
Monophosphoryl Lipid A and Poly I:C Combination Adjuvant Promoted Ovalbumin-Specific Cell Mediated Immunity in Mice Model


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Reagents
2.2. Immunization
2.3. Sample Preparation
2.4. Serum Antigen (Ag)-Specific Antibody ELISA
2.5. Cytokine ELISA
2.6. Memory B Cell Response
2.7. Flow Cytometry
2.8. Bone Marrow-Derived Dendritic Cells (DCs) and Macrophage Culture and Mixed Lymphocyte Reaction (MLR)
2.9. Statistical Analysis
3. Results
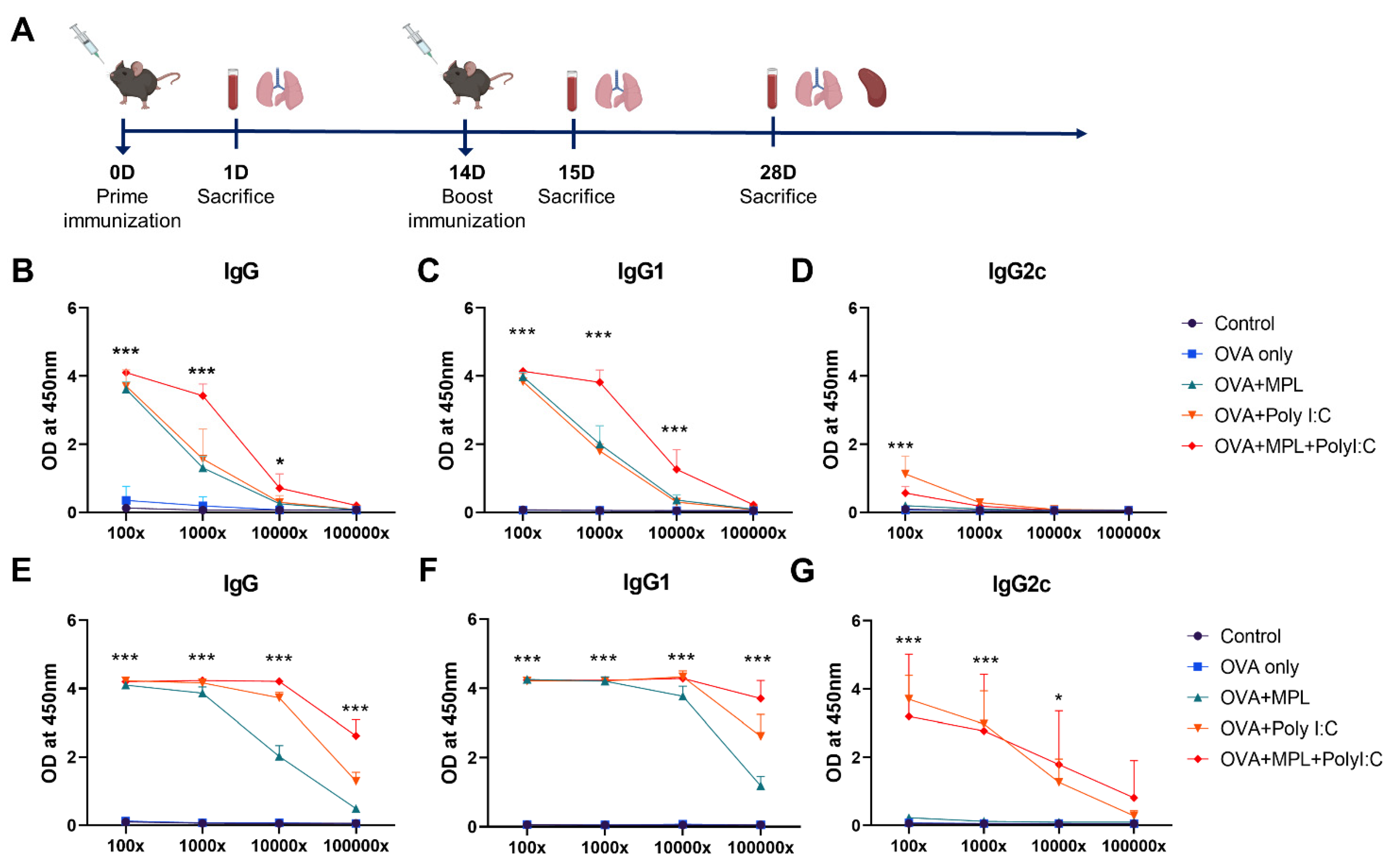
3.1. Combination of MPL and Poly I:C Enhanced the OVA-Specific IgG Antibody Responses
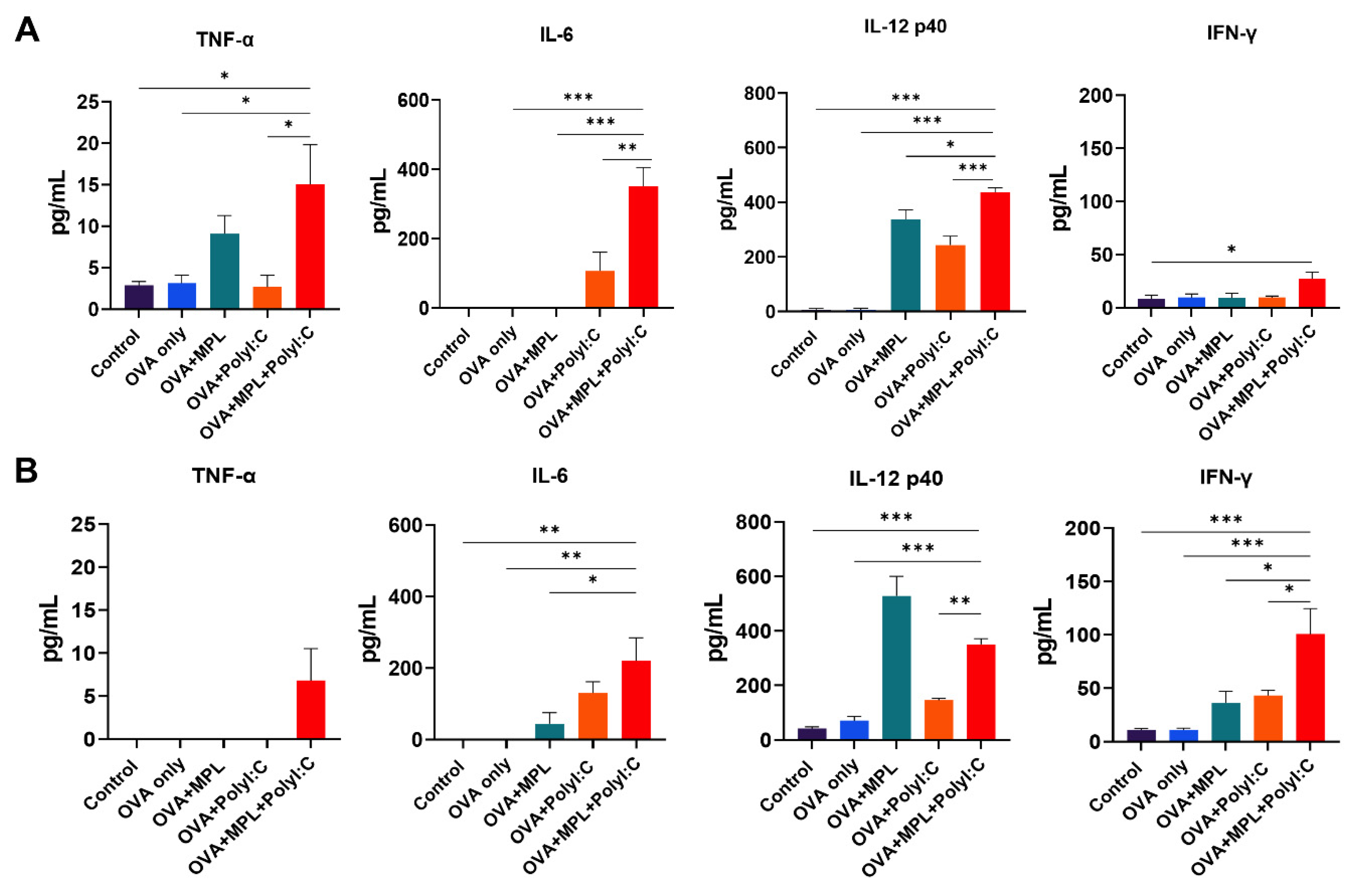
3.2. Combination of MPL and Poly I:C Promoted the Induction of Initial Inflammatory Cytokines after Immunization
3.3. OVA Immunization with the MPL and Poly I:C Combination Adjuvant Recruited Inflammatory Cells to the Site of Immunization
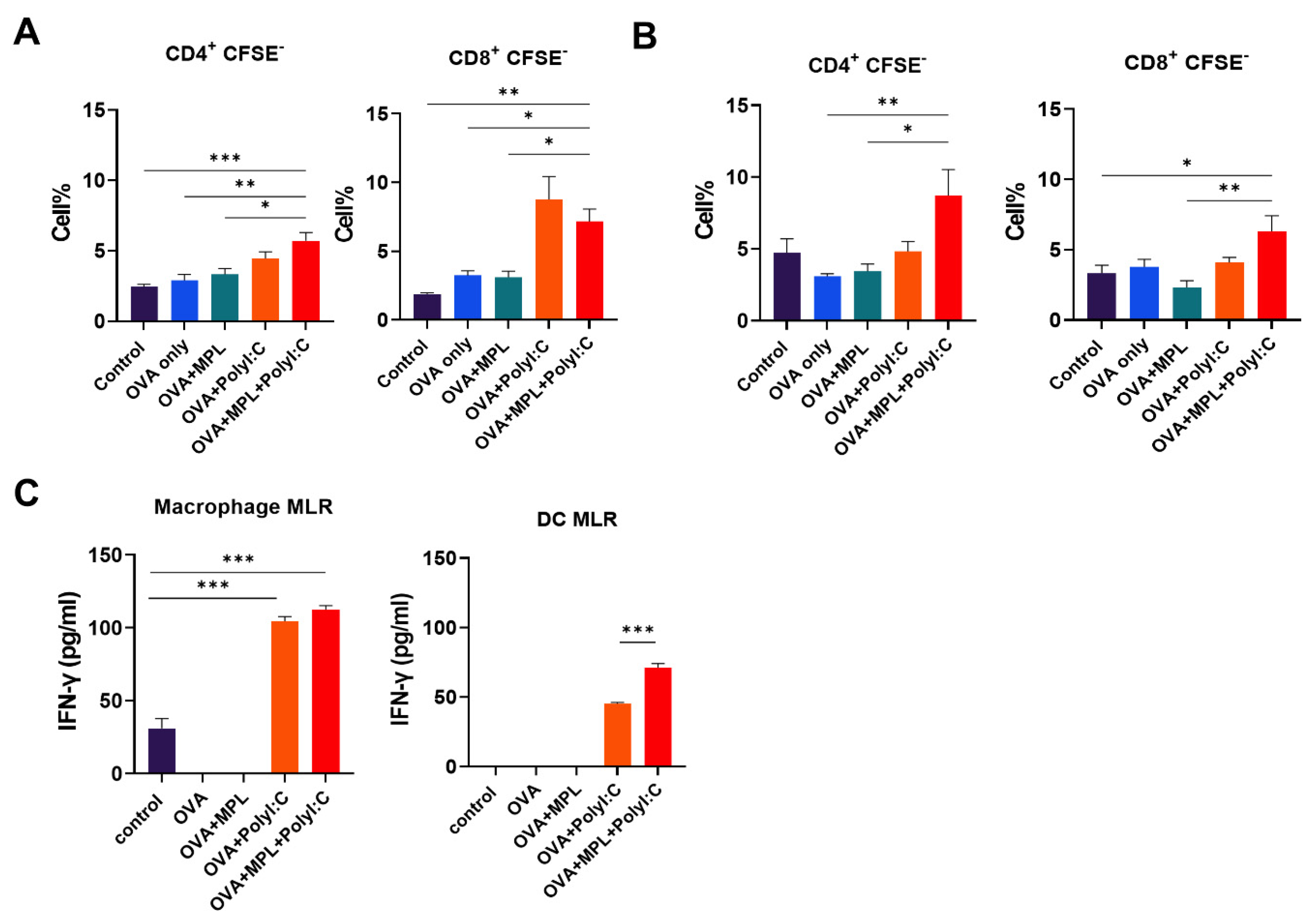
3.4. Adjuvants Promote In Vitro APC Activity to Proliferate Lymphocytes
3.5. Effects of the Adjuvants on the Memory T Cell Population and Proliferation of Antigen-Specific T Cells
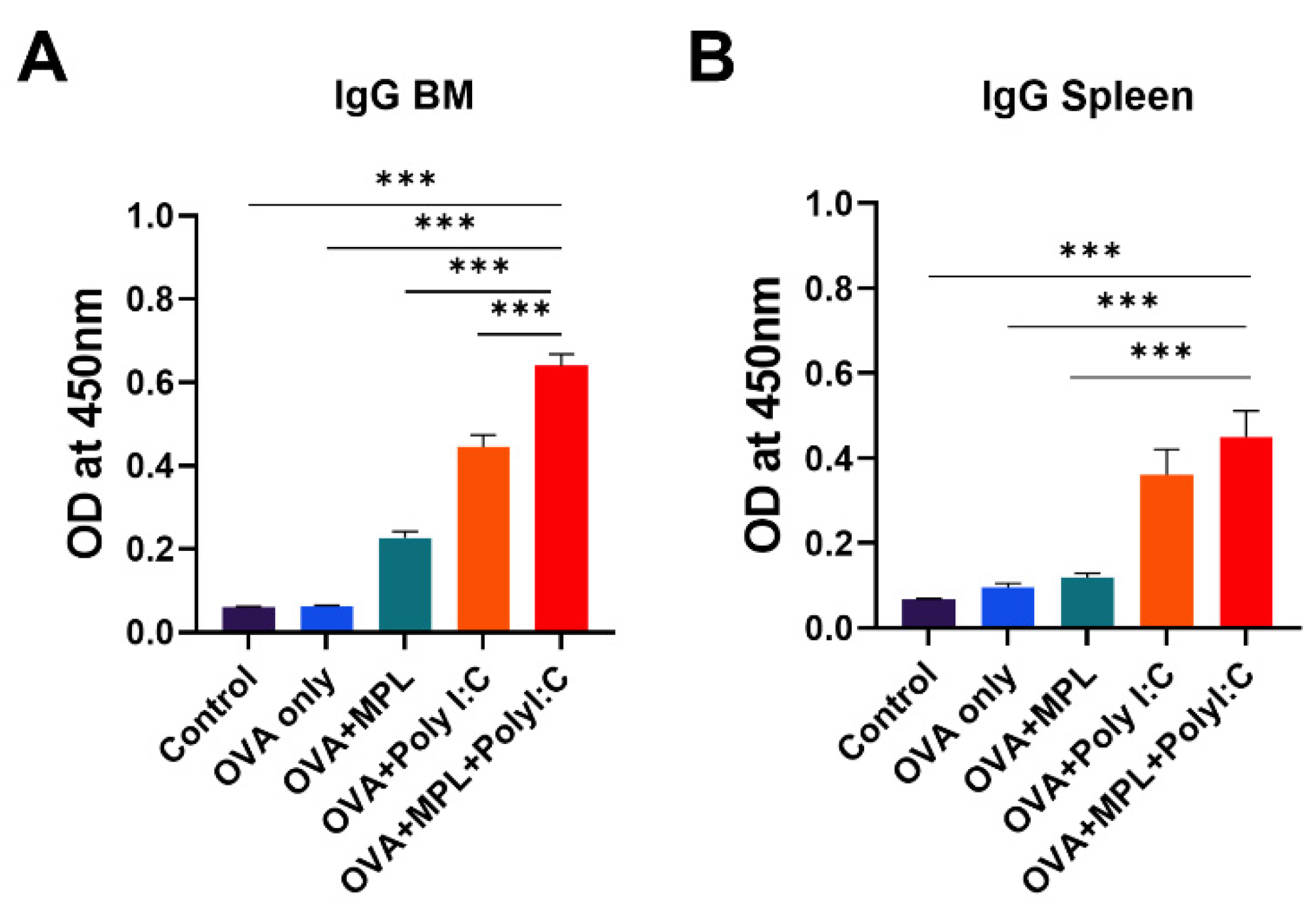
3.6. Combination of MPL and Poly I:C Enhanced the Antigen-Specific Memory B Cell Responses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thomas, P.G.; Keating, R.; Hulse-Post, D.J.; Doherty, P.C. Cell-mediated protection in influenza infection. Emerg. Infect. Dis. 2006, 12, 48–54. [Google Scholar] [CrossRef] [PubMed]
- McMichael, A.J.; Rowland-Jones, S.L. Cellular immune responses to HIV. Nat. Cell. Biol. 2001, 410, 980–987. [Google Scholar] [CrossRef]
- Coffman, R.L.; Sher, A.; Seder, R.A. Vaccine adjuvants: Putting innate immunity to work. Immunity 2010, 33, 492–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olive, C. Pattern recognition receptors: Sentinels in innate immunity and targets of new vaccine adjuvants. Expert Rev. Vaccines 2012, 11, 237–256. [Google Scholar] [CrossRef] [PubMed]
- Wille-Reece, U.; Flynn, B.J.; Lore, K.; Koup, R.A.; Miles, A.P.; Saul, A.; Kedl, R.M.; Mattapallil, J.J.; Weiss, W.R.; Roederer, M.; et al. Toll-like receptor agonists influence the magnitude and quality of memory T cell responses after prime-boost immunization in nonhuman primates. J. Exp. Med. 2006, 203, 1249–1258. [Google Scholar] [CrossRef]
- Sallusto, F.; Geginat, J.; Lanzavecchia, A. Central memory and effector memory T cell subsets: Function, generation, and maintenance. Annu Rev. Immunol 2004, 22, 745–763. [Google Scholar] [CrossRef] [PubMed]
- Lindenstrom, T.; Agger, E.M.; Korsholm, K.S.; Darrah, P.A.; Aagaard, C.; Seder, R.A.; Rosenkrands, I.; Andersen, P. Tuberculosis subunit vaccination provides long-term protective immunity characterized by multifunctional CD4 memory T cells. J. Immunol. 2009, 182, 8047–8055. [Google Scholar] [CrossRef]
- Rai, D.; Martin, M.D.; Badovinac, V.P. The longevity of memory CD8 T cell responses after repetitive antigen stimulations. J. Immunol. 2014, 192, 5652–5659. [Google Scholar] [CrossRef] [Green Version]
- Mbow, M.L.; De Gregorio, E.; Valiante, N.M.; Rappuoli, R. New adjuvants for human vaccines. Curr. Opin. Immunol. 2010, 22, 411–416. [Google Scholar] [CrossRef]
- Spellberg, B.; Edwards, J.E., Jr. Type 1/Type 2 immunity in infectious diseases. Clin. Infect. Dis. 2001, 32, 76–102. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Nguyen, M.T. Recent advances of vaccine adjuvants for infectious diseases. Immune Netw. 2015, 15, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Garcon, N.; Chomez, P.; Van Mechelen, M. GlaxoSmithKline Adjuvant Systems in vaccines: Concepts, achievements and perspectives. Expert Rev. Vaccines 2007, 6, 723–739. [Google Scholar] [CrossRef]
- Didierlaurent, A.M.; Laupeze, B.; Di Pasquale, A.; Hergli, N.; Collignon, C.; Garcon, N. Adjuvant system AS01: Helping to overcome the challenges of modern vaccines. Expert Rev. Vaccines 2017, 16, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Reed, S.G.; Tomai, M.; Gale, M.J., Jr. New horizons in adjuvants for vaccine development. Curr. Opin. Immunol. 2020, 65, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Wack, A.; Baudner, B.C.; Hilbert, A.K.; Manini, I.; Nuti, S.; Tavarini, S.; Scheffczik, H.; Ugozzoli, M.; Singh, M.; Kazzaz, J.; et al. Combination adjuvants for the induction of potent, long-lasting antibody and T-cell responses to influenza vaccine in mice. Vaccine 2008, 26, 552–561. [Google Scholar] [CrossRef] [PubMed]
- Levast, B.; Awate, S.; Babiuk, L.; Mutwiri, G.; Gerdts, V.; van Drunen Littel-van den Hurk, S. Vaccine Potentiation by Combination Adjuvants. Vaccines 2014, 2, 297–322. [Google Scholar] [CrossRef] [Green Version]
- Ko, E.-J.; Lee, Y.; Lee, Y.-T.; Kim, Y.-J.; Kim, K.-H.; Kang, S.-M. MPL and CpG combination adjuvants promote homologous and heterosubtypic cross protection of inactivated split influenza virus vaccine. Antivir. Res. 2018, 156, 107–115. [Google Scholar] [CrossRef]
- Ko, E.-J.; Lee, Y.-T.; Lee, Y.; Kim, K.-H.; Kang, S.-M. Distinct Effects of Monophosphoryl Lipid A, Oligodeoxynucleotide CpG, and Combination Adjuvants on Modulating Innate and Adaptive Immune Responses to Influenza Vaccination. Immune Netw. 2017, 17, 326–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.-Q.; Bazin-Lee, H.; Evans, J.T.; Casella, C.R.; Mitchell, T.C. MPL Adjuvant Contains Competitive Antagonists of Human TLR4. Front. Immunol. 2020, 11, 577823. [Google Scholar] [CrossRef]
- Kalali, B.N.; Kollisch, G.; Mages, J.; Muller, T.; Bauer, S.; Wagner, H.; Ring, J.; Lang, R.; Mempel, M.; Ollert, M. Double-stranded RNA induces an antiviral defense status in epidermal keratinocytes through TLR3-, PKR-, and MDA5/RIG-I-mediated differential signaling. J. Immunol. 2008, 181, 2694–2704. [Google Scholar] [CrossRef] [Green Version]
- Ngoi, S.M.; Tovey, M.G.; Vella, A.T. Targeting poly(I:C) to the TLR3-independent pathway boosts effector CD8 T cell differentiation through IFN-α/β. J. Immunol. 2008, 181, 7670–7680. [Google Scholar] [CrossRef]
- Navabi, H.; Jasani, B.; Reece, A.; Clayton, A.; Tabi, Z.; Donninger, C.; Mason, M.; Adams, M. A clinical grade poly I:C-analogue (Ampligen®) promotes optimal DC maturation and Th1-type T cell responses of healthy donors and cancer patients in vitro. Vaccine 2009, 27, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Apostolico, J.S.; Lunardelli, V.A.S.; Yamamoto, M.M.; Cunha-Neto, E.; Boscardin, S.B.; Rosa, D.S. Poly(I:C) Potentiates T Cell Immunity to a Dendritic Cell Targeted HIV-Multiepitope Vaccine. Front. Immunol. 2019, 10, 843. [Google Scholar] [CrossRef]
- Park, H.; Adamson, L.; Ha, T.; Mullen, K.; Hagen, S.I.; Nogueron, A.; Sylwester, A.W.; Axthelm, M.K.; Legasse, A.; Piatak, M., Jr.; et al. Polyinosinic-polycytidylic acid is the most effective TLR adjuvant for SIV Gag protein-induced T cell responses in nonhuman primates. J. Immunol. 2013, 190, 4103–4115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacLeod, M.K.L.; McKee, A.S.; David, A.; Wang, J.; Mason, R.; Kappler, J.W.; Marrack, P. Vaccine adjuvants aluminum and monophosphoryl lipid A provide distinct signals to generate protective cytotoxic memory CD8 T cells. Proc. Nat. Acad. Sci. USA 2011, 108, 7914–7919. [Google Scholar] [CrossRef] [Green Version]
- Ravindran, R.; Bhowmick, S.; Das, A.; Ali, N. Comparison of BCG, MPL and cationic liposome adjuvant systems in leishmanial antigen vaccine formulations against murine visceral leishmaniasis. BMC Microbiol. 2010, 10, 181. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Quan, N. Immune Cell Isolation from Mouse Femur Bone Marrow. Bio-Protocol 2015, 5, e1631. [Google Scholar] [CrossRef] [PubMed]
- GeurtsvanKessel, C.H.; Lambrecht, B.N. Division of labor between dendritic cell subsets of the lung. Mucosal Immunol. 2008, 1, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Rose, S.; Misharin, A.; Perlman, H. A novel Ly6C/Ly6G-based strategy to analyze the mouse splenic myeloid compartment. Cytom. Part A 2012, 81, 343–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.F.; Berger, H.; Su, I.-H. Isolation and Activation of Murine Lymphocytes. J. Vis. Exp. 2016, e54596. [Google Scholar] [CrossRef]
- Cyster, J.G.; Allen, C.D.C. B Cell Responses: Cell Interaction Dynamics and Decisions. Cell 2019, 177, 524–540. [Google Scholar] [CrossRef] [Green Version]
- Salerno-Goncalves, R.; Sztein, M.B. Cell-mediated immunity and the challenges for vaccine development. Trends Microbiol. 2006, 14, 536–542. [Google Scholar] [CrossRef] [PubMed]
- Saravia, J.; Chapman, N.M.; Chi, H. Helper T cell differentiation. Cell. Mol. Immunol. 2019, 16, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Mata-Haro, V. The vaccine adjuvant monophosphoryl lipid A as a TRIF-biased agonist of TLR4. Science 2007, 317, 1036. [Google Scholar] [CrossRef]
- Akira, S.; Takeda, K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004, 4, 499–511. [Google Scholar] [CrossRef]
- Casella, C.R.; Mitchell, T.C. Putting endotoxin to work for us: Monophosphoryl lipid A as a safe and effective vaccine adjuvant. Cell. Mol. Life Sci. 2008, 65, 3231–3240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiscott, J. Triggering the innate antiviral response through IRF-3 activation. J. Biol. Chem. 2007, 282, 15325–15329. [Google Scholar] [CrossRef] [Green Version]
- Martins, K.A.O.; Cooper, C.L.; Stronsky, S.M.; Norris, S.L.W.; Kwilas, S.A.; Steffens, J.T.; Benko, J.G.; van Tongeren, S.A.; Bavari, S. Adjuvant-enhanced CD4 T Cell Responses are Critical to Durable Vaccine Immunity. EBioMedicine 2016, 3, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Bravo, M.; Ardavin, C. In vivo induction of immune responses to pathogens by conventional dendritic cells. Immunity 2008, 29, 343–351. [Google Scholar] [CrossRef] [Green Version]
- Ismaili, J.; Rennesson, J.; Aksoy, E.; Vekemans, J.; Vincart, B.; Amraoui, Z.; Van Laethem, F.; Goldman, M.; Dubois, P.M. Monophosphoryl lipid A activates both human dendritic cells and T cells. J. Immunol. 2002, 168, 926–932. [Google Scholar] [CrossRef]
- Vidyarthi, A.; Khan, N.; Agnihotri, T.; Negi, S.; Das, D.K.; Aqdas, M.; Chatterjee, D.; Colegio, O.R.; Tewari, M.K.; Agrewala, J.N. TLR-3 Stimulation Skews M2 Macrophages to M1 Through IFN-αβ Signaling and Restricts Tumor Progression. Front. Immunol. 2018, 9, 1650. [Google Scholar] [CrossRef] [PubMed]
- Watad, A.; Quaresma, M.; Brown, S.; Tervaert, J.W.C.; Rodriguez-Pint, I.; Cervera, R.; Perricone, C.; Shoenfeld, Y. Autoimmune/inflammatory syndrome induced by adjuvants (Shoenfeld’s syndrome)‒An update. Lupus 2017, 26, 675–681. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahn, S.Y.; Le, C.T.T.; Ko, E.-J. Monophosphoryl Lipid A and Poly I:C Combination Adjuvant Promoted Ovalbumin-Specific Cell Mediated Immunity in Mice Model. Biology 2021, 10, 908. https://doi.org/10.3390/biology10090908
Ahn SY, Le CTT, Ko E-J. Monophosphoryl Lipid A and Poly I:C Combination Adjuvant Promoted Ovalbumin-Specific Cell Mediated Immunity in Mice Model. Biology. 2021; 10(9):908. https://doi.org/10.3390/biology10090908
Chicago/Turabian StyleAhn, So Yeon, Chau Thuy Tien Le, and Eun-Ju Ko. 2021. "Monophosphoryl Lipid A and Poly I:C Combination Adjuvant Promoted Ovalbumin-Specific Cell Mediated Immunity in Mice Model" Biology 10, no. 9: 908. https://doi.org/10.3390/biology10090908
APA StyleAhn, S. Y., Le, C. T. T., & Ko, E.-J. (2021). Monophosphoryl Lipid A and Poly I:C Combination Adjuvant Promoted Ovalbumin-Specific Cell Mediated Immunity in Mice Model. Biology, 10(9), 908. https://doi.org/10.3390/biology10090908