


Anthocyanins from Opuntia ficus-indica Modulate Gut Microbiota Composition and Improve Short-Chain Fatty Acid Production


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Animal Experiment
2.3. Animal Behavioral Observation
2.4. Calculation of the Coefficient of Each Tissue Organ
2.5. DNA Extraction
2.6. 16S rDNA Gene Amplification and Sequencing
2.7. Bioinformatics Analysis of Gut Microbiota Profiles
2.8. Detection of Short-Chain Fatty Acids in the Cecum by Gas Chromatography
2.8.1. Treatment of Cecal Contents
2.8.2. Gas Chromatography Conditions
2.9. Statistical Analysis
3. Results
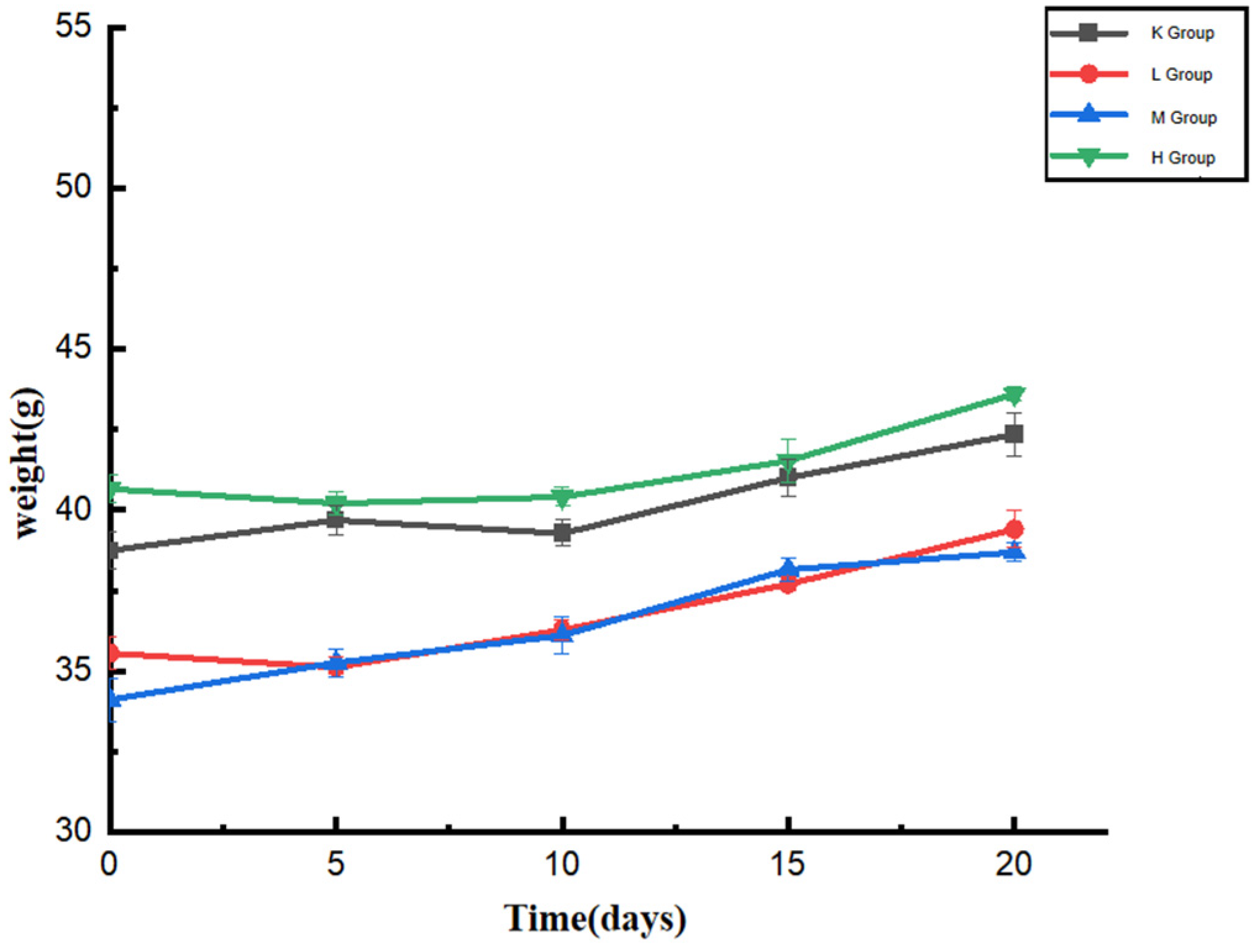
3.1. Effect of Opuntia ficus-indica Anthocyanins on the Body Weight of Mice
3.2. Effects of Opuntia ficus-indica Anthocyanins on the Organ Coefficient of Mice
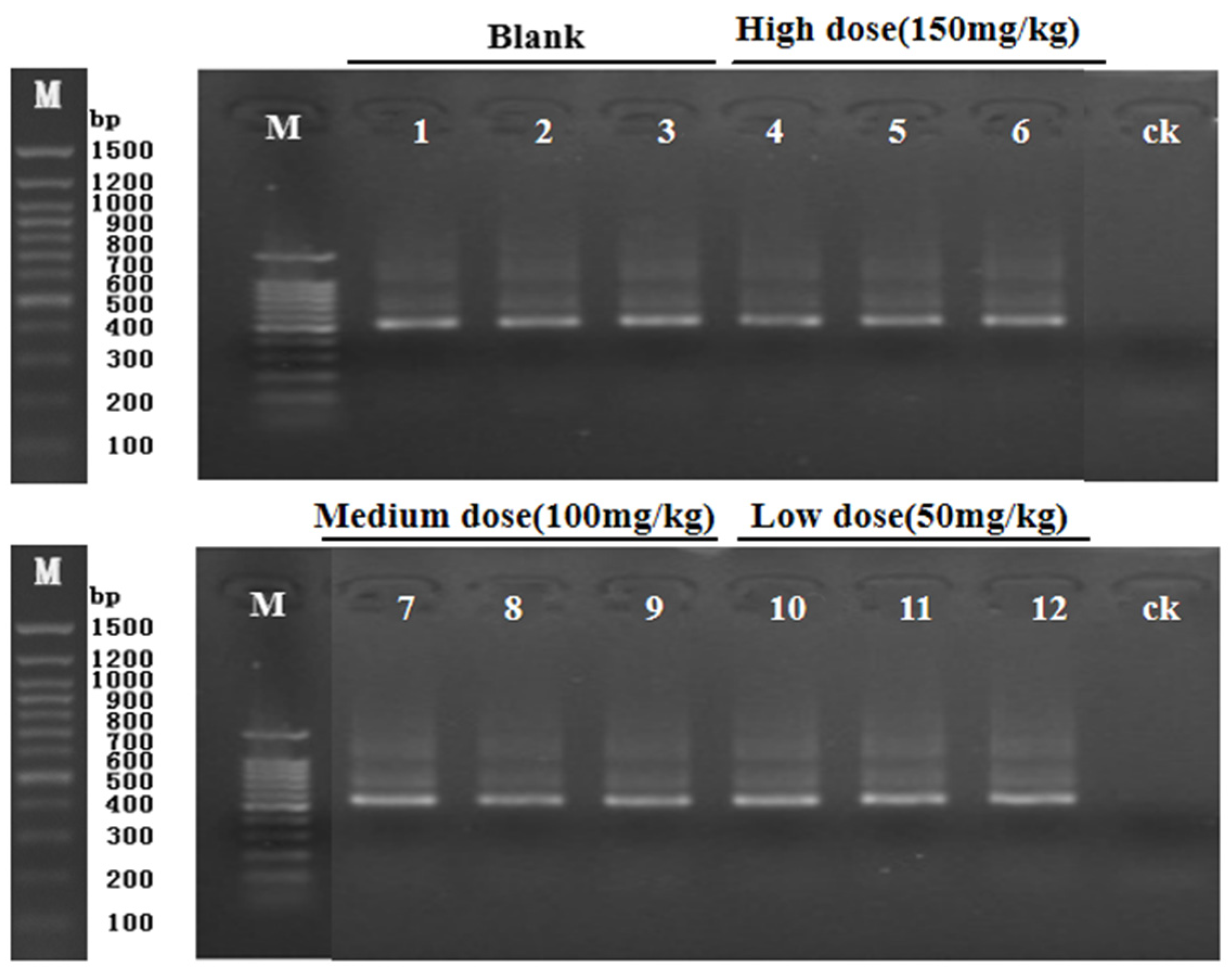
3.3. PCR Amplification Gel Electrophoresis Results
3.4. α-Diversity
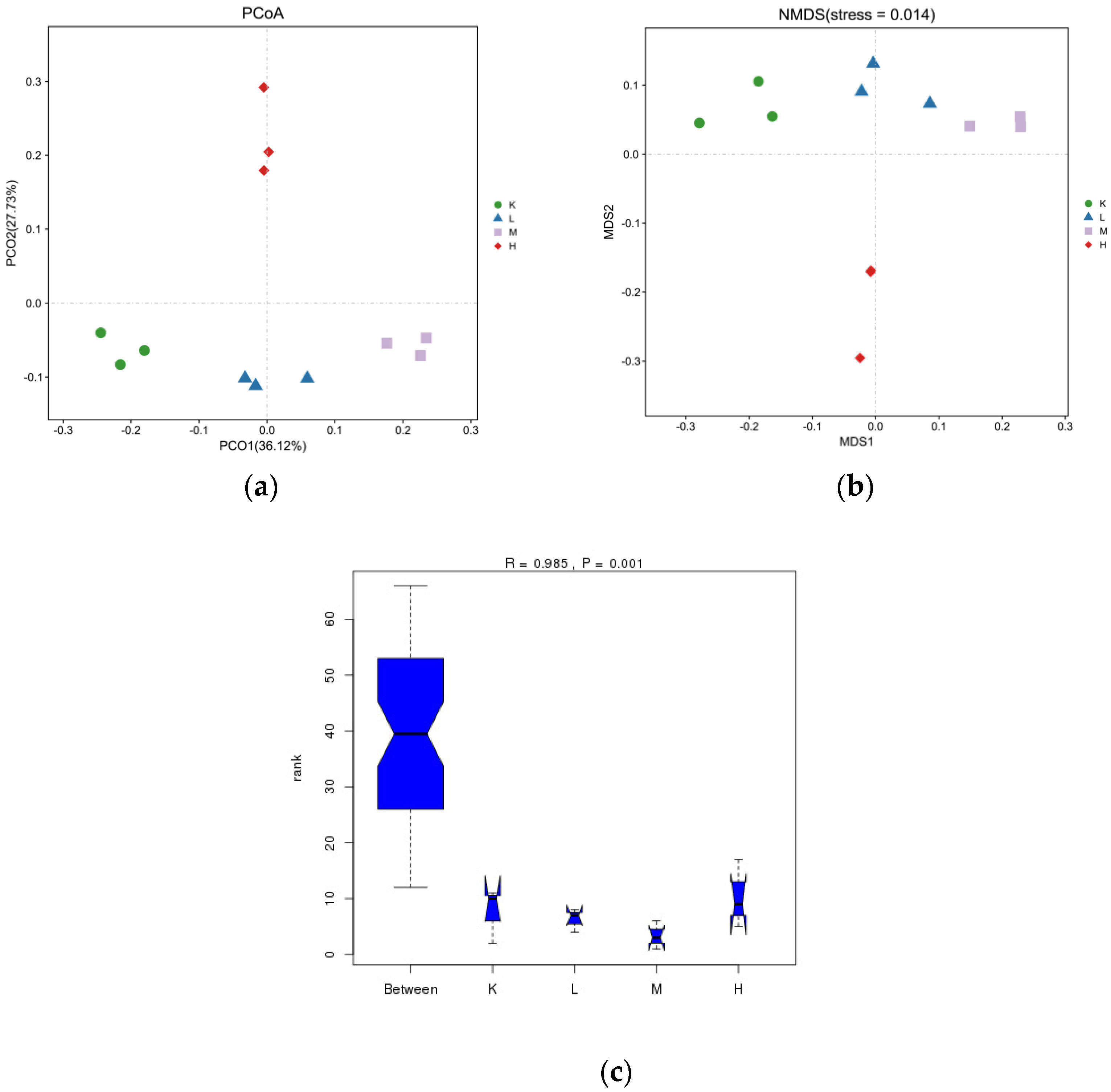
3.5. β-Diversity
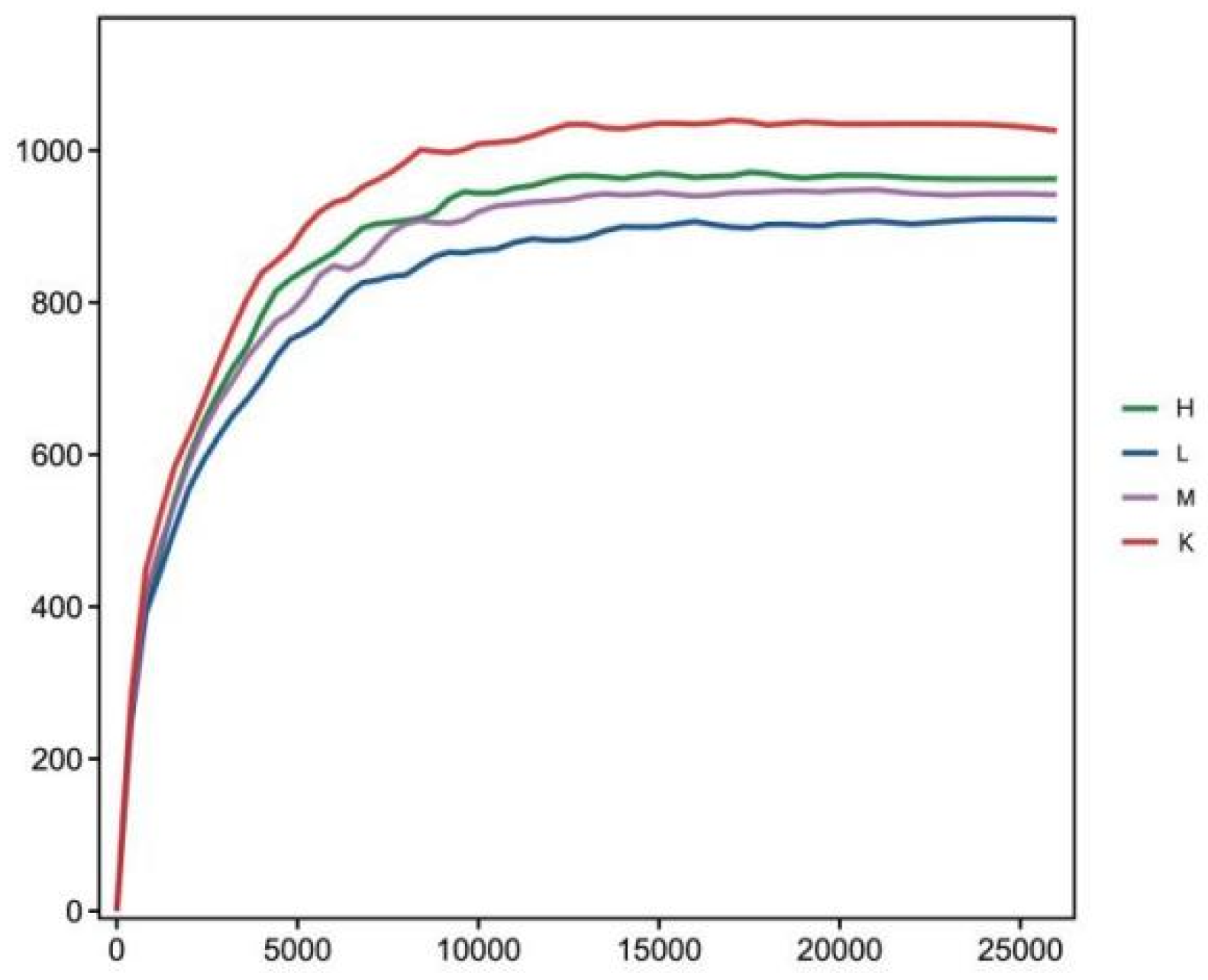
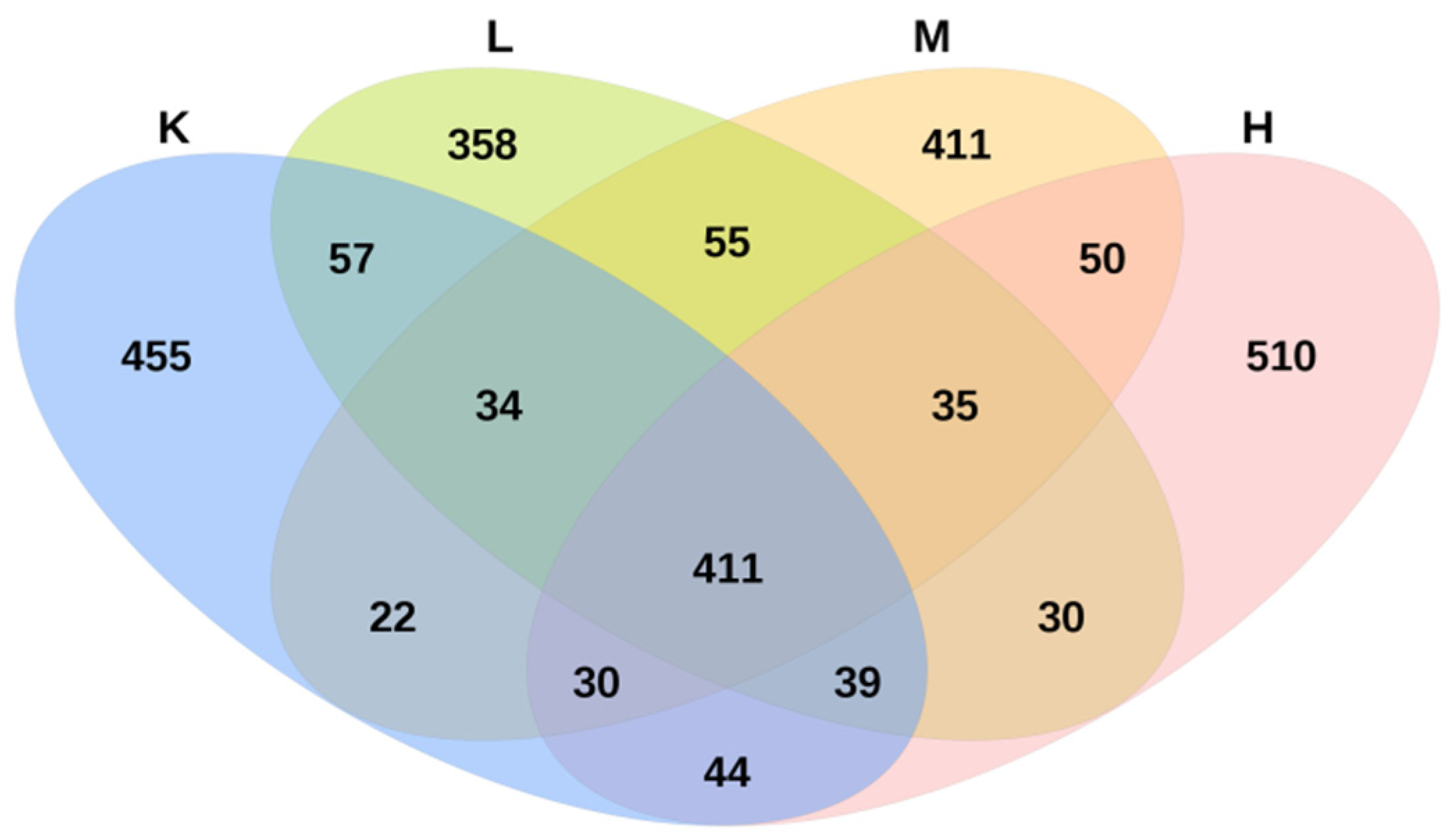
3.6. OTU Numbers
3.7. Effect of Opuntia ficus-indica Anthocyanins on Microbiota Taxonomic Composition
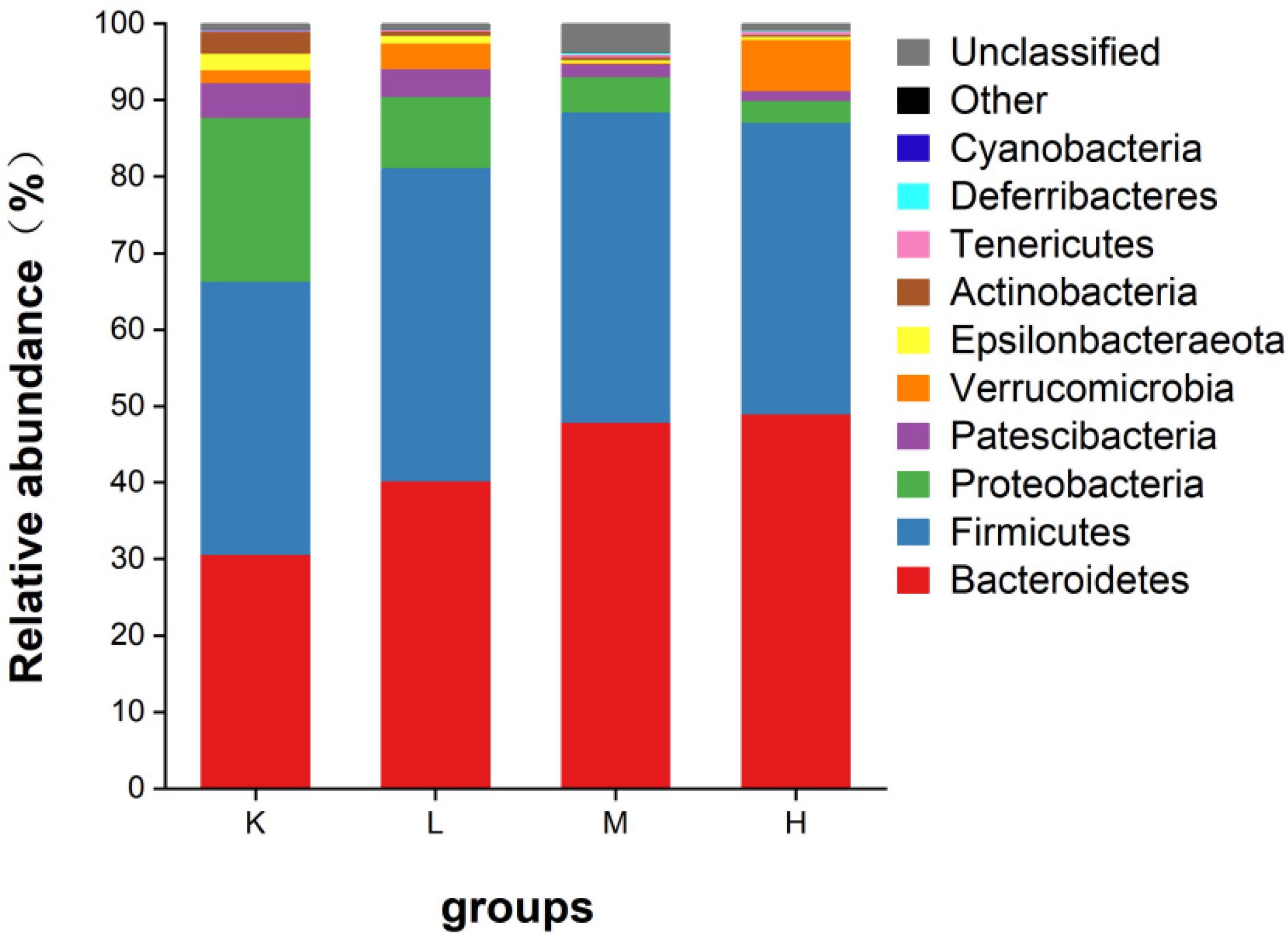
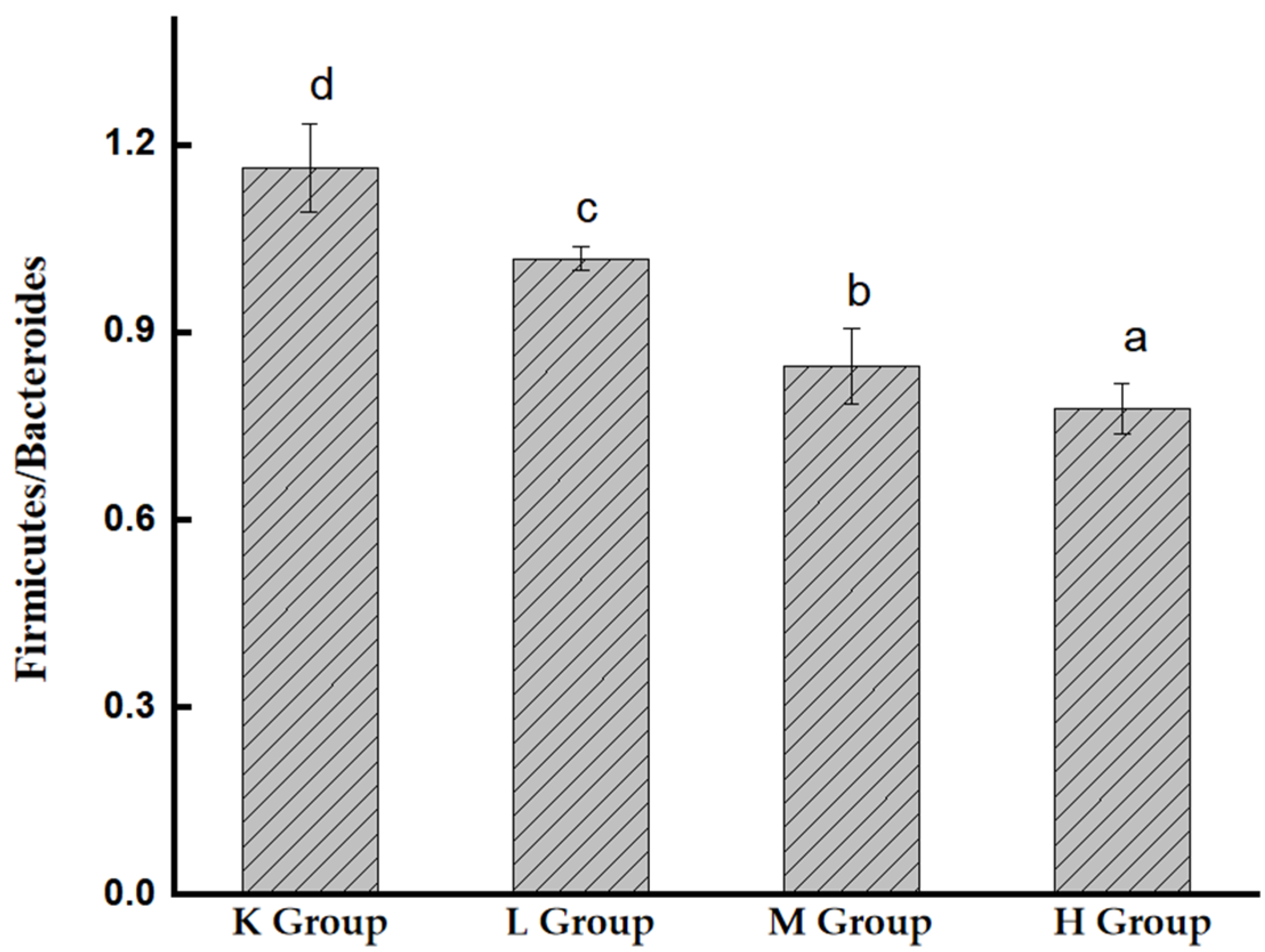
3.7.1. Species Composition Analysis at the Phylum Level
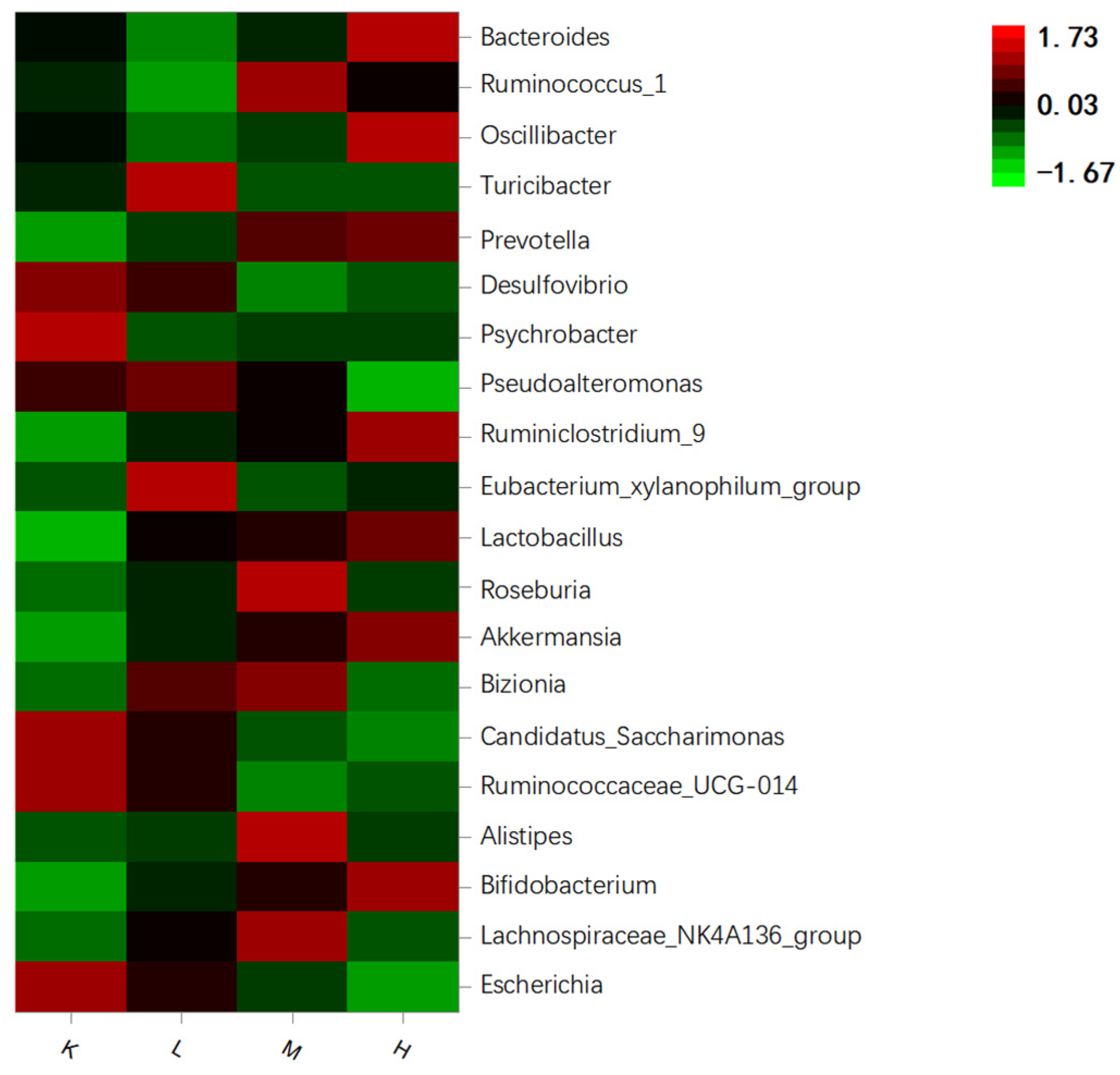
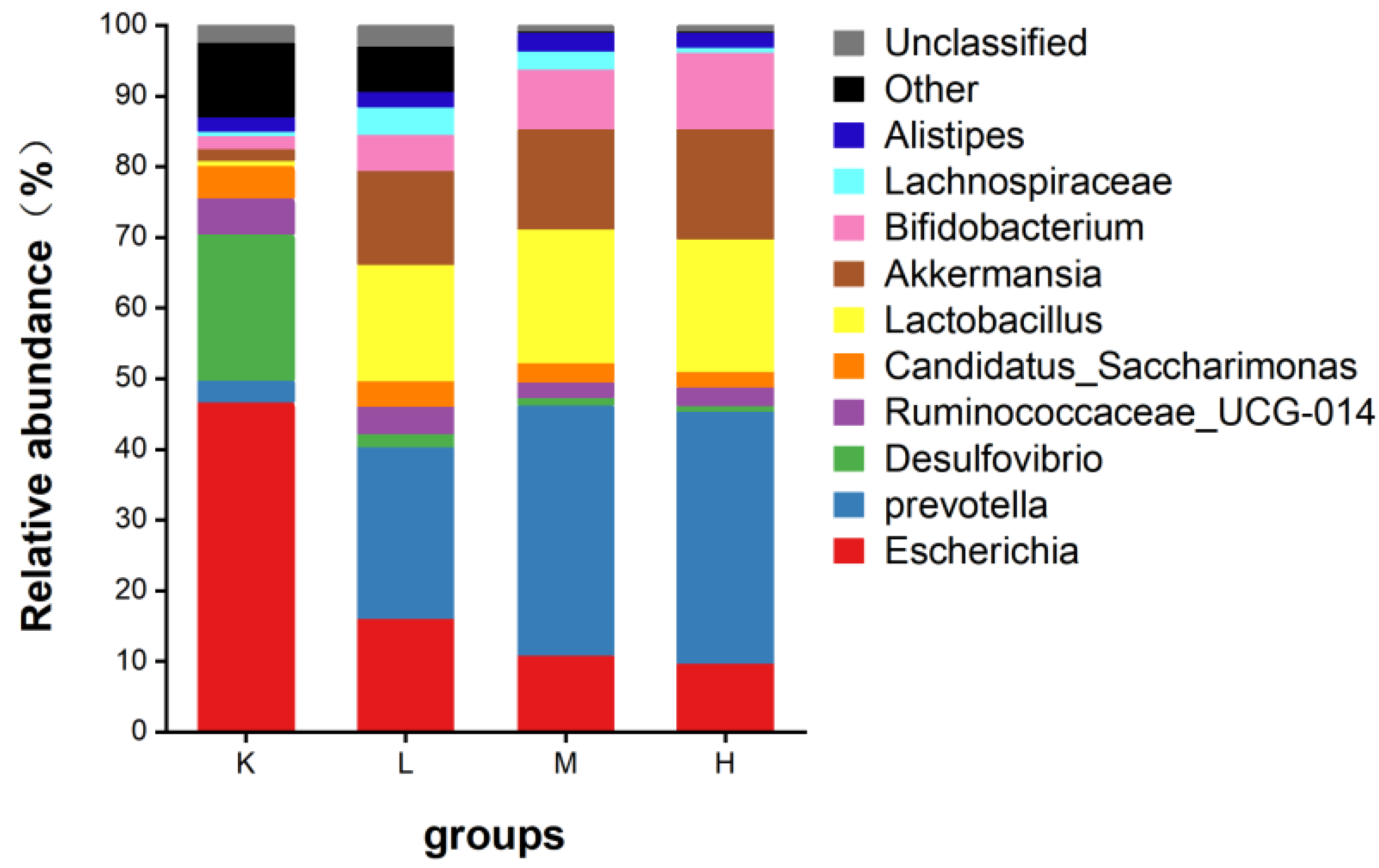
3.7.2. Species Composition Analysis at the Genus Level
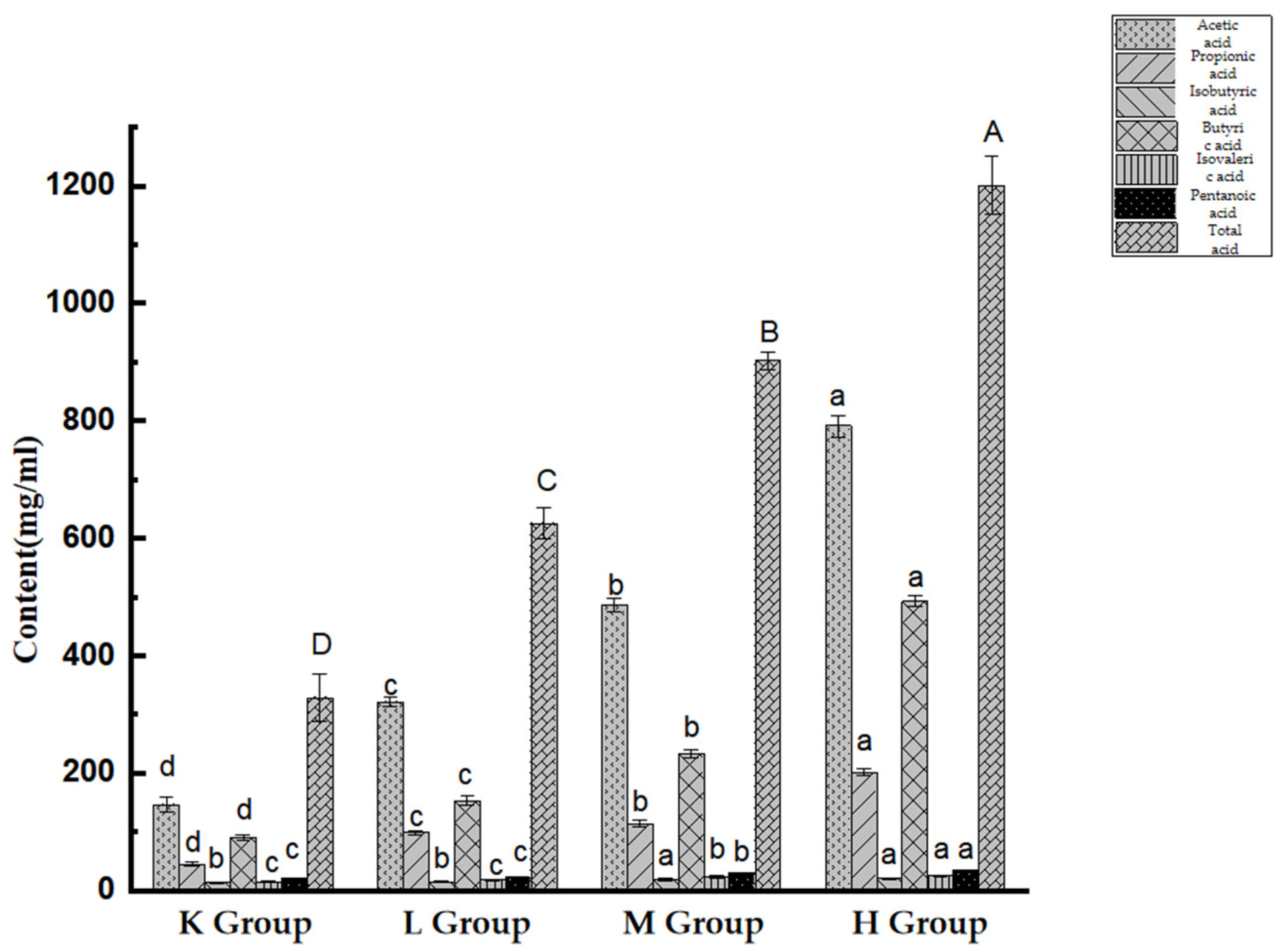
3.8. Effect of Opuntia ficus-indica Anthocyanins: SCFA production
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, H.T. Research progress on the relationship between intestinal microorganisms and human health and its influencing factors. J. Food Saf. Qual. 2022, 13, 175–181. [Google Scholar]
- Walsh, C.J.; Guinane, C.M.; Toole, P.W. Beneficial modulation of the gut microbiota. FEBS Lett. 2016, 588, 4120–4130. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, R.; Kopela, K.; Liu, X. Early Life Microbiota—Impact of Delivery Mode and Infant Feeding. Ann. Nutr. Metab. 2021, 2, 25–38. [Google Scholar]
- Fang, Q.Y.; Hu, J.L.; Nie, Q.X.; Nie, S.P. Effects of polysaccharides on glycometabolism based on gut microbiota alteration. Trends Food Sci. Technol. 2019, 92, 65–70. [Google Scholar] [CrossRef]
- Gao, J. Separation, Purification, Structural Characterization and Physiological Activity of Polysaccharides from Opuntia dillenii Haw Fruits. Master’s Thesis, Hefei University of Technology, Anhui, China, 2015. [Google Scholar]
- Yahia, E.M.; Saenz, C. Cactus pear (Opuntia species). Postharvest Biol. Technol. Trop. Subtrop. Fruits 2011, 19, 290–329. [Google Scholar]
- Ana, F.; Lva, F.; Sonia, N.; Nuno, M. Interaction of anthocyanins with gut microbes. J. Agric. Food Chem. 2014, 62, 231–243. [Google Scholar]
- Tian, L.M.; Tan, Y.S.; Chen, G.W.; Wang, G.; Sun, J.X. Metabolism of anthocyanins and consequent effects on the gut microbiota. Crit. Rev. Food Sci. Nutr. 2018, 42, 102–110. [Google Scholar] [CrossRef]
- Shao, S.; Zhao, J.; Zhang, L.; Wang, R.Y.; Chu, Z.; Zhang, Y. The effects of anthocyanins from Opuntia ficus-indica on gut micobiota and metabolites SCFAs were studied based on in vitro fermentation. Sci. Technol. Food Ind. 2022. [Google Scholar] [CrossRef]
- Zhang, Y.; Shao, S.; Ji, X.H.; Zhang, R.Y.; Zhang, S.W. Study on Purification of Anthocyanins from Opuntia ficus-indica. China Food Addit. 2022, 33, 103–110. [Google Scholar]
- Kong, Q.H.; Zhang, R.F.; You, L.J.; Ma, Y.X.; Liao, L. In vitro fermentation characteristics of polysaccharide from Sargassum fusiforme and its modulation effects on gut microbiota. Food Chem. Toxicol. 2021, 151, 112145. [Google Scholar] [CrossRef]
- Xu, L.L.; Cheng, W.K.; Zhou, X.N. Regulatory effect of Pu-erh tea polysaccharide on short-chain fatty acid metabolism and gut microbiota in mice. Food Ferment. Ind. 2021, 47, 115–122. [Google Scholar]
- Jia, Y.Q.; Ye, F.Y.; Wang, S.; Zhang, Y.Q.; Bai, S.S. Extraction and Determination of Short-chain Fatty Acids in Biological Samples. Res. Explor. Lab. 2012, 31, 262–264. [Google Scholar]
- Peixoto, F.M.; Fernandes, I.; Ana, C.M.S. Simulation of in vitro digestion coupled to gastric and intestinal transport models to estimate absorption of anthocyanins from peel powder of jabuticaba, jamelao and jamb fruits. J. Funct. Foods 2016, 24, 373–381. [Google Scholar] [CrossRef]
- Budiene, J.; Guclu, G.; Oussou, K.F.; Kelebek, H.; Selli, S. Elucidation of Volatiles, Anthocyanins, Antioxidant and Sensory Properties of cv. Caner Pomegranate (Punica granatum L.) Juices Produced from Three Juice Extraction Methods. Foods 2021, 10, 1497. [Google Scholar] [CrossRef]
- Stoner, G.D.; Wang, L.S.; Zikri, N.; Chen, T.; Stephen, S.H. Cancer prevention with freeze-dried berries and berry components. Semin. Cancer Biol. 2007, 17, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Leichtweis, M.G.; Pereira, C.; Prleto, M.A.; Maria, F.B.; Ilton, J.B. Ultrasound as a Rapid and Low-Cost Extraction Procedure to Obtain Anthocyanin-Based Colorants from Prunus spinosa L. Fruit Epicarp: Comparative Study with Conventional Heat-Based Extraction. Molecules 2019, 24, 573. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Mazza, G. Inhibitory effects of anthocyanins and other phenolic compounds on nitric oxide production in LPS/IFN-gamma-activated RAW 264.7 macrophages. J. Agric. Food Chem. 2002, 50, 850–857. [Google Scholar] [CrossRef]
- Li, A.; Xiao, R.S.; He, S.J.; An, X.Y.; He, Y. Research Advances of Purple Sweet Potato Anthocyanins: Extraction, Identification, Stability, Bioactivity, Application, and Biotransformation. Molecules 2019, 24, 3816. [Google Scholar] [CrossRef]
- Chao, Z. Preparation of Sodium Alginate-Anthocyanin Self-Assembled Particles and Its Improvement of Food Allergy in Mice. Master’s Thesis, Jimei University, Fujian, China, 2020. [Google Scholar]
- Bai, W.B.; Zhu, C.J.; Hu, Y.F.; Jiao, R.; Wu, S.; Sun, J.X. Advance in Molecular Mechanism of Nutrition Interventions of Anthocyanins for Chronic Disease. J. Food Sci. Biotechnol. 2016, 35, 1009–1019. [Google Scholar]
- Peng, Y.J.; Yan, Y.M.; Wang, P.; Dong, W.; Huang, K.Y. Effects of long-term intake of anthocyanins from Lycium ruthenicum Murray on the organism health and gut microbiota in vivo. Food Res. Int. 2020, 130, 108952. [Google Scholar] [CrossRef]
- Yu, W.C.; Gao, J.; Hao, R.B.; Yang, J.; Wei, J. Effects of simulated digestion on black chokeberry (Aronia melanocarpa (Michx.) Elliot) anthocyanins and intestinal flora. J. Food Sci. Technol. 2020, 58, 1511–1523. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.L. Study on the interaction between acylated purple sweet potato anthocyanins and intestinal bacteria. Ph.D. Thesis, Tianjin University of Science & Technology, Tianjin, China, 2018. [Google Scholar]
- Xie, S.Z.; Liu, B.; Ye, H.Y.; Li, Q.M.; Pan, X.Q. Dendrobium huoshanense polysaccharide regionally regulates intestinal mucosal barrier function and intestinal microbiota in mice. Carbohydr. Polym. 2019, 206, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Ou, Y.W.; Zhou, Q.; Zhang, Y.; Hu, X.S. Nutrient Uptake of Anthocyanins and Their Impacts on Colon Cancer. Food Sci. 2017, 38, 301–305. [Google Scholar]
- Wang, W.; Liu, F.; Xu, C.; Liu, Z.J.; Ma, J.G.; Gu, L.Y.; Jiang, Z.M.; Hou, J.C. Lactobacillus plantarum 69-2 combined with galacto-oligosaccharides alleviates d-galactose-induced aging by regulating the AMPK/SIRT1 signaling pathway and gut microbiota in mice. J. Agric. Food Chem. 2021, 69, 2745–2757. [Google Scholar] [CrossRef]
- Blesso, C.N. Dietary Anthocyanins and Human Health. Nutrients 2019, 11, 2107. [Google Scholar] [CrossRef]
- Gowd, V.; Tao, B.; Wei, C. Antioxidant potential and phenolic profile of blackberry anthocyanin extract followed by human gut microbiota fermentation. Food Res. Int. 2019, 120, 523–533. [Google Scholar] [CrossRef]
- Tang, C.; Sun, J.; Zhou, B.; Jin, C.H.; Liu, J. Effects of polysaccharides from purple sweet potatoes on immune response and gut microbiota composition in normal and cyclophosphamide treated mice. Food Funct. 2018, 9, 937–950. [Google Scholar] [CrossRef]
- Wang, M.X.; Chen, Y.X.; Wang, Y.Y.; Li, Y.; Zheng, H.H. The effect of probiotics and polysaccharides on the gut microbiota composition and function of weaned rats. Food Funct. 2018, 9, 1864–1877. [Google Scholar] [CrossRef]
- Foley, M.H.; Cockburn, D.W.; Koropatkin, N.M. The Sus operon: A model system for starch uptake by the human gut Bacteroidetes. Cell. Mol. Life Sci. 2016, 73, 2603–2617. [Google Scholar] [CrossRef]
- Ganesan, K.; Chung, S.K.; Vanamala, J.; Xu, B.J. Causal Relationship Between Diet-induced Gut Microbiota Changes and Diabetes: A Novel Strategy to Transplant Faecalibacterium prausnitzii in Preventing Diabetes. Int. J. Mol. Sci. 2018, 19, 3720. [Google Scholar] [CrossRef]
- Liu, L.; Wang, Q.; Wu, X.; Qi, H.M.; Ranjit, D. Vancomycin exposure caused opportunistic pathogens bloom in intestinal microbiome by simulator of the human intestinal microbial ecosystem (SHIME). Environ. Pollut. 2020, 265, 114399. [Google Scholar] [CrossRef] [PubMed]
- Filoppo, C.; Cavalieri, D.; Paola, M.; Ramazzotti, M.; Poullet, J.B. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [PubMed]
- Jian, H.L.; Wang, J.H.; Zou, Y.H.; Erika, K.; Han, Y.Z.; Ning, M.; Zhen, Y.C. Blueberry and cranberry anthocyanin extracts reduce bodyweight and modulate gut microbiota in C57BL/6J mice fed with a high-fat diet. Eur. J. Nutr. 2021, 60, 2735–2746. [Google Scholar]
- Wang, W.; Xu, C.; Liu, Z.J.; Gu, L.Y.; Ma, J.G.; Hou, J.C.; Jiang, Z.M. Physicochemical properties and bioactivity of polysaccharides from Isaria cicadae Miquel with different extraction processes: Effects on gut microbiota and immune response in mice. Food Funct. 2022, 13, 9268–9284. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, D.; Ji, Y.L.; Liu, Y.J.; Xu, L.; Guo, Y.T. Dietary Supplementation of Black Rice Anthocyanin Extract Regulates Cholesterol Metabolism and Improves Gut Microbiota Dysbiosis in C57BL/6J Mice Fed a High-Fat and Cholesterol Diet. Mol. Nutr. Food Res. 2020, 64, 876–883. [Google Scholar] [CrossRef] [PubMed]
- Muriel, D.; Clara, B.; Willem, M. Akkermansia muciniphila and its role in regulating host functions. Microb. Pathog. 2017, 106, 171–181. [Google Scholar]
- Plovier, H.; Everard, A.; Druart, C.; Depommier, C.; Van Hul, M. A purified membrane protein from Akkermansia muciniphila or the pasteurized bacterium improves metabolism in obese and diabetic mice. Nat. Med. 2016, 23, 107. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.B.; Zhang, W.; Wu, Y.X.; Zhang, X.L.; Li, C.J.; Wang, J.Z.; Yan, B. Mitigation of Obesity-Related Systemic Low-Grade Inflammation and Gut Microbial Dysbiosis in Mice with Nanosilver Supplement. ACS Appl. Bio Mater. 2021, 4, 2570–2582. [Google Scholar] [CrossRef]
- Lloyd, J.R.; Thomas, G.H.; Finlay, J.A.; Cole, J.A.; Macaskie, L.E. Microbial reduction of technetium by Escherichia coli and Desulfovibrio desulfuricans: Enhancement via the use of high-activity strains and effect of process parameters. Biotechnol. Bioeng. 2015, 66, 122–130. [Google Scholar] [CrossRef]
- Sawin, E.A.; De Wolfe, T.J.; Aktas, B.; Stroup, B.M.; Murali, S.G.; Steele, J.L.; Ney, D.M. Glycomacropeptide is a prebiotic that reduces Desulfovibrio bacteria, increases cecal short-chain fatty acids, and is anti-inflammatory in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, 590–601. [Google Scholar] [CrossRef]
- Wen, C.Y. Study and Development on Functional Ingredients of Cactus Fruit. Master’s Thesis, Hainan University, Hainan, China, 2020. [Google Scholar]
- Canfora, E.E.; Meex, R.C.R.; Venema, K.; Blaak, E.E. Gut microbial metabolites in obesity, NAFLD and T2DM. Nat. Rev. Endocrinol. 2019, 15, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Ikuo, K. Host energy regulation via SCFAs receptors, as dietary nutrition sensors, by gut microbiota. Yakugaku Zasshi 2014, 134, 1037–1042. [Google Scholar]
- Ana, M.V.; Jens, W.; Eran, S.; Tim, D.S. Role of the gut microbiota in nutrition and health. BMJ 2018, 361, k2179. [Google Scholar]
- Sa’ad, H.A.; Maikel, P.P.; Roelofsen, H.; Roel, J.V.; Roel, V. Biological efects of propionic acid in humans; metabolism, potential applications and underlying mechanisms. Biochim. Biophys. Acta 2010, 1801, 1175–1183. [Google Scholar]
- Hirschberg, S.; Gisevius, B.; Duscha, A.; Haghikia, A. Implications of diet and the gut microbiome in neuroinfammatory and neurodegenerative diseases. Int. J. Mol. Sci. 2019, 20, 3109. [Google Scholar] [CrossRef]
- Jian, G.L.; Lu, W.H.; Cui, W.; Xue, Y.J.; Xue, M.Q.; Chang, X.W. Short term intrarectal administration of sodium propionate induces antidepressant-like efects in rats exposed to chronic unpredictable mild stress. Front. Psychiatry 2018, 9, 454. [Google Scholar]
- Gail, A.C.; Bryan, G.; Megan, R.M.; Wei, X.; Daniella, A.; Laura, E.N. Prophylactic tributyrin treatment mitigates chronic-binge ethanol-induced intestinal barrier and liver injury. J Gastroenterol. Hepatol. 2017, 32, 1587–1597. [Google Scholar]
- Sun, J.; Wang, F.; Hong, G.; Pang, M.; Xu, H.; Li, H. Antidepressant-like efects of sodium butyrate and its possible mechanisms of action in mice exposed to chronic unpredictable mild stress. Neurosci. Lett. 2016, 618, 159–166. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Cardiac Coefficient% | Liver Coefficient% | Spleen Coefficient% | Lung Coefficient% | Kidney Coefficient% |
|---|---|---|---|---|---|
| K group | 0.487 ± 0.04 | 4.663 ± 0.38 | 0.303 ± 0.08 | 0.586 ± 0.08 | 1.447 ± 0.19 |
| L group | 0.584 ± 0.07 | 4.661 ± 0.58 | 0.337 ± 0.11 | 0.698 ± 0.12 | 1.599 ± 0.16 |
| M group | 0.529 ± 0.04 | 4.742 ± 0.58 | 0.344 ± 0.06 | 0.744 ± 0.19 | 1.456 ± 0.11 |
| H group | 0.543 ± 0.06 | 4.844 ± 0.46 | 0.387 ± 0.21 | 0.638 ± 0.11 | 1.439 ± 0.18 |
| Group | Chao1 | ACE | Shannon | Simpson |
|---|---|---|---|---|
| K group | 1023.41 ± 43.58 | 2052.29 ± 48.65 | 7.08 ± 0.07 | 0.98 ± 0.004 |
| L group | 905.81 ± 9.51 ** | 929.96 ± 13.27 ** | 7.53 ± 0.04 * | 1.53 ± 0.001 * |
| M group | 938.84 ± 37.39 ** | 967.38 ± 40.00 ** | 7.82 ± 0.03 * | 1.95 ± 0.001 * |
| H group | 958.26 ± 32.89 ** | 987.78 ± 32.94 ** | 8.31 ± 0.18 * | 2.42 ± 0.003 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Chang, H.; Shao, S.; Zhao, L.; Zhang, R.; Zhang, S. Anthocyanins from Opuntia ficus-indica Modulate Gut Microbiota Composition and Improve Short-Chain Fatty Acid Production. Biology 2022, 11, 1505. https://doi.org/10.3390/biology11101505
Zhang Y, Chang H, Shao S, Zhao L, Zhang R, Zhang S. Anthocyanins from Opuntia ficus-indica Modulate Gut Microbiota Composition and Improve Short-Chain Fatty Acid Production. Biology. 2022; 11(10):1505. https://doi.org/10.3390/biology11101505
Chicago/Turabian StyleZhang, Yun, Huan Chang, Shuai Shao, Lin Zhao, Ruiying Zhang, and Shouwen Zhang. 2022. "Anthocyanins from Opuntia ficus-indica Modulate Gut Microbiota Composition and Improve Short-Chain Fatty Acid Production" Biology 11, no. 10: 1505. https://doi.org/10.3390/biology11101505
APA StyleZhang, Y., Chang, H., Shao, S., Zhao, L., Zhang, R., & Zhang, S. (2022). Anthocyanins from Opuntia ficus-indica Modulate Gut Microbiota Composition and Improve Short-Chain Fatty Acid Production. Biology, 11(10), 1505. https://doi.org/10.3390/biology11101505