
Early Starvation Contributes to the Adaptive Capacity of Corythucha marmorata (Uhler), an Emerging Pest in China


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Insects and Host Plants
2.2. Observation of Development on Four Host Plants of C. marmorata
2.3. Measurement of Early Starvation on Development of C. marmorata
2.4. Determination of the Supercooling and Freezing Point
2.5. Data Analysis
3. Results
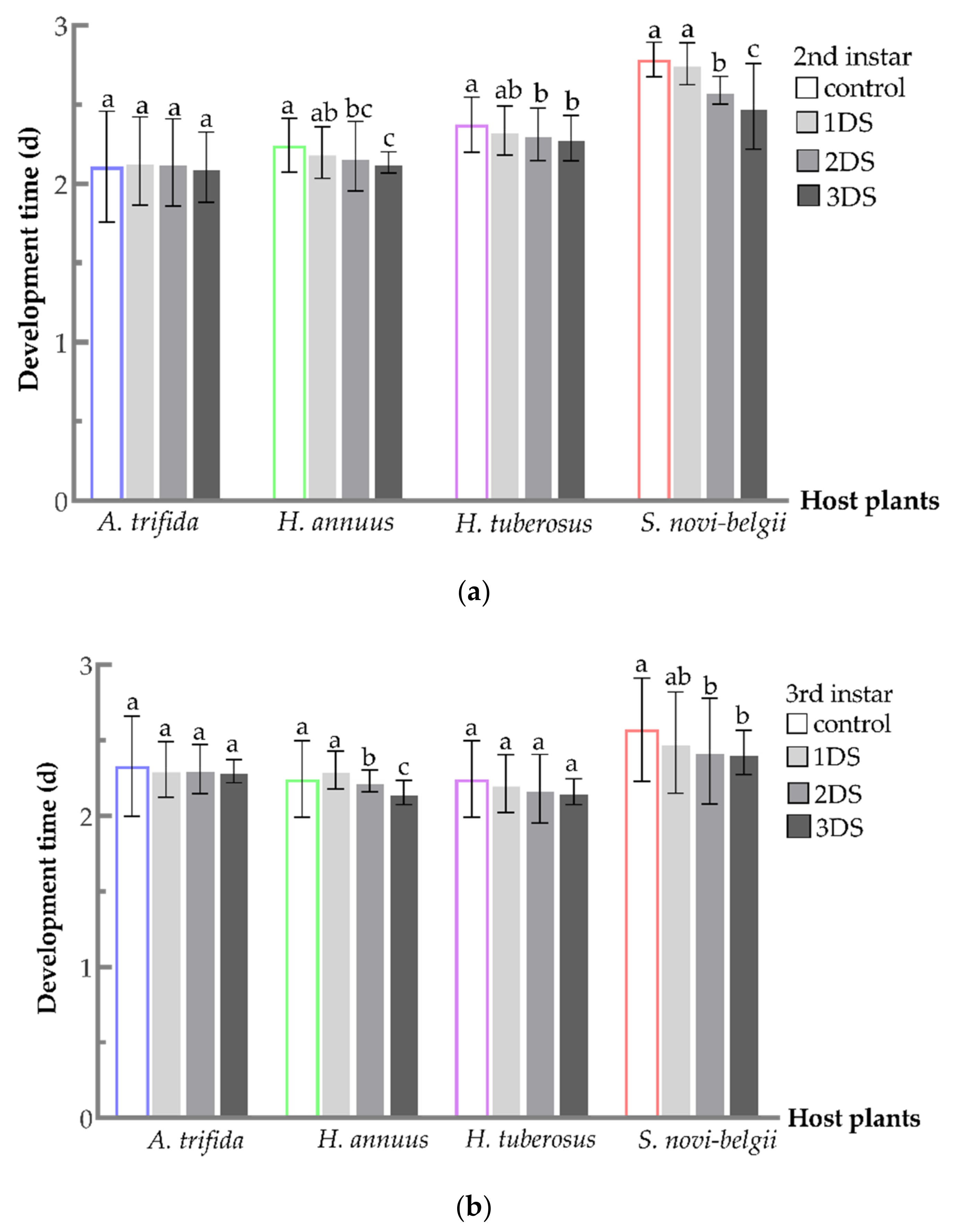
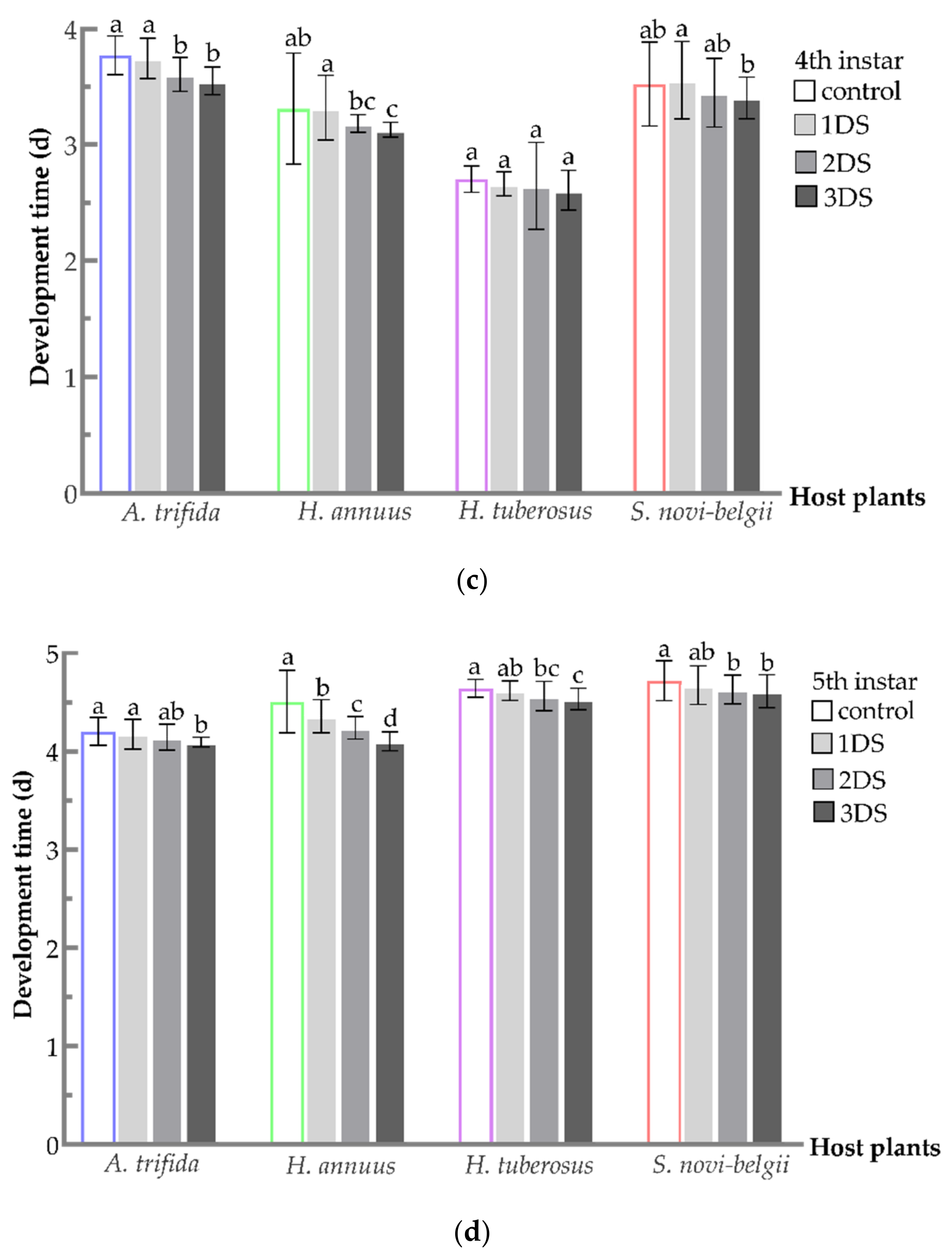
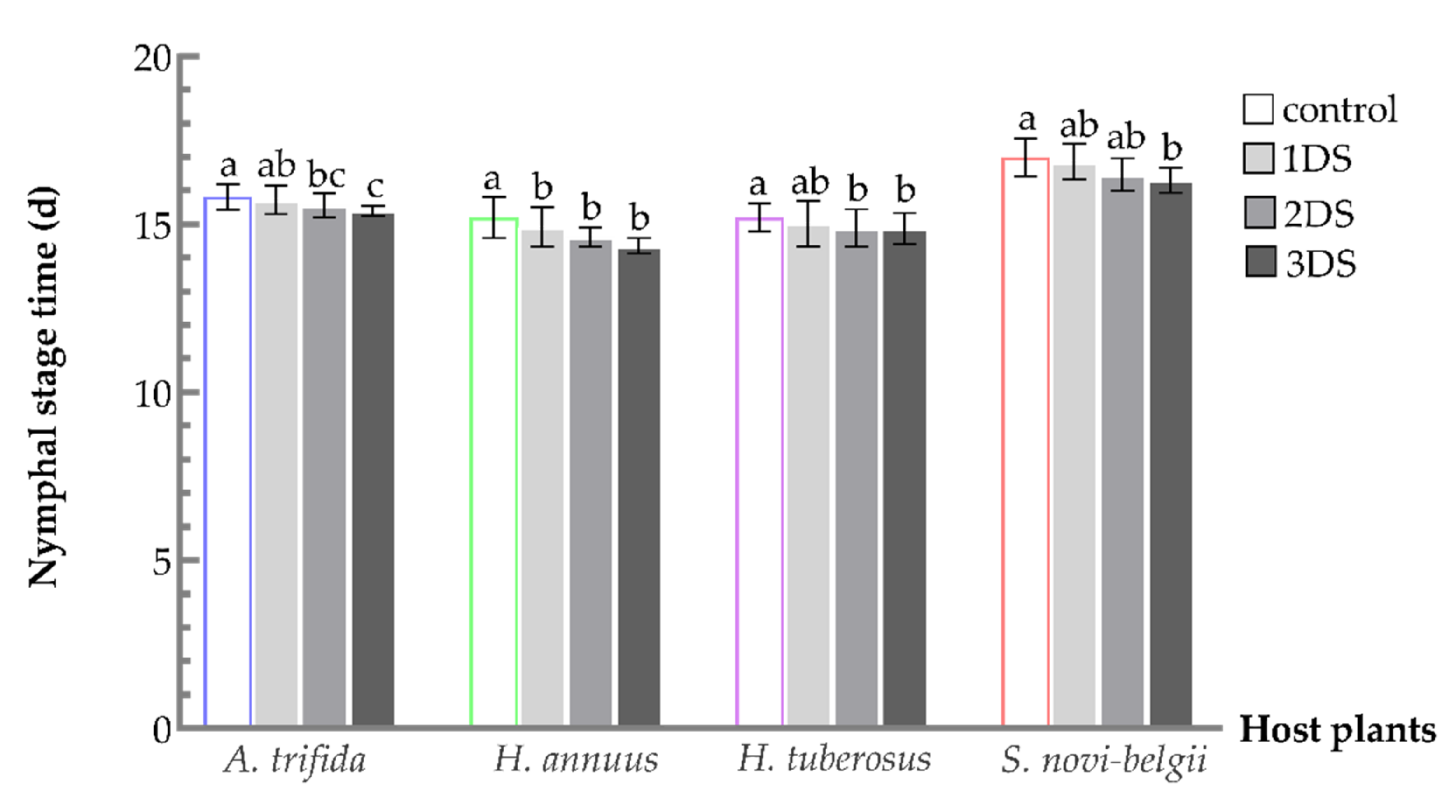
3.1. Shortened Development Time of C. marmorata Suffering from Early Starvation
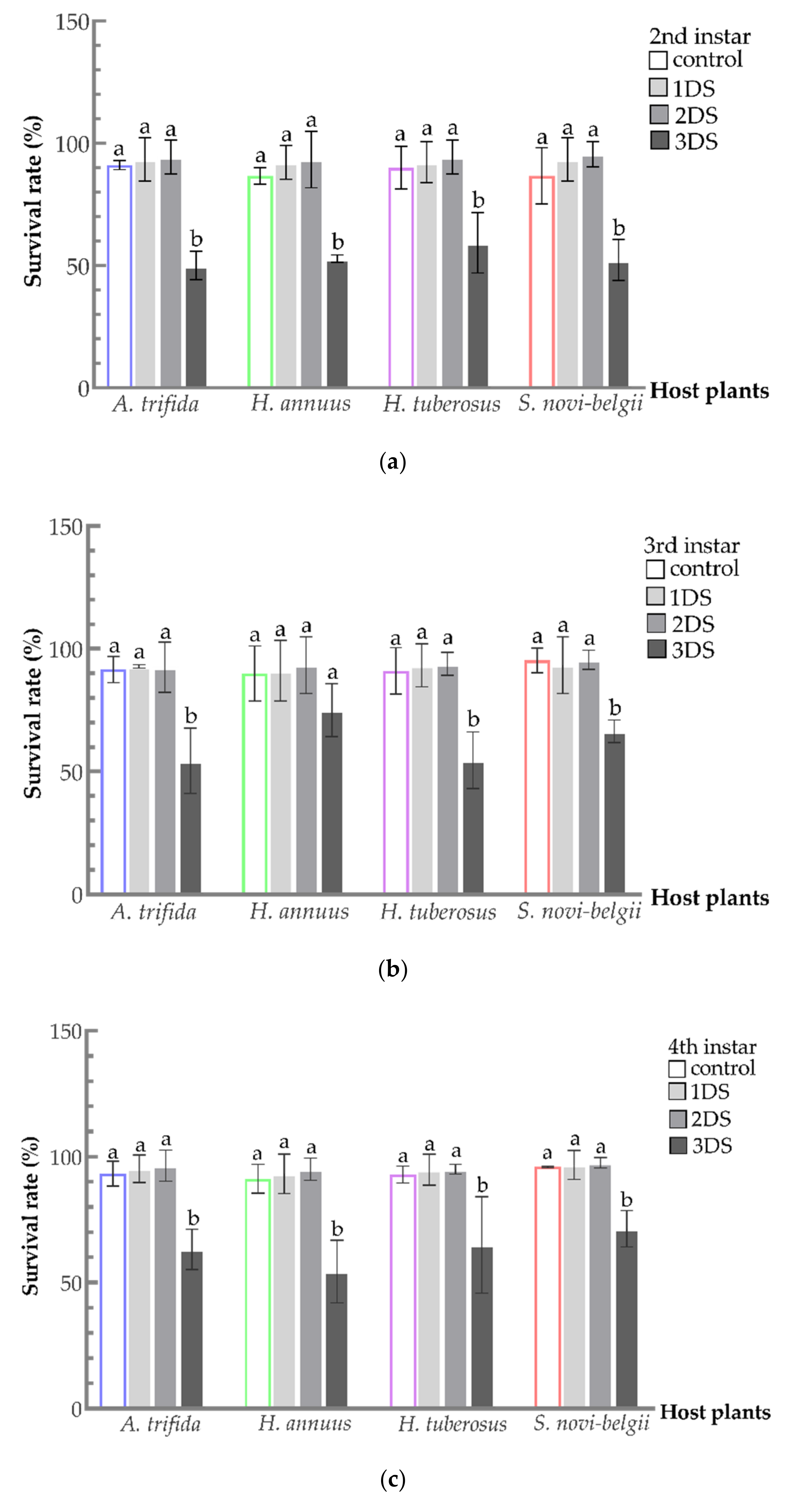
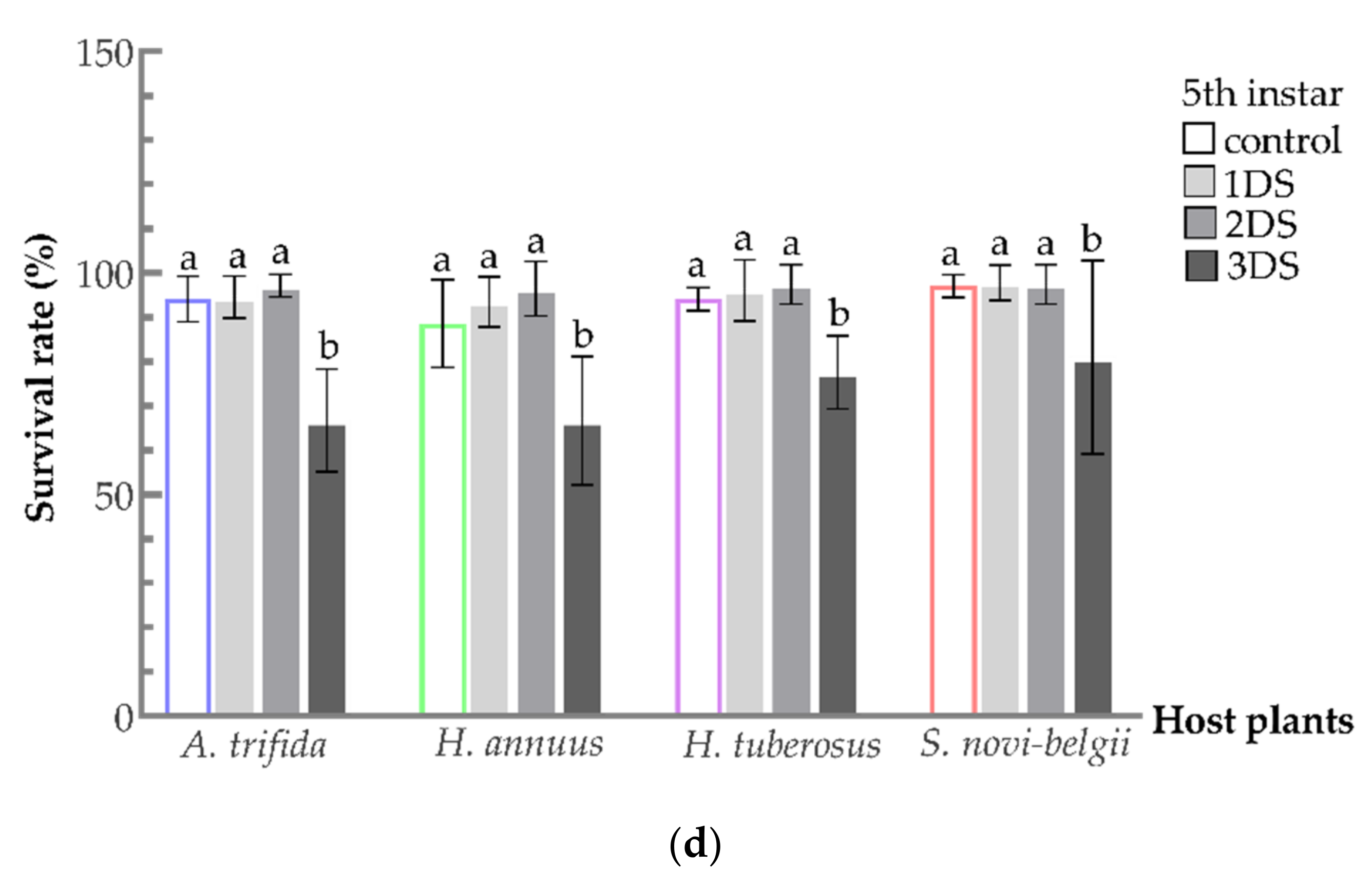
3.2. Sustained High Survival Rate of C. marmorata Suffering from Early Starvation
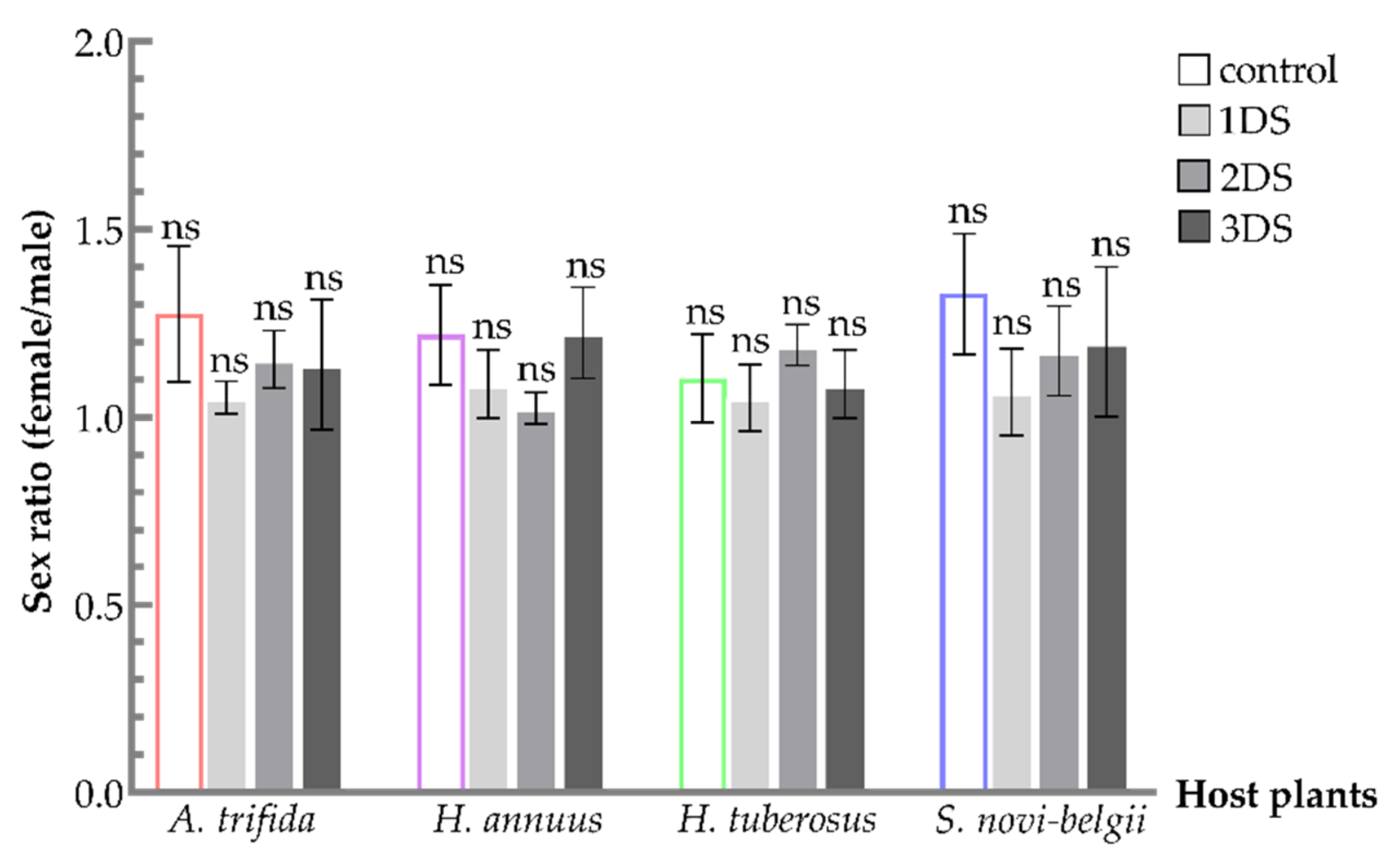
3.3. Similar Sex Ratio of C. marmorata Suffering from Early Starvation
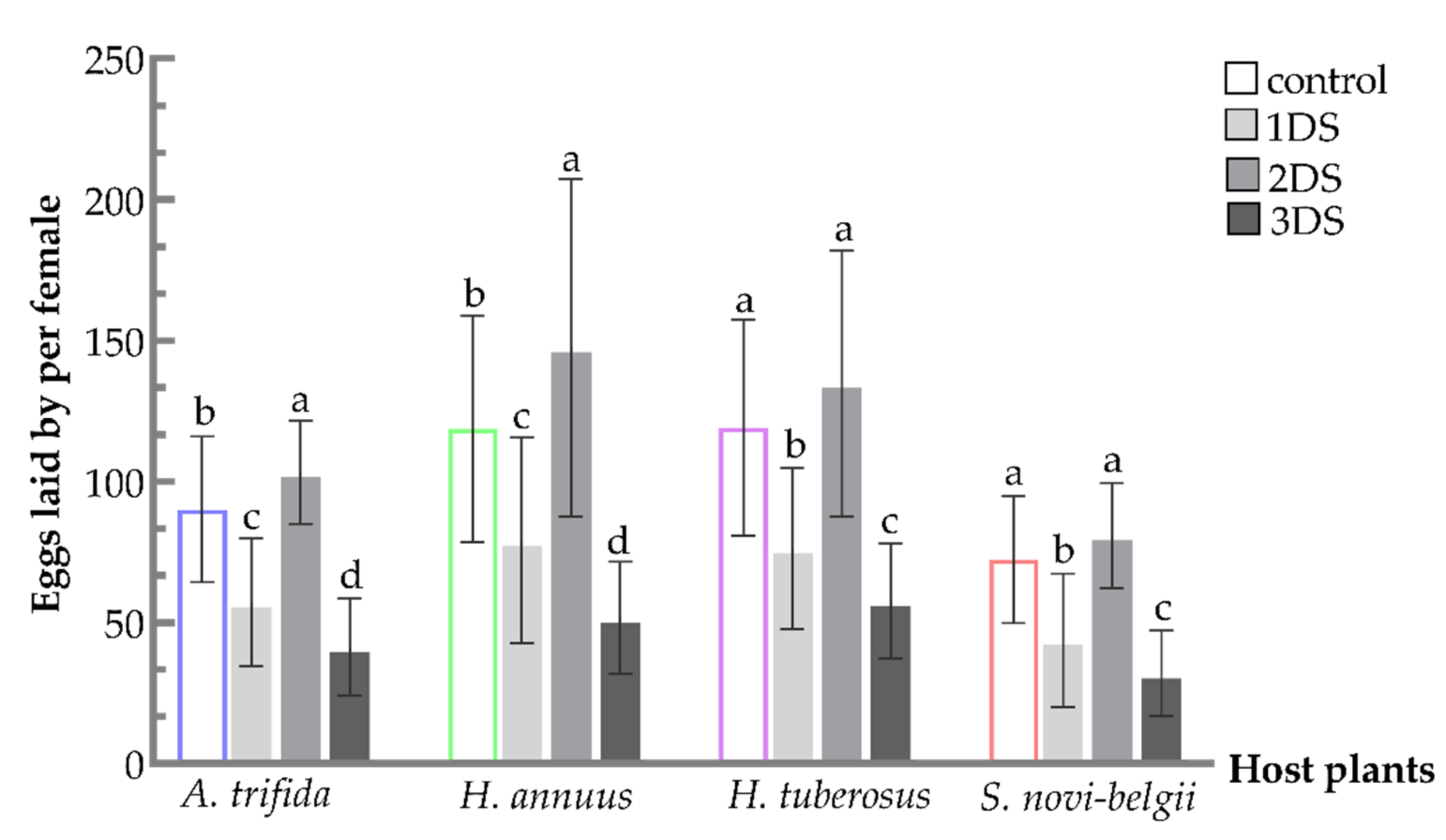
3.4. Increased Number of Eggs Laid by Females Suffering from Early Starvation
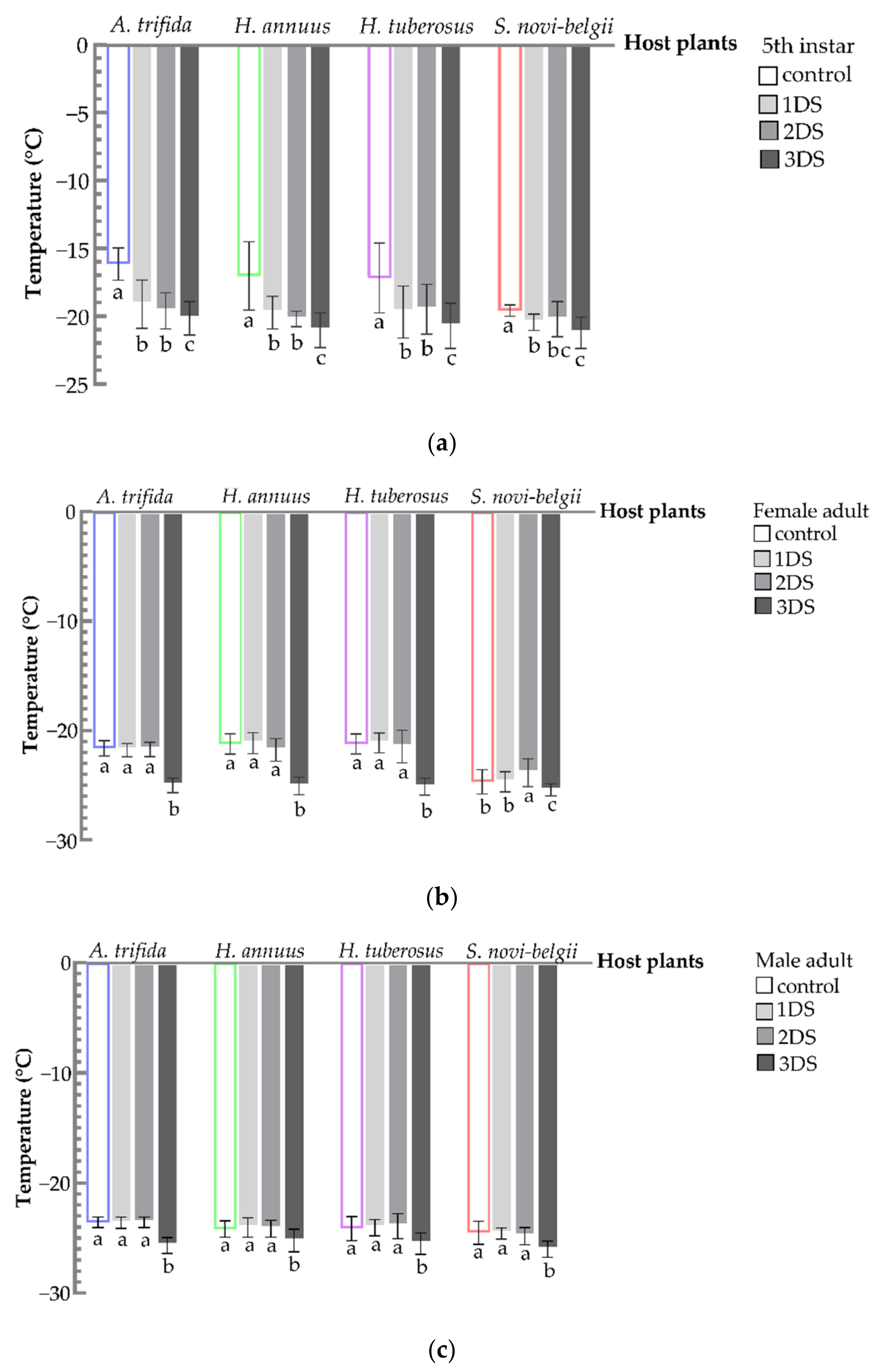
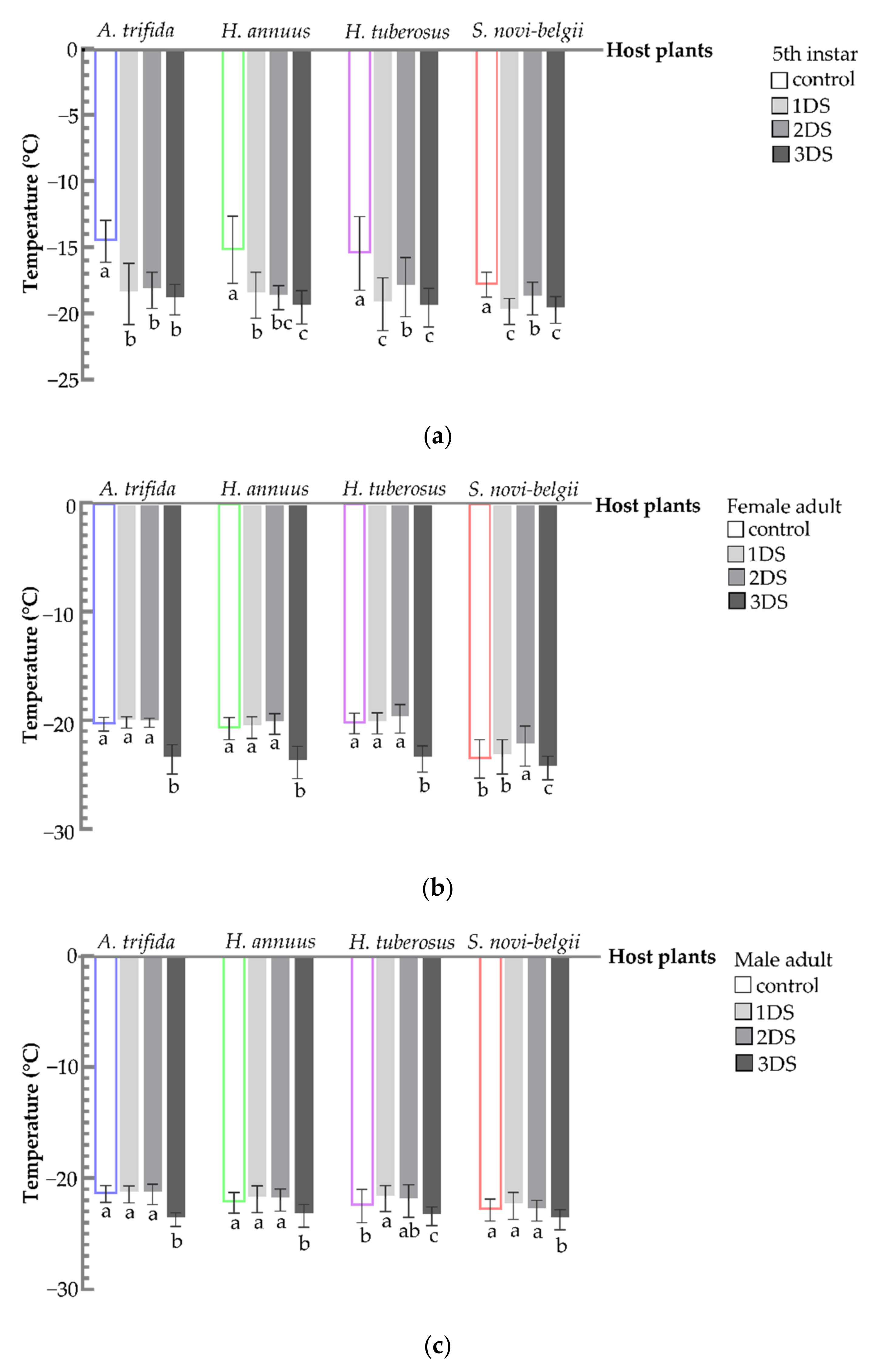
3.5. Decreased SCP and FP of C. marmorata Suffering from Early Starvation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chaby, L.E.; Sheriff, M.J.; Hirrlinger, A.M.; Braithwaite, V.A. Does early stress prepare individuals for a stressful future? Stress during adolescence improves foraging under threat. Anim. Behav. 2015, 105, 37–45. [Google Scholar] [CrossRef]
- Jobson, M.A.; Jordan, J.M.; Sandrof, M.A.; Hibshman, J.D.; Lennox, A.L.; Baugh, L.R. Transgenerational effects of early life starvation on growth, reproduction, and stress resistance in Caenorhabditis elegans. Genetics 2015, 201, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Klosin, A.; Casas, E.; Hidalgo-Carcedo, C.; Vavouri, T.; Lehner, B. Transgenerational transmission of environmental information in C. elegans. Science 2017, 356, 320–323. [Google Scholar] [CrossRef] [Green Version]
- Bauerfeind, S.S.; Fischer, K. Effects of food stress and density in different life stages on reproduction in a butterfly. Oikos 2005, 111, 514–524. [Google Scholar] [CrossRef]
- Boggs, C.L.; Freeman, K.D. Larval food limitation in butterflies: Effects on adult resource allocation and fitness. Oecologia 2005, 144, 353–361. [Google Scholar] [CrossRef]
- Deng, W.H.; Wu, J.Y.; Chen, T.Y.; Yin, Y.M.; Zhu, F. Starvation tolerance and biological characteristics after starvation and refeeding of Hermetia illucens (Diptera: Stratiomyidae) larvae. Acta Entomol. Sin. 2019, 62, 461–467. [Google Scholar] [CrossRef]
- McDougall, S.; Mills, N. The influence of hosts, temperature and food sources on the longevity of Trichogramma platneri. Entomol. Exp. Appl. 1997, 83, 195–203. [Google Scholar] [CrossRef]
- Colinet, H.; Vernon, P.; Hance, T. Does thermal-related plasticity in size and fat reserves influence supercooling abilities and cold-tolerance in Aphidius colemani (Hymenoptera: Aphidiinae) mummies? J. Therm. Biol. 2007, 32, 374–382. [Google Scholar] [CrossRef]
- Benoit, J.B.; Oyen, K.; Finch, G.; Gantz, J.; Wendeln, K.; Arya, T.; Lee, R.E., Jr. Cold hardening improves larval tick questing under low temperatures at the expense of longevity. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2021, 257, 110966. [Google Scholar] [CrossRef]
- Renault, D.; Salin, C.; Vannier, G.; Vernon, P. Survival at low temperatures in insects: What is the ecological significance of the supercooling point? CryoLetters 2002, 23, 217–228. [Google Scholar]
- Ju, R.T.; Du, Y.Z. Mensuration of super-cooling point and principles of cold hardiness of insects. Wuyi Sci. J. 2002, 18, 252–257. [Google Scholar] [CrossRef]
- Pechenik, J.A. Larval experience and latent effects—Metamorphosis is not a new beginning. Integr. Comp. Biol. 2006, 46, 323–333. [Google Scholar] [CrossRef]
- Grettenberger, I.M.; Joseph, S.V. Influence of starvation on walking behavior of Bagrada hilaris (Hemiptera: Pentatomidae). PLoS ONE 2019, 14, e0215446. [Google Scholar] [CrossRef]
- Heijmans, B.T.; Tobi, E.W.; Stein, A.D.; Putter, H.; Blauw, G.J.; Susser, E.S.; Slagboom, P.E.; Lumey, L.H. Persistent epigenetic differences associated with prenatal exposure to famine in humans. Proc. Natl. Acad. Sci. USA 2008, 105, 17046–17049. [Google Scholar] [CrossRef] [Green Version]
- Kohl, K.D.; Amaya, J.; Passement, C.A.; Dearing, M.D.; McCue, M.D. Unique and shared responses of the gut microbiota to prolonged fasting: A comparative study across five classes of vertebrate hosts. FEMS Microbiol. Ecol. 2014, 90, 883–894. [Google Scholar] [CrossRef]
- Urbinati, E.C.; Sarmiento, S.J.; Takahashi, L.S. Short-term cycles of feed deprivation and refeeding promote full compensatory growth in the Amazon fish matrinxã (Brycon amazonicus). Aquaculture 2014, 433, 430–433. [Google Scholar] [CrossRef]
- Drew, W.A.; Arnold, D.C. Tingoidea of Oklahoma (Hemiptera). Proc. Okla. Acad. Sci. 1977, 57, 29–31. [Google Scholar]
- Tallamy, D.W.; Denno, R.F. Alternative life history patterns in risky environments: An example from lacebugs. In Insect Life History Patterns; Springer: Berlin/Heidelberg, Germany, 1981; pp. 129–147. [Google Scholar]
- Lawson, R.H. Economic importance and trends in ornamental horticulture. In Proceedings of the IX International Symposium on Virus Diseases of Ornamental Plants, Herzliya, Israel, 17–22 March 1996; Volume 432, pp. 226–237. [Google Scholar]
- Van Tuyl, J.M.; Arens, P.; Miller, W.B.; Anderson, N.O. The role of ornamentals in human life. In Horticulture: Plants for People and Places, Volume 1: Production Horticulture; Dixon, G.R., Aldous, D.E., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 407–433. [Google Scholar]
- Kato, A.; Ohbayashi, N. Habitat expansion of an exotic lace bug, Corythucha marmorata (Uhler) (Hemiptera: Tingidae), on the Kii Peninsula and Shikoku Island in western Japan. Entomol. Sci. 2009, 12, 130–134. [Google Scholar] [CrossRef]
- Dang, K.; Gao, L.; Zhu, J. First record of the chrysanthemum lace bug, Corythucha marmorata (Uhler, 1878) from China. Dongwu Fenlei Xuebao 2012, 37, 894–898. [Google Scholar]
- Yoon, C.-S.; Kim, H.-G.; Choi, W.-Y.; Cheong, S.-W. First record on the exotic lace bug of Asteraceae, Corythucha marmorata Uhler (Hemiptera: Tingidae) in Korea. J. Environ. Sci. Int. 2013, 22, 1611–1614. [Google Scholar] [CrossRef]
- Dong, L.K.; Wang, Z.H.; Zhang, H.; Yu, J.Y.; Liu, C. The distribution and occurrence of the Corythucha marmorata in Wuhan. Hubei Agric. Sci. 2015, 54, 5299–5302. [Google Scholar] [CrossRef]
- Watanabe, K.; Shimizu, N. Identification of a sex pheromone of the chrysanthemum lace bug Corythucha marmorata (Hemiptera: Tingidae). Sci. Rep. 2017, 7, 7302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J. Distribution and hazards of new record Corythucha marmorata (Uhler, 1878) in Shanghai. J. Anhui Agric. Sci. 2018, 46, 153–156. [Google Scholar] [CrossRef]
- Yang, L.; He, Q.S.; Corscadden, K.; Udenigwe, C.C. The prospects of Jerusalem artichoke in functional food ingredients and bioenergy production. Biotechnol. Rep. 2015, 5, 77–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braman, S.K.; Nair, S.; Carr, E. Influence of temperature, CO2 concentration, and species on survival and development of lace bugs (Hemiptera: Tingidae). J. Entomol. Sci. 2013, 48, 251–254. [Google Scholar] [CrossRef]
- Sakata, Y.; Craig, T.P. An exotic herbivore reinforces competition between exotic and native plants. J. Ecol. 2021, 109, 2740–2753. [Google Scholar] [CrossRef]
- Kim, D.E.; Kil, J. Geographical distribution and host plants of Corythucha marmorata (Uhler) (Hemiptera: Tingidae) in Korea. Korean J. Appl. Entomol. 2014, 53, 185–191. [Google Scholar] [CrossRef]
- Cho, G.; Kim, W.; Kwon, Y.S.; Lee, S. Check list of lace bugs (Hemiptera: Tingidae) of Korea. J. Asia Pac. Entomol. 2020, 23, 736–745. [Google Scholar] [CrossRef]
- Cappuccino, N.; Root, R.B. The significance of host patch edges to the colonization and development of Corythucha marmorata (Hemiptera: Tingidae). Ecol. Entomol. 1992, 17, 109–113. [Google Scholar] [CrossRef]
- Luo, Y.; Xiao, F.; Zhao, R.N.; Yin, Z.Y.; Li, T.M.; Guo, F.; Wu, X.S.; Chen, W.L. Predation on five species of Tingidae (Hemiptera) by Stethoconus japonicas. Chin. J. Appl. Entomol. 2020, 57, 413–420. [Google Scholar] [CrossRef]
- Shen, J.S.; Zhu, M.; Cui, X.H.; Li, L.J. Life table and biological characteristics of an exotic lace bug, Corythucha marmorata (Uhler). Chin. J. Appl. Ecol. 2016, 27, 1657–1662. [Google Scholar] [CrossRef]
- Yu, G.Y. Corythucha marmorata. Plant Prot. 2014, 40, 7. [Google Scholar] [CrossRef]
- Lu, H.P.; Liu, X.; He, Y.M.; Guo, D.C. Corythucha marmorata—An invasive pest newly recorded at Ganzhou city, China. Biol. Disaster Sci. 2018, 41, 55–57. [Google Scholar] [CrossRef]
- Cheng, Q.; Wu, X.Q.; Ye, J.R.; Lin, S.X. Distribution of Corythucha marmorata in Nanjing and its risk analysis in China. J. Nanjing For. Univ. 2020, 44, 125–130. [Google Scholar] [CrossRef]
- Pan, P.L.; Zhang, F.M.; Yin, J.; Liu, H.M.; Zhou, S.Y.; Zhi, Y.N. Preliminary studies on image recognition technology for female and male adults of Corythucha marmorata (Uhler) (Hemiptera: Tingidae). Plant Prot. 2017, 43, 70–75. [Google Scholar] [CrossRef]
- Picciau, L.; Alma, A.; Ferracini, C. Effect of different feeding sources on lifespan and fecundity in the biocontrol agent Torymus sinensis. Biol. Control 2019, 134, 45–52. [Google Scholar] [CrossRef]
- Murillo, H.; Hunt, D.W.A.; VanLaerhoven, S.L. Fecundity and life table parameters of Campoletis sonorensis (Hymenoptera: Ichneumonidae), an Endoparasitoid of the Cabbage Looper Trichoplusia ni Hübner (Lepidoptera: Noctuidae), under laboratory conditions. Biocontrol Sci. Technol. 2012, 22, 125–134. [Google Scholar] [CrossRef]
- Yu, C.; Yao, M.Y.; Zhao, R.N.; Wang, L.; Chen, W.L. Effects of breeding density on growth and development of Eocanthecona Furcellata (Wolff.). J. Mt. Agric. Biol. 2021, 40, 75–79. [Google Scholar] [CrossRef]
- Wang, Z.H.; Yu, J.Y.; Shen, J.; Liang, Y.T.; Zhang, X.Q.; Dong, L.K.; Yu, H.F. Prediction of potential distribution of the invasive Chrysanthemum Lace Bug, Corythucha marmorata in China based on Maxent. J. Environ. Entomol. 2019, 41, 626–633. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, W.; Chen, W. Early Starvation Contributes to the Adaptive Capacity of Corythucha marmorata (Uhler), an Emerging Pest in China. Biology 2022, 11, 80. https://doi.org/10.3390/biology11010080
Zhou W, Chen W. Early Starvation Contributes to the Adaptive Capacity of Corythucha marmorata (Uhler), an Emerging Pest in China. Biology. 2022; 11(1):80. https://doi.org/10.3390/biology11010080
Chicago/Turabian StyleZhou, Wei, and Wenlong Chen. 2022. "Early Starvation Contributes to the Adaptive Capacity of Corythucha marmorata (Uhler), an Emerging Pest in China" Biology 11, no. 1: 80. https://doi.org/10.3390/biology11010080
APA StyleZhou, W., & Chen, W. (2022). Early Starvation Contributes to the Adaptive Capacity of Corythucha marmorata (Uhler), an Emerging Pest in China. Biology, 11(1), 80. https://doi.org/10.3390/biology11010080