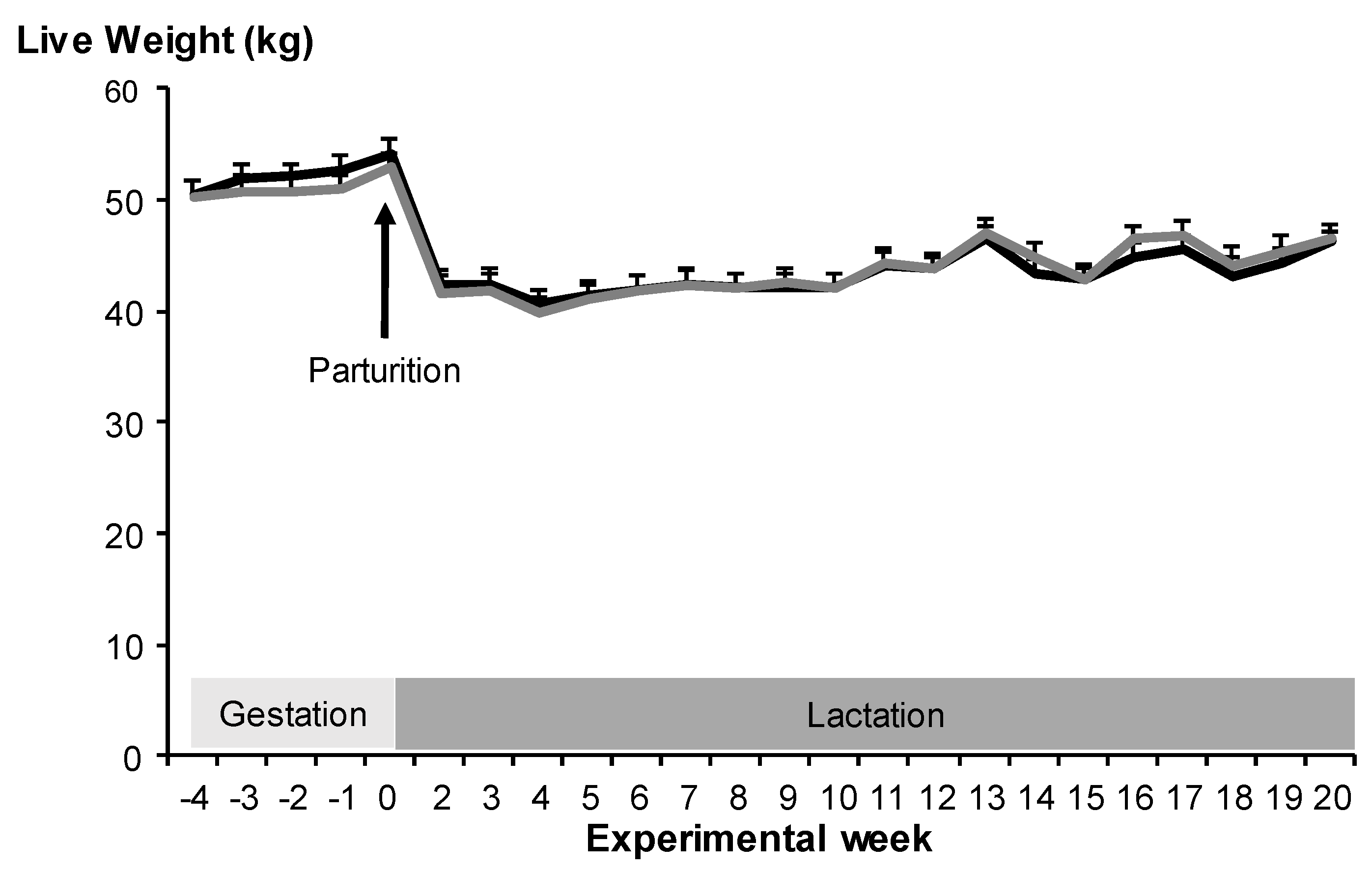
3.1. Maternal Live Weight Change
3.2. Milk Yield and Chemical Composition
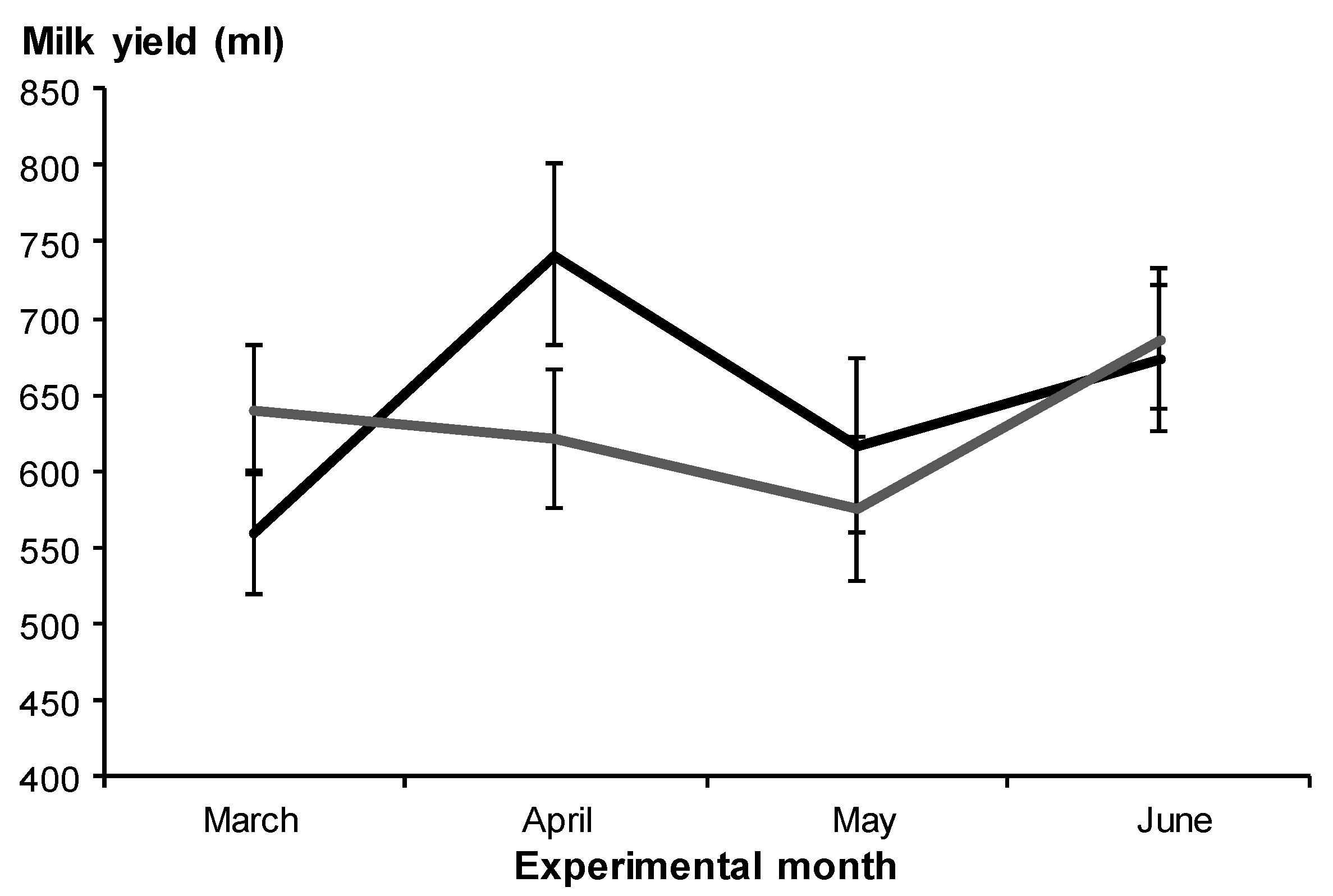
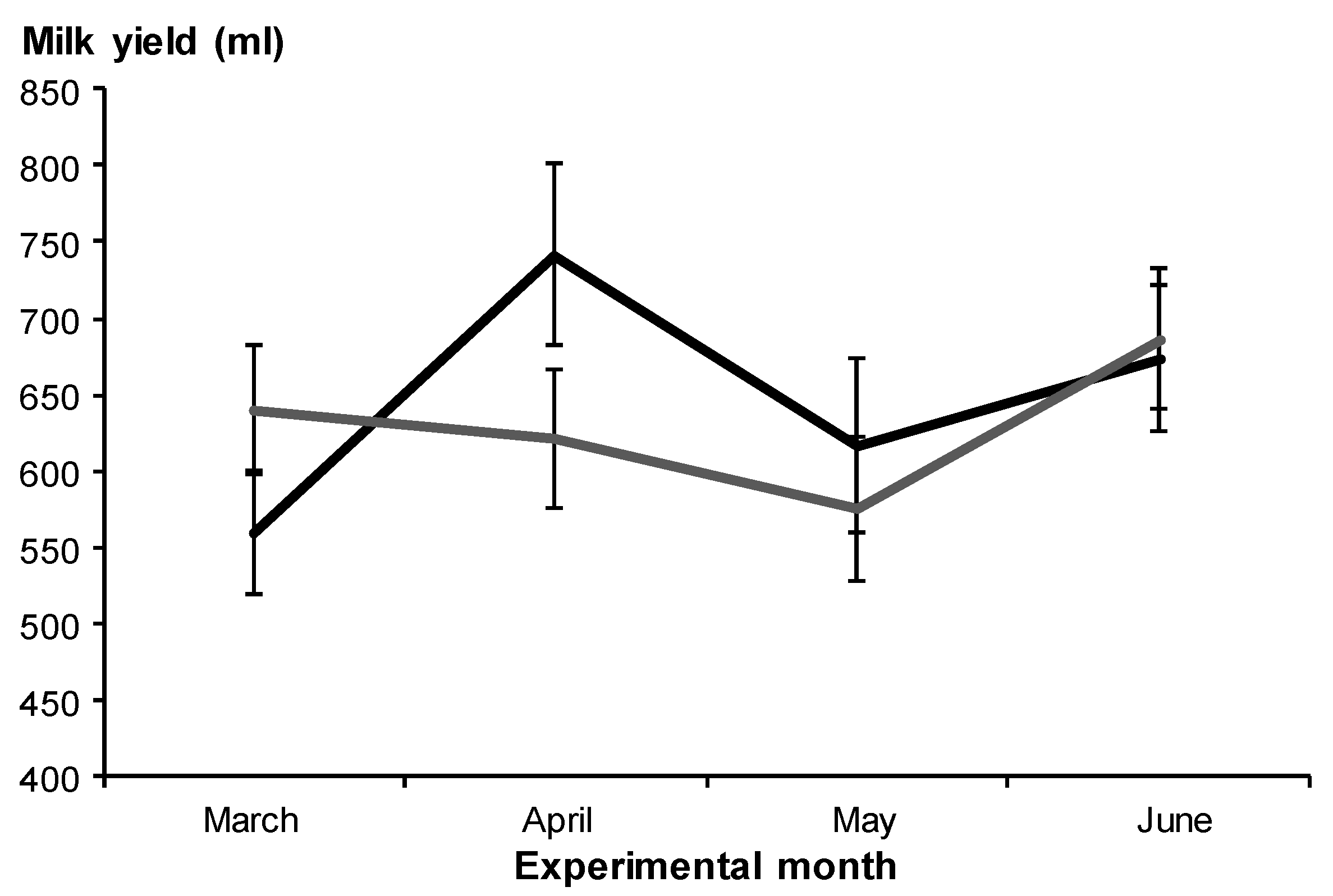
3.2.1. Milk Yield
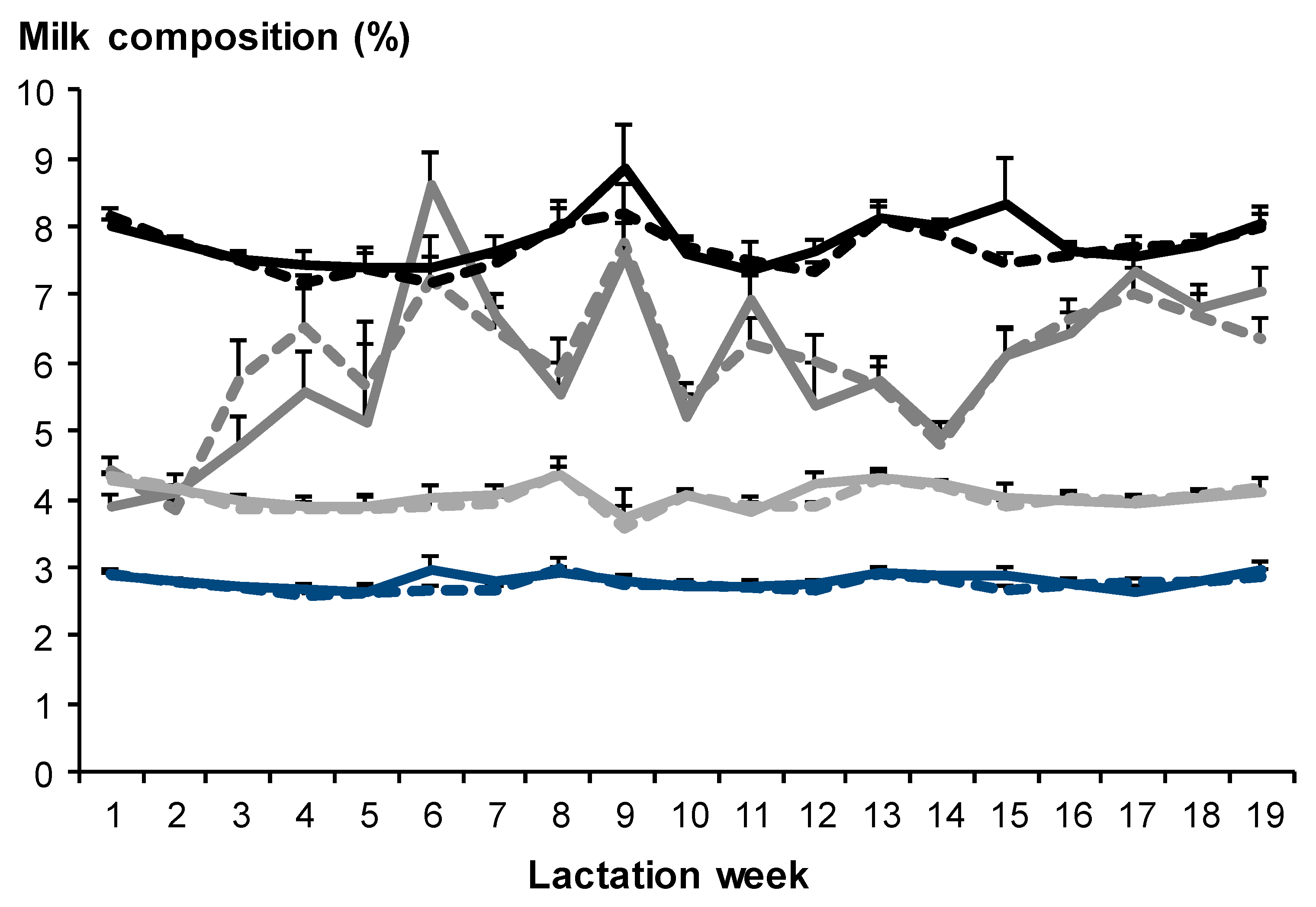
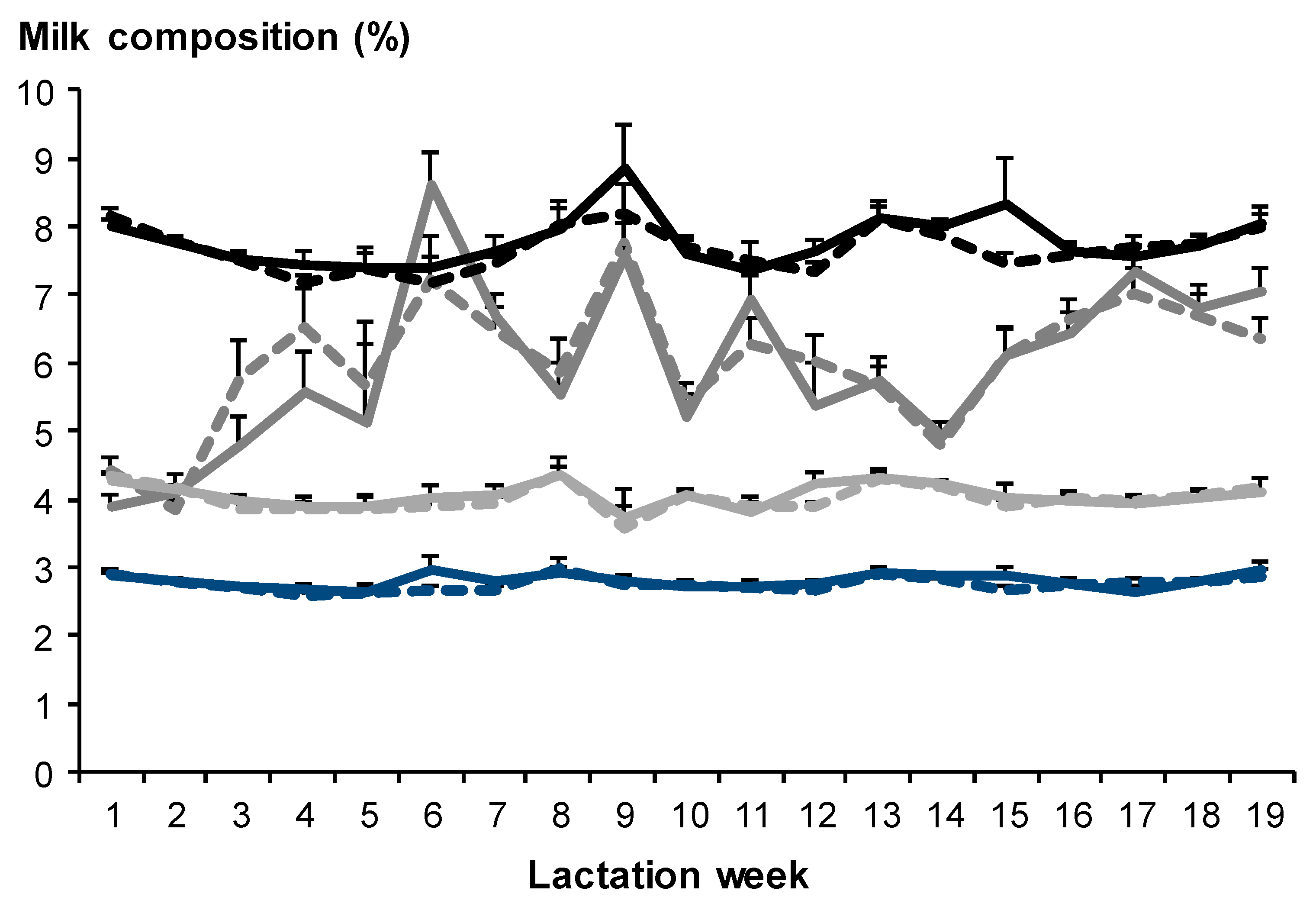
3.2.2. Milk Composition
3.3. Offspring Outcomes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Du, M.; Tong, J.; Zhao, J.; Underwood, K.R.; Zhu, M.; Ford, S.P.; Nathanielsz, P.W. Fetal programming of skeletal muscle development in ruminant animals. J. Anim. Sci. 2010, 88, E51–E60. [Google Scholar] [CrossRef] [Green Version]
- Ochoa Cordero, M.; Meza Herrera, C.A.; Vázquez García, J.M.; Stewart, C.A.; Rosales Nieto, C.A.; Ochoa Alfaro, A.E.; Purvis, I.A.; Cuevas Reyes, V.; Lee, H.; Martin, G.B. Pregnancy and litter size, but not lamb sex, affect feed intake and wool production by Merino-type ewes. Animals 2019, 9, 214. [Google Scholar] [CrossRef] [Green Version]
- Drackley, J.K. Biology of dairy cows during the transition period: The Final Frontier? J. Dairy Sci. 1999, 82, 2259–2273. [Google Scholar] [CrossRef]
- Bauman, D.E.; Bruce Currie, W. Partitioning of Nutrients During Pregnancy and Lactation: A review of mechanisms involving homeostasis and homeorhesis. J. Dairy Sci. 1980, 63, 1514–1529. [Google Scholar] [CrossRef]
- Godfrey, K.; Robinson, S.; Barker, D.; Osmond, C.; Cox, V. Maternal nutrition in early and late pregnancy in relation to placental and fetal growth. BMJ 1996, 312, 410. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Bazer, F.W.; Cudd, T.A.; Meininger, C.J.; Spencer, T.E. Maternal Nutrition and Fetal Development. J. Nutr. 2004, 134, 2169–2172. [Google Scholar] [CrossRef]
- He, Z.X.; Wu, D.Q.; Sun, Z.H.; Tan, Z.L.; Qiao, J.Y.; Ran, T.; Tang, S.X.; Zhou, C.S.; Han, X.F.; Wang, M.; et al. Protein or energy restriction during late gestation alters fetal growth and visceral organ mass: An evidence of intrauterine programming in goats. Anim. Rep. Sci. 2013, 137, 177–182. [Google Scholar] [CrossRef]
- Heasman, L.; Brameld, J.; Mostyn, A.; Budge, H.; Dawson, J.; Buttery, P.; Stephenson, T.; Symonds, M.E. Maternal nutrient restriction during early to mid gestation alters the relationship between insulin-like growth factor I and bodyweight at term in fetal sheep. Reprod. Fertil. Dev. 2000, 12, 345–350. [Google Scholar] [CrossRef]
- Thorn, S.R.; Regnault, T.R.H.; Brown, L.D.; Rozance, P.J.; Keng, J.; Roper, M.; Wilkening, R.B.; Hay, W.W., Jr.; Friedman, J.E. Intrauterine growth restriction increases fetal hepatic gluconeogenic capacity and reduces messenger ribonucleic acid translation initiation and nutrient sensing in fetal liver and skeletal muscle. Endocrinology 2009, 150, 3021–3030. [Google Scholar] [CrossRef]
- Pillai, S.M.; Jones, A.K.; Hoffman, M.L.; McFadden, K.K.; Reed, S.A.; Zinn, S.A.; Govoni, K.E. Fetal and organ development at gestational days 45, 90, 135 and at birth of lambs exposed to under- or over-nutrition during gestation. Transl. Anim. Sci. 2017, 1, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Vázquez-García, J.M.; Álvarez-Fuentes, G.; Orozco-Gregorio, H.O.; García-López, J.C.; González-Hernández, M.; Rosales-Nieto, C.A. Energy supplementation during the last third of gestation improves mother–young bonding in goats. Animals 2021, 11, 287. [Google Scholar] [CrossRef]
- Chadio, S.; Katsafadou, A.; Kotsampasi, B.; Michailidis, G.; Mountzouris, K.C.; Kalogiannis, D.; Christodoulou, V. Effects of maternal undernutrition during late gestation and/or lactation on colostrum synthesis and immunological parameters in the offspring. Reprod. Fertil. Dev. 2016, 28, 384–393. [Google Scholar] [CrossRef]
- Seyed Almoosavi, S.M.M.; Ghoorchi, T.; Naserian, A.A.; Khanaki, H.; Drackley, J.K.; Ghaffari, M.H. Effects of late-gestation heat stress independent of reduced feed intake on colostrum, metabolism at calving, and milk yield in early lactation of dairy cows. J. Dairy Sci. 2021, 104, 1744–1758. [Google Scholar] [CrossRef]
- Ramírez-Bribiesca, J.E.; Tórtora, J.L.; Hernández, L.M.; Huerta, M. Main causes of mortalities in dairy goat kids from the Mexican plateau. Small Rum. Res. 2001, 41, 77–80. [Google Scholar] [CrossRef]
- Hinch, G.N.; Brien, F. Lamb survival in Australian flocks: A review. Anim. Prod. Sci. 2014, 54, 656–666. [Google Scholar]
- Rosales Nieto, C.A.; Ferguson, M.; MacLeay, C.; Briegel, J.; Wood, D.; Martin, G.; Thompson, A. Ewe lambs with higher breeding values for growth achieve higher reproductive performance when mated at age 8 months. Theriogenology 2013, 80, 427–435. [Google Scholar] [CrossRef]
- Mossa, F.; Walsh, S.W.; Ireland, J.J.; Evans, A.C.O. Early nutritional programming and progeny performance: Is reproductive success already set at birth? Anim. Front. 2015, 5, 18–24. [Google Scholar] [CrossRef] [Green Version]
- Mellado, M. Dietary selection by goats and the implications for range management in the Chihuahuan Desert: A review. Rangel. J. 2016, 38, 331. [Google Scholar] [CrossRef]
- García-Monjaras, S.; Santos-Díaz, R.; Flores-Najera, M.; Cuevas-Reyes, V.; Meza-Herrera, C.; Mellado, M.; Chay-Canul, A.; Rosales-Nieto, C. Diet selected by goats on xerophytic shrubland with different milk yield potential. J. Arid Environ. 2021, 186, 104429. [Google Scholar] [CrossRef]
- Rosales-Nieto, C.A.; Gamez-Vazquez, H.G.; Gudino-Reyes, J.; Reyes-Ramirez, E.A.; Eaton, M.; Stanko, R.L.; Meza-Herrera, C.A.; Gonzalez-Bulnes, A. Nutritional and metabolic modulation of the male effect on the resumption of ovulatory activity in goats. Anim. Prod. Sci. 2011, 51, 115–122. [Google Scholar] [CrossRef]
- Urrutia Morales, J.; Rosales Nieto, C.A.; Vera Ávila, H.R.; Villagomez Amezcua Manjarres, E. Resumption of ovarian activity is modified by non-photoperiodic environmental cues in Criollo goats in tropical latitudes. Small Rum. Res. 2016, 137, 9–16. [Google Scholar] [CrossRef]
- Mellado, M.; Rodríguez, I.J.; Alvarado-Espino, A.; Véliz, F.G.; Mellado, J.; García, J.E. Short communication: Reproductive response to concentrate supplementation of mixed-breed goats on rangeland. Trop. Anim. Health Prod. 2020, 52, 2737–2741. [Google Scholar] [CrossRef]
- Gámez-Vázquez, H.G.; Rosales-Nieto, C.A.; Urrutia-Morales, J.; Mellado, M.; Meza-Herrera, C.A.; Vázquez-García, J.M.; Hernández-Arteaga, L.E.S.; Negrete-Sánchez, L.O.; Loredo-Osti, C.; Rivas-Jacobo, M.A.; et al. Effect of replacing sorghum stubble with Tillandsia Recurvata (L.) on liveweight change, blood metabolites, and hematic biometry of goats. Biology 2022, 11, 517. [Google Scholar] [CrossRef]
- Cuevas Reyes, V.; Santiago Hernandez, F.; de Jesus, M.F.N.; Vazquez Garcia, J.M.; Urrutia Morales, J.; Hosseini-Ghaffari, M.; Chay-Canul, A.; Meza-Herrera, C.A.; Gonzalez-Bulnes, A.; Martin, G.B.; et al. Intake of spineless cladodes of Opuntia ficus-indica during late pregnancy improves progeny performance in underfed sheep. Animals 2020, 10, 995. [Google Scholar] [CrossRef]
- Rosales-Nieto, C.A.; Rodríguez-Aguilar, M.; Santiago-Hernandez, F.; Cuevas-Reyes, V.; Flores-Najera, M.J.; Vázquez-García, J.M.; Urrutia-Morales, J.; Ghaffari, M.H.; Meza-Herrera, C.A.; González-Bulnes, A.; et al. Periconceptional nutrition with;spineless cactus (Opuntia ficus-indica) improves metabolomic profiles and pregnancy outcomes in sheep. Sci. Rep. 2021, 11, 7214. [Google Scholar] [CrossRef]
- Brosnan, J.T.; Brosnan, M.E. The Sulfur-Containing Amino Acids: An Overview. J. Nutr. 2006, 136, 1636S–1640S. [Google Scholar] [CrossRef] [Green Version]
- Rezaei, R.; Wu, Z.; Hou, Y.; Bazer, F.W.; Wu, G. Amino acids and mammary gland development: Nutritional implications for milk production and neonatal growth. J. Anim. Sci. Biotechnol. 2016, 7, 20. [Google Scholar] [CrossRef] [Green Version]
- Lobley, G.E. Control of the metabolic fate of amino acids in ruminants: A review. J. Anim. Sci. 1992, 70, 3264–3275. [Google Scholar] [CrossRef]
- Monsen, A.-L.B.; Schneede, J.; Ueland, P.M. Mid-trimester amniotic fluid methionine concentrations: A predictor of birth weight and length. Metabolism 2006, 55, 1186–1191. [Google Scholar] [CrossRef]
- Ji, Y.; Wu, Z.; Dai, Z.; Sun, K.; Wang, J.; Wu, G. Nutritional epigenetics with a focus on amino acids: Implications for the development and treatment of metabolic syndrome. J. Nutr. Biochem. 2016, 27, 1–8. [Google Scholar] [CrossRef]
- Caso, G.; Garlick, P.J. Control of muscle protein kinetics by acid-base balance. Curr. Opin. Clin. Nutr. Metab. Care 2005, 8, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Oster, M.; Nuchchanart, W.; Trakooljul, N.; Muráni, E.; Zeyner, A.; Wirthgen, E.; Hoeflich, A.; Ponsuksili, S.; Wimmers, K. Methylating micronutrient supplementation during pregnancy influences foetal hepatic gene expression and IGF signalling and increases foetal weight. Eur. J. Nutr. 2016, 55, 1717–1727. [Google Scholar] [CrossRef] [PubMed]
- Jacometo, C.B.; Zhou, Z.; Luchini, D.; Corrêa, M.N.; Loor, J.J. Maternal supplementation with rumen-protected methionine increases prepartal plasma methionine concentration and alters hepatic mRNA abundance of 1-carbon, methionine, and transsulfuration pathways in neonatal Holstein calves. J. Dairy Sci. 2017, 100, 3209–3219. [Google Scholar] [CrossRef] [PubMed]
- Mousaie, A.; Valizadeh, R.; Chamsaz, M. Selenium-methionine and chromium-methionine supplementation of sheep around parturition: Impacts on dam and offspring performance. Arch. Anim. Nutr. 2017, 71, 134–149. [Google Scholar] [CrossRef] [PubMed]
- Alharthi, A.S.; Batistel, F.; Abdelmegeid, M.K.; Lascano, G.; Parys, C.; Helmbrecht, A.; Trevisi, E.; Loor, J.J. Maternal supply of methionine during late-pregnancy enhances rate of Holstein calf development in utero and postnatal growth to a greater extent than colostrum source. J. Anim. Sci. Biotechnol. 2018, 9, 83. [Google Scholar] [CrossRef]
- Alharthi, A.S.; Lopreiato, V.; Dai, H.; Bucktrout, R.; Abdelmegeid, M.; Batistel, F.; Parys, C.; Shen, X.; Ballou, M.A.; Trevisi, E.; et al. Short communication: Supply of methionine during late pregnancy enhances whole-blood innate immune response of Holstein calves partly through changes in mRNA abundance in polymorphonuclear leukocytes. J. Dairy Sci. 2019, 102, 10599–10605. [Google Scholar] [CrossRef] [Green Version]
- Osorio, J.S.; Ji, P.; Drackley, J.K.; Luchini, D.; Loor, J.J. Smartamine M and MetaSmart supplementation during the peripartal period alter hepatic expression of gene networks in 1-carbon metabolism, inflammation, oxidative stress, and the growth hormone–insulin-like growth factor 1 axis pathways. J. Dairy Sci. 2014, 97, 7451–7464. [Google Scholar] [CrossRef]
- Giallongo, F.; Harper, M.T.; Oh, J.; Lopes, J.C.; Lapierre, H.; Patton, R.A.; Parys, C.; Shinzato, I.; Hristov, A.N. Effects of rumen-protected methionine, lysine, and histidine on lactation performance of dairy cows. J. Dairy Sci. 2016, 99, 4437–4452. [Google Scholar] [CrossRef] [Green Version]
- Nuproxa Switzerland Ltd. Available online: https://nuproxa.ch/products/optimethione/ (accessed on 31 January 2022).
- Martin, G.B.; Milton, J.T.B.; Davidson, R.H.; Banchero Hunzicker, G.E.; Lindsay, D.R.; Blache, D. Natural methods for increasing reproductive efficiency in small ruminants. Anim. Rep. Sci. 2004, 82–83, 231–245. [Google Scholar] [CrossRef]
- FASS. Guide for the Care and Use of Agricultural Animals in Agricultural Research and Teaching, 3rd ed.; Federation Animal Science Society: Champaign, IL, USA, 2010; p. 177. [Google Scholar]
- NAM-National Academy of Medicine. Guide for the Care and Use of Laboratory Animals, 1st ed.; Co-Produced by the National Academy of Medicine—Mexico and the Association for Assessment and Accreditation of Laboratory Animal Care International; Harlan: Mexico City, Mexico, 2010. [Google Scholar]
- Medina García, G.; Díaz Padilla, G.; Osti, C.L.; Altamirano, V.S.; García, M.A.C. Estadísticas Climatológicas Básicas del Estado de San Luis Potosí (Período 1961–2001); Libro Técnico No 2; Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, Campo Experimental San Luis: San Luis Potosí, México, 2005; 319p. [Google Scholar]
- Rosales Nieto, C.A.; Mantey, A.; Makela, B.; Byrem, T.; Ehrhardt, R.; Veiga-Lopez, A. Shearing during late pregnancy increases size at birth but does not alter placental endocrine responses in sheep. Animal 2020, 14, 799–806. [Google Scholar] [CrossRef]
- N.R.C. Nutrient Requirements of Small Ruminants: Sheep, Goats, Cervids and New World Camelids; National Academies Press: Washington, DC, USA, 2007; 292p. [Google Scholar]
- Bárcena-Gama, J.R.; Martínez-Aispuro, J.A.; Mendoza-Martínez, G.D.; Cordero-Mora, J.L.; Sánchez-Torres, M.T.; Figueroa-Velasco, J.L.; Ayala-Monter, M.A. Evaluation of polyherbal methionine and choline in feedlot rations for lambs. S. Afr. J. Anim. Sci. 2020, 50, 731–737. [Google Scholar] [CrossRef]
- Rosales Nieto, C.A.; Meza-Herrera, C.A.; Morón Cedillo, F.J.; Flores Najera, M.J.; Gámez Vázquez, H.G.; Cuevas Reyes, V.; Liu, S.M. Effects of vitamin E supply during late gestation and early lactation upon colostrum composition, milk production and quality in nutritional restricted ewes. Small Rum. Res. 2015, 133, 77–81. [Google Scholar] [CrossRef]
- Tanaka, T.; Akaboshi, N.; Inoue, Y.; Kamomae, H.; Kaneda, Y. Fasting-induced suppression of pulsatile luteinizing hormone secretion is related to body energy status in ovariectomized goats. Anim. Rep. Sci. 2002, 72, 185–196. [Google Scholar] [CrossRef]
- Rosales-Nieto, C.A.; Ehrhardt, R.; Mantey, A.; Makela, B.; Byrem, T.; Veiga-Lopez, A. Preconceptional diet manipulation and fetus number can influence placenta endocrine function in sheep. Dom. Anim. Endocrinol. 2021, 74, 106577. [Google Scholar] [CrossRef]
- SAS Institute. SAS/Stat User’s Guide; Version 9.3; SAS Institute Inc.: Cary, NC, USA, 2010. [Google Scholar]
- Silanikove, N. The physiological basis of adaptation in goats to harsh environments. Small Rum. Res. 2000, 35, 181–193. [Google Scholar] [CrossRef]
- Flores-Najera, M.J.; Cuevas-Reyes, V.; Vázquez-García, J.M.; Beltrán-López, S.; Meza-Herrera, C.A.; Mellado, M.; Negrete-Sánchez, L.O.; Rivas-Jacobo, M.A.; Rosales-Nieto, C.A. Milk yield and composition of mixed-breed goats on rangeland during the dry season and the effect on the growth of their progeny. Biology 2021, 10, 220. [Google Scholar] [CrossRef]
- Mellado, M.; Véliz, F.G.; Macías-Cruz, U.; Avendaño-Reyes, L.; García, J.E.; Rosales-Nieto, C.A. Effect of breed and management practices on reproductive and milking performance of rangeland goats. Trop. Anim. Health Prod. 2022, 54, 193. [Google Scholar] [CrossRef]
- Lu, L.-M.; Li, Q.-Z.; Huang, J.-G.; Gao, X.-J. Proteomic and Functional Analyses Reveal MAPK1 Regulates Milk Protein Synthesis. Molecules 2013, 18, 263–275. [Google Scholar] [CrossRef]
- Hu, L.; Chen, Y.; Cortes, I.M.; Coleman, D.N.; Dai, H.; Liang, Y.; Parys, C.; Fernandez, C.; Wang, M.; Loor, J.J. Supply of methionine and arginine alters phosphorylation of mechanistic target of rapamycin (mTOR), circadian clock proteins, and α-s1-casein abundance in bovine mammary epithelial cells. Food Funct. 2020, 11, 883–894. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Lei, J.; Hancock, S.; Scanlan, V.; Broomfield, S.; Currie, A.; Thompson, A. Lamb survival, glutathione redox state and immune function of neonates and lambs from periparturient Merino ewes supplemented with rumen-protected methionine. Arch. Anim. Nutr. 2016, 70, 389–401. [Google Scholar] [CrossRef]
- Williams, A.; Murison, R.; Padgett, J. Metabolism of Sulfur-containing Amino Acids by Pregnant Merino Ewes. Aust. J. Biol. Sci. 1988, 41, 247–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, C.; Masters, D.; Williams, I.; Connell, P. Changes in plasma amino acid patterns and wool growth in response to abomasal injections of amino acids during late pregnancy and early lactation. J. Agric. Res. 1993, 44, 959–971. [Google Scholar] [CrossRef]
- Lan, X.; Cretney, E.; Kropp, J.; Khateeb, K.; Berg, M.; Peñagaricano, F.; Magness, R.; Radunz, A.; Khatib, H. Maternal diet during pregnancy induces gene expression and DNA methylation changes in fetal tissues in sheep. Front. Genet. 2013, 4, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larson, D.M.; Martin, J.L.; Adams, D.C.; Funston, R.N. Winter grazing system and supplementation during late gestation influence performance of beef cows and steer progeny. J. Anim. Sci. 2009, 87, 1147–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montaldo, H.H.; Torres-Hernández, G.; Valencia-Posadas, M. Goat breeding research in Mexico. Small Rum. Res. 2010, 89, 155–163. [Google Scholar] [CrossRef]
- Flores-Najera, M.J.; Vélez-Monroy, L.I.; Sánchez-Duarte, J.I.; Cuevas-Reyes, V.; Mellado, M.; Rosales-Nieto, C.A. Milk yield and composition and body weight of offsprings of mixed-breed goats on semiarid rangelands with different rainfall. Trop. Anim. Health Prod. 2020, 52, 3799–3808. [Google Scholar] [CrossRef]
- Rosales Nieto, C.A.; Meza-Herrera, C.A.; Moron Cedillo, F.d.J.; Flores Najera, M.d.J.; Gámez Vázquez, H.G.; Ventura Pérez, F.d.J.; Liu, S. Vitamin E supplementation of undernourished ewes pre- and post-lambing reduces weight loss of ewes and increases weight of lambs. Trop. Anim. Health Prod. 2016, 48, 613–618. [Google Scholar] [CrossRef]
- Rosales Nieto, C.A.; Ferguson, M.B.; Macleay, C.A.; Briegel, J.R.; Wood, D.A.; Martin, G.B.; Bencini, R.; Thompson, A.N. Milk production and composition, and progeny performance in young ewes with high merit for rapid growth and muscle and fat accumulation. Animal 2018, 12, 2292–2299. [Google Scholar] [CrossRef]
- Jacometo, C.B.; Alharthi, A.S.; Zhou, Z.; Luchini, D.; Loor, J.J. Maternal supply of methionine during late pregnancy is associated with changes in immune function and abundance of microRNA and mRNA in Holstein calf polymorphonuclear leukocytes. J. Dairy Sci. 2018, 101, 8146–8158. [Google Scholar] [CrossRef]
- Tripodi, F.; Castoldi, A.; Nicastro, R.; Reghellin, V.; Lombardi, L.; Airoldi, C.; Falletta, E.; Maffioli, E.; Scarcia, P.; Palmieri, L.; et al. Methionine supplementation stimulates mitochondrial respiration. Biochim. Biophys. Acta. Mol. Cell Res. 2018, 1865, 1901–1913. [Google Scholar]
- Liu, H.; Yang, G.; Degen, A.; Ji, K.; Jiao, D.; Liang, Y.; Xiao, L.; Long, R.; Zhou, J. Effect of feed level and supplementary rumen protected lysine and methionine on growth performance, rumen fermentation, blood metabolites and nitrogen balance in growing Tan lambs fed low protein diets. Anim. Feed Sci. Technol. 2021, 279, 115024. [Google Scholar] [CrossRef]
- Gardner, D.S.; Buttery, P.J.; Daniel, Z.; Symonds, M.E. Factors affecting birth weight in sheep: Maternal environment. Reproduction 2007, 133, 297–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatford, K.; Quinn, K.; Walton, P.; Grant, P.; Hosking, B.; Egan, A.; Owens, P. Ontogenic and nutritional changes in circulating insulin-like growth factor (IGF)-I, IGF-II and IGF-binding proteins in growing ewe and ram lambs. J. Endocrinol. 1997, 155, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Clifton, V.L. Review: Sex and the Human Placenta: Mediating Differential Strategies of Fetal Growth and Survival. Placenta 2010, 31, S33–S39. [Google Scholar] [CrossRef] [PubMed]
- Bencini, R.; Pulina, G. The quality of sheep milk: A review. Aust. J. Exp. Agric. 1997, 37, 485–504. [Google Scholar] [CrossRef]
- Flores, A.; Mendoza, G.; Pinos-Rodriguez, J.M.; Plata, F.; Vega, S.; Bárcena, R. Effects of rumen-protected methionine on milk production of dairy goats. Italian J. Anim. Sci. 2009, 8, 271–275. [Google Scholar] [CrossRef]
- McCarthy, R.D.; Porter, G.A.; Griel, L.C. Bovine Ketosis and Depressed Fat Test in Milk: A Problem of Methionine Metabolism and Serum Lipoprotein Aberration1. J. Dairy Sci. 1968, 51, 459–462. [Google Scholar] [CrossRef]
- Samuelson, D.J.; Denise, S.K.; Roffler, R.; Ax, R.L.; Armstrong, D.V.; Romagnolo, D.F. Response of Holstein and Brown Swiss Cows Fed Alfalfa Hay-Based Diets to Supplemental Methionine at Two Stages of Lactation. J. Dairy Sci. 2001, 84, 917–928. [Google Scholar] [CrossRef]
- Zamuner, F.; DiGiacomo, K.; Cameron, A.W.N.; Leury, B.J. Endocrine and metabolic status of commercial dairy goats during the transition period. J. Dairy Sci. 2020, 103, 5616–5628. [Google Scholar] [CrossRef]
- Van der Drift, S.G.A.; Houweling, M.; Schonewille, J.T.; Tielens, A.G.M.; Jorritsma, R. Protein and fat mobilization and associations with serum β-hydroxybutyrate concentrations in dairy cows. J. Dairy Sci. 2012, 95, 4911–4920. [Google Scholar] [CrossRef]
- Patton, R.A.; Hristov, A.N.; Lapierre, H. Protein Feeding and Balancing for Amino Acids in Lactating Dairy Cattle. Vet. Clin. Food Anim. Pract. 2014, 30, 599–621. [Google Scholar] [CrossRef] [PubMed]
- Patton, R.A. Effect of rumen-protected methionine on feed intake, milk production, true milk protein concentration, and true milk protein yield, and the factors that influence these effects: A meta-analysis. J. Dairy Sci. 2010, 93, 2105–2118. [Google Scholar] [CrossRef]
- Lundquist, R.G.; Otterby, D.E.; Linn, J.G. Influence of three concentrations of dl-methionine or methionine hydroxy analog on milk yield and milk composition. J. Dairy Sci. 1985, 68, 3350–3354. [Google Scholar] [CrossRef]
- Chilliard, Y.; Doreau, M. Influence of supplementary fish oil and rumen-protected methionine on milk yield and composition in dairy cows. J. Dairy Res. 1997, 64, 173–179. [Google Scholar] [CrossRef]
- Chen, Z.H.; Broderick, G.A.; Luchini, N.D.; Sloan, B.K.; Devillard, E. Effect of feeding different sources of rumen-protected methionine on milk production and N-utilization in lactating dairy cows1. J. Dairy Sci. 2011, 94, 1978–1988. [Google Scholar] [CrossRef] [PubMed]
- Osorio, J.S.; Ji, P.; Drackley, J.K.; Luchini, D.; Loor, J.J. Supplemental Smartamine M or MetaSmart during the transition period benefits postpartal cow performance and blood neutrophil function. J. Dairy Sci. 2013, 96, 6248–6263. [Google Scholar] [CrossRef] [Green Version]
- Batistel, F.; Arroyo, J.M.; Bellingeri, A.; Wang, L.; Saremi, B.; Parys, C.; Trevisi, E.; Cardoso, F.C.; Loor, J.J. Ethyl-cellulose rumen-protected methionine enhances performance during the periparturient period and early lactation in Holstein dairy cows. J. Dairy Sci. 2017, 100, 7455–7467. [Google Scholar] [CrossRef] [Green Version]
- Junior, V.C.; Lopes, F.; Schwab, C.G.; Toledo, M.Z.; Collao-Saenz, E.A. Effects of rumen-protected methionine supplementation on the performance of high production dairy cows in the tropics. PLoS ONE 2021, 16, e0243953. [Google Scholar] [CrossRef]
- Titi, H.H. Effect of long-term rumen-protected methionine supplementation on performance of Shami goats and growth performance of their kids. Anim. Prod. Sci. 2017, 57, 1713–1718. [Google Scholar] [CrossRef]
- Papadomichelakis, G.; Koutsotolis, K.; Zabeli, G.; Zervas, G. The effect of lactating dairy ewes’ diet supplementation with ALIMET (liquid methionine) on milk yield and milk composition. Italian J. Anim. Sci. 2002, 1, 301–305. [Google Scholar] [CrossRef] [Green Version]
- Bagnicka, E.; Distl, O.; Hamann, H.; Lukaszewicz, M. Heritabilities and genetic correlations between the dairy traits in goats estimated in first vs. later lactations. Anim. Sci. Pap. Rep. 2004, 2, 205–213. [Google Scholar]
- Torres-Vázquez, J.A.; Valencia-Posadas, M.; Castillo-Juárez, H.; Montaldo, H.H. Genetic and phenotypic parameters of milk yield, milk composition and age at first kidding in Saanen goats from Mexico. Livest. Sci. 2009, 126, 147–153. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Diet | |||
|---|---|---|---|
| Ingredient Composition (% in Diet) | Chemical Composition | ||
| Alfalfa hay | 24.2 | Dry Matter (%) | 72.3 |
| Oats hay | 45.5 | Crude Protein (%) | 8.9 |
| Maize silage | 30.3 | Metabolizable Energy (Mcal/kg) | 2.2 |
| Calcium (%) | 0.35 | ||
| Phosphorus (%) | 0.23 | ||
| Variable | Treatment | p-Value | |
|---|---|---|---|
| T-CTL | T-MET | ||
| n | 30 | 30 | |
| Enrolment weight (kg) | 50.5 ± 1.1 | 50.3 ± 1.3 | 0.897 |
| Final weight (kg) | 54.2 ± 1.3 | 52.8 ± 0.1.4 | 0.472 |
| Live weight change (g/d) | 73 ± 10 | 51 ± 7 | 0.068 |
| Total Birth Weight (kg) | 5.4 ± 0.3 | 5.1 ± 0.3 | 0.434 |
| Fetal daily gain (g/d) | 108 ± 6 | 102 ± 5 | 0.434 |
| Total Maternal Gain (g/d) | −31 ± 10 | −47 ± 8 | 0.231 |
| Variable | Treatment | p-Value | Interactions | ||
|---|---|---|---|---|---|
| T-CTL | T-MET | TRT*BT | TRT*Sex | ||
| n | 43 | 48 | |||
| Birth weight (kg) | 3.1 ± 0.1 | 3.5 ± 0.1 | 0.001 | 0.199 | 0.913 |
| Daily live weight gain (g d−2) | 121 ± 6 | 142 ± 6 | 0.010 | 0.729 | 0.919 |
| Weaning weight (kg) | 8.3 ± 0.2 | 9.3 ± 0.3 | 0.004 | 0.979 | 0.593 |
| BMI at birth (units kg m−2) | 0.28 ± 0.01 | 0.30 ± 0.01 | 0.010 | 0.160 | 0.240 |
| BMI at wean (units kg m−2) | 0.85 ± 0.03 | 1.00 ± 0.06 | 0.030 | 0.862 | 0.868 |
| Sex | p-value | ||||
| Female | Male | ||||
| n | 46 | 45 | |||
| Birth weight (kg) | 3.1 ± 0.1 | 3.5 ± 0.1 | 0.004 | ||
| Daily live weight gain (g d−2) | 122 ± 6 | 144 ± 6 | 0.010 | ||
| Weaning weight (kg) | 8.5 ± 0.3 | 9.2 ± 0.3 | 0.040 | ||
| BMI at birth (units kg m−2) | 0.27 ± 0.01 | 0.31 ± 0.01 | 0.001 | ||
| BMI at wean (units kg m−2) | 0.90 ± 0.06 | 0.96 ± 0.04 | 0.420 | ||
| Birth type | |||||
| Singleton | Twins | ||||
| n | 25 | 66 | |||
| Birth weight (kg) | 3.7 ± 0.1 | 3.1 ± 0.1 | 0.001 | ||
| Daily live weight gain (g d−2) | 165 ± 9 | 120 ± 4 | 0.001 | ||
| Weaning weight (kg) | 10.3 ± 0.3 | 8.3 ± 0.2 | 0.001 | ||
| BMI at birth (units kg m−2) | 0.32 ± 0.01 | 0.28 ± 0.01 | 0.002 | ||
| BMI at wean (units kg m−2) | 1.14 ± 0.11 | 0.86 ± 0.03 | 0.001 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castillo-Gutierrez, D.; Hernández-Arteaga, L.E.S.; Flores-Najera, M.J.; Cuevas-Reyes, V.; Vázquez-García, J.M.; Loredo-Osti, C.; Beltrán-López, S.; Ballesteros-Rodea, G.; Gonzalez-Bulnes, A.; Meza-Herrera, C.A.; et al. Methionine Supplementation during Pregnancy of Goats Improves Kids’ Birth Weight, Body Mass Index, and Postnatal Growth Pattern. Biology 2022, 11, 1065. https://doi.org/10.3390/biology11071065
Castillo-Gutierrez D, Hernández-Arteaga LES, Flores-Najera MJ, Cuevas-Reyes V, Vázquez-García JM, Loredo-Osti C, Beltrán-López S, Ballesteros-Rodea G, Gonzalez-Bulnes A, Meza-Herrera CA, et al. Methionine Supplementation during Pregnancy of Goats Improves Kids’ Birth Weight, Body Mass Index, and Postnatal Growth Pattern. Biology. 2022; 11(7):1065. https://doi.org/10.3390/biology11071065
Chicago/Turabian StyleCastillo-Gutierrez, Diego, Luisa E. S. Hernández-Arteaga, Manuel J. Flores-Najera, Venancio Cuevas-Reyes, Juan M. Vázquez-García, Catarina Loredo-Osti, Sergio Beltrán-López, Gilberto Ballesteros-Rodea, Antonio Gonzalez-Bulnes, Cesar A. Meza-Herrera, and et al. 2022. "Methionine Supplementation during Pregnancy of Goats Improves Kids’ Birth Weight, Body Mass Index, and Postnatal Growth Pattern" Biology 11, no. 7: 1065. https://doi.org/10.3390/biology11071065
APA StyleCastillo-Gutierrez, D., Hernández-Arteaga, L. E. S., Flores-Najera, M. J., Cuevas-Reyes, V., Vázquez-García, J. M., Loredo-Osti, C., Beltrán-López, S., Ballesteros-Rodea, G., Gonzalez-Bulnes, A., Meza-Herrera, C. A., & Rosales-Nieto, C. A. (2022). Methionine Supplementation during Pregnancy of Goats Improves Kids’ Birth Weight, Body Mass Index, and Postnatal Growth Pattern. Biology, 11(7), 1065. https://doi.org/10.3390/biology11071065