
Isolation, Identification and Hyperparasitism of a Novel Cladosporium cladosporioides Isolate Hyperparasitic to Puccinia striiformis f. sp. tritici, the Wheat Stripe Rust Pathogen

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation of the Hyperparasite from Pst-Infected Leaves
2.2. Morphological Identification
2.3. Molecular Characterization
2.4. Phylogenetic Analysis
2.5. Pathogenicity and Hyperparasite Tests
3. Results
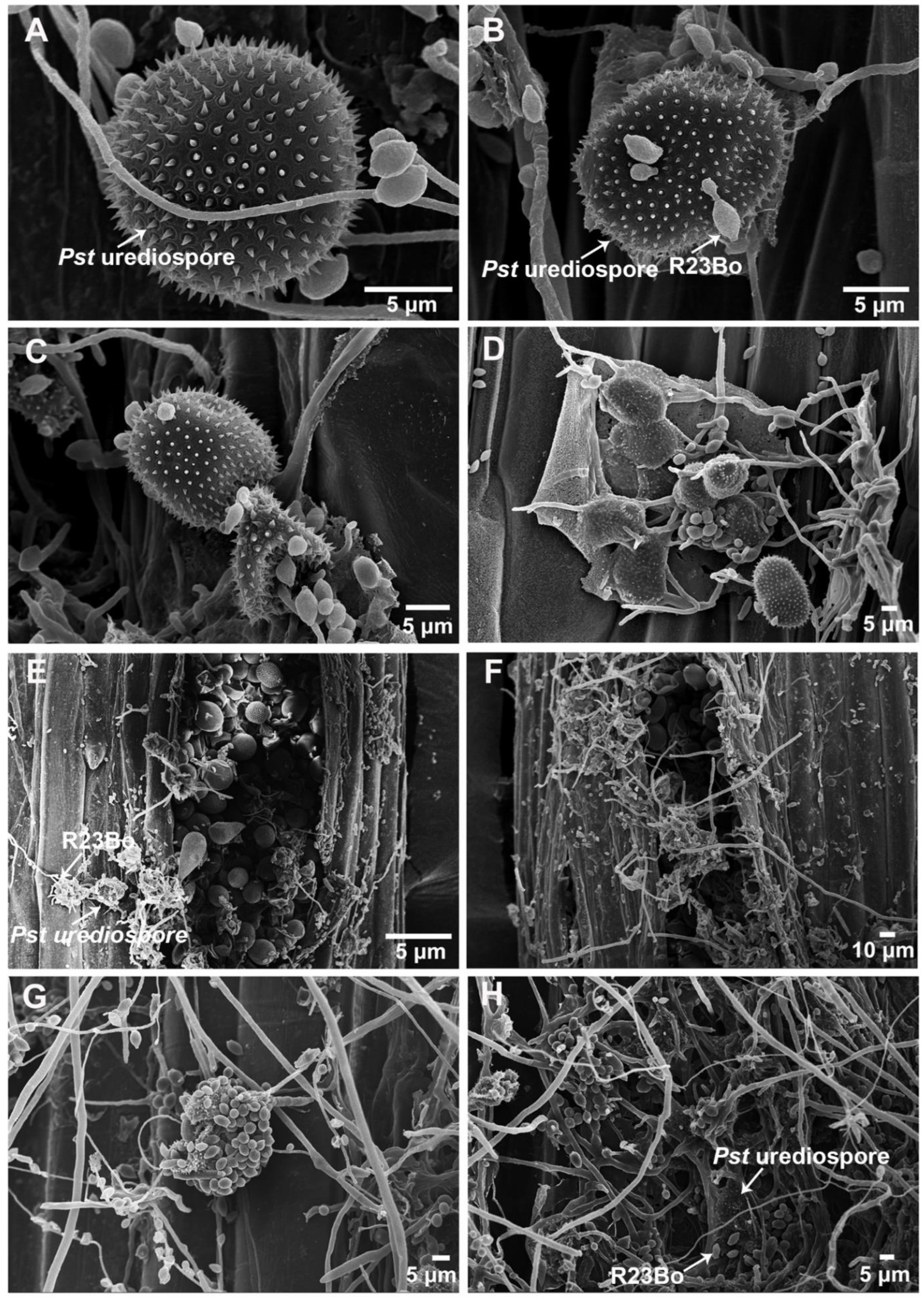
3.1. SEM Observations of Pst Uredinia Parasitized by the Hyperparasite
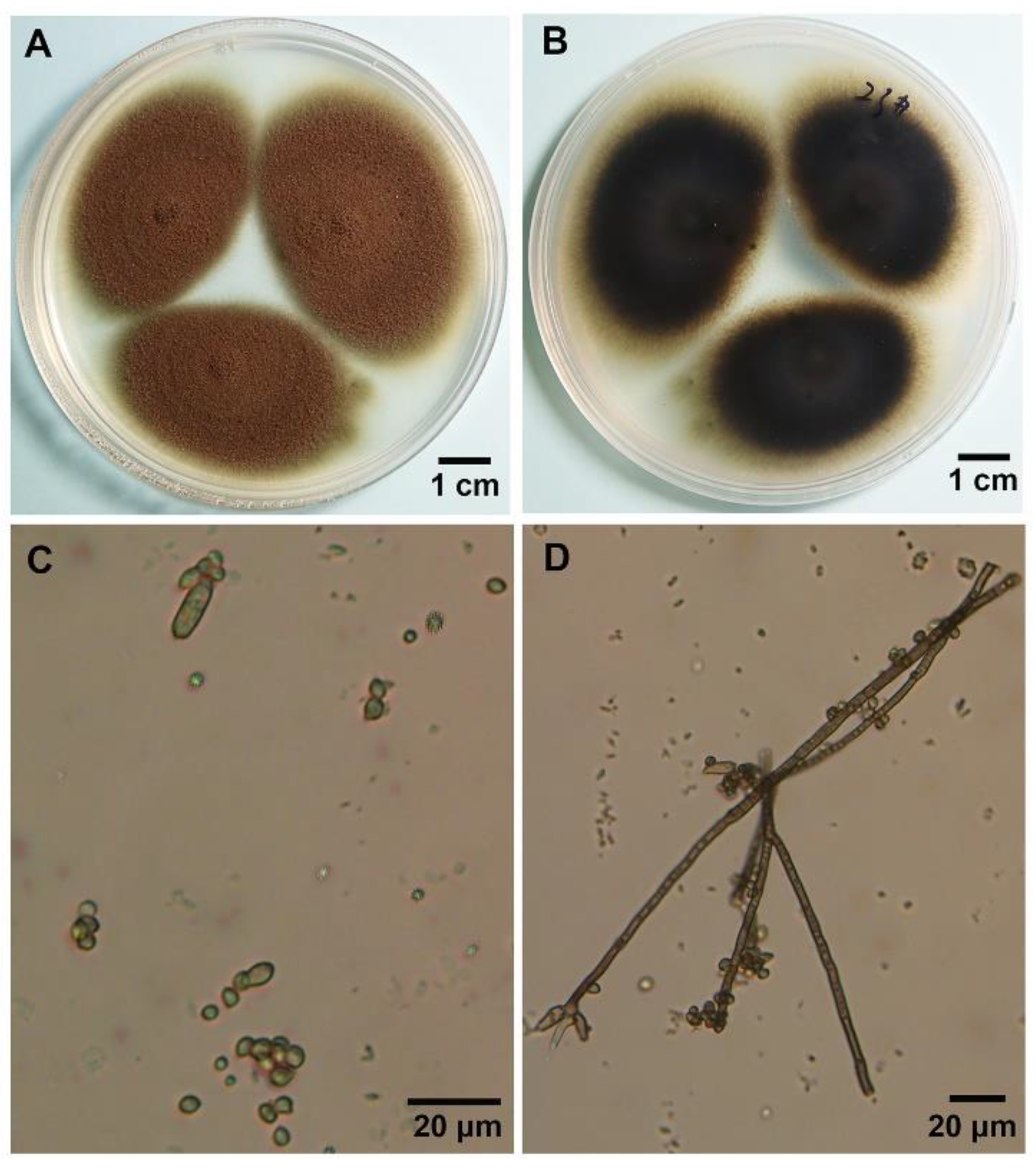
3.2. Morphological Characterization of the Hyperparasite
3.3. Molecular Characterization of the C. cladosporioides Isolate
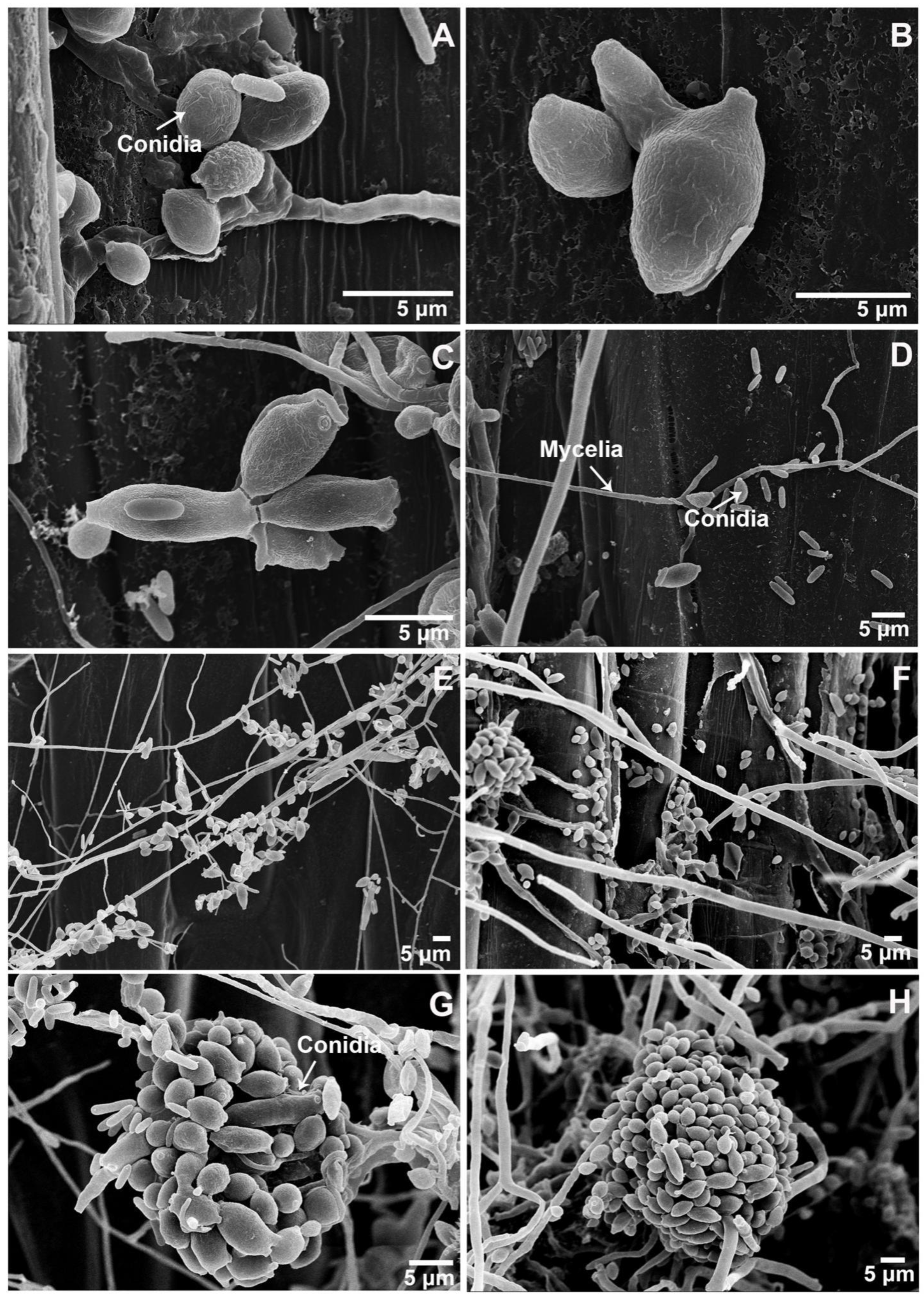
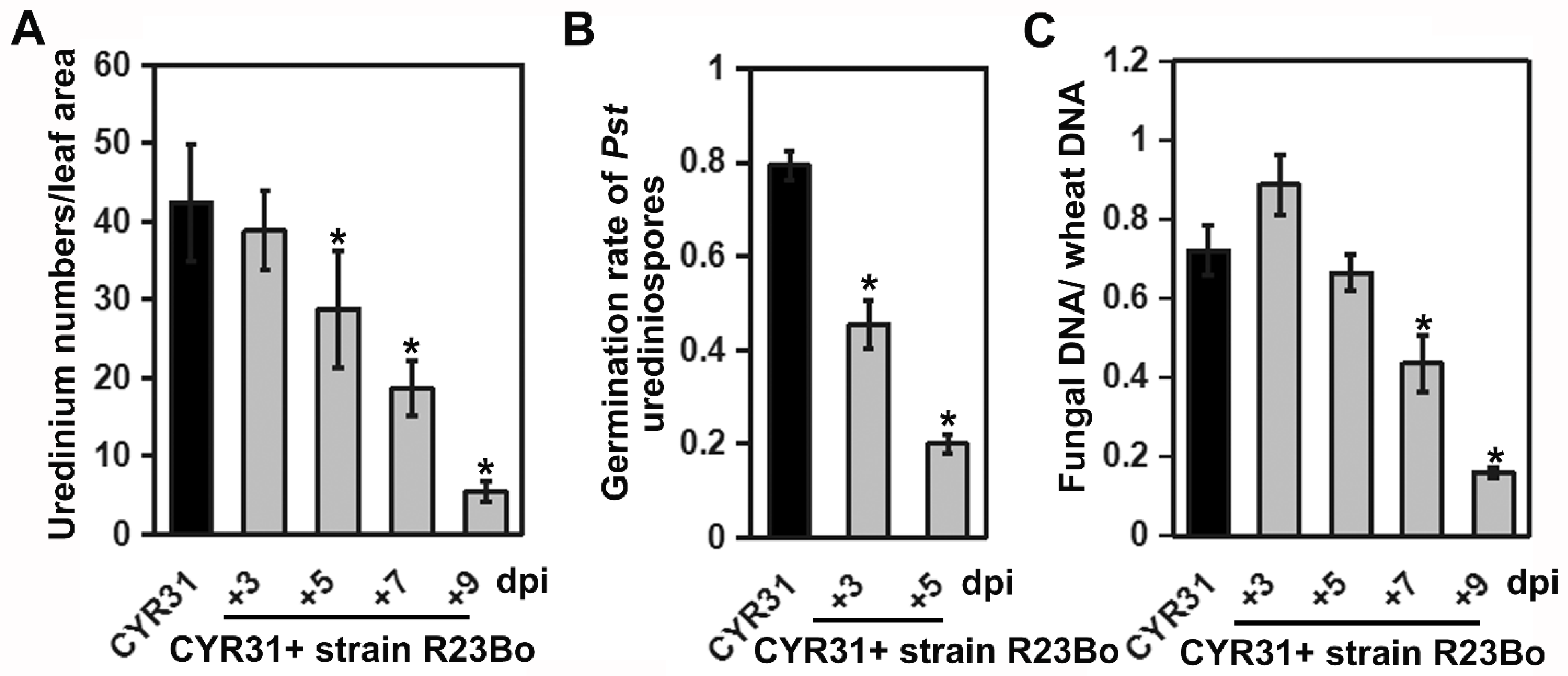
3.4. Confirmation of the C. cladosporioides Isolate Parasitizing Pst
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wellings, C.R. Global status of stripe rust: A review of historical and current threats. Euphytica 2011, 179, 129–141. [Google Scholar] [CrossRef]
- Wellings, C.R. Puccinia striiformis in Australia: A review of the incursion, evolution, and adaptation of stripe rust in the period 1979–2006. Aust. J. Agric. Res. 2007, 58, 567–575. [Google Scholar] [CrossRef]
- Beddow, J.M.; Pardey, P.G.; Chai, Y.; Hurley, T.M.; Kriticos, D.J.; Braun, H.J.; Park, R.F.; Cuddy, W.S.; Yonow, T. Research investment implications of shifts in the global geography of wheat stripe rust. Nat. Plants 2015, 1, 15132. [Google Scholar] [CrossRef]
- Chen, W.; Wu, L.; Liu, T.; Xu, S.; Jin, S.; Peng, Y.; Wang, B. Race Dynamics, Diversity, and Virulence Evolution in Puccinia striiformis f. sp. tritici, the Causal Agent of Wheat Stripe Rust in China from 2003 to 2007. Plant Dis. 2009, 93, 1093–1101. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Yu, H.X.; Kang, X.H.; Shen, H.M.; Li, C.; Liu, T.G.; Liu, B.; Chen, W.Q. Development of SCAR Markers and an SYBR Green Assay to Detect Puccinia striiformis f. sp. tritici in Infected Wheat Leaves. Plant Dis. 2016, 100, 1840–1847. [Google Scholar]
- Chen, X.; Kang, Z. Integrated Control of Stripe Rust. In Stripe Rust; Springer: Dordrecht, The Netherlands, 2017. [Google Scholar]
- Ansari, I.; El-Kady, M.M.; Arora, C.; Sundararajan, M.; Maiti, D.; Khan, A. A review on the fatal impact of pesticide toxicity on environment and human health. Glob. Clim. Chang. 2021, 361–391. [Google Scholar] [CrossRef]
- Aktar, W.; Sengupta, D.; Chowdhury, A. Impact of pesticides use in agriculture: Their benefits and hazards. Interdiscip. Toxicol. 2009, 2, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ainsworth, G.C.; Bisby, G.R. Ainsworth & Bisby’s dictionary of the fungi. Mycologist 1995, 17, 17–19. [Google Scholar]
- Köhl, J.; Molhoek, W.; Haas, G.D.; Geijn, G. Selection and orchard testing of antagonists suppressing conidia production of the apple scab pathogen Venturia inaequalis. Eur. J. Plant Pathol. 2009, 123, 401–414. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Kalkhove, S.I.C.; Lugones, L.G.; Baars, J.P.; Wösten, H.A.M.; Bakker, P.A. Effects of fluorescent Pseudomonas spp. isolated from mushroom cultures on Lecanicillium fungicola. Biol. Control 2012, 63, 210–221. [Google Scholar] [CrossRef]
- Adhikari, A.; Nandi, S.; Dutta, S.; Bhattacharya, I.; Mandal, T. Study of morphology and Mycoparasitism of some antagonists of Trichoderma sp. from West Bengal, India. J. Mt. Sinai Hosp. N. Y. 2014, 22, 91. [Google Scholar]
- Zhong, X.; Li, S.S.; Peng, Q.Y.; Zhang, J.S.; Kan, X.T.; Zhang, G.R.; Liu, X. A Polycephalomyces hyperparasite of Ophiocordyceps sinensis leads to shortened duration of production and reduced numbers of host ascospores. Fungal Ecol. 2016, 21, 24–31. [Google Scholar] [CrossRef]
- Kiss, L.; Russell, J.C.; Szentiványi, O.; Xu, X.; Jeffries, P. Biology and biocontrol potential of Ampelomyces mycoparasites, natural antagonists of powdery mildew fungi. Biocontrol Sci. Technol. 2004, 14, 635–651. [Google Scholar] [CrossRef]
- Ikeda, S.; Shimizu, A.; Shimizu, M.; Takahashi, H.; Takenaka, S. Biocontrol of black scurf on potato by seed tuber treatment with Pythium oligandrum. Biol. Control 2012, 60, 297–304. [Google Scholar] [CrossRef]
- Kapooria, R.G.; Sinha, S. Phylloplane mycoflora of pearl millet and its influence on the development of Puccinia penniseti. Trans. Br. Mycol. Soc. 1969, 53, 153–155. [Google Scholar] [CrossRef]
- Koçl, N.K.; Défago, G. Studies on the Host Range of the Hyperparasite Aphanocladium album. J. Phytopathol. 2010, 107, 214–218. [Google Scholar]
- Cummins, G.B.; Littlefield, L.J. Biology of the plant rusts: An introduction. Bioence 1984, 34, 116. [Google Scholar]
- Moricca, S.; Ragazzi, A.; Mitchelson, K.R.; Assante, G. Antagonism of the Two-Needle Pine Stem Rust Fungi Cronartium flaccidum and Peridermium pini by Cladosporium tenuissimum In Vitro and In Planta. Phytopathology 2001, 91, 457–468. [Google Scholar] [CrossRef] [Green Version]
- Tsuneda, A.; Hiratsuka, Y.; Maruyama, P.J. Hyperparasitism of Scytalidium uredinicola on western gall rust, Endocronartium harknessii. Can. J. Bot. 1980, 58, 1154–1159. [Google Scholar] [CrossRef]
- Li, Z.; Jie, Z.; Liang, X.; Zhan, G.; Kang, Z. Identification of a Novel Alternaria alternata Strain Able to Hyperparasitize Puccinia striiformis f. sp. tritici, the Causal Agent of Wheat Stripe Rust. Front. Microbiol. 2017, 8, 71. [Google Scholar]
- Zhan, G.; Tian, Y.; Wang, F.; Chen, X.; Guo, J.; Jiao, M.; Huang, L.; Kang, Z. A novel fungal hyperparasite of Puccinia striiformis f. sp. tritici, the causal agent of wheat stripe rust. PLoS ONE 2014, 9, e111484. [Google Scholar]
- Wang, N.; Fan, X.; Zhang, S.; Liu, B.; Wang, X. Identification of a Hyperparasitic Simplicillium obclavatum strain affecting the infection dynamics of Puccinia striiformis f. sp. tritici on wheat. Front. Microbiol. 2020, 11, 1277. [Google Scholar] [CrossRef]
- Bomblies, K.; Shukla, V.; Graham, C. Scanning Electron Microscopy (SEM) of Plant Tissues. Cold Spring Harb. Protoc. 2008, 2008, pdb.prot4933. [Google Scholar] [CrossRef]
- Wang, Y.B.; Yu, H.; Dai, Y.D.; Wu, C.K.; Zeng, W.B.; Yuan, F.; Liang, Z.Q. Polycephalomyces agaricus, a new hyperparasite of Ophiocordyceps sp. infecting melolonthid larvae in southwestern China. Mycol. Prog. 2015, 14, 70. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.S.; Wang, W.L.; Li, J.C.; Wang, Y.Y.; Tsay, J.G. First Report of Papaya Scab Caused by Cladosporium cladosporioides in Taiwan. Plant Dis. 2009, 93, 426. [Google Scholar] [CrossRef]
- Bensch, K.; Groenewald, J.Z.; Dijksterhuis, J.; Starink-Willemse, M.; Andersen, B.; Summerell, B.A.; Shin, H.D.; Dugan, F.M.; Schroers, H.J.; Braun, U. Species and ecological diversity within the Cladosporium cladosporioides complex (Davidiellaceae, Capnodiales). Stud. Mycol. 2010, 67, 1–94. [Google Scholar] [CrossRef]
- Shao, M.W.; Lu, Y.H.; Miao, S.; Zhang, Y.; Chen, T.T.; Zhang, Y.L. Diversity, Bacterial Symbionts and Antibacterial Potential of Gut-Associated Fungi Isolated from the Pantala flavescens Larvae in China. PLoS ONE 2015, 10, e0134542. [Google Scholar] [CrossRef]
- Schubert, K.; Groenewald, J.Z.; Braun, U.; Dijksterhuis, J.; Starink, M.; Hill, C.F.; Zalar, P.; de Hoog, G.S.; Crous, P.W. Biodiversity in the Cladosporium herbarum complex (Davidiellaceae, Capnodiales), with standardisation of methods for Cladosporium taxonomy and diagnostics. Stud. Mycol. 2007, 58, 105–156. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Vandermeer, J.; Perfecto, I.; Liere, H. Evidence for hyperparasitism of coffee rust (Hemileia vastatrix) by the entomogenous fungus, Lecanicillium lecanii, through a complex ecological web. Plant Pathol. 2010, 58, 636–641. [Google Scholar] [CrossRef] [Green Version]
- Dugan, F.; Braun, U.; Groenewald, J.; Crous, P. Morphological plasticity in Cladosporium sphaerospermum. Persoonia 2008, 21, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Heather, W.A.; Sharma, J.K. Some Aspects of Poplar Rust Research in Australia. Aust. For. 1977, 40, 28–43. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Accession Number | GenBank Number (ITS, EF, ACT) | Substrate (Including Host) | Country | Reference |
|---|---|---|---|---|---|
| C. cladosporioides | Cc-6 | EU935608.1 | Carica papaya | China | Chen et al., 2009 [27] |
| C. cladosporioides | UM15 | HQ148094.1 | Echinacea purpurea | USA | Bensch et al., 2010 [28] |
| C. cladosporioides | QTYC16 | KM103301.1 | Pantala flavescens larvae | China | Shao et al., 2015 [29] |
| C. cladosporioides | FCBP:1493 | KY290222.1 | Solanum melongena | Pakistan | - |
| C. cladosporioides | CBS 112388 | HM148003 HM148244 HM148490 | Indoor air | Germany | Bensch et al., 2010 [28] |
| C. cladosporioides | CBS 143.35 | HM148011 HM148252 HM148498 | Pisum sativum | South Africa | Bensch et al., 2010 [28] |
| C. cladosporioides | CBS 144.35 | HM148012 HM148253 HM148499 | P. sativum | USA | Bensch et al., 2010 [28] |
| C. cladosporioides | CBS 101367 | HM148002 HM148489 HM148243 | Soil | Brazil | Bensch et al., 2010 [28] |
| C. cladosporioides | CPC 14018 MRC 10810 | HM148040 HM148281 HM148527 | Triticum aestivum | South Africa | Bensch et al., 2010 [28] |
| C. cladosporioides | CPC 14244 | HM148044 HM148285 HM148531 | Magnolia spp. | USA | Bensch et al., 2010 [28] |
| C. cladosporioides | CBS 145.35 | HM148013 HM148254 HM148500 | P. sativum | Germany | Bensch et al., 2010 [28] |
| C. cladosporioides | 14PI001 CGMCC 7.175 | KJ598781 KM281945 KJ598781 | Puccinia striiformis | China | Zhan et al., 2014 [22] |
| C. herbarum | CBS 121621 | EF679363 | Hordeum vulgare | Netherlands | Schubert et al., 2007 [30] |
| C. herbarum | CBS 132.29 | HM148010 HM148251 HM148497 | -a | - | Bensch et al., 2010 [28] |
| C. herbarum | CBS 170.54 | MH857281 IMI025324 NCTC6740 | - | England | Vu et al., 2018 [31] |
| C. oxysporum | CBS 125991 CPC 14371 BA 1738 | HM148118 HM148362 HM148607 | Soil, near the terracotta | China | Bensch et al., 2010 [28] |
| C. oxysporum | CBS 126351 CPC 14308 BA 1707 | HM148119 HM148363 HM148608 | Indoor air | Venezuela | Bensch et al., 2010 [28] |
| C. pseudocladosporioides | CBS 149.66 | HM148161 HM148405 HM148650 | T. aestivum | USA | Bensch et al., 2010 [28] |
| C. pseudocladosporioides | CBS 176.82 | HM148162 HM148406 HM148651 | Pteridium aquilinum | Romania | Bensch et al., 2010 [28] |
| C. salinae | CBS 119413 EXF-335 | DQ780374 JN906993 EF101390 | Hypersaline water | Slovenia | Bensch et al., 2010 [28] |
| C. tenuissimum | CPC 12223 | HM148208 HM148453 HM148698 | Rust | Brazil | Bensch et al., 2010 [28] |
| C. tenuissimum | CBS 126501 CPC 14410 | HM148199 HM148444 HM148689 | Musa spp. | Ivory Coast | Bensch et al., 2010 [28] |
| C. xylophilum | CBS 126588 | HM148231 HM148477 HM148722 | Salix viminalis twigs | Italy | Bensch et al., 2010 [28] |
| C. xylophilum | CPC 14281 | HM148233 HM148479 HM148724 | Leaves | France | Bensch et al., 2010 [28] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; He, M.; Fan, X.; Dai, L.; Zhang, S.; Hu, Z.; Wang, N. Isolation, Identification and Hyperparasitism of a Novel Cladosporium cladosporioides Isolate Hyperparasitic to Puccinia striiformis f. sp. tritici, the Wheat Stripe Rust Pathogen. Biology 2022, 11, 892. https://doi.org/10.3390/biology11060892
Zhang H, He M, Fan X, Dai L, Zhang S, Hu Z, Wang N. Isolation, Identification and Hyperparasitism of a Novel Cladosporium cladosporioides Isolate Hyperparasitic to Puccinia striiformis f. sp. tritici, the Wheat Stripe Rust Pathogen. Biology. 2022; 11(6):892. https://doi.org/10.3390/biology11060892
Chicago/Turabian StyleZhang, Hongjuan, Mengying He, Xin Fan, Lu Dai, Shan Zhang, Zeyu Hu, and Ning Wang. 2022. "Isolation, Identification and Hyperparasitism of a Novel Cladosporium cladosporioides Isolate Hyperparasitic to Puccinia striiformis f. sp. tritici, the Wheat Stripe Rust Pathogen" Biology 11, no. 6: 892. https://doi.org/10.3390/biology11060892
APA StyleZhang, H., He, M., Fan, X., Dai, L., Zhang, S., Hu, Z., & Wang, N. (2022). Isolation, Identification and Hyperparasitism of a Novel Cladosporium cladosporioides Isolate Hyperparasitic to Puccinia striiformis f. sp. tritici, the Wheat Stripe Rust Pathogen. Biology, 11(6), 892. https://doi.org/10.3390/biology11060892