
Reproductive and Developmental Biology of Acroclisoides sinicus, a Hyperparasitoid of Scelionid Parasitoids


 ,
, 
Abstract
Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Origin of the Insects for Studies in Europe
2.2. Establishment of Acroclisoides sinicus Laboratory Colonies in Europe
2.3. Rearing Acroclisoides sinicus in the USA under Laboratory and Field Conditions
2.4. Acroclisoides sinicus Reproductive Biology Studies in Europe
2.5. Acroclisoides sinicus Immature Development Study in Europe
2.6. Development of Acroclisoides sinicus in Association with Native European Pentatomids
2.7. Acroclisoides sinicus Longevity Tests in Europe
2.8. Polyphenism and Female Egg Load of Acroclisoides sinicus Study in Europe
3. Results
3.1. Establishment of Laboratory-Reared Colonies in Europe
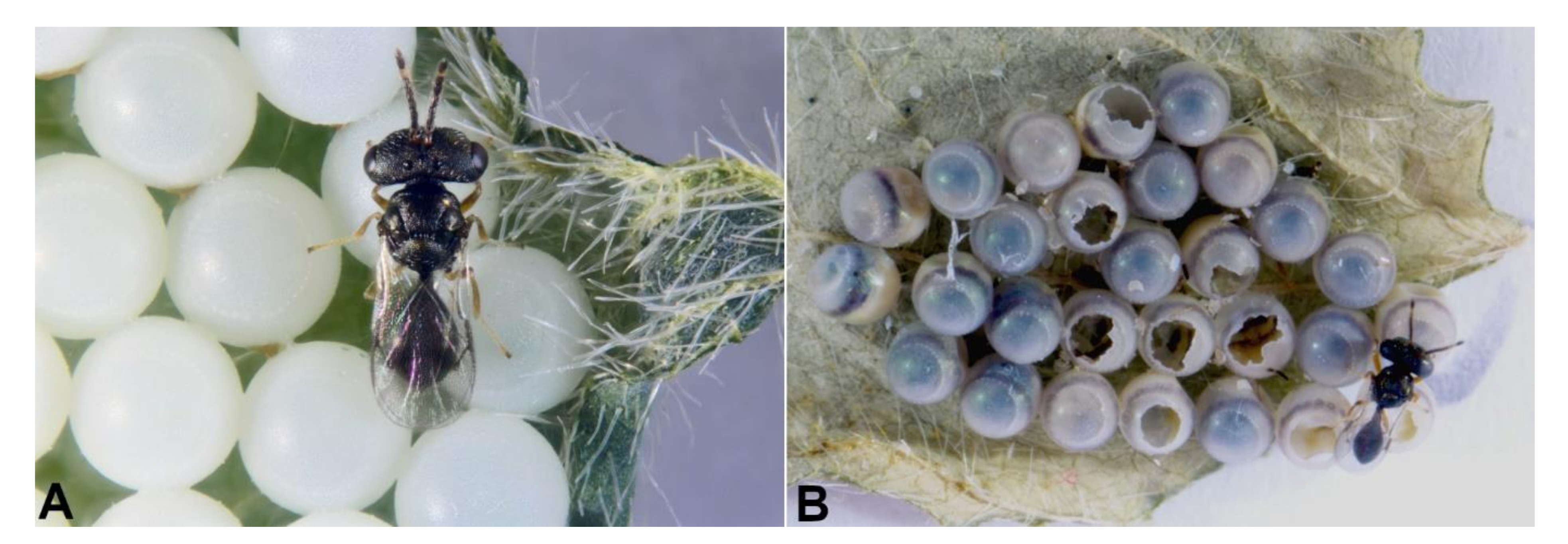
3.2. Reproductive Biological Traits of Acroclisoides sinicus in Europe and the USA
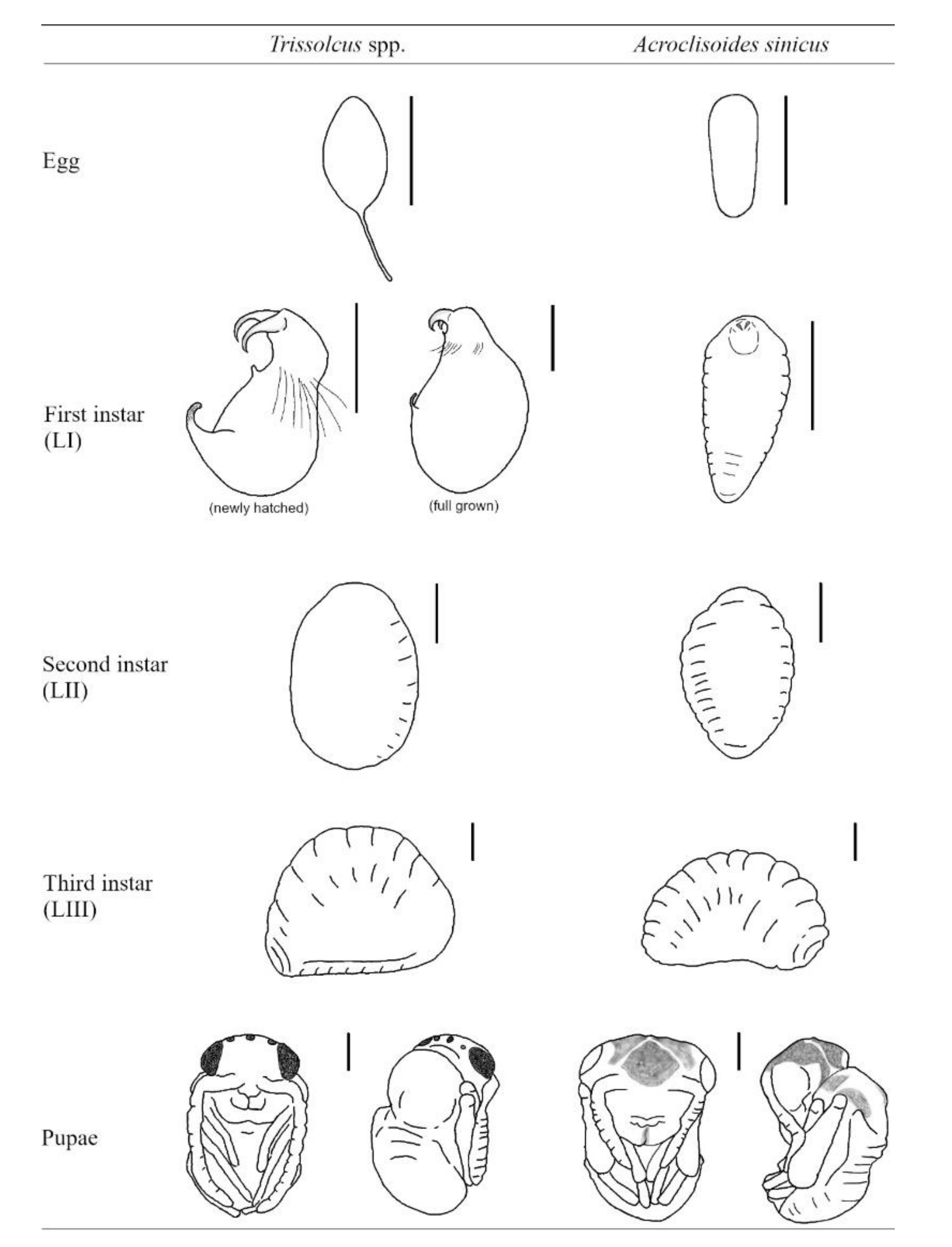
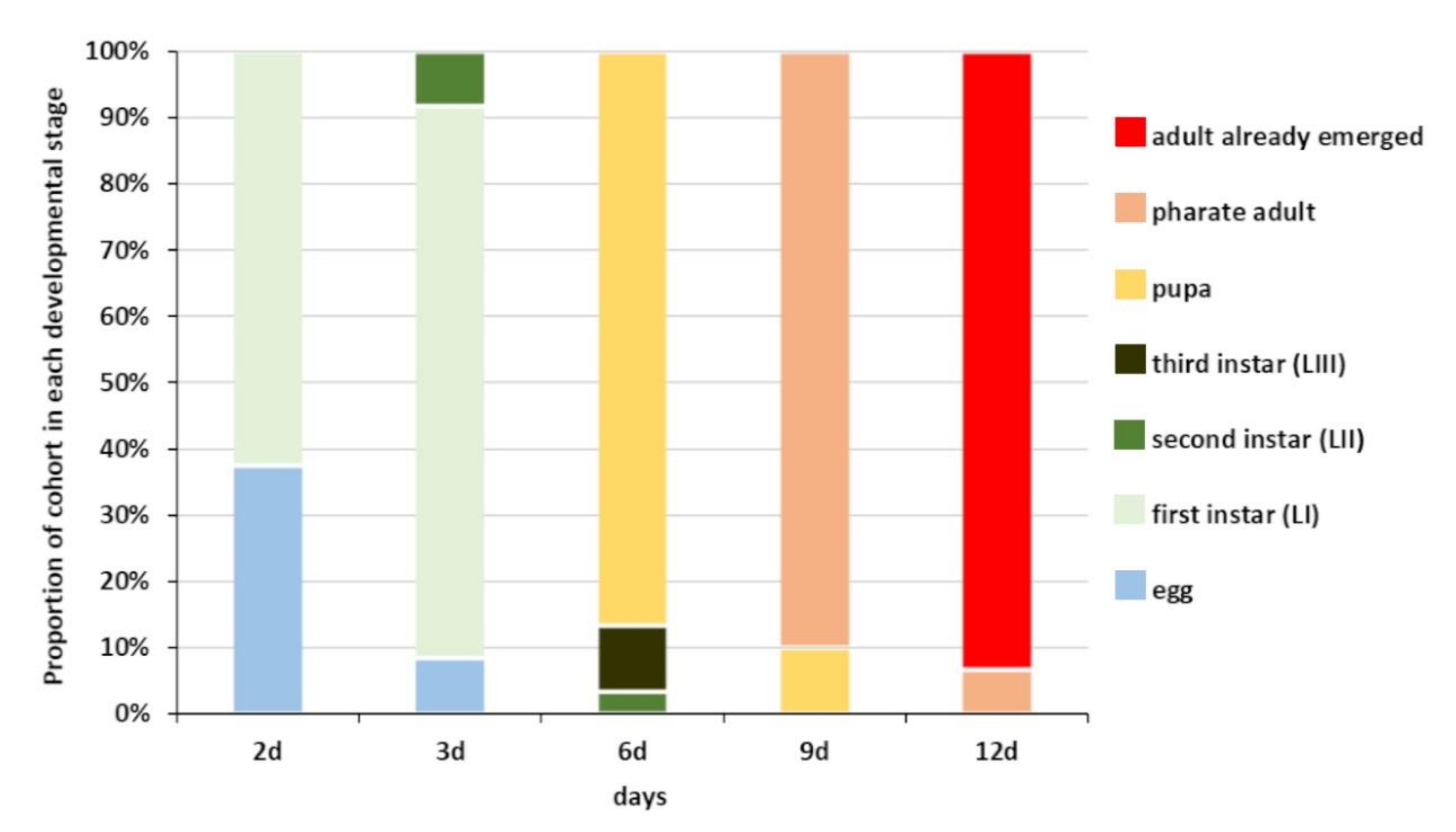
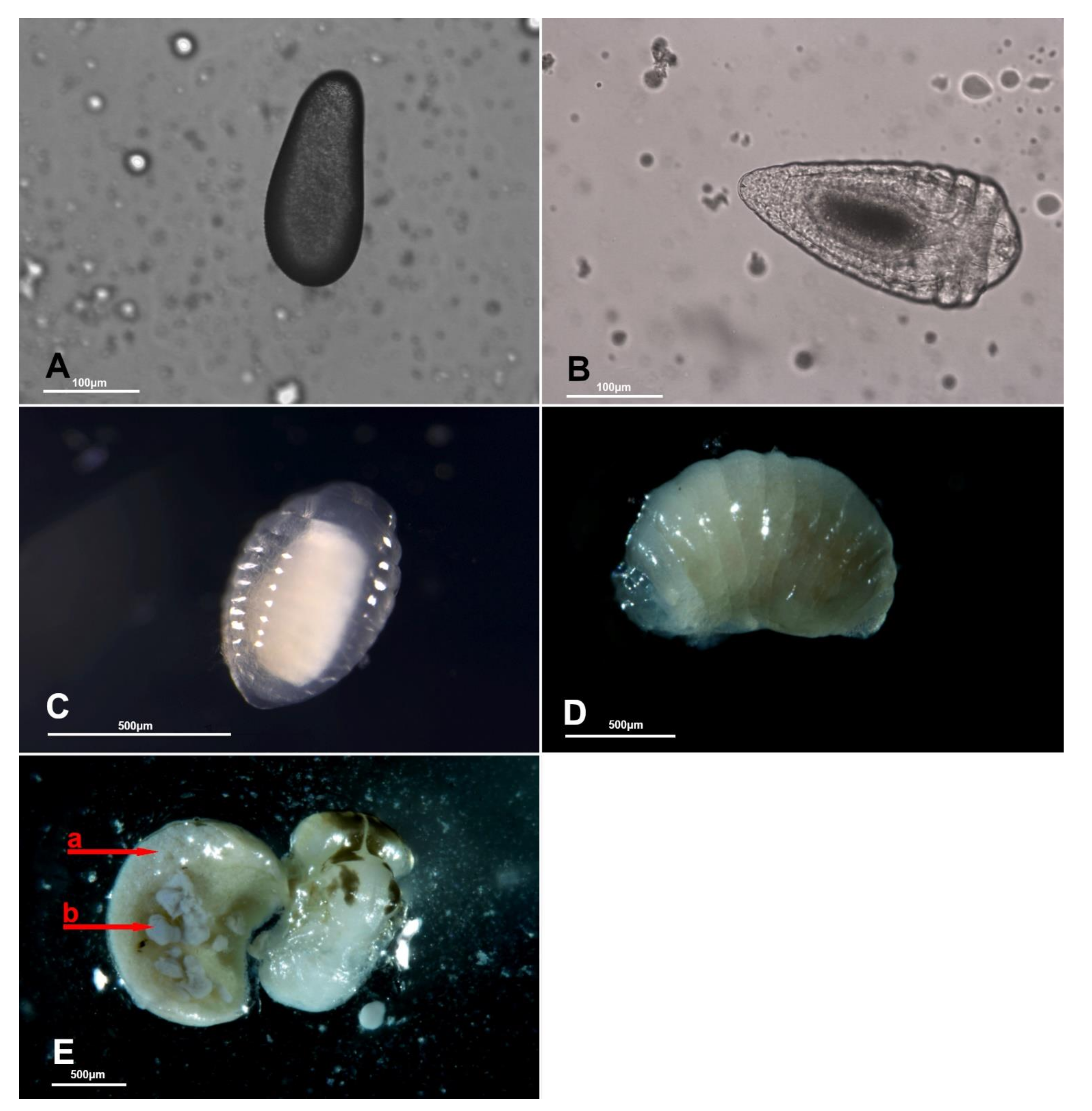
3.3. Acroclisoides sinicus Immature Development Studies in Europe
3.4. Development of Acroclisoides sinicus in Combinations with European Species
3.5. Acroclisoides sinicus Longevity Tests in Europe
3.6. Polyphenism and Female Egg Load of Acroclisoides sinicus
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethical Information
References
- Bouček, Z.; Heydon, S.L. Pteromalidae. In Annotated Keys to the Genera of Nearctic Chalcidoidea (Hymenoptera); Gibson, G.A.P., Huber, J.T., Woolley, J.B., Eds.; NRC Research Press: Ottawa, ON, Canada, 1997; pp. 541–692. [Google Scholar]
- Bouček, Z.; Rasplus, J.Y. Illustrated Key to West-Palearctic Genera of Pteromalidae (Hymenoptera: Chalcidoidea); Institut National de la Recherche Agronomique (INRA): Paris, France, 1991; pp. 1–141. [Google Scholar]
- Sureshan, P.M.; Narendran, T.C. The species of Metasteanus Walker and Acroclisoides Girault & Dodd (Hymenoptera: Chalcidoidea: Pteromalidae). Rec. Zool. Surv. India 2002, 100, 123–131. [Google Scholar]
- Grissell, E.E.; Smith, D.R. First report of Acroclisoides Girault and Dodd (Hymenoptera: Pteromalidae) in the Western Hemisphere, with description of a new species. Proc. Entomol. Soc. Washing. 2006, 108, 923–929. [Google Scholar]
- Noyes, J.S. Universal Chalcidoidea Database. Available online: http://www.nhm.ac.uk/chalcidoids (accessed on 8 October 2020).
- Rider, D.A. Family Pentatomidae. In Catalogue of the Heteroptera of the Palaearctic Region; Pentatomomorpha II; Aukema, B., Rieger, C., Eds.; The Netherland Entomological Society: Amsterdam, The Netherlands, 2006; Volume 5, pp. 233–402. [Google Scholar]
- Clarke, A.R.; Seymour, J.E. Two species of Acroclisoides Girault and Dodd (Hymenoptera: Pteromalidae) parasitic on Trissolcus basalis (Wollaston) (Hymenoptera: Scelionidae), a parasitoid of Nezara viridula (L.) (Hemiptera: Pentatomidae). Aust. J. Entomol. 1992, 31, 299–300. [Google Scholar] [CrossRef]
- Sabbatini Peverieri, G.; Mitroiu, M.-D.; Bon, M.-C.; Balusu, R.; Benvenuto, L.; Bernardinelli, I.; Fadamiro, H.; Falagiarda, M.; Fusu, L.; Grove, E.; et al. Surveys of stink bug egg parasitism in Asia, Europe and North America, morphological taxonomy, and molecular analysis reveal the Holarctic distribution of Acroclisoides sinicus (Huang & Liao) (Hymenoptera, Pteromalidae). J. Hym. Res. 2019, 74, 123–151. [Google Scholar]
- Huang, D.W.; Liao, D.X. A new genus and a new species of Pteromalidae (Hymenoptera Chalcidoidea). Acta Entomol. Sinica 1988, 31, 426–428. [Google Scholar]
- Xiao, H.; Huang, D.W. Taxonomic studies on genus Acroclisoides Girault and Dodd from China (Hymenoptera: Pteromalidae). Acta Zool. Sinica 2000, 25, 98–99. [Google Scholar]
- Ko, G.H.; Park, D.Y.; Lee, J.W. Taxonomic review of subfamily Pteromalinae (Hymenoptera, Chalcidoidea) with 25 newly recorded species in South Korea. J. Asia Pacific Biodiv. 2018, 11, e87–e122. [Google Scholar] [CrossRef]
- Moraglio, S.T.; Tortorici, F.; Pansa, M.G.; Castelli, G.; Pontini, M.; Scovero, S.; Visentin, S.; Tavella, L. A 3-year survey on parasitism of Halyomorpha halys by egg parasitoids in northern Italy. J. Pest Sci. 2019, 9, 1–12. [Google Scholar]
- Sabbatini Peverieri, G.; Talamas, E.; Bon, M.C.; Marianelli, L.; Bernardinelli, I.; Malossini, G.; Benvenuto, L.; Roversi, P.F.; Hoelmer, K. Two Asian egg parasitoids of Halyomorpha halys (Stal) (Hemiptera, Pentatomidae) emerge in northern Italy: Trissolcus mitsukurii (Ashmead) and Trissolcus japonicus (Ashmead) (Hymenoptera, Scelionidae). J. Hym. Res. 2018, 67, 37–53. [Google Scholar] [CrossRef]
- Stahl, J.; Tortorici, F.; Pontini, M.; Bon, M.C.; Hoelmer, K.; Marazzi, C.; Tavella, L.; Haye, T. First discovery of adventive populations of Trissolcus japonicus (Ashmead) in Europe. J. Pest Sci. 2019, 92, 371–379. [Google Scholar] [CrossRef]
- Onagbola, E.O.; Fadamiro, H.J. Morphology and development of Pteromalus cerealellae (Ashmead) (Hymenoptera: Pteromalidae) on Callosobruchus maculatus (F.) (Coleoptera: Chrysomelidae). BioControl 2008, 53, 737–750. [Google Scholar] [CrossRef]
- Tormos, J.; Beitia, F.; Böckmann, E.A.; Asís, J.D. The preimaginal stages and development of Spalangia cameroni Perkins (Hymenoptera: Pteromalidae) on Ceratitis capitata (Wiedemann) (Diptera: Tephritidae). Micron 2009, 40, 646–658. [Google Scholar] [CrossRef]
- Talamas, E.J.; Buffington, M.L.; Hoelmer, K. Revision of Palearctic Trissolcus Ashmead (Hymenoptera, Scelionidae). J. Hym. Res. 2017, 56, 3–185. [Google Scholar]
- Tillman, P.G.; Toews, M.; Blaauw, B.; Sial, A.; Cottrell, T.E.; Talamas, E.; Buntin, D.; Joseph, S.; Balusu, R.; Fadamiro, H.; et al. Parasitism and predation of sentinel eggs of the invasive brown marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae) in the southeastern US. Biol. Control 2020, 145, 104247. [Google Scholar] [CrossRef]
- Gerling, D. The developmental biology of Telenomus remus Nixon (Hym., Scelionidae). Bull. Entomol. Res 1972, 61, 385–388. [Google Scholar] [CrossRef]
- Navasero, R.C.; Oatman, E.R. Life history, immature morphology and adult behavior of Telenomus solitus [Hymenoptera: Scelionidae]. Entomophaga 1989, 34, 165–177. [Google Scholar] [CrossRef]
- Sousa, J.M. Development of Tiphodytes gerriphagus (Hymenoptera: Scelionidae) in Limnoporus dissortis eggs (Hemiptera: Gerridae). Can. Entomol. 1999, 131, 219–228. [Google Scholar] [CrossRef]
- Volkoff, N.; Colazza, S. Growth patterns of teratocytes in the immature stages of Trissolcus basalis (Woll.) (Hymenoptera: Scelionidae), an egg parasitoid of Nezara viridula (L.) (Heteroptera: Pentatomidae). Int. J. Morphol. Embryol. 1992, 21, 323–336. [Google Scholar] [CrossRef]
- Sabbatini-Peverieri, G.; Giovannini, L.; Benvenuti, C.; Madonni, L.; Hoelmer, K.A.; Roversi, P.F. Characteristics of the meconia of European egg parasitoids of Halyomorpha halys. J. Hym. Res. 2020, 77, 187–201. [Google Scholar] [CrossRef]
- Abram, P.K.; Parent, J.-P.; Brodeur, J.; Boivin, G. Size-induced phenotypic reaction norms in a parasitoid wasp: An examination of life-history and behavioural traits. Biol. J. Linn. Soc. 2015, 117, 620–632. [Google Scholar] [CrossRef]
- Botch, P.S.; Delfosse, E.S. Host-acceptance behavior of Trissolcus japonicus (Hymenoptera: Scelionidae) reared on the invasive Halyomorpha halys (Heteroptera: Pentatomidae) and nontarget species. Environ. Entomol. 2018, 47, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Clausen, C.P. Entomophagous Insects; McGraw Hill: New York, NY, USA, 1940; pp. 1–688. [Google Scholar]
- Steidle, J.L.M.; VanLoon, J.J.A. Chemoecology of parasitoid and predator oviposition behaviour. In Chemoecology of Insect Eggs and Egg Deposition; Hilker, M., Meiners, T., Eds.; Publisher Blackwell: Berlin, Germany, 2002; pp. 291–317. [Google Scholar]
- Viggiani, G. Lotta Biologica e Integrata Nella Difesa Fitosanitaria; Liguori Editore: Napoli, Italy, 1994; pp. 1–517. [Google Scholar]
- Konopka, J.K.; Haye, T.; Gariepy, T.D.; McNeil, J.N. Possible coexistence of native and exotic parasitoids and their impact on control of Halyomorpha halys. J. Pest. Sci. 2017, 90, 1119–1125. [Google Scholar] [CrossRef]
- Stahl, J.M.; Babendreier, D.; Foti, M.C.; Colazza, S.; Haye, T. Intrinsic competition between two European egg parasitoids of the brown marmorated stink bug. J. Appl. Entomol. 2020, 144, 1–9. [Google Scholar] [CrossRef]
- Hokyo, F.; Kiritani, K.; Nakasuji, F.; Shiga, M. Comparative biology of the two scelionid egg parasites of Nezara viridula L. (Hemiptera: Pentatomidae). Appl. Entomol. Zool. 1966, 1, 94–102. [Google Scholar] [CrossRef][Green Version]
- Yang, Z.-Q.; Yao, Y.-X.; Qiu, L.-F.; Li, Z.-X. A new species of Trissolcus (Hymenoptera: Scelionidae) parasitizing eggs of Halyomorpha halys (Heteroptera: Pentatomidae) in China with comments on its biology. Ann. Entomol. Soc. Am. 2009, 102, 39–47. [Google Scholar] [CrossRef]
- Sabbatini Peverieri, G.; Furlan, P.; Simoni, S.; Strong, W.B.; Roversi, P.F. Laboratory evaluation of Gryon pennsylvanicum (Ashmead) (Hymenoptera, Platygastridae) as biological control agent of Leptoglossus occidentalis Heidemann (Heteroptera, Coreidae). Biol. Control 2012, 61, 104–111. [Google Scholar] [CrossRef]
- Sabbatini-Peverieri, G.; Dieckhoff, C.; Giovannini, L.; Marianelli, L.; Roversi, P.F.; Hoelmer, K. Rearing Trissolcus japonicus and Trissolcus mitsukurii for biological control of Halyomorpha halys. Insects 2020, 11, 787. [Google Scholar] [CrossRef]
- Stahl, J.M.; Babendreier, D.; Haye, T. Life history of Anastatus bifasciatus, a potential biological control agent of the brown marmorated stink bug in Europe. Biol. Control 2019, 129, 178–186. [Google Scholar] [CrossRef]
- Arakawa, R.; Miura, M.; Fujita, M. Effects of host species on the body size, fecundity, and longevity of Trissolcus mitsukurii (Hymenoptera: Scelionidae), a solitary egg parasitoid of stink bugs. Appl. Entomol. Zool. 2004, 39, 177–181. [Google Scholar] [CrossRef]
- Boyle, S.M.; Weber, D.C.; Hough-Goldstein, J.; Hoelmer, K.A. Parental host species affects behavior and parasitism by the pentatomid egg parasitoid, Trissolcus japonicus (Hymenoptera: Scelionidae). Biol. Control 2020, 149, 104324. [Google Scholar] [CrossRef]
- Nofemela, R.S. The effect of obligate hyperparasitoids on biological control: Differential vulnerability of primary parasitoids to hyperparasitism can mitigate trophic cascades. Biol. Control 2013, 65, 218. [Google Scholar] [CrossRef][Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
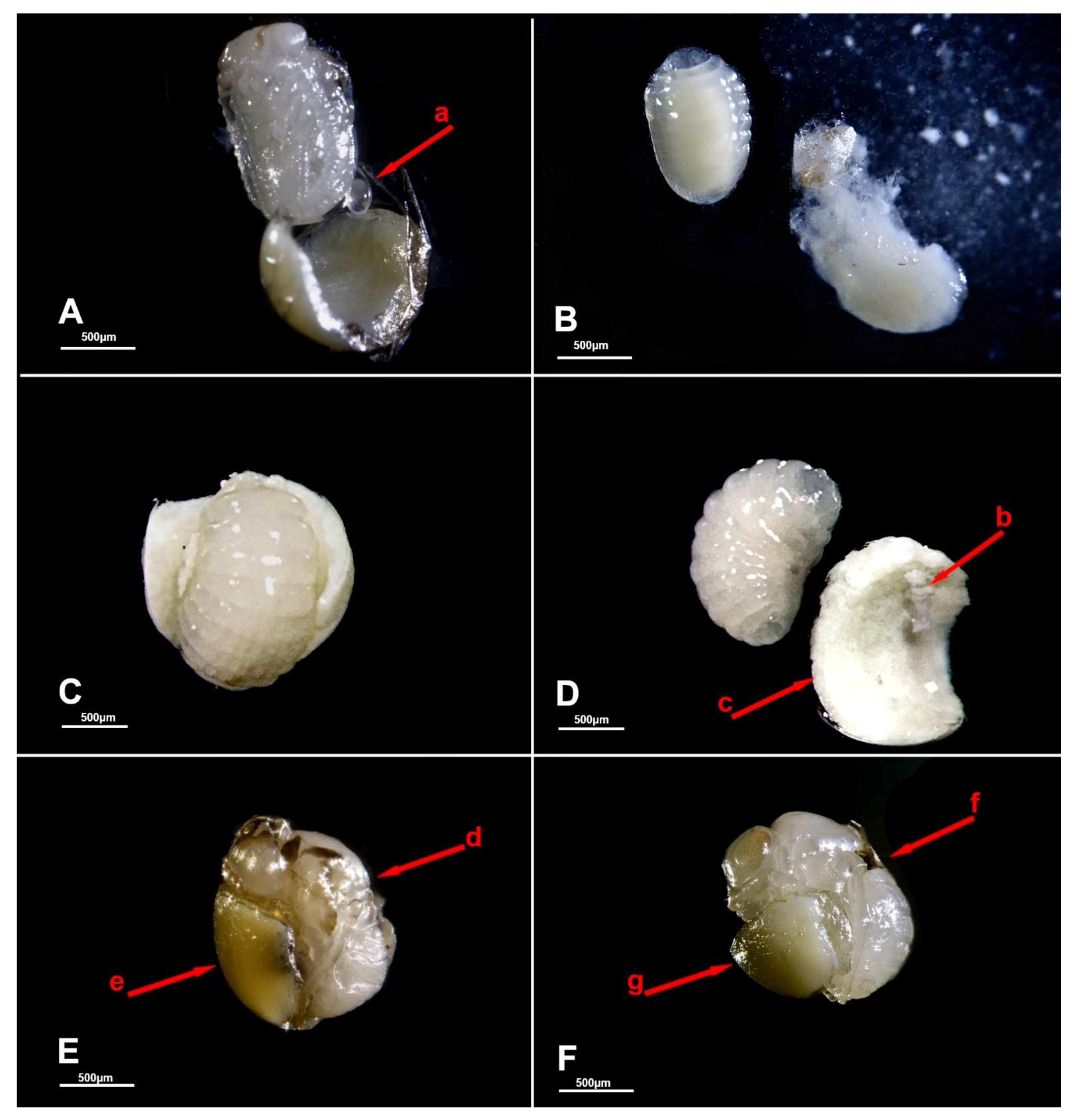{kind=link}
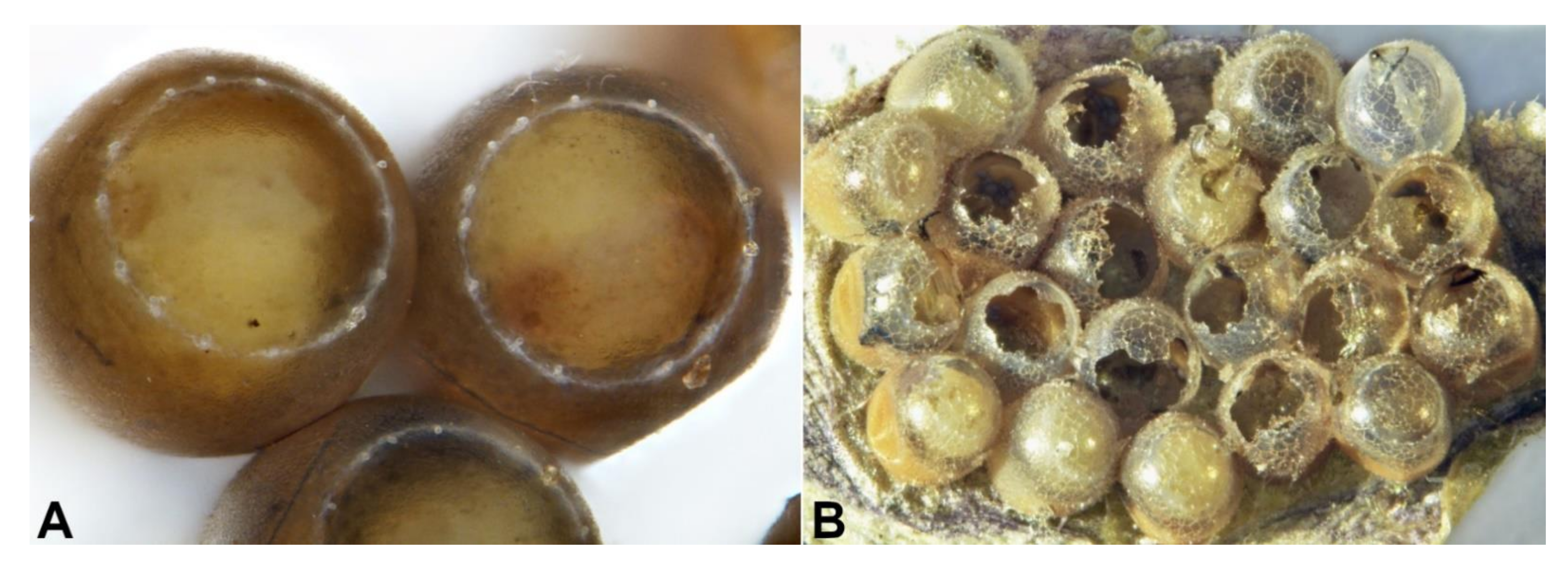{kind=link}
{kind=link}
| Trophic Chain | Pentatomid Eggs | Primary Parasitization | Aged Primary Parasitized Host | Hyperparasitization | Hyperparasitized Egg Masses | ||||
|---|---|---|---|---|---|---|---|---|---|
 |  |  |  |  |  |  |  |  | |
| Fresh (<24 h) H. halys Egg Masses | Trissolcusspp. Parasitize Host Eggs | Trissolcusspp. Juveniles Develop Inside Its Host | A. sinicusParasitize Its Host Trissolcus spp. inside Eggs of H. halys | A. sinicusJuveniles Develop by Feeding on Its Host Trissolcus spp. inside H. halys Eggs | |||||
| Parasitization tests | H. halys egg masses | Exposure to parasitization for 24 h | Ageing for 1, 3, 5, 7, and 9 days | Exposure for 24 (all treatments) and 72 h (except 9 days ageing) and 216 h (only for 1 day) | Rearing egg masses to emergence of adult parasitoids | ||||
| Juvenile development study | H. halys egg masses | Exposure to parasitization for 24 h | Ageing for 5 days | Exposure for 48 h and 72 h | Egg dissection at the end of exposure (48 h = “2d”; 72 h = “3d”) | ||||
| Exposure for 72 h | Eggs rearing for additional 3 (“6d”), 6 (“9d”), and 9 days (“12d”) before dissection |
| Exposure Combination (Egg Mass Age/Exposure Time) | T. japonicus Development Stage | n. of Eggs/Egg Mass | Adult Emergence Rate (%) | Dead H. halys Eggs | Sex Ratio (% Female) | ||
|---|---|---|---|---|---|---|---|
| T. japonicus | A. sinicus | T. japonicus | A. sinicus | ||||
| 1 day × 24 h | Egg | 27.00 ± 0.70 | 93.68 ± 4.22 | 0.00 | 6.46 ± 4.23 | 89.84 ± 2.74 | - |
| 1 day × 72 h | Egg–LI | 27.50 ± 0.28 | 99.07 ± 0.92 | 0.00 | 1.85 ± 0.00 | 92.65 ± 2.65 | - |
| 3 days × 24 h | LI | 27.50 ± 0.50 | 97.11 ± 2.88 | 0.00 | 2.88 ± 0.00 | 92.27 ± 1.96 | - |
| 3 days × 72 h | LI–LII–LIII | 25.25 ± 0.85 | 91.65 ± 4.82 | 0.00 | 8.34 ± 4.82 | 95.99 ± 1.53 | - |
| 5 days × 24 h | LIII | 27.75 ± 0.25 | 94.60 ± 3.08 | 0.00 | 5.39 ± 3.08 | 94.32 ± 1.81 | - |
| 5 days × 72 h | LIII–pupa | 28.00 ± 0.00 | 59.82 ± 7.95 | 19.64 ± 5.15 | 20.24 ± 1.79 | 88.10 ± 4.76 | 80.44 ± 8.33 |
| 7 days × 24 h | Pupa | 26.75 ± 0.75 | 98.07 ± 1.92 | 0.00 | 1.92 ± 0.00 | 91.38 ± 2.56 | - |
| 7 days × 72 h | Pupa–pharate adult | 27.40 ± 0.60 | 89.74 ± 2.88 | 1.43 ± 1.43 | 8.11 ± 1.87 | 89.29 ± 1.24 | 50.00 ± 0.00 |
| 9 days × 24 h | Pharate adult | 27.50 ± 0.57 | 92.75 ± 3.73 | 0.00 | 7.24 ± 3.73 | 95.19 ± 0.79 | - |
| 1 day × 216 h | From egg to pharate adult | 25.56 ± 1.50 | 46.48 ± 4.95 | 23.21 ± 11.08 | 30.30 ± 6.46 | 53.50 ± 8.53 | 66.43 ± 17.34 |
| Exposure Combination (Egg Mass Age/Exposure Time) | T. mitsukurii Development Stage | n. of Eggs/Egg Mass | Adult Emergence Rate (%) | Dead H. halys Eggs | Sex Ratio (% Female) | ||
|---|---|---|---|---|---|---|---|
| T. mitsukurii | A. sinicus | T. mitsukurii | A. sinicus | ||||
| 1 day × 24 h | Egg | 27.50 ± 0.51 | 100.00 | 0.00 | 0.00 | 61.57 ± 13.43 | - |
| 1 day × 72 h | Egg–LI | 27.00 ± 0.00 | 100.00 | 0.00 | 0.00 | 66.07 ± 19.64 | - |
| 3 days × 24 h | LI | 27.50 ± 0.52 | 100.00 | 0.00 | 0.00 | 52.98 ± 13.69 | - |
| 3 days × 72 h | LI–LII–LIII | 26.80 ± 0.73 | 100.00 | 0.00 | 0.00 | 87.42 ± 5.07 | - |
| 5 days × 24 h | LIII | 27.50 ± 0.57 | 95.01 ± 1.35 | 0.00 | 4.98 ± 2.88 | 79.62 ± 4.76 | - |
| 5 days × 72 h | LIII–pupa | 25.80 ± 0.66 | 16.90 ± 0.00 | 33.01 ± 9.14 | 50.06 ± 10.56 | 100.00 | 83.79 ± 7.95 |
| 7 days × 24 h | Pupa | 28.00 ± 0.57 | 77.90 ± 18.58 | 1.23 ± 0.00 | 22.09 ± 18.58 | 77.90 ± 18.58 | - |
| 7 days × 72 h | Pupa–pharate adult | 26.20 ± 1.11 | 47.21 ± 18.32 | 27.00 ± 10.51 | 26.25 ± 17.51 | 96.87 ± 1.99 | 61.79 ± 17.25 |
| 9 days × 24 h | Pharate adult | 27.50 ± 0.57 | 100.00 | 0.00 | 0.00 | 93.69 ± 1.13 | - |
| 1 day × 216 h | From egg to pharate adult | 25.40 ± 1.20 | 14.40 ± 0.00 | 85.60 ± 14.40 | 0.00 | 94.44 ± 0.00 | 90.69 ± 1.76 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giovannini, L.; Sabbatini-Peverieri, G.; Tillman, P.G.; Hoelmer, K.A.; Roversi, P.F. Reproductive and Developmental Biology of Acroclisoides sinicus, a Hyperparasitoid of Scelionid Parasitoids. Biology 2021, 10, 229. https://doi.org/10.3390/biology10030229
Giovannini L, Sabbatini-Peverieri G, Tillman PG, Hoelmer KA, Roversi PF. Reproductive and Developmental Biology of Acroclisoides sinicus, a Hyperparasitoid of Scelionid Parasitoids. Biology. 2021; 10(3):229. https://doi.org/10.3390/biology10030229
Chicago/Turabian StyleGiovannini, Lucrezia, Giuseppino Sabbatini-Peverieri, Patricia Glynn Tillman, Kim Alan Hoelmer, and Pio Federico Roversi. 2021. "Reproductive and Developmental Biology of Acroclisoides sinicus, a Hyperparasitoid of Scelionid Parasitoids" Biology 10, no. 3: 229. https://doi.org/10.3390/biology10030229
APA StyleGiovannini, L., Sabbatini-Peverieri, G., Tillman, P. G., Hoelmer, K. A., & Roversi, P. F. (2021). Reproductive and Developmental Biology of Acroclisoides sinicus, a Hyperparasitoid of Scelionid Parasitoids. Biology, 10(3), 229. https://doi.org/10.3390/biology10030229