
High DAPK1 Expression Promotes Tumor Metastasis of Gastric Cancer
_Kwok.png)
 and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Mining from Public Databases
2.2. Cells Culture and Transfection
2.3. Cell Migration Assay
2.4. Cell Invasion Assay
2.5. CCK-8 Assay
2.6. WGCNA (Weighted Gene Co-Expression Network Analysis) Analysis
2.7. Risk Score Module Construction
2.8. Functional Enrichment Analysis
2.9. Correlogram Analyses
2.10. Statistical Analysis
3. Results
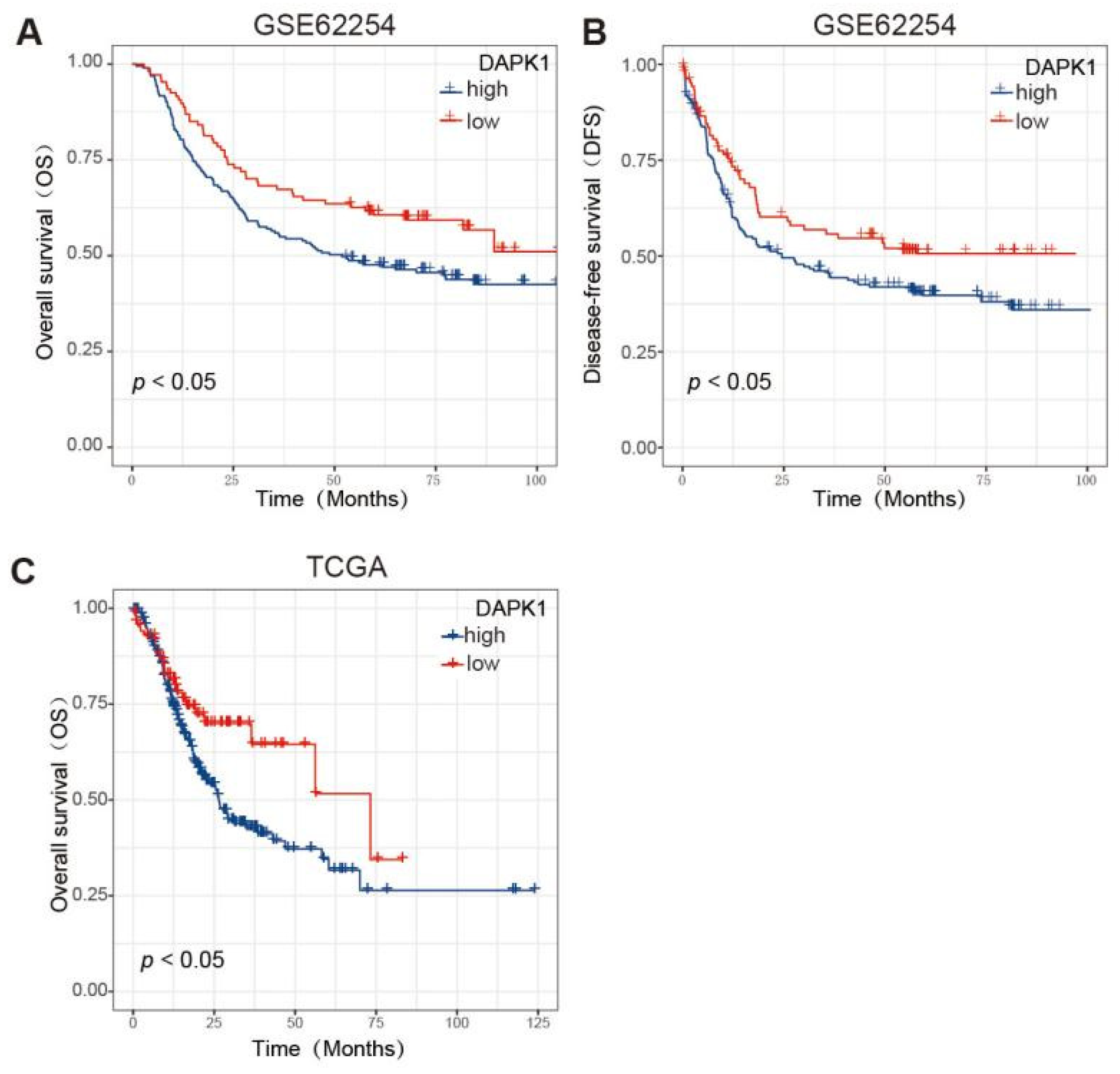
3.1. The Expression Pattern and Prognosis Analysis of DAPK1 in GC Patients
3.2. The Relationships of DAPK1 Expression with DNA Methylation and Transcription Factors
3.3. DAPK1 Displays Oncogenic Features in GC Cells
3.4. Construction of a Co-Expression Network for Weighted Genes of DAPK1 in GC
3.5. Prognosis Model of DAPK1-Related Genes Constructed
3.6. Construction of a Clinical Prognostic Prediction Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Ang, T.L.; Fock, K.M. Clinical epidemiology of gastric cancer. Singap. Med. J. 2014, 55, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Lin, Y.; Zhong, W.; Jiang, Y.; Lin, Y. Regulatory Non-coding RNAs for Death Associated Protein Kinase Family. Front. Mol. Biosci. 2021, 8, 649100. [Google Scholar] [CrossRef]
- Farag, A.K.; Roh, E.J. Death-associated protein kinase (DAPK) family modulators: Current and future therapeutic outcomes. Med. Res. Rev. 2019, 39, 349–385. [Google Scholar] [CrossRef]
- Song, Z.; Li, Z.; Han, W.; Zhu, C.; Lou, N.; Li, X.; Luo, G.; Peng, S.; Li, G.; Zhao, Y.; et al. Low DAPK1 expression correlates with poor prognosis and sunitinib resistance in clear cell renal cell carcinoma. Aging 2020, 13, 1842–1858. [Google Scholar] [CrossRef]
- Li, Y.; Zhu, M.; Zhang, X.; Cheng, D.; Ma, X. Clinical significance of DAPK promoter hypermethylation in lung cancer: A meta-analysis. Drug Des. Dev. Ther. 2015, 9, 1785–1796. [Google Scholar] [CrossRef][Green Version]
- Ivanovska, J.; Zlobec, I.; Forster, S.; Karamitopoulou, E.; Dawson, H.; Koelzer, V.H.; Agaimy, A.; Garreis, F.; Söder, S.; Laqua, W.; et al. Correction: DAPK loss in colon cancer tumor buds: Implications for migration capacity of disseminating tumor cells. Oncotarget 2017, 8, 18615–18616. [Google Scholar] [CrossRef]
- Peng, D.F.; Kanai, Y.; Sawada, M.; Ushijima, S.; Hiraoka, N.; Kitazawa, S.; Hirohashi, S. DNA methylation of multiple tumor-related genes in association with overexpression of DNA methyltransferase 1 (DNMT1) during multistage carcinogenesis of the pancreas. Carcinogenesis 2006, 27, 1160–1168. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, C.; Li, K.; Ye, Y.; Shen, A.; Guo, L.; Chen, P.; Meng, C.; Wang, Q.; Yang, X.; et al. Death-associated protein kinase 1 suppresses hepatocellular carcinoma cell migration and invasion by upregulation of DEAD-box helicase 20. Cancer Sci. 2020, 111, 2803–2813. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Ye, Y.; Lin, R.; Weng, S.; Cai, F.; Zou, M.; Niu, H.; Ge, L.; Lin, Y. Analysis of the expression, function, prognosis and co-expression genes of DDX20 in gastric cancer. Comput. Struct. Biotechnol. J. 2020, 18, 2453–2462. [Google Scholar] [CrossRef]
- Guo, Z.; Zhou, C.; Zhou, L.; Wang, Z.; Zhu, X.; Mu, X. Overexpression of DAPK1-mediated inhibition of IKKβ/CSN5/PD-L1 axis enhances natural killer cell killing ability and inhibits tumor immune evasion in gastric cancer. Cell. Immunol. 2022, 372, 104469. [Google Scholar] [CrossRef]
- Wang, Q.S.; Li, F.; Liao, Z.Q.; Li, K.; Yang, X.L.; Lin, Y.Y.; Zhao, Y.L.; Weng, S.Y.; Xia, Y.; Ye, Y.; et al. Low level of Cyclin-D1 correlates with worse prognosis of clear cell renal cell carcinoma patients. Cancer Med. 2019, 8, 4100–4109. [Google Scholar] [CrossRef] [PubMed]
- Niu, H.; Li, F.; Wang, Q.; Ye, Z.; Chen, Q.; Lin, Y. High expression level of MMP9 is associated with poor prognosis in patients with clear cell renal carcinoma. PeerJ 2018, 6, e5050. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, D.S.; Bashel, B.; Balasubramanya, S.A.H.; Creighton, C.J.; Ponce-Rodriguez, I.; Chakravarthi, B.; Varambally, S. UALCAN: A Portal for Facilitating Tumor Subgroup Gene Expression and Survival Analyses. Neoplasia 2017, 19, 649–658. [Google Scholar] [CrossRef]
- Zhang, X.; Ni, Z.; Duan, Z.; Xin, Z.; Wang, H.; Tan, J.; Wang, G.; Li, F. Overexpression of E2F mRNAs associated with gastric cancer progression identified by the transcription factor and miRNA co-regulatory network analysis. PLoS ONE 2015, 10, e0116979. [Google Scholar] [CrossRef]
- Cheng, L.; Zhang, Q.; Yang, S.; Yang, Y.; Zhang, W.; Gao, H.; Deng, X.; Zhang, Q. A 4-gene panel as a marker at chromosome 8q in Asian gastric cancer patients. Genomics 2013, 102, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhang, X.; Chen, L.; Weng, S.; Xia, Y.; Ye, Y.; Li, K.; Liao, Z.; Chen, P.; Alsamman, K.; et al. Regulation of the Expression of DAPK1 by SUMO Pathway. Biomolecules 2019, 9, 151. [Google Scholar] [CrossRef] [PubMed]
- Nong, S.; Wei, Z.; Wang, Z.; Ma, L.; Guan, Y.; Ni, J. Reduced DAPK1 Expression Promotes Stem Cell-Like Characteristics of Prostate Cancer Cells by Activating ZEB1 via Hippo/YAP Signaling Pathway. Stem Cells Dev. 2021, 30, 934–945. [Google Scholar] [CrossRef]
- Wei, Z.; Li, P.; He, R.; Liu, H.; Liu, N.; Xia, Y.; Bi, G.; Du, Q.; Xia, M.; Pei, L.; et al. DAPK1 (death associated protein kinase 1) mediates mTORC1 activation and antiviral activities in CD8+ T cells. Cell. Mol. Immunol. 2021, 18, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Movahhed, P.; Saberiyan, M.; Safi, A.; Arshadi, Z.; Kazerouni, F.; Teimori, H. The impact of DAPK1 and mTORC1 signaling association on autophagy in cancer. Mol. Biol. Rep. 2022, 49, 4959–4964. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.Y.; Chen, P.C.; Zhang, J.L.; Gao, Z.S.; Neves, H.; Zhang, S.D.; Wen, Q.; Chen, W.D.; Kwok, H.F.; Lin, Y. The prognostic significance of DAPK1 in bladder cancer. PLoS ONE 2017, 12, e0175290. [Google Scholar] [CrossRef]
- Steinmann, S.; Kunze, P.; Hampel, C.; Eckstein, M.; Bertram Bramsen, J.; Muenzner, J.K.; Carlé, B.; Ndreshkjana, B.; Kemenes, S.; Gasparini, P.; et al. DAPK1 loss triggers tumor invasion in colorectal tumor cells. Cell Death Dis. 2019, 10, 895. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Wu, X.Y. Expression and clinical significance of DAPK1 and CD147 in esophageal squamous cell carcinoma. Zhonghua Zhong Liu Za Zhi [Chin. J. Oncol.] 2012, 34, 44–48. [Google Scholar]
- Zhao, J.; Zhao, D.; Poage, G.M.; Mazumdar, A.; Zhang, Y.; Hill, J.L.; Hartman, Z.C.; Savage, M.I.; Mills, G.B.; Brown, P.H. Death-associated protein kinase 1 promotes growth of p53-mutant cancers. J. Clin. Investig. 2015, 125, 2707–2720. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Ma, C.; Zhang, Z.; Zeng, S.; Liu, A.; Tang, S.; Ren, Q.; Sun, Y.; Xu, C. DAPK Promoter Methylation and Bladder Cancer Risk: A Systematic Review and Meta-Analysis. PLoS ONE 2016, 11, e0167228. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Yu, T.; Cao, X.; An, Q.; Yang, H. Clinical effect of DAPK promoter methylation in gastric cancer: A systematic meta-analysis. Medicine 2016, 95, e5040. [Google Scholar] [CrossRef] [PubMed]
- Cai, F.; Xiao, X.; Niu, X.; Zhong, Y. Association between promoter methylation of DAPK gene and HNSCC: A meta-analysis. PLoS ONE 2017, 12, e0173194. [Google Scholar] [CrossRef] [PubMed]
- Spisák, S.; Kalmár, A.; Galamb, O.; Wichmann, B.; Sipos, F.; Péterfia, B.; Csabai, I.; Kovalszky, I.; Semsey, S.; Tulassay, Z.; et al. Genome-wide screening of genes regulated by DNA methylation in colon cancer development. PLoS ONE 2012, 7, e46215. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Yu, J.; Zhang, X.; Dai, Z.; Fang, A. CircSEC31A Promotes the Malignant Progression of Non-Small Cell Lung Cancer Through Regulating SEC31A Expression via Sponging miR-376a. Cancer Manag. Res. 2020, 12, 11527–11539. [Google Scholar] [CrossRef]
- Chen, M.; Wei, L.; Law, C.T.; Tsang, F.H.; Shen, J.; Cheng, C.L.; Tsang, L.H.; Ho, D.W.; Chiu, D.K.; Lee, J.M.; et al. RNA N6-methyladenosine methyltransferase-like 3 promotes liver cancer progression through YTHDF2-dependent posttranscriptional silencing of SOCS2. Hepatology 2018, 67, 2254–2270. [Google Scholar] [CrossRef] [PubMed]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for functional genomics data sets—Update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Tang, W.; Xu, Y.; Xu, Y.; Xu, B.; Fu, S.; Wang, Y.; Chen, F.; Chen, Y.; Han, Y.; et al. METTL3/YTHDF2 m6A axis accelerates colorectal carcinogenesis through epigenetically suppressing YPEL5. Mol. Oncol. 2021, 15, 2172–2184. [Google Scholar] [CrossRef] [PubMed]
- Ripke, S.; O’Dushlaine, C.; Chambert, K.; Moran, J.L.; Kähler, A.K.; Akterin, S.; Bergen, S.E.; Collins, A.L.; Crowley, J.J.; Fromer, M.; et al. Genome-wide association analysis identifies 13 new risk loci for schizophrenia. Nat. Genet. 2013, 45, 1150–1159. [Google Scholar] [CrossRef]
- Lencz, T.; Knowles, E.; Davies, G.; Guha, S.; Liewald, D.C.; Starr, J.M.; Djurovic, S.; Melle, I.; Sundet, K.; Christoforou, A.; et al. Molecular genetic evidence for overlap between general cognitive ability and risk for schizophrenia: A report from the Cognitive Genomics consorTium (COGENT). Mol. Psychiatry 2014, 19, 168–174. [Google Scholar] [CrossRef]
- Chang, C.C.; Kuo, H.Y.; Chen, S.Y.; Lin, W.T.; Lu, K.M.; Saito, T.; Liu, F.C. Developmental Characterization of Schizophrenia-Associated Gene Zswim6 in Mouse Forebrain. Front. Neuroanat. 2021, 15, 669631. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | All Patients | Low DAPK1 | High DAPK1 | p Value | |
|---|---|---|---|---|---|
| n = 300 | n = 107 | n = 193 | |||
| No. | (%) | No. | No. | ||
| Age | 0.1168 | ||||
| ≤50 | 41 | 13.7 | 10 | 31 | |
| >50 | 259 | 86.3 | 97 | 162 | |
| Sex | 0.0110 | ||||
| Male | 199 | 66.3 | 81 | 118 | |
| Female | 101 | 33.7 | 26 | 75 | |
| Tumor Stage | 0.0105 | ||||
| I + II | 127 | 42.3 | 56 | 71 | |
| III + IV | 173 | 57.7 | 51 | 122 | |
| Pathology | 0.2796 | ||||
| Diffuse | 135 | 45 | 42 | 93 | |
| Intestinal | 146 | 48.7 | 58 | 88 | |
| Mixed | 17 | 5.6 | 5 | 12 |
| Variables | OS | DFS | ||
|---|---|---|---|---|
| HR (95% CI) | p | HR (95% CI) | p | |
| Age (years) | ||||
| ≤50 | Reference | Reference | ||
| >50 | 1.24 (0.8–1.92) | 0.330 | 1.19 (0.78–1.83) | 0.420 |
| Sex | ||||
| Female | Reference | Reference | ||
| Male | 1.11(0.79–1.55) | 0.560 | 1.02(0.74–1.42) | 0.900 |
| Tumor Stage | ||||
| I + II | Reference | Reference | ||
| III + IV | 3.47 (2.39–5.05) | <0.001 | 3.46 (2.39–5.01) | <0.001 |
| Pathology | ||||
| Diffuse | Reference | Reference | ||
| Intestinal | 0.6 (0.43–0.83) | 0.002 | 0.68 (0.49–0.94) | 0.019 |
| DAPK1 expression | ||||
| Low | Reference | Reference | ||
| High | 1.49 (1.05–2.11) | 0.025 | 1.41 (1.00–1.99) | 0.046 |
| Variables | OS | DFS | ||
|---|---|---|---|---|
| HR (95% CI) | p | HR (95% CI) | p | |
| Age (years) | ||||
| ≤50 | NA | NA | ||
| >50 | ||||
| Sex | ||||
| Female | NA | NA | ||
| Male | ||||
| Tumor Stage | ||||
| I + II | Reference | Reference | ||
| III + IV | 3.52 (2.33–5.31) | <0.001 | 3.59 (2.40–5.38) | <0.001 |
| Pathology | ||||
| diffuse | Reference | Reference | ||
| intestinal | 0.79 (0.56–1.12) | 0.182 | 0.88 (0.62–1.25) | 0.471 |
| DAPK1 expression | ||||
| Low | Reference | Reference | ||
| High | 1.25 (0.87–1.81) | 0.23 | 1.29 (0.89–1.86) | 0.179 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Weng, S.; Sun, Y.; Lin, Y.; Zhong, W.; Kwok, H.F.; Lin, Y. High DAPK1 Expression Promotes Tumor Metastasis of Gastric Cancer. Biology 2022, 11, 1488. https://doi.org/10.3390/biology11101488
Wang Q, Weng S, Sun Y, Lin Y, Zhong W, Kwok HF, Lin Y. High DAPK1 Expression Promotes Tumor Metastasis of Gastric Cancer. Biology. 2022; 11(10):1488. https://doi.org/10.3390/biology11101488
Chicago/Turabian StyleWang, Qingshui, Shuyun Weng, Yuqin Sun, Youyu Lin, Wenting Zhong, Hang Fai Kwok, and Yao Lin. 2022. "High DAPK1 Expression Promotes Tumor Metastasis of Gastric Cancer" Biology 11, no. 10: 1488. https://doi.org/10.3390/biology11101488
APA StyleWang, Q., Weng, S., Sun, Y., Lin, Y., Zhong, W., Kwok, H. F., & Lin, Y. (2022). High DAPK1 Expression Promotes Tumor Metastasis of Gastric Cancer. Biology, 11(10), 1488. https://doi.org/10.3390/biology11101488