
The Role of ZIP9 and Androgen Receptor in the Establishment of Tight Junctions between Adult Rat Sertoli Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Primary Adult Rat Sertoli Cells
2.2. Measurement of Transepithelial Resistance (TER)
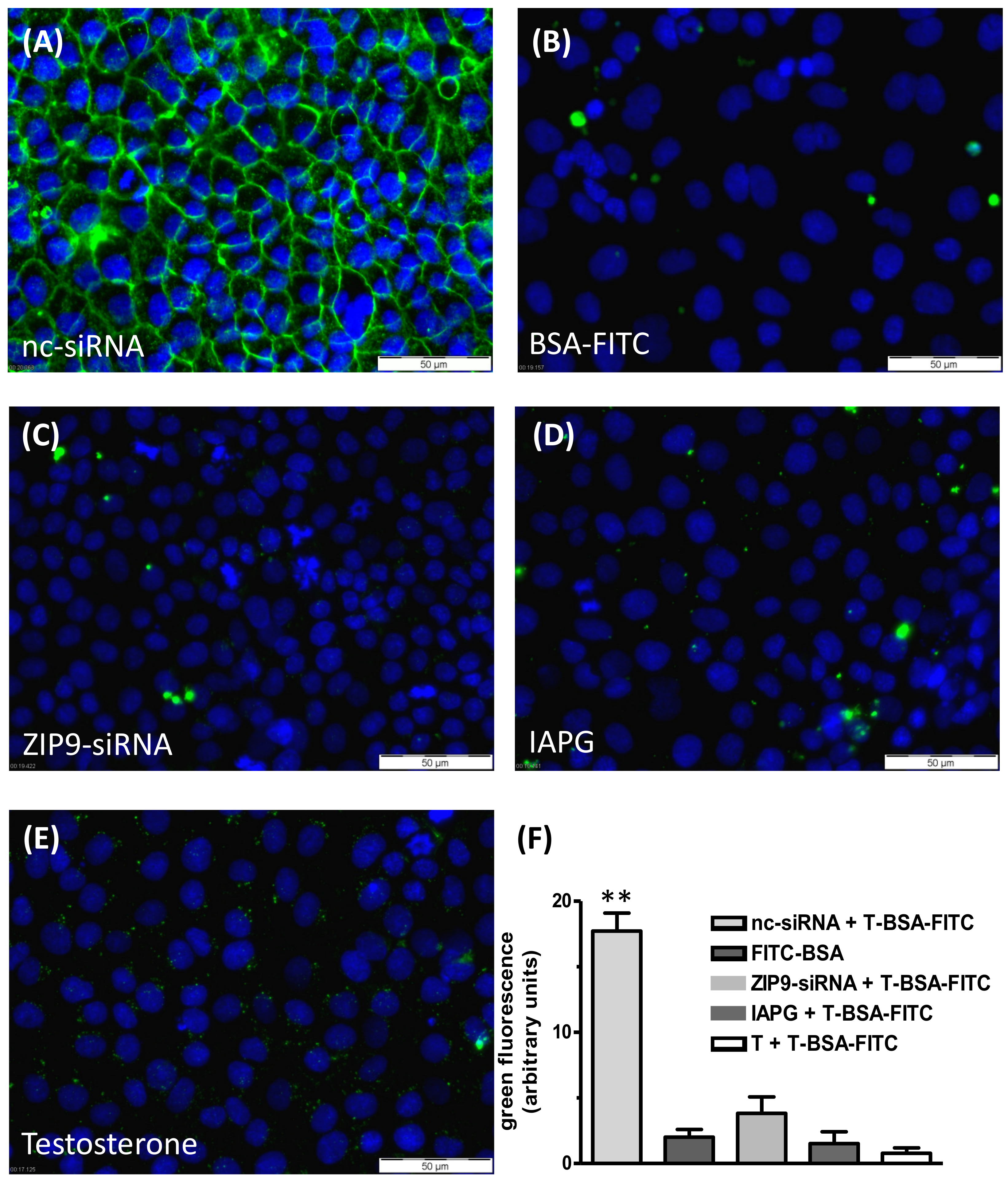
2.3. Plasma Membrane Labeling with Testosterone-BSA-FITC
2.4. Preparation of Cell Lysates from PASC1
2.5. Western Blotting
2.6. Immunofluorescence
2.7. RNA Extraction and Quantitative Real Time PCR
2.8. Silencing Expression of ZIP9 or AR via siRNA
2.9. Statistical Analysis
3. Results
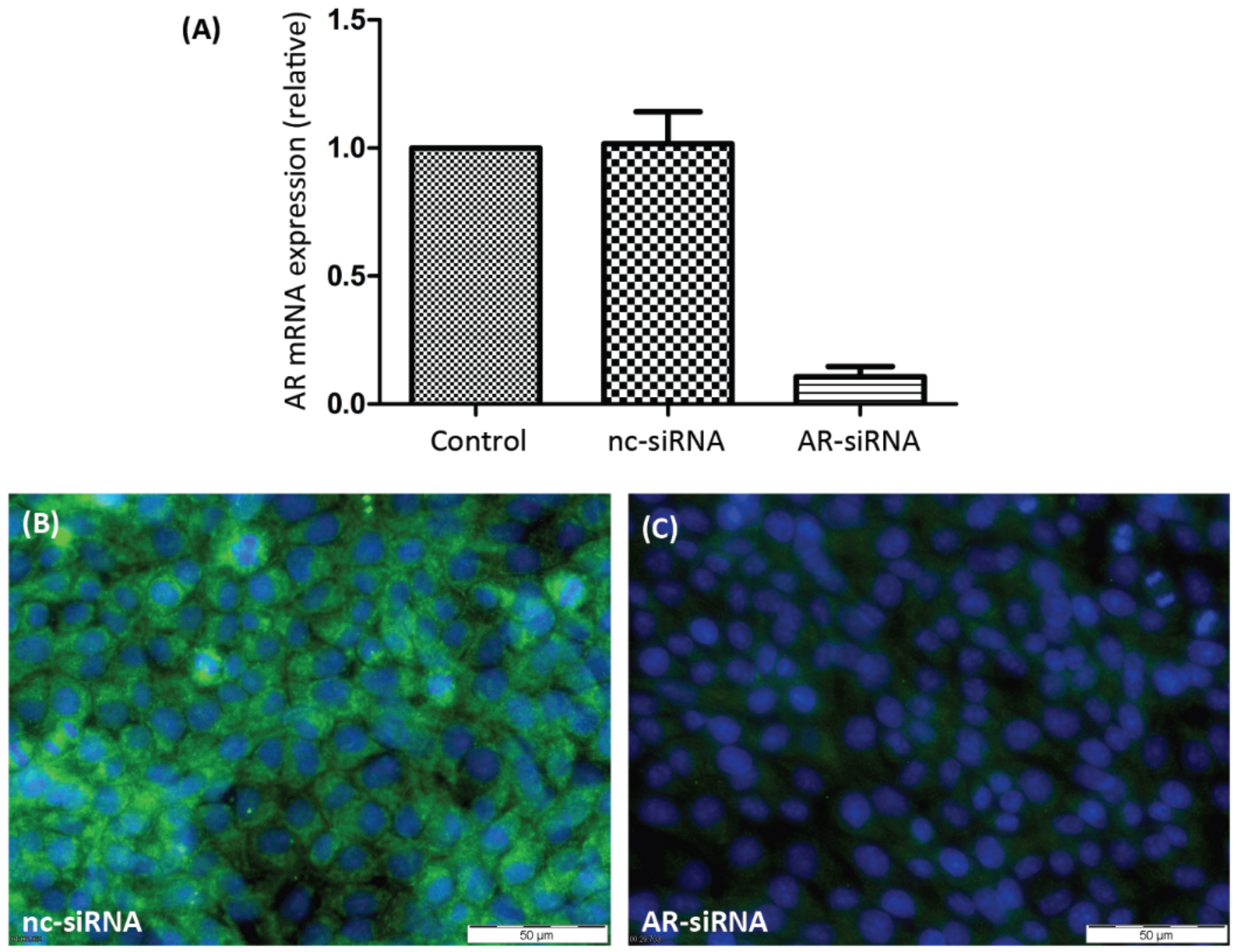
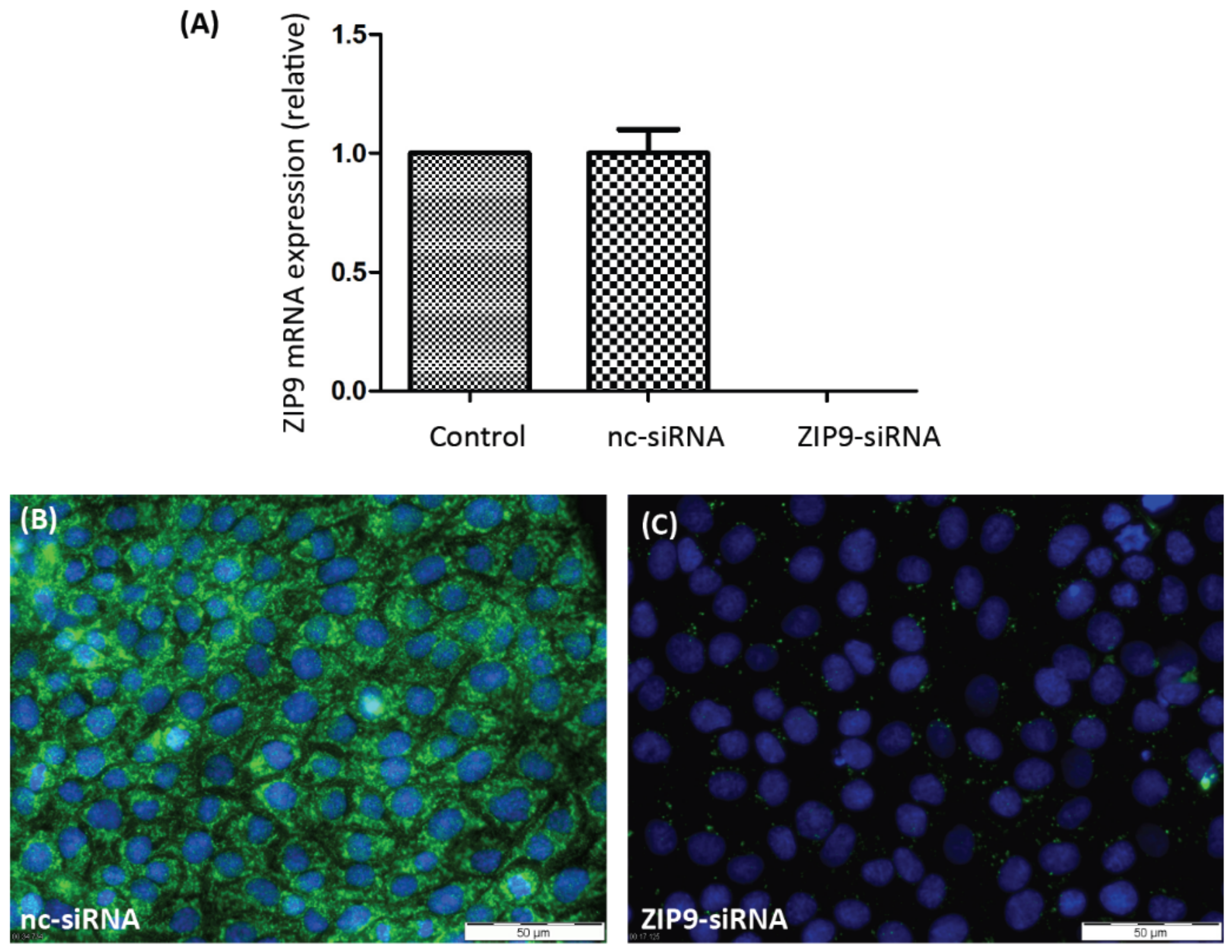
3.1. Silencing AR or ZIP9 Expression by siRNA
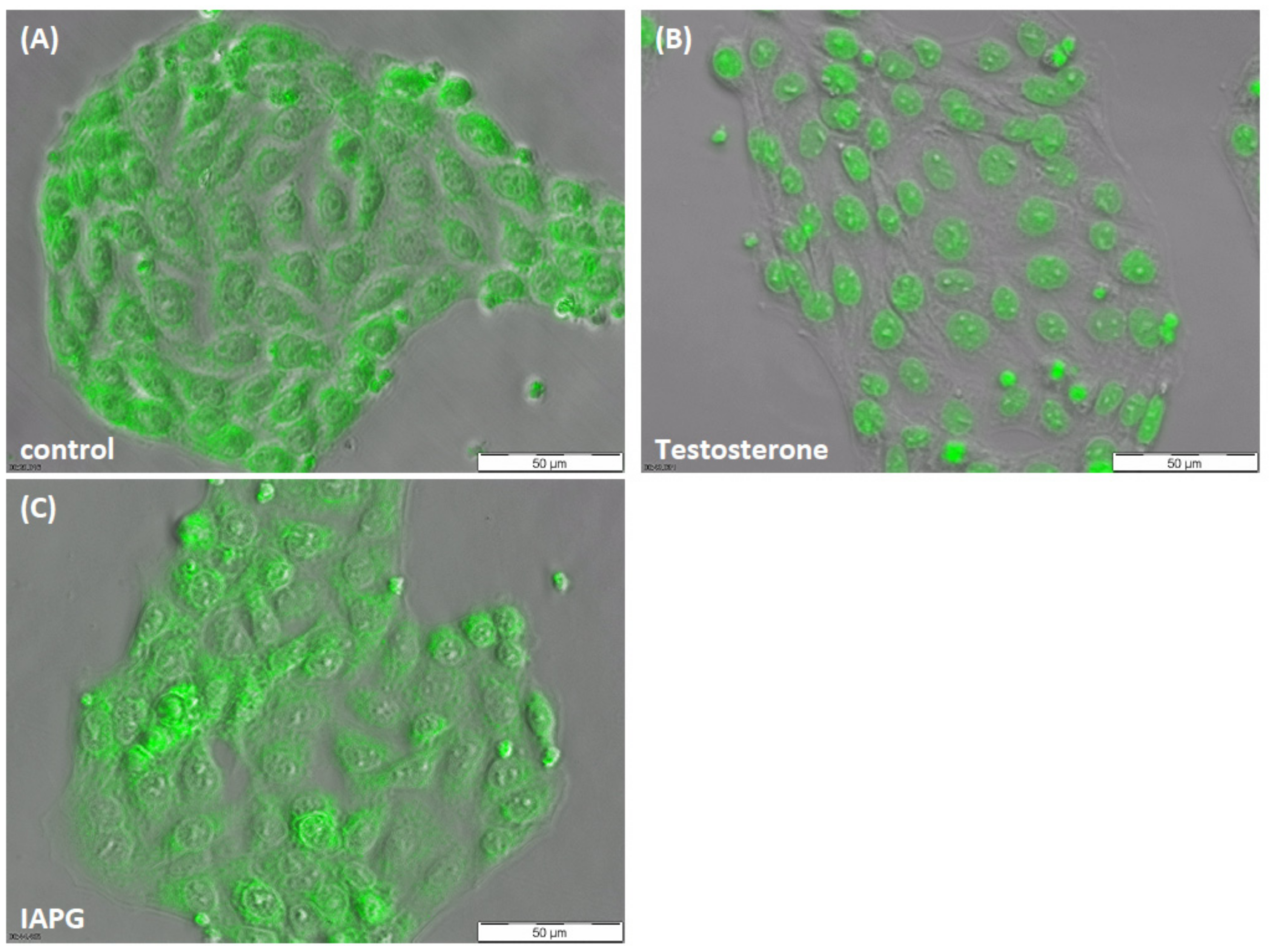
3.2. Responsiveness of PASC1 Cell AR towards Testosterone or the ZIP9-Targeting Peptide IAPG
3.3. Binding of the IAPG Peptide or Testosterone to the Androgen Binding Site of ZIP9
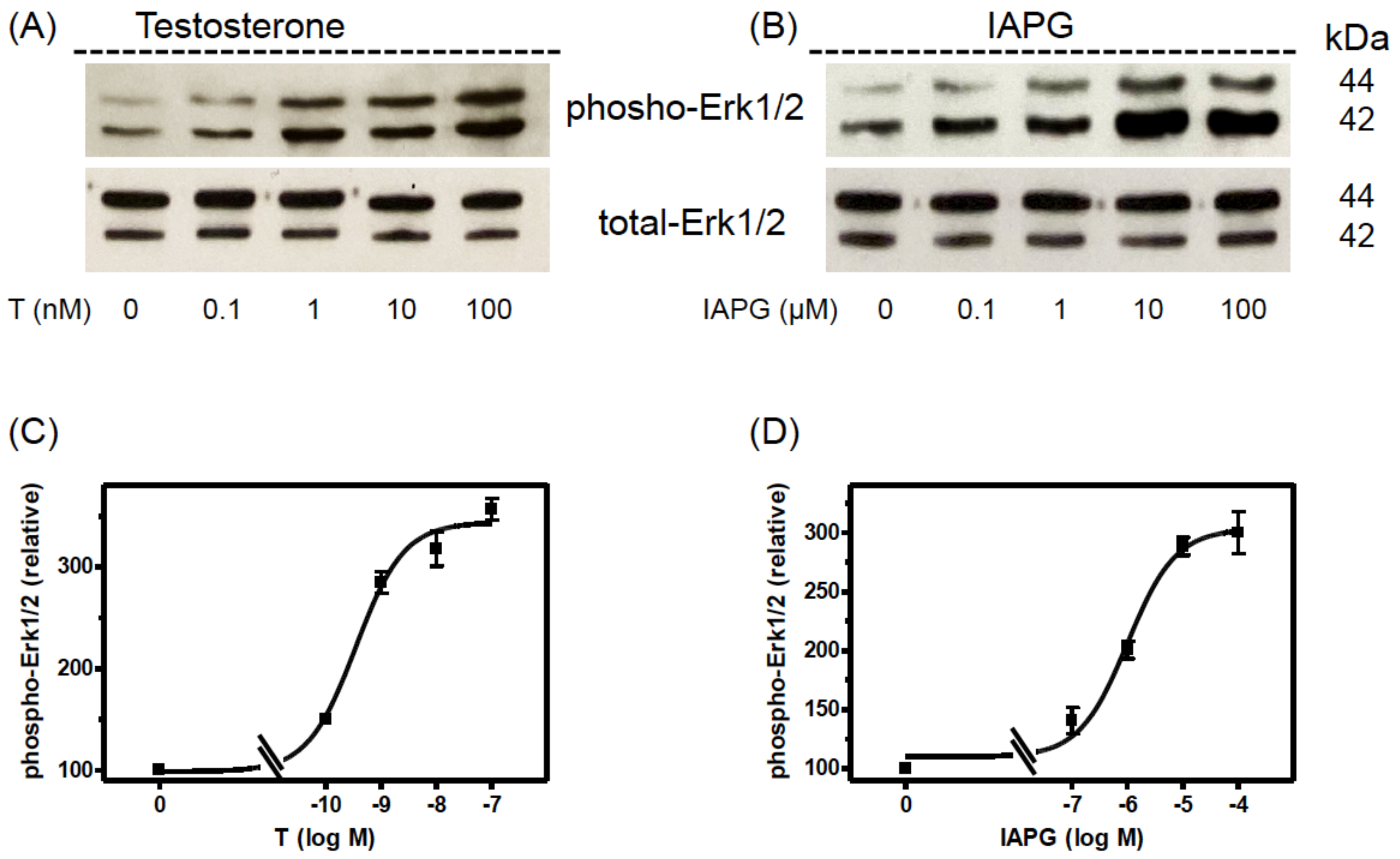
3.4. Stimulation of Erk1/2 Phosphorylation by Testosterone or IAPG
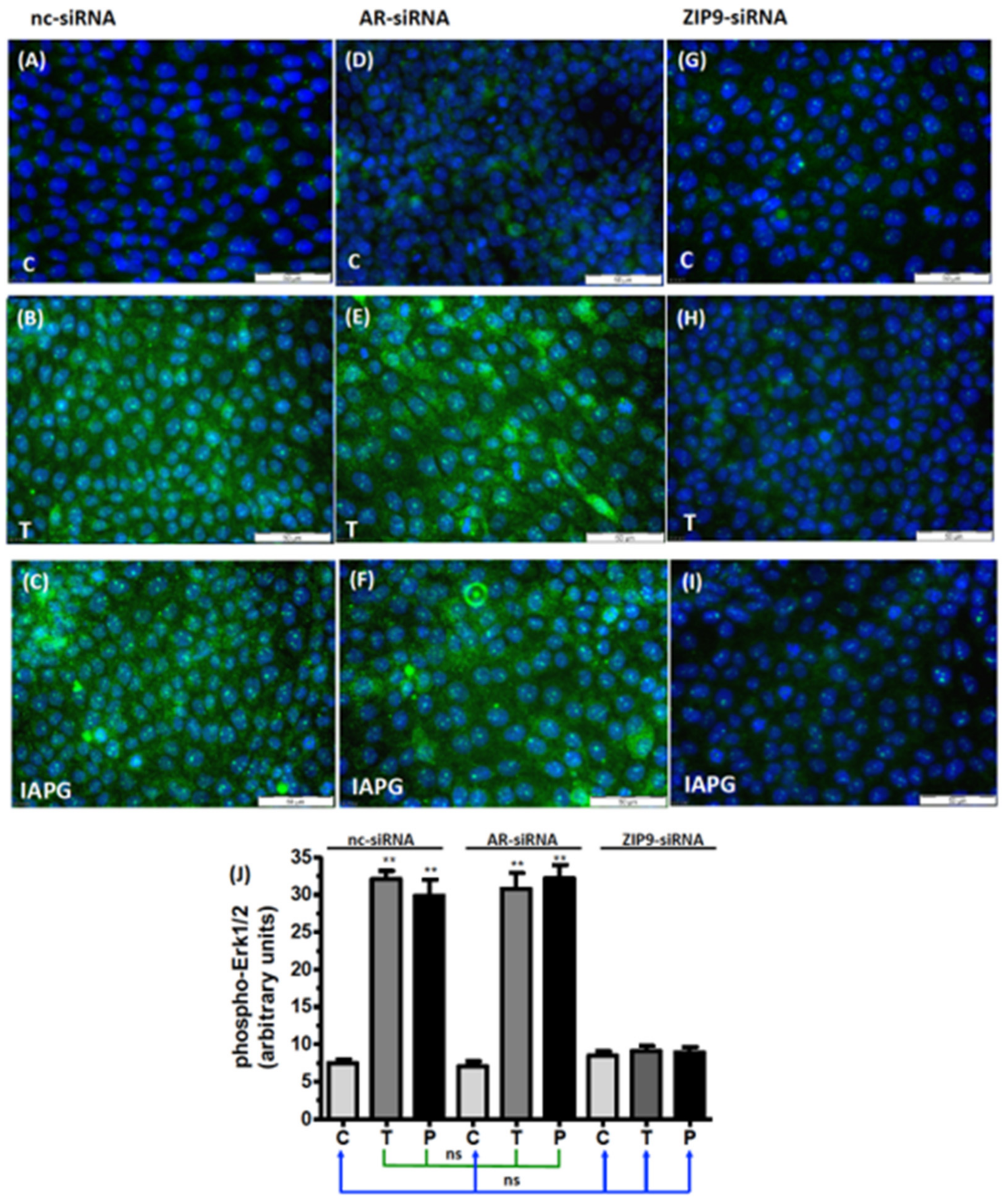
3.5. Identification of the Androgen Receptor Involved in Erk1/2 Phosphorylation
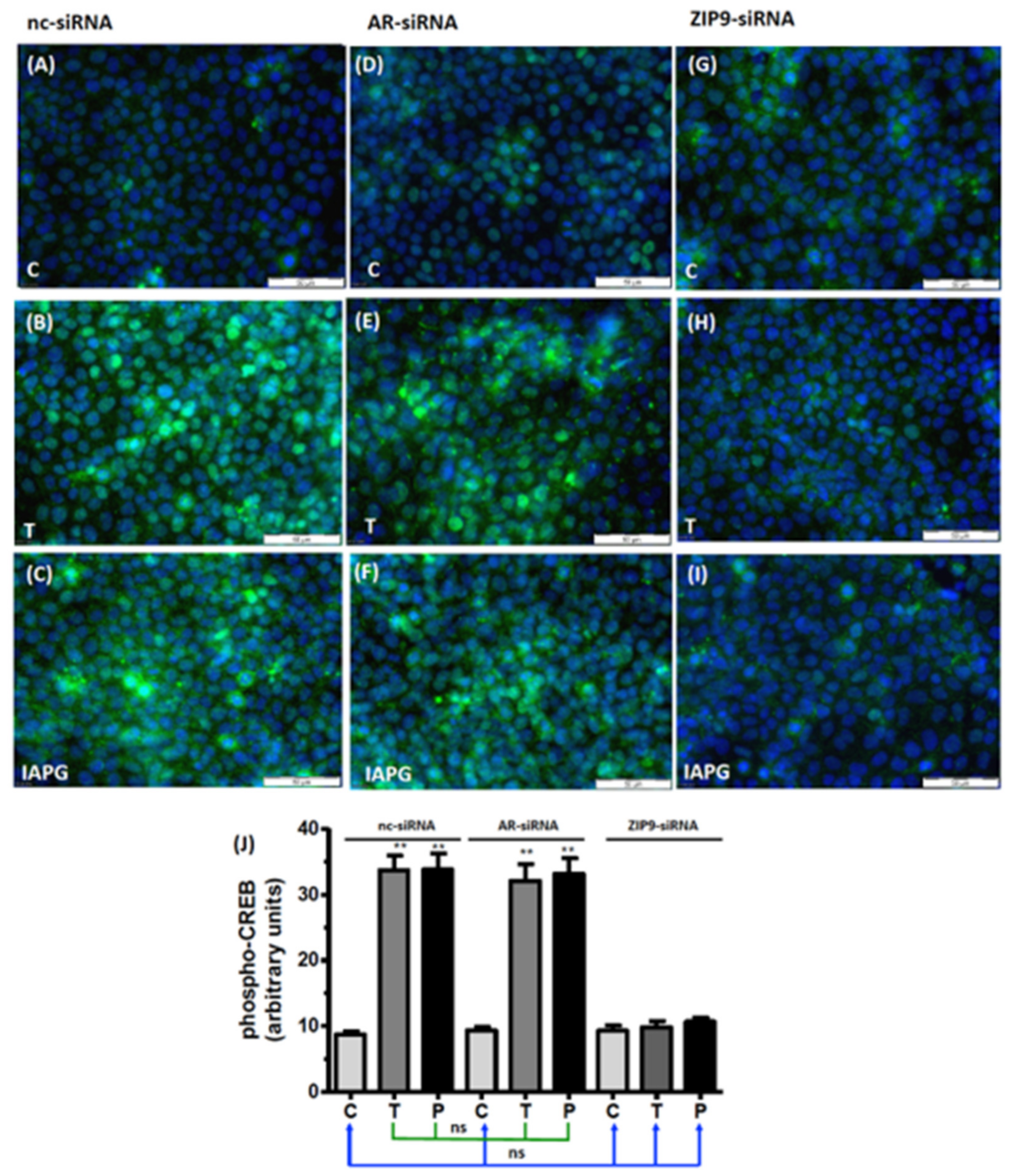
3.6. Involvement of AR or ZIP9 in the Stimulation of CREB/ATF-1 Phosphorylation by Testosterone or IAPG
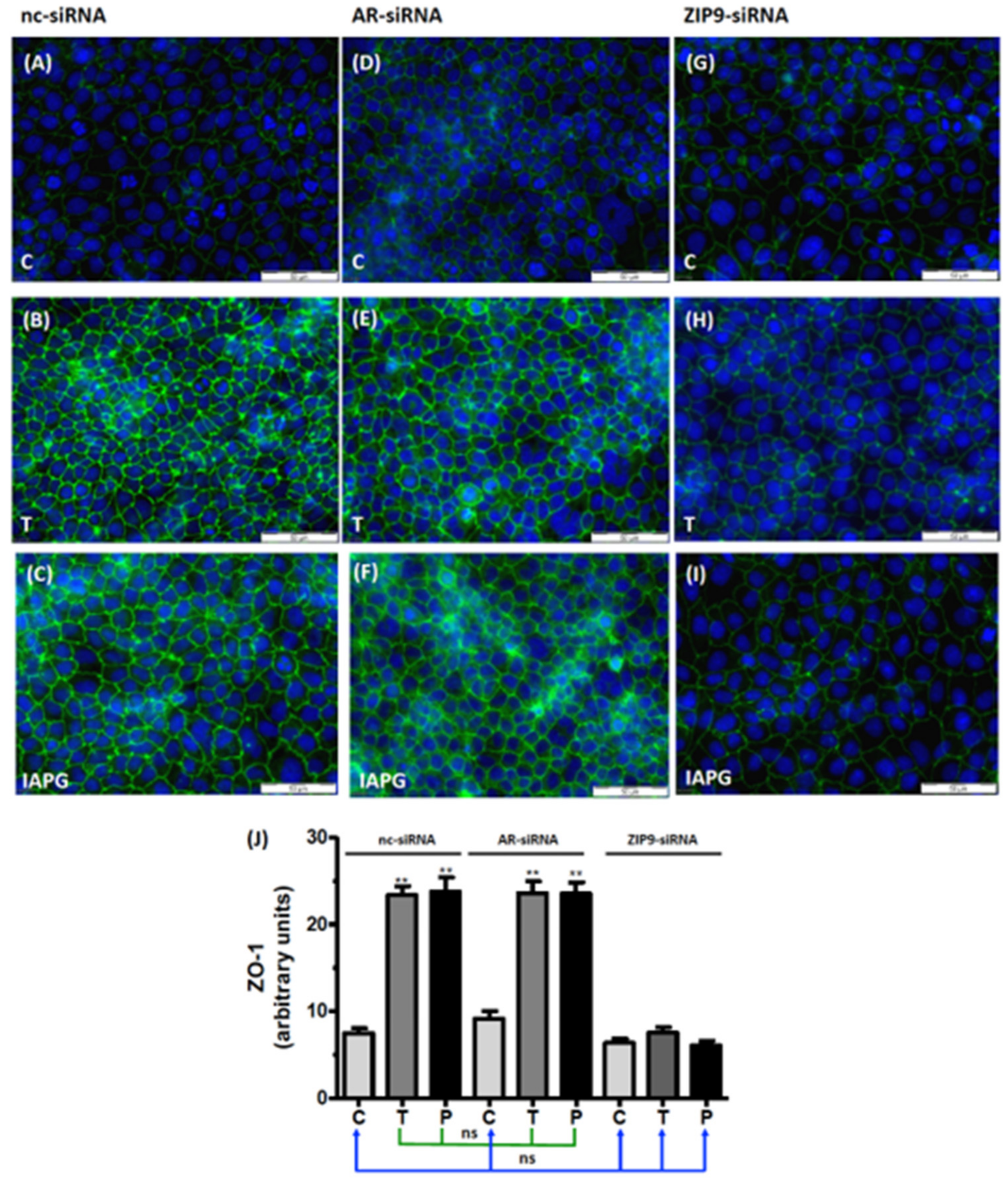
3.7. Involvement of AR or ZIP9 in the Stimulation of ZO-1 Expression by Testosterone or IAPG
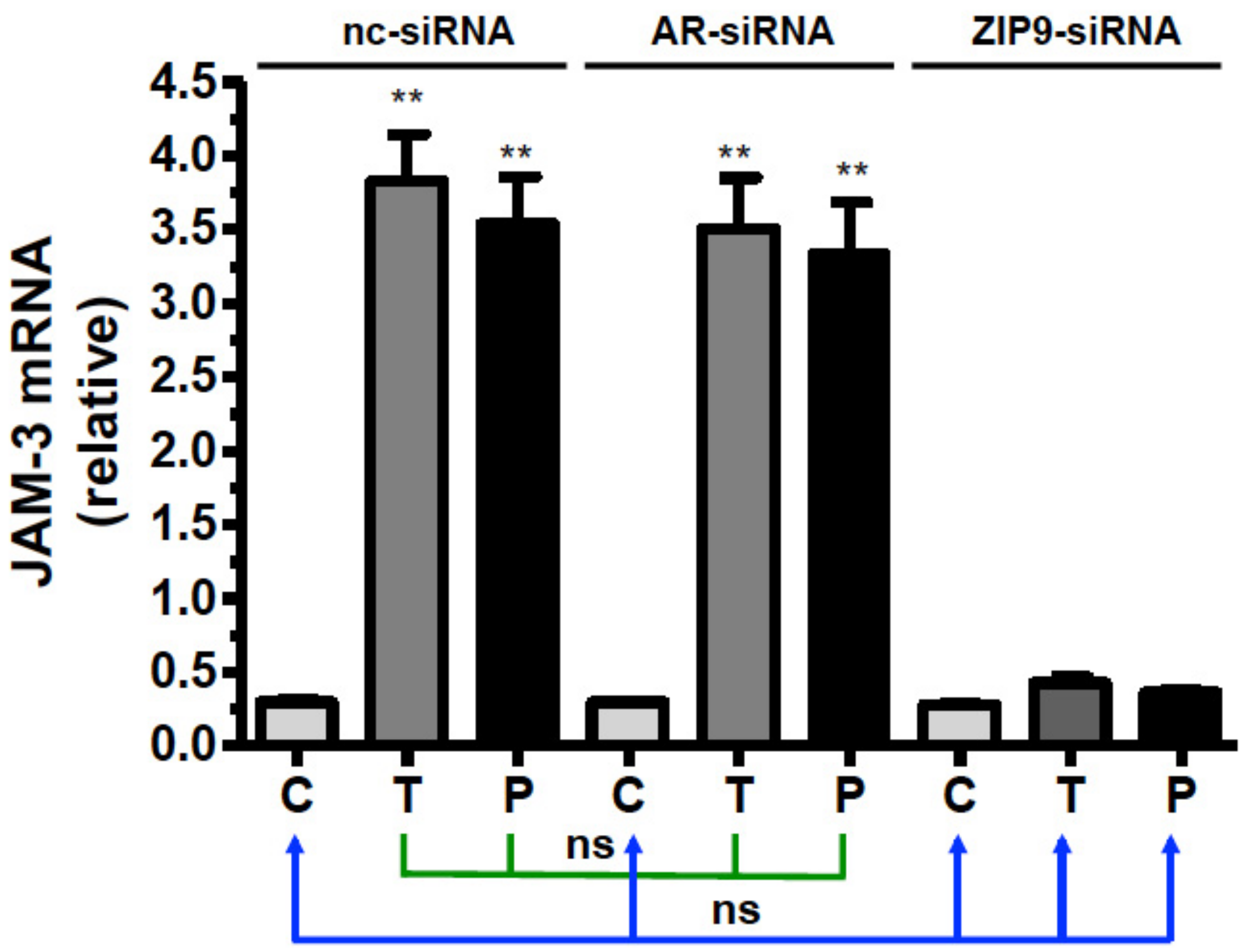
3.8. Participation of AR or ZIP9 in the Stimulation of Cldn-1 and JAM-3 Expression by Testosterone or IAPG
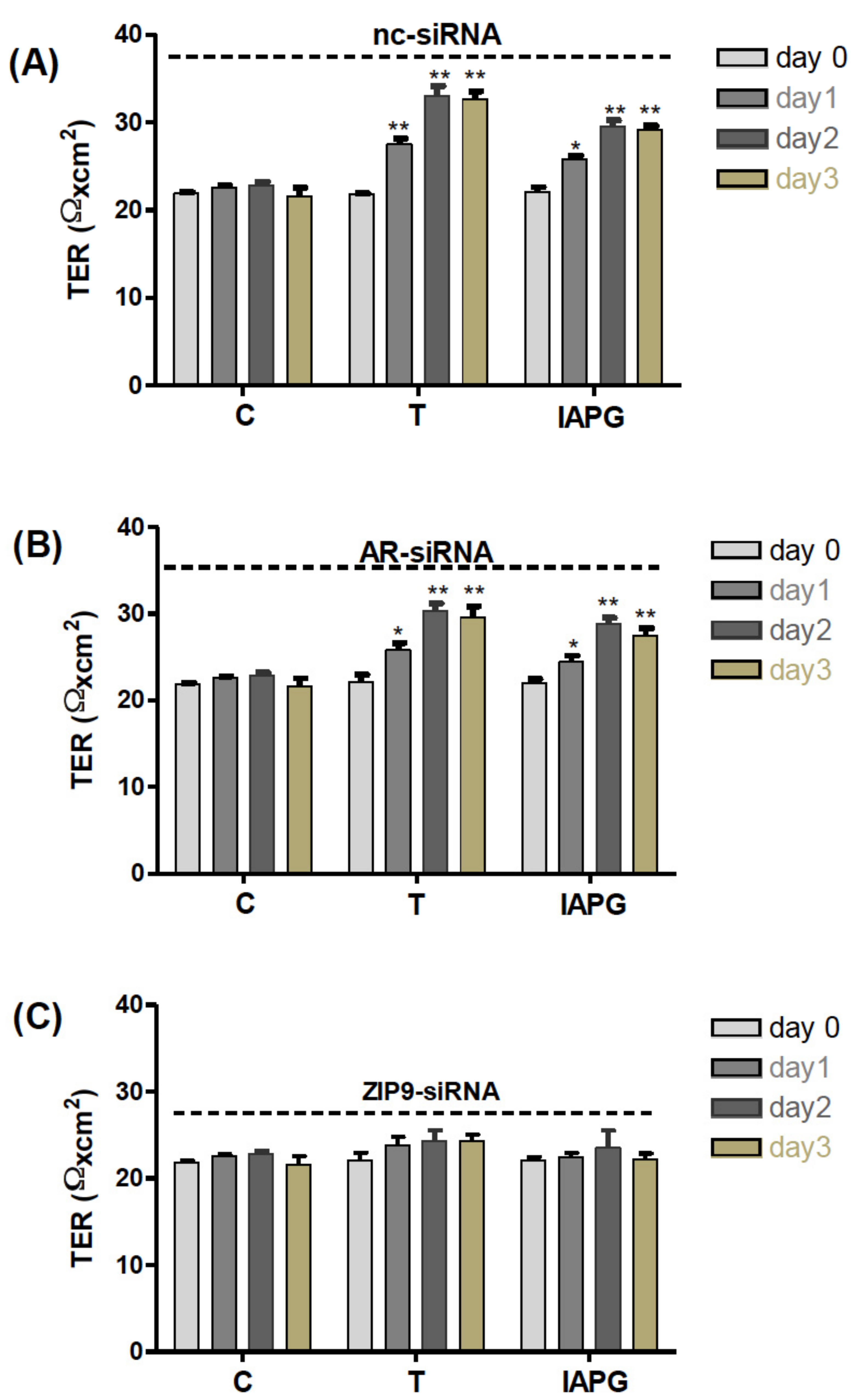
3.9. Involvement of AR or ZIP9 in Testosterone- or IAPG-Induced TJ Formation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheng, C.Y.; Mruk, D.D. The blood-testis barrier and its implications for male contraception. Pharmacol. Rev. 2012, 64, 16–64. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Holdcraft, R.W.; Shima, J.E.; Griswold, M.D.; Braun, R.E. Androgens regulate the permeability of the blood-testis barrier. Proc. Natl. Acad. Sci. USA 2005, 102, 16696–16700. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.S.; Yeh, S.; Chen, L.M.; Lin, H.Y.; Zhang, C.; Ni, J.; Wu, C.C.; di Sant’Agnese, P.A.; deMesy-Bentley, K.L.; Tzeng, C.R.; et al. Androgen receptor in sertoli cell is essential for germ cell nursery and junctional complex formation in mouse testes. Endocrinology 2006, 147, 5624–5633. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Mostaghel, E.A.; Vakar-Lopez, F.; Montgomery, B.; True, L.; Nelson, P.S. Testosterone regulates tight junction proteins and influences prostatic autoimmune responses. Horm. Cancer 2011, 2, 145–156. [Google Scholar] [CrossRef]
- Meinhardt, A.; Hedger, M.P. Immunological, paracrine and endocrine aspects of testicular immune privilege. Mol. Cell Endocrinol. 2011, 335, 60–68. [Google Scholar] [CrossRef]
- Fijak, M.; Bhushan, S.; Meinhardt, A. Immunoprivileged sites: The testis. Methods Mol. Biol. 2011, 677, 459–470. [Google Scholar] [CrossRef]
- Wong, E.W.; Mruk, D.D.; Lee, W.M.; Cheng, C.Y. Regulation of blood-testis barrier dynamics by TGF-beta3 is a Cdc42-dependent protein trafficking event. Proc. Natl. Acad. Sci. USA 2010, 107, 11399–11404. [Google Scholar] [CrossRef]
- Yan, H.H.; Mruk, D.D.; Lee, W.M.; Cheng, C.Y. Blood-testis barrier dynamics are regulated by testosterone and cytokines via their differential effects on the kinetics of protein endocytosis and recycling in Sertoli cells. FASEB J. 2008, 22, 1945–1959. [Google Scholar] [CrossRef]
- Ilani, N.; Armanious, N.; Lue, Y.H.; Swerdloff, R.S.; Baravarian, S.; Adler, A.; Tsang, C.; Jia, Y.; Cui, Y.G.; Wang, X.H.; et al. Integrity of the blood-testis barrier in healthy men after suppression of spermatogenesis with testosterone and levonorgestrel. Hum. Reprod. 2012, 27, 3403–3411. [Google Scholar] [CrossRef][Green Version]
- McCabe, M.J.; Allan, C.M.; Foo, C.F.; Nicholls, P.K.; McTavish, K.J.; Stanton, P.G. Androgen initiates Sertoli cell tight junction formation in the hypogonadal (hpg) mouse. Biol. Reprod. 2012, 87, 38. [Google Scholar] [CrossRef]
- Walker, W.H. Non-classical actions of testosterone and spermatogenesis. Philos. Trans. R Soc. Lond B Biol. Sci. 2010, 365, 1557–1569. [Google Scholar] [CrossRef]
- Bulldan, A.; Dietze, R.; Shihan, M.; Scheiner-Bobis, G. Non-classical testosterone signaling mediated through ZIP9 stimulates claudin expression and tight junction formation in Sertoli cells. Cell Signal. 2016, 28, 1075–1085. [Google Scholar] [CrossRef]
- Möller, M.-L.; Bulldan, A.; Scheiner-Bobis, G. Tetrapeptides Modelled to the Androgen Binding Site of ZIP9 Stimulate Expression of Tight Junction Proteins and Tight Junction Formation in Sertoli Cells. Biology 2022, 11, 55. [Google Scholar] [CrossRef]
- Berg, A.H.; Rice, C.D.; Rahman, M.S.; Dong, J.; Thomas, P. Identification and characterization of membrane androgen receptors in the ZIP9 zinc transporter subfamily: I. Discovery in female atlantic croaker and evidence ZIP9 mediates testosterone-induced apoptosis of ovarian follicle cells. Endocrinology 2014, 155, 4237–4249. [Google Scholar] [CrossRef]
- Thomas, P.; Pang, Y.; Dong, J.; Berg, A.H. Identification and characterization of membrane androgen receptors in the ZIP9 zinc transporter subfamily: II. Role of human ZIP9 in testosterone-induced prostate and breast cancer cell apoptosis. Endocrinology 2014, 155, 4250–4265. [Google Scholar] [CrossRef]
- Bulldan, A.; Malviya, V.N.; Upmanyu, N.; Konrad, L.; Scheiner-Bobis, G. Testosterone/bicalutamide antagonism at the predicted extracellular androgen binding site of ZIP9. Biochim. Biophys Acta 2017, 1864, 2402–2414. [Google Scholar] [CrossRef]
- Kabbesh, H.; Riaz, M.A.; Jensen, A.D.; Scheiner-Bobis, G.; Konrad, L. Long-Term Maintenance of Viable Adult Rat Sertoli Cells Able to Establish Testis Barrier Components and Function in Response to Androgens. Cells 2021, 10, 2405. [Google Scholar] [CrossRef]
- Malviya, V.N.; Bulldan, A.; Wende, R.C.; Kabbesh, H.; Moller, M.L.; Schreiner, P.R.; Scheiner-Bobis, G. The Effects of Tetrapeptides Designed to Fit the Androgen Binding Site of ZIP9 on Myogenic and Osteogenic Cells. Biology 2021, 11, 19. [Google Scholar] [CrossRef]
- Mruk, D.D.; Cheng, C.Y. Enhanced chemiluminescence (ECL) for routine immunoblotting: An inexpensive alternative to commercially available kits. Spermatogenesis 2011, 1, 121–122. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Zhou, X. Roles of androgen receptor in male and female reproduction: Lessons from global and cell-specific androgen receptor knockout (ARKO) mice. J. Androl. 2010, 31, 235–243. [Google Scholar] [CrossRef]
- Davey, R.A.; Grossmann, M. Androgen Receptor Structure, Function and Biology: From Bench to Bedside. Clin. Biochemist. Rev. 2016, 37, 3–15. [Google Scholar]
- Gaetjens, E.; Pertschuk, L.P. Synthesis of fluorescein labelled steroid hormone-albumin conjugates for the fluorescent histochemical detection of hormone receptors. J. Steroid. Biochem. 1980, 13, 1001–1003. [Google Scholar] [CrossRef]
- Benten, W.P.; Lieberherr, M.; Giese, G.; Wrehlke, C.; Stamm, O.; Sekeris, C.E.; Mossmann, H.; Wunderlich, F. Functional testosterone receptors in plasma membranes of T cells. FASEB J. 1999, 13, 123–133. [Google Scholar] [CrossRef]
- Shihan, M.; Bulldan, A.; Scheiner-Bobis, G. Non-classical testosterone signaling is mediated by a G-protein-coupled receptor interacting with Gnalpha11. Biochim. Biophys Acta 2014, 1843, 1172–1181. [Google Scholar] [CrossRef]
- Shihan, M.; Chan, K.H.; Konrad, L.; Scheiner-Bobis, G. Non-classical testosterone signaling in spermatogenic GC-2 cells is mediated through ZIP9 interacting with Gnalpha11. Cell Signal. 2015, 27, 2077–2086. [Google Scholar] [CrossRef]
- Cheng, J.; Watkins, S.C.; Walker, W.H. Testosterone activates mitogen-activated protein kinase via Src kinase and the epidermal growth factor receptor in sertoli cells. Endocrinology 2007, 148, 2066–2074. [Google Scholar] [CrossRef]
- Wen-Hua, Z.; Quirion, R. Insulin-like growth factor-1 (IGF-1) induces the activation/phosphorylation of Akt kinase and cAMP response element-binding protein (CREB) by activating different signaling pathways in PC12 cells. BMC Neurosci. 2006, 7, 51. [Google Scholar] [CrossRef]
- Stevenson, B.R.; Siliciano, J.D.; Mooseker, M.S.; Goodenough, D.A. Identification of ZO-1: A high molecular weight polypeptide associated with the tight junction (zonula occludens) in a variety of epithelia. J. Cell Biol. 1986, 103, 755–766. [Google Scholar] [CrossRef]
- Muller, S.L.; Portwich, M.; Schmidt, A.; Utepbergenov, D.I.; Huber, O.; Blasig, I.E.; Krause, G. The tight junction protein occludin and the adherens junction protein alpha-catenin share a common interaction mechanism with ZO-1. J. Biol. Chem. 2005, 280, 3747–3756. [Google Scholar] [CrossRef]
- Odenwald, M.A.; Choi, W.; Buckley, A.; Shashikanth, N.; Joseph, N.E.; Wang, Y.; Warren, M.H.; Buschmann, M.M.; Pavlyuk, R.; Hildebrand, J.; et al. ZO-1 interactions with F-actin and occludin direct epithelial polarization and single lumen specification in 3D culture. J. Cell Sci. 2017, 130, 243–259. [Google Scholar] [CrossRef] [PubMed]
- Ebnet, K.; Schulz, C.U.; Meyer Zu Brickwedde, M.K.; Pendl, G.G.; Vestweber, D. Junctional adhesion molecule interacts with the PDZ domain-containing proteins AF-6 and ZO-1. J. Biol. Chem. 2000, 275, 27979–27988. [Google Scholar] [CrossRef] [PubMed]
- Ahn, C.; Shin, D.-H.; Lee, D.; Kang, S.-M.; Seok, J.-H.; Kang, H.Y.; Jeung, E.-B. Expression of claudins, occludin, junction adhesion molecule A and zona occludens 1 in canine organs. Mol. Med. Rep. 2016, 14, 3697–3703. [Google Scholar] [CrossRef] [PubMed]
- Gye, M.C. Expression of claudin-1 in mouse testis. Arch. Androl. 2003, 49, 271–279. [Google Scholar] [CrossRef]
- Lui, W.Y.; Cheng, C.Y. Transcriptional regulation of cell adhesion at the blood-testis barrier and spermatogenesis in the testis. Adv. Exp. Med. Biol. 2012, 763, 281–294. [Google Scholar] [CrossRef]
- De Gendt, K.; Verhoeven, G.; Amieux, P.S.; Wilkinson, M.F. Genome-wide identification of AR-regulated genes translated in Sertoli cells in vivo using the RiboTag approach. Mol. Endocrinol. 2014, 28, 575–591. [Google Scholar] [CrossRef]
- Landon, G.V.; Pryor, J.P. The blood-testis barrier in men of diverse fertility status: An ultrastructural study. Virchows Arch. A Pathol. Anat. Histol. 1981, 392, 355–364. [Google Scholar] [CrossRef]
- Cavicchia, J.C.; Sacerdote, F.L.; Ortiz, L. The human blood-testis barrier in impaired spermatogenesis. Ultrastruct. Pathol. 1996, 20, 211–218. [Google Scholar] [CrossRef]
- Jiang, X.H.; Bukhari, I.; Zheng, W.; Yin, S.; Wang, Z.; Cooke, H.J.; Shi, Q.H. Blood-testis barrier and spermatogenesis: Lessons from genetically-modified mice. Asian. J. Androl. 2014, 16, 572–580. [Google Scholar] [CrossRef]
- Willems, A.; Batlouni, S.R.; Esnal, A.; Swinnen, J.V.; Saunders, P.T.; Sharpe, R.M.; Franca, L.R.; De Gendt, K.; Verhoeven, G. Selective ablation of the androgen receptor in mouse sertoli cells affects sertoli cell maturation, barrier formation and cytoskeletal development. PLoS ONE 2010, 5, e14168. [Google Scholar] [CrossRef]
- Payne, J.R.; Kotwinski, P.J.; Montgomery, H.E. Cardiac effects of anabolic steroids. Heart 2004, 90, 473–475. [Google Scholar] [CrossRef]
- Nascimento, J.H.; Medei, E. Cardiac effects of anabolic steroids: Hypertrophy, ischemia and electrical remodelling as potential triggers of sudden death. Mini. Rev. Med. Chem. 2011, 11, 425–429. [Google Scholar] [CrossRef]
- Basaria, S. Male hypogonadism. Lancet 2014, 383, 1250–1263. [Google Scholar] [CrossRef]
- Patel, A.S.; Leong, J.Y.; Ramos, L.; Ramasamy, R. Testosterone Is a Contraceptive and Should Not Be Used in Men Who Desire Fertility. World J. Mens. Health 2019, 37, 45–54. [Google Scholar] [CrossRef]
- Morrow, C.M.; Mruk, D.; Cheng, C.Y.; Hess, R.A. Claudin and occludin expression and function in the seminiferous epithelium. Philos. Trans R Soc. Lond B Biol. Sci. 2010, 365, 1679–1696. [Google Scholar] [CrossRef]
- Singh, A.P.; Harada, S.; Mishina, Y. Downstream genes of Sox8 that would affect adult male fertility. Sex Dev. 2009, 3, 16–25. [Google Scholar] [CrossRef]
- Singh, A.P.; Cummings, C.A.; Mishina, Y.; Archer, T.K. SOX8 regulates permeability of the blood-testes barrier that affects adult male fertility in the mouse. Biol. Reprod. 2013, 88, 133. [Google Scholar] [CrossRef]
- Aydin, S.; Billur, D.; Kizil, S.; Ozkavukcu, S.; Topal Celikkan, F.; Aydos, K.; Erdemli, E. Evaluation of blood-testis barrier integrity in terms of adhesion molecules in nonobstructive azoospermia. Andrologia 2020, 52, e13636. [Google Scholar] [CrossRef]
- Ebling, F.J.; Nwagwu, M.O.; Baines, H.; Myers, M.; Kerr, J.B. The hypogonadal (hpg) mouse as a model to investigate the estrogenic regulation of spermatogenesis. Hum. Fertil. (Camb) 2006, 9, 127–135. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kabbesh, H.; Bulldan, A.; Konrad, L.; Scheiner-Bobis, G. The Role of ZIP9 and Androgen Receptor in the Establishment of Tight Junctions between Adult Rat Sertoli Cells. Biology 2022, 11, 668. https://doi.org/10.3390/biology11050668
Kabbesh H, Bulldan A, Konrad L, Scheiner-Bobis G. The Role of ZIP9 and Androgen Receptor in the Establishment of Tight Junctions between Adult Rat Sertoli Cells. Biology. 2022; 11(5):668. https://doi.org/10.3390/biology11050668
Chicago/Turabian StyleKabbesh, Hassan, Ahmed Bulldan, Lutz Konrad, and Georgios Scheiner-Bobis. 2022. "The Role of ZIP9 and Androgen Receptor in the Establishment of Tight Junctions between Adult Rat Sertoli Cells" Biology 11, no. 5: 668. https://doi.org/10.3390/biology11050668
APA StyleKabbesh, H., Bulldan, A., Konrad, L., & Scheiner-Bobis, G. (2022). The Role of ZIP9 and Androgen Receptor in the Establishment of Tight Junctions between Adult Rat Sertoli Cells. Biology, 11(5), 668. https://doi.org/10.3390/biology11050668