
The Pharmacological Inhibition of CaMKII Regulates Sodium Chloride Cotransporter Activity in mDCT15 Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. SiRNA Transduction
2.3. Immunofluorescence
2.4. Mice
2.5. Immunoprecipitation
2.6. SDS-PAGE and Western Blotting
2.7. Assessment of the NCC Function in the Cells
2.8. Statistical Analysis
3. Results
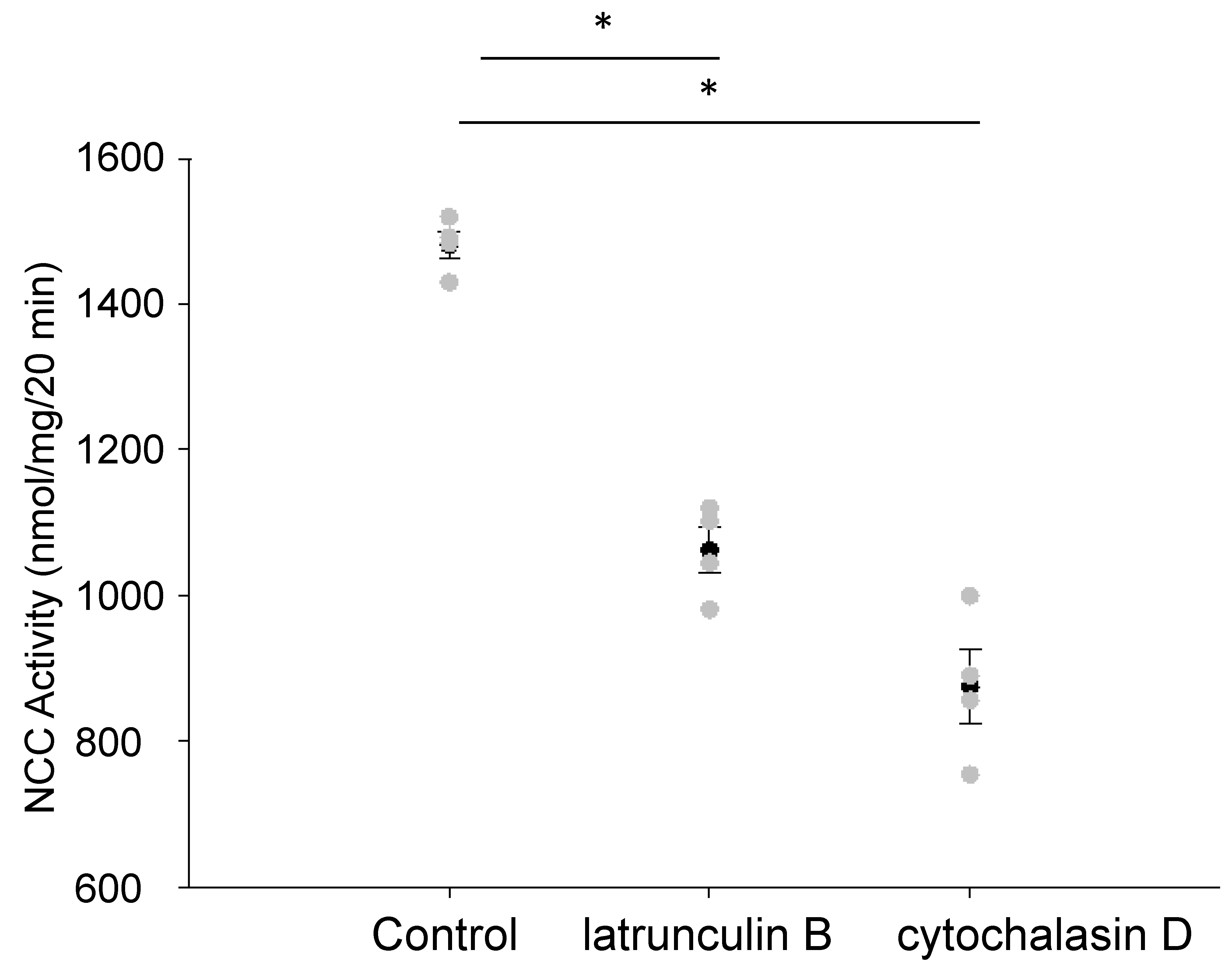
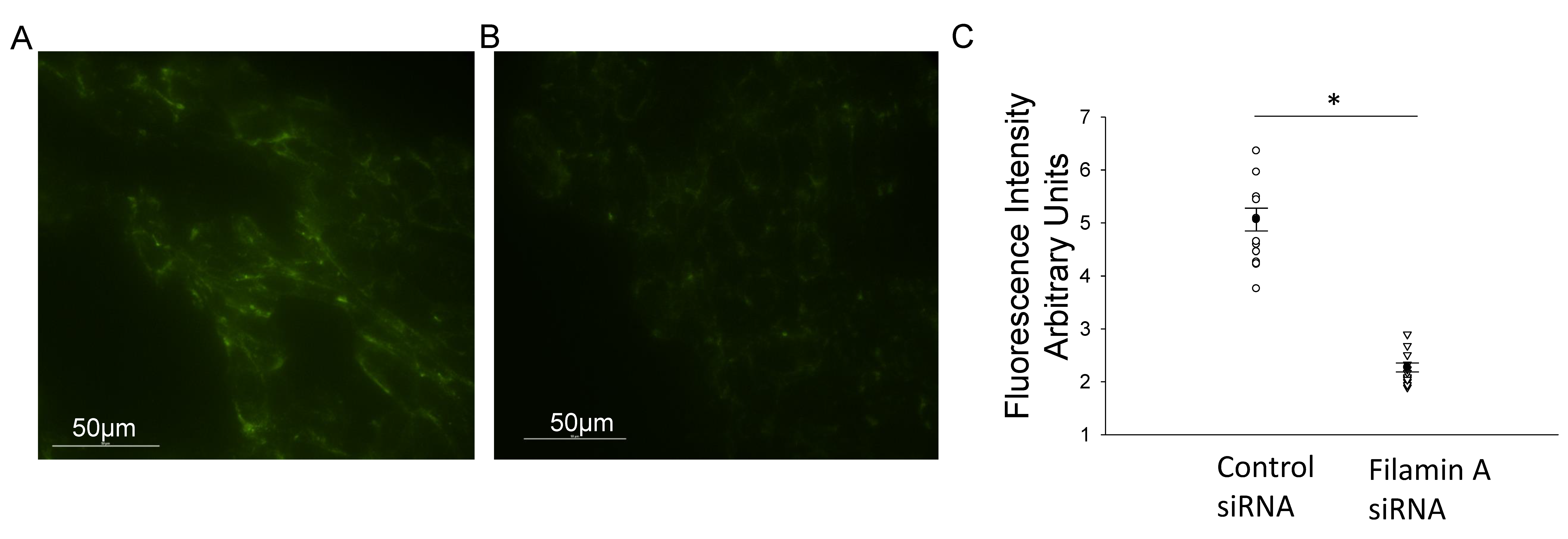
3.1. Disruption of the Actin Cytoskeleton Decreases NCC Activity in mDCT15 Cells
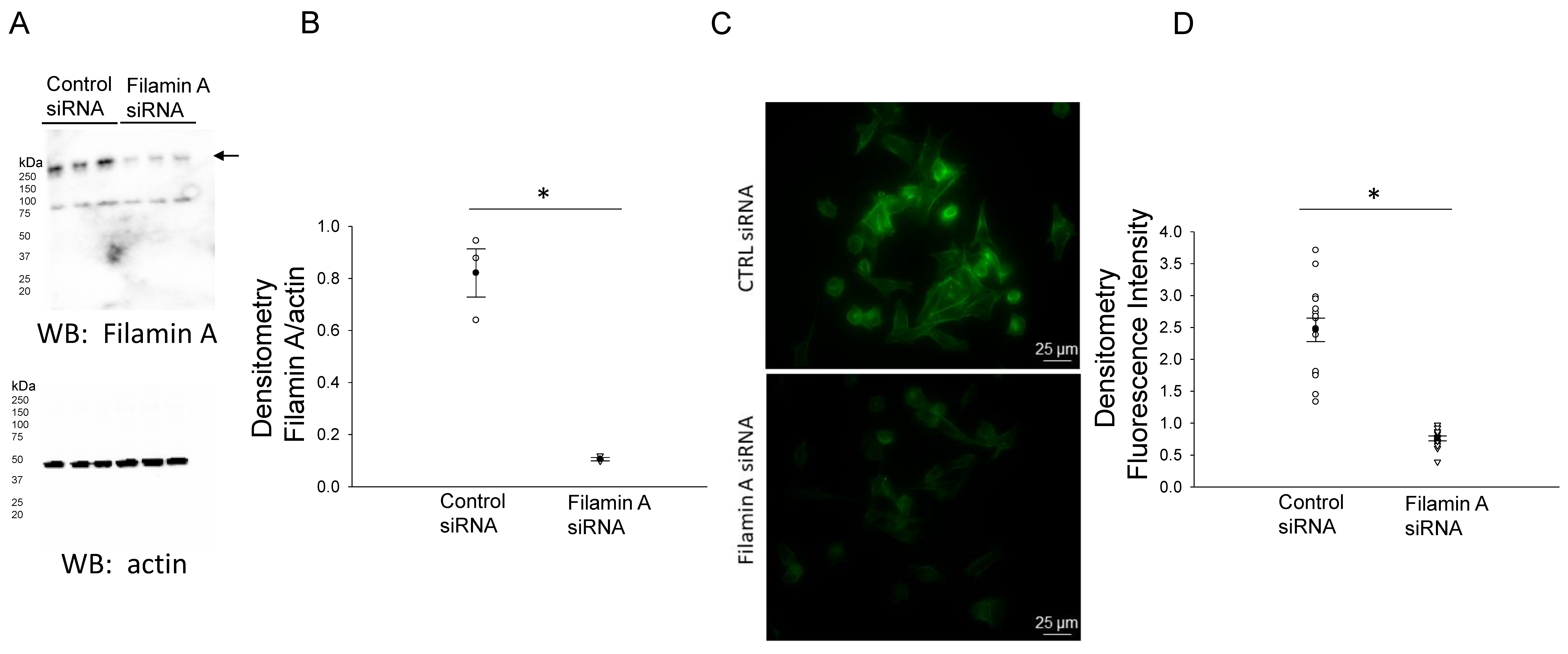
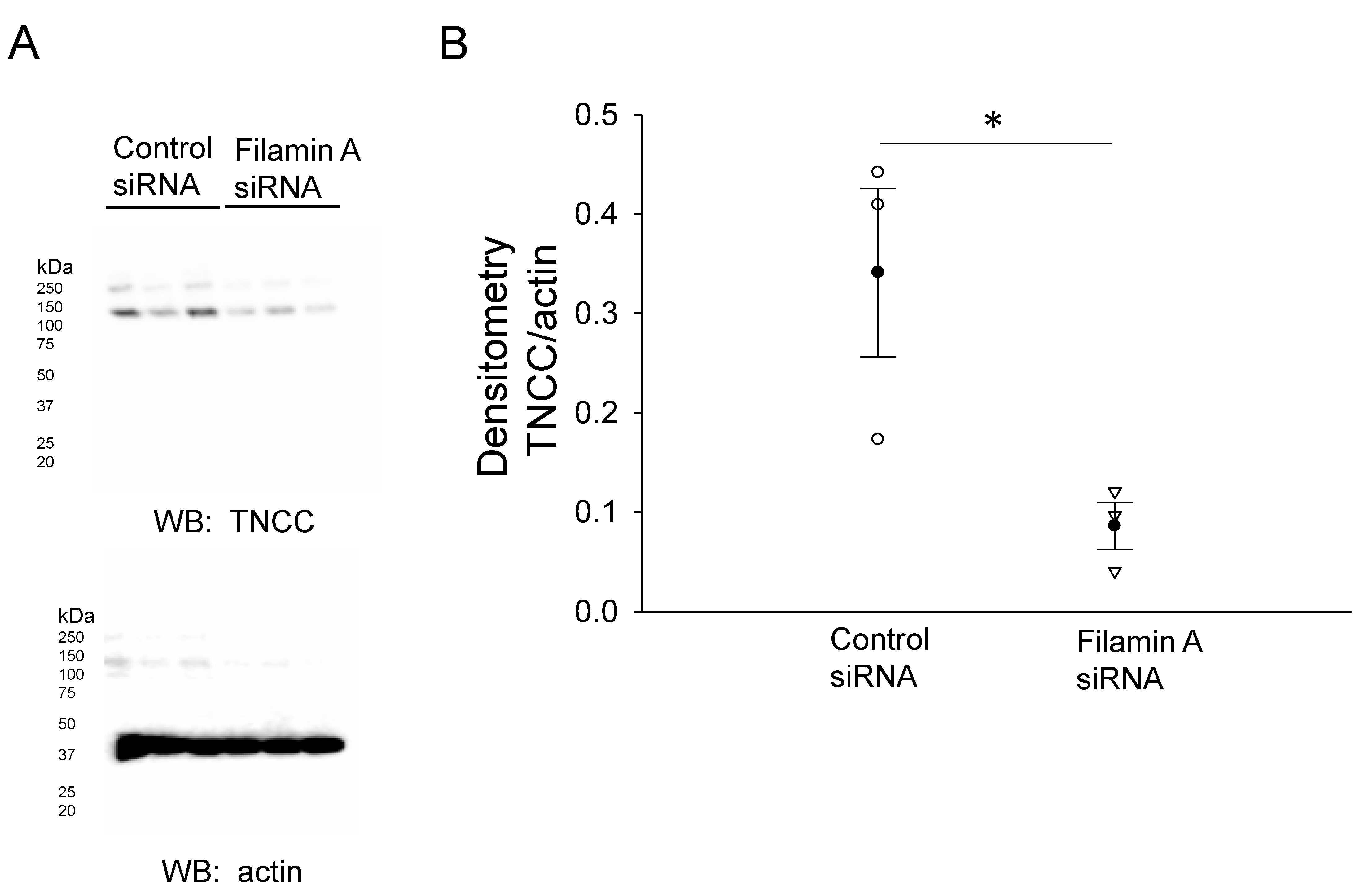
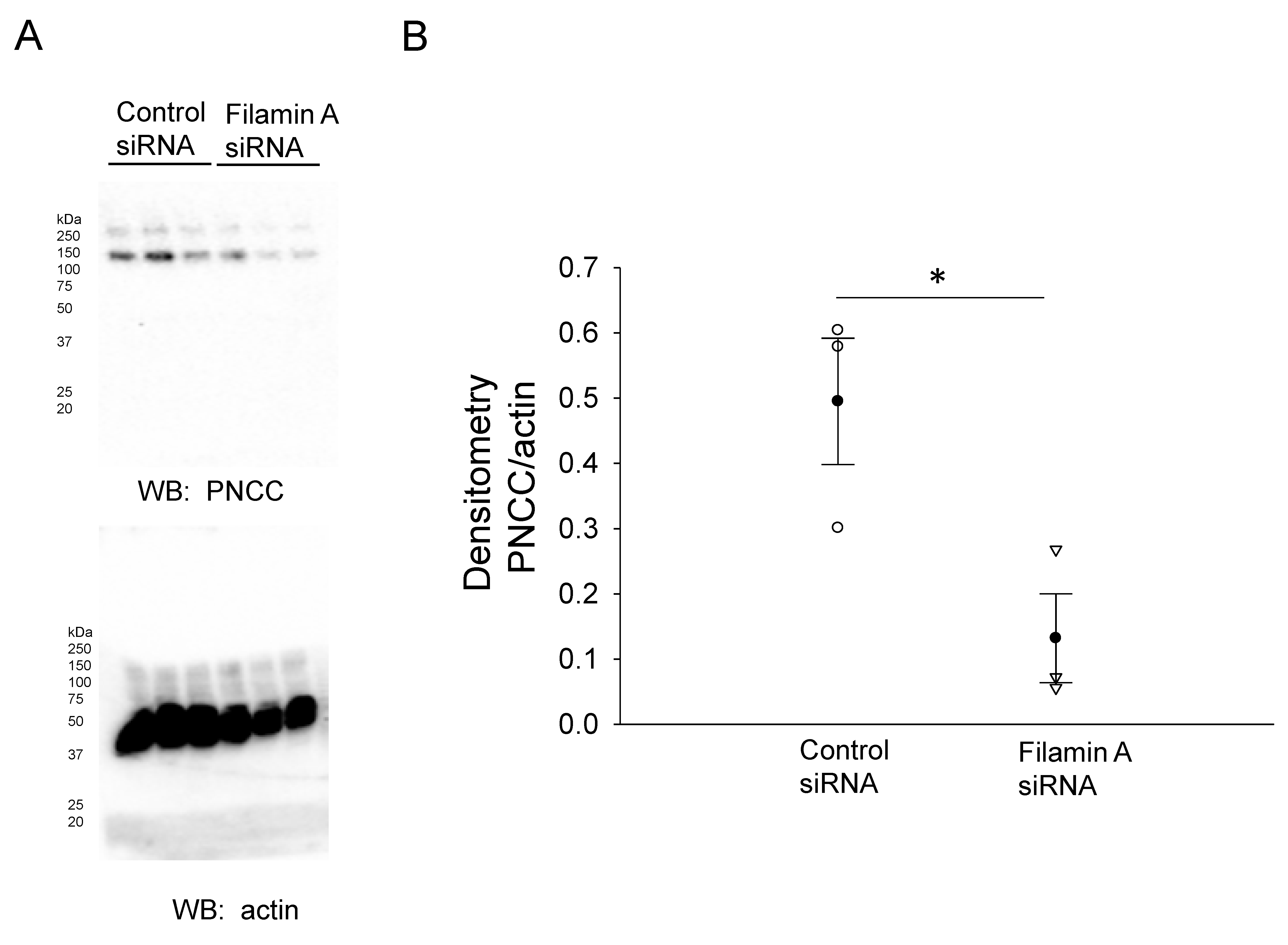
3.2. siRNA-Mediated Knockdown of Filamin A Reduces the Total NCC and Phospho-NCC Expression in mDCT15 Cells
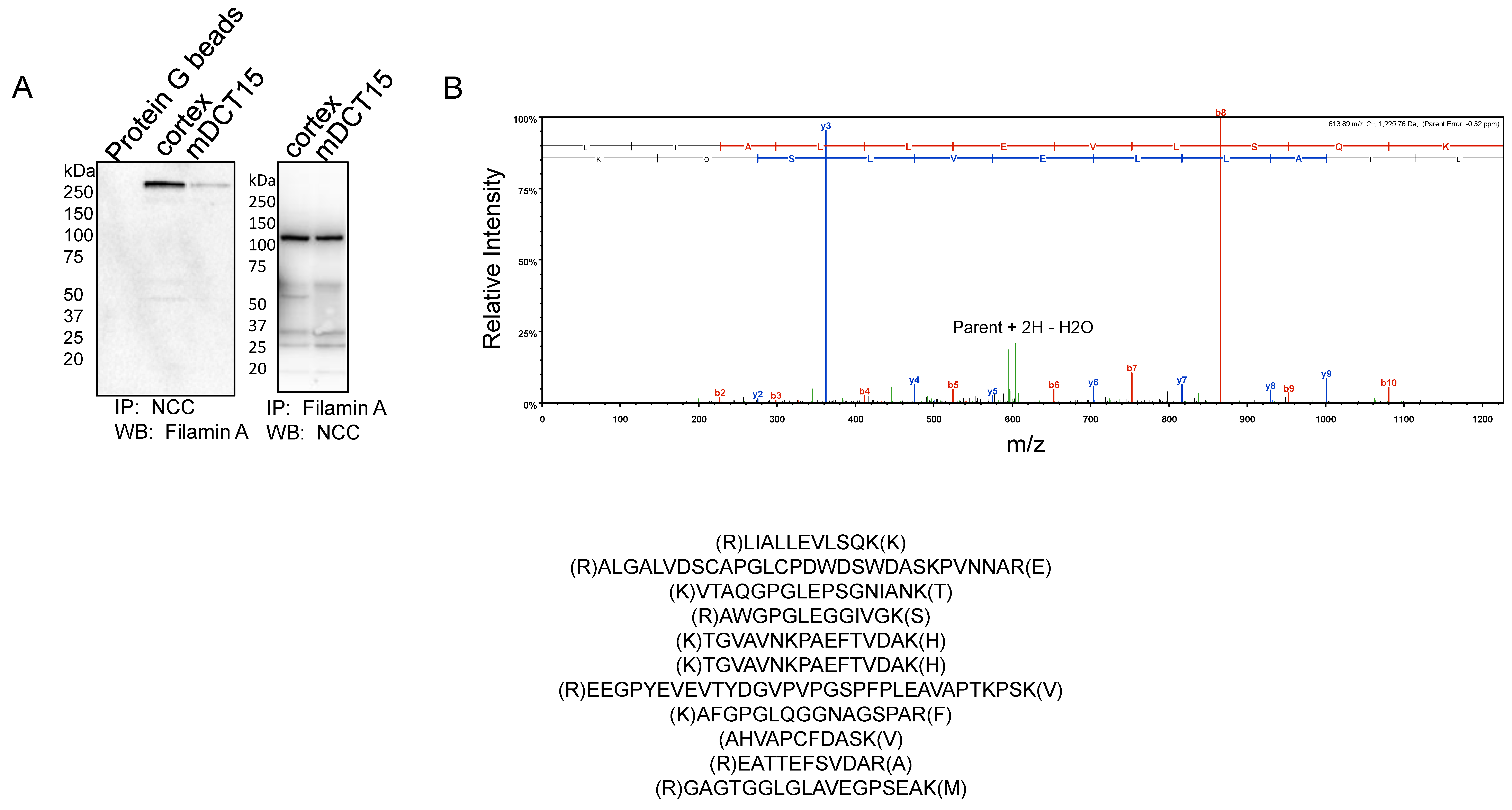
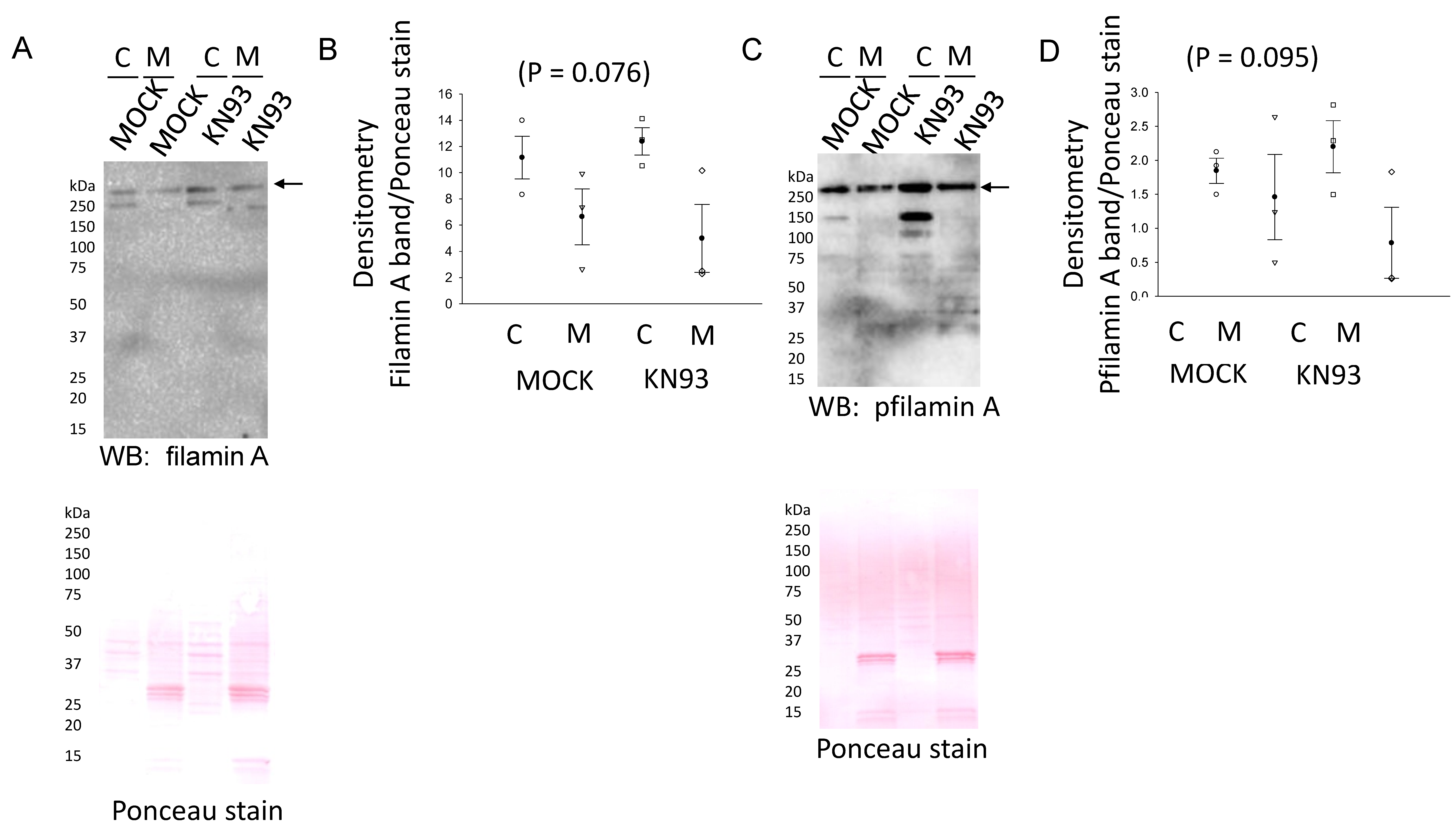
3.3. NCC and Filamin A Directly Interact in mDCT15 Cells
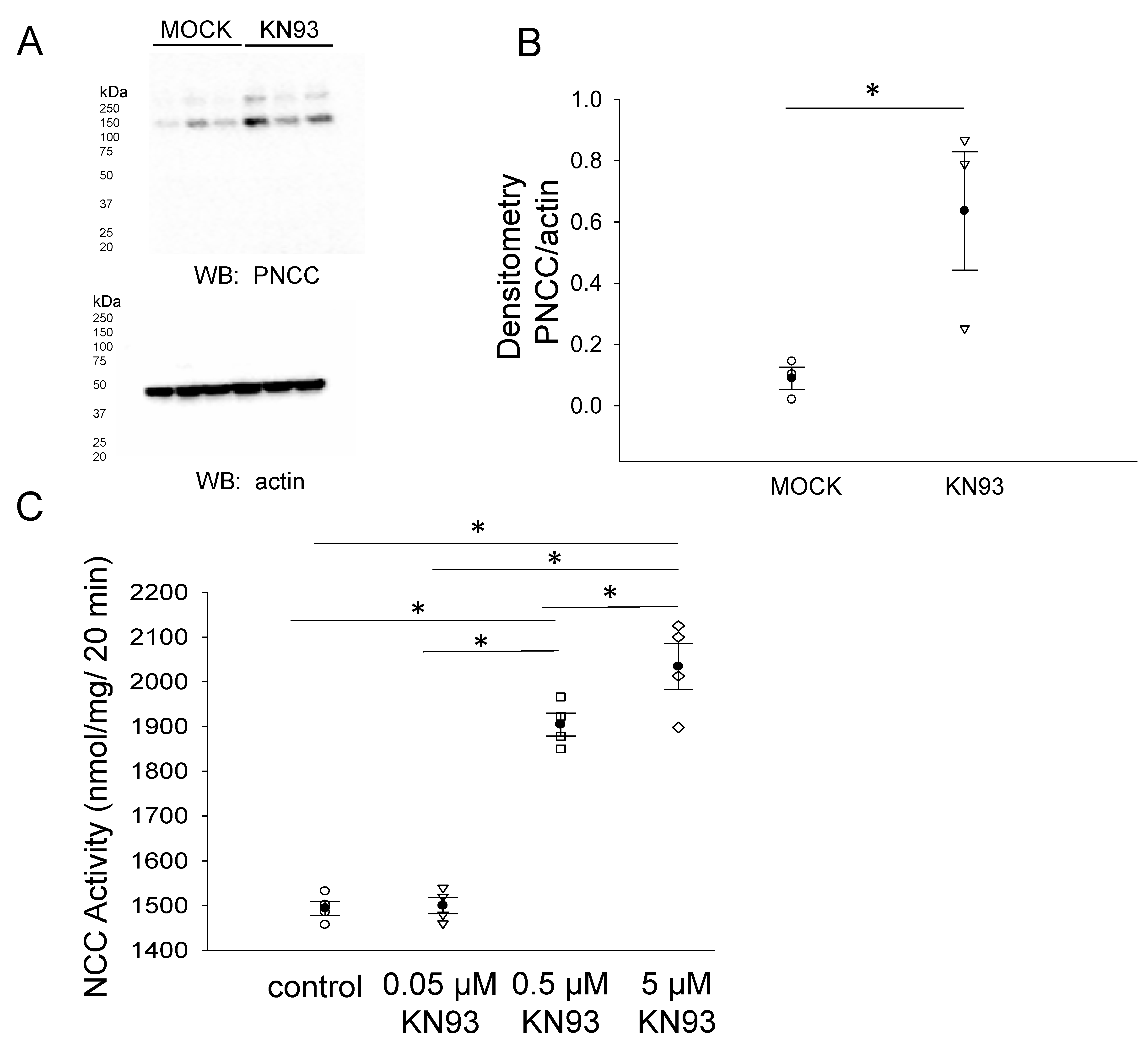
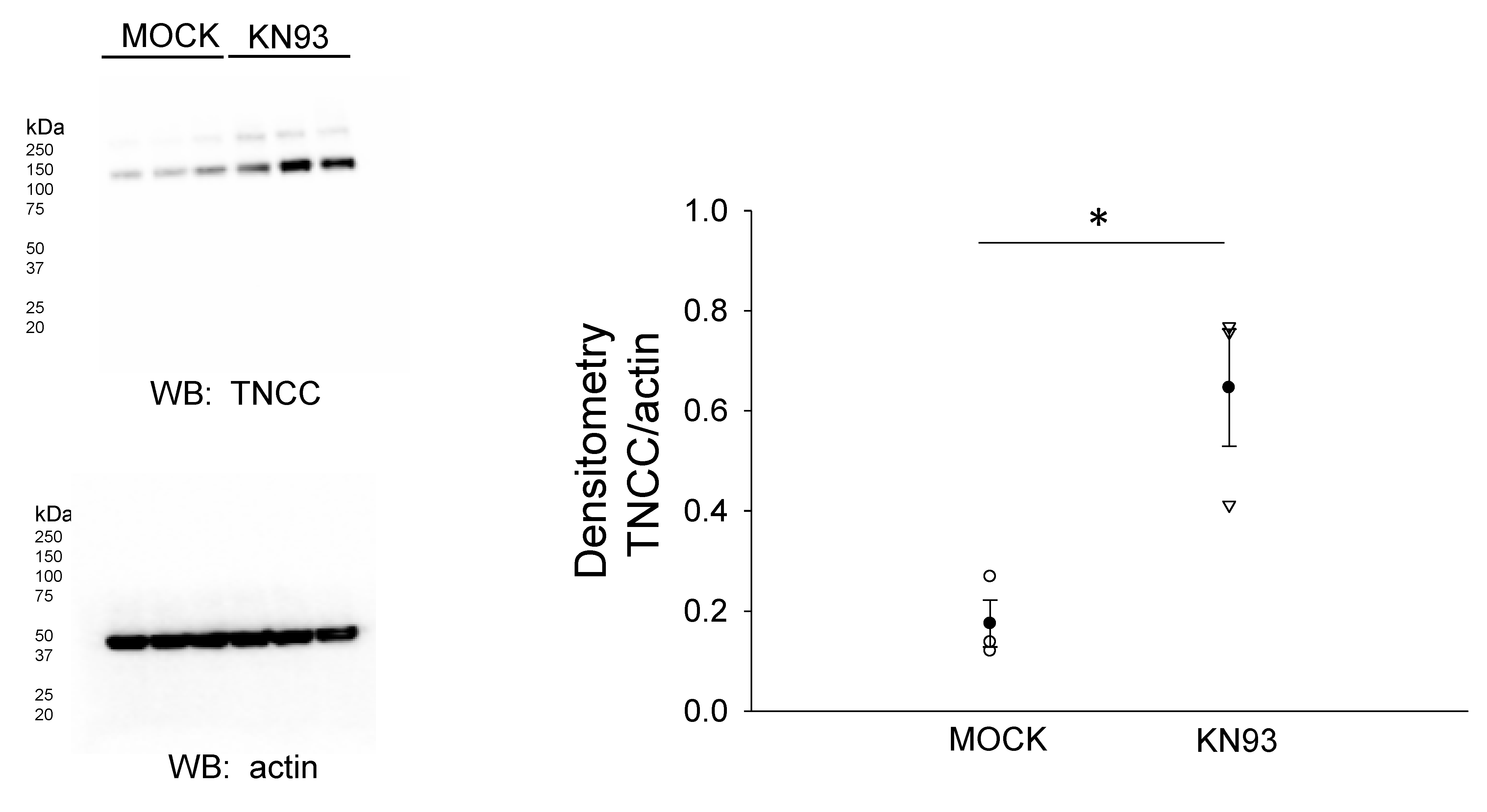
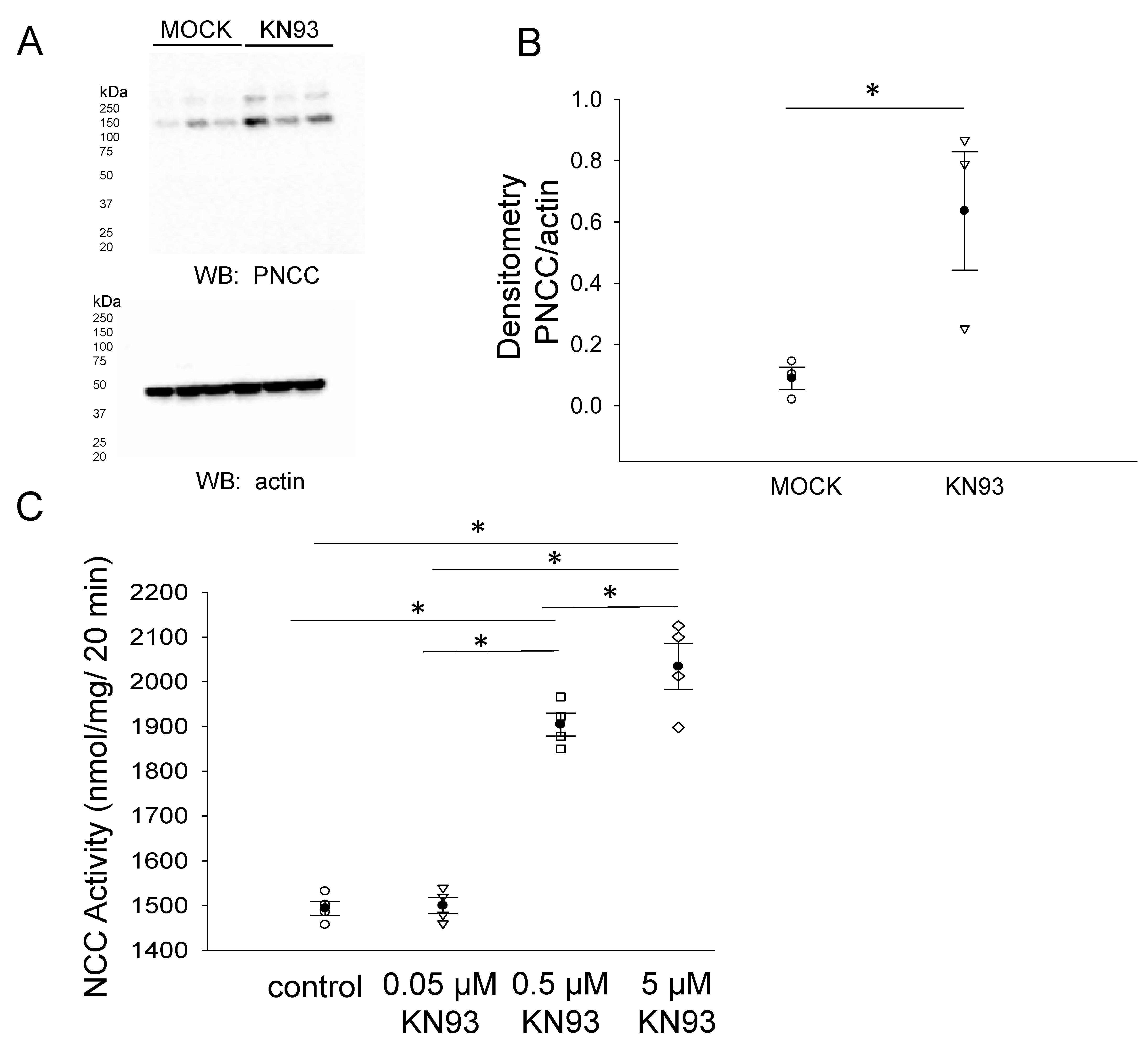
3.4. CaMKII Inhibition Augments the Total NCC Protein Expression in mDCT15 Cells
3.5. CaMKII Inhibition Increases Phospho-NCC and Its Activity in mDCT15 Cells
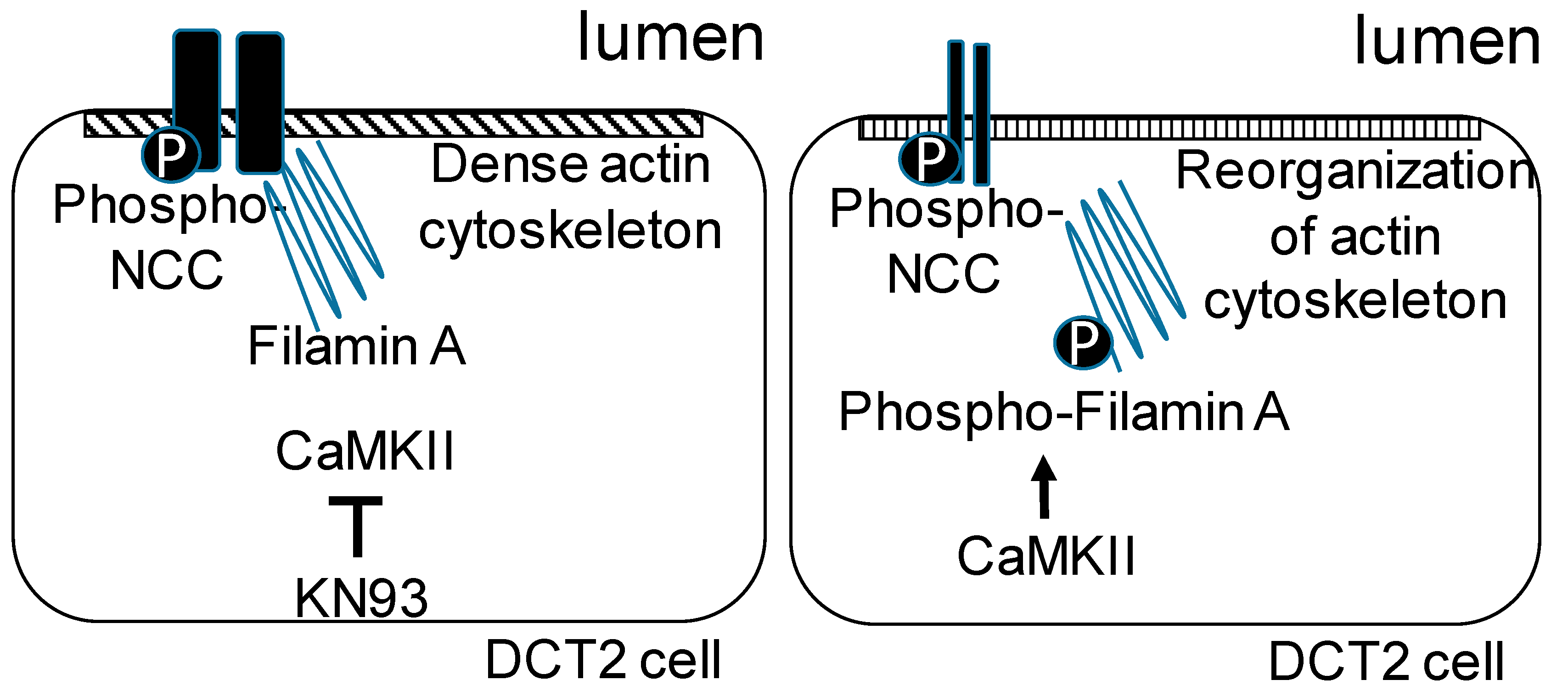
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jones, C.A.; Krolewski, A.S.; Rogus, J.; Xue, J.L.; Collins, A.; Warram, J.H. Epidemic of end-stage renal disease in people with diabetes in the United States population: Do we know the cause? Kidney Int. 2005, 67, 1684–1691. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention. Vital signs: Awareness and treatment of uncontrolled hypertension among adults—United States, 2003–2010. MMWR Morb. Mortal Wkly. Rep. 2012, 61, 703–709. [Google Scholar]
- Kearney, P.M.; Whelton, M.; Reynolds, K.; Muntner, P.; Whelton, P.K.; He, J. Global burden of hypertension: Analysis of worldwide data. Lancet 2005, 365, 217–223. [Google Scholar] [CrossRef]
- Guyton, A.C. Blood Pressure Control—Special Role of the Kidneys and Body Fluids. Science 1991, 252, 1813–1816. [Google Scholar] [CrossRef]
- Seyberth, H.W.; Weber, S.; Komhoff, M. Bartter’s and Gitelman’s syndrome. Curr. Opin. Pediatr. 2017, 29, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Gamba, G. Regulation of the renal Na+-Cl− cotransporter by phosphorylation and ubiquitylation. Am. J. Physiol. Physiol. 2012, 303, F1573–F1583. [Google Scholar] [CrossRef] [Green Version]
- Khurana, S. Role of Actin Cytoskeleton in Regulation of Ion Transport: Examples from Epithelial Cells. J. Membr. Biol. 2000, 178, 73–87. [Google Scholar] [CrossRef] [PubMed]
- Alli, A.A.; Bao, H.-F.; Alli, A.A.; Aldrugh, Y.; Song, J.Z.; Ma, H.-P.; Yu, L.; Al-Khalili, O.; Eaton, D.C. Phosphatidylinositol phosphate-dependent regulation of Xenopus ENaC by MARCKS protein. Am. J. Physiol.-Renal Physiol. 2012, 303, F800–F811. [Google Scholar] [CrossRef] [Green Version]
- Alli, A.A.; Bao, H.-F.; Liu, B.-C.; Yu, L.; Aldrugh, S.; Montgomery, D.S.; Ma, H.-P.; Eaton, D.C. Calmodulin and CaMKII modulate ENaC activity by regulating the association of MARCKS and the cytoskeleton with the apical membrane. Am. J. Physiol. Physiol. 2015, 309, F456–F463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montgomery, D.S.; Yu, L.; Ghazi, Z.M.; Thai, T.L.; Al-Khalili, O.; Ma, H.-P.; Eaton, D.C.; Alli, A.A. ENaC activity is regulated by calpain-2 proteolysis of MARCKS proteins. Am. J. Physiol. Physiol. 2017, 313, C42–C53. [Google Scholar] [CrossRef]
- Reifenberger, M.S.; Yu, L.; Bao, H.-F.; Duke, B.J.; Liu, B.-C.; Ma, H.-P.; Alli, A.A.; Eaton, D.; Alli, A.A. Cytochalasin E alters the cytoskeleton and decreases ENaC activity in Xenopus 2F3 cells. Am. J. Physiol. Physiol. 2014, 307, F86–F95. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, S.; Yui, N.; Noda, Y. Actin directly interacts with different membrane channel proteins and influences channel activities: AQP2 as a model. Biochim. Biophys. Acta (BBA)-Biomembr. 2014, 1838, 514–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbattiscianni, A.C.; Favia, M.; Mancini, M.T.; Cardone, R.A.; Guerra, L.; Monterisi, S.; Castellani, S.; Laselva, O.; Di Sole, F.; Conese, M.; et al. Correctors of mutant CFTR enhance subcortical cAMP-PKA signaling through modulating ezrin phosphorylation and cytoskeleton organization. J. Cell Sci. 2016, 129, 1128–1140. [Google Scholar] [PubMed] [Green Version]
- Edelman, A. Cytoskeleton and CFTR. Int. J. Biochem. Cell Biol. 2014, 52, 68–72. [Google Scholar] [CrossRef]
- Dimke, H.; San-Cristobal, P.; de Graaf, M.; Lenders, J.W.; Deinum, J.; Hoenderop, J.G.; Bindels, R.J. gamma-Adducin stimulates the thiazide-sensitive NaCl cotransporter. J. Am. Soc. Nephrol. 2011, 22, 508–517. [Google Scholar] [CrossRef] [Green Version]
- Stossel, T.P.; Condeelis, J.; Cooley, L.; Hartwig, J.H.; Noegel, A.; Schleicher, M.; Shapiro, S.S. Filamins as integrators of cell mechanics and signalling. Nat. Rev. Mol. Cell Biol. 2001, 2, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Sampson, L.J.; Leyland, M.L.; Dart, C. Direct Interaction between the Actin-binding Protein Filamin-A and the Inwardly Rectifying Potassium Channel, Kir2.1. J. Biol. Chem. 2003, 278, 41988–41997. [Google Scholar] [CrossRef] [Green Version]
- van der Flier, A.; Sonnenberg, A. Structural and functional aspects of filamins. Biochim. Biophys. Acta 2001, 1538, 99–117. [Google Scholar] [CrossRef] [Green Version]
- Ko, B.; Mistry, A.C.; Hanson, L.; Mallick, R.; Cooke, L.L.; Hack, B.K.; Cunningham, P.; Hoover, R.S. A new model of the distal convoluted tubule. Am. J. Physiol. Physiol. 2012, 303, F700–F710. [Google Scholar] [CrossRef] [Green Version]
- Hatano, R.; Fujii, E.; Segawa, H.; Mukaisho, K.; Matsubara, M.; Miyamoto, K.-I.; Hattori, T.; Sugihara, H.; Asano, S. Ezrin, a membrane cytoskeletal cross-linker, is essential for the regulation of phosphate and calcium homeostasis. Kidney Int. 2013, 83, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, K.; Yoshida, S.; Hatano, R.; Asano, S. Pathophysiological Roles of Ezrin/Radixin/Moesin Proteins. Biol. Pharm. Bull. 2017, 40, 381–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Jin, W.W.; Tsuji, K.; Chen, Y.; Nomura, N.; Su, L.; Yui, N.; Arthur, J.; Cotecchia, S.; Paunescu, T.G.; et al. Ezrin directly interacts with AQP2 and promotes its endocytosis. J. Cell Sci. 2017, 130, 2914–2925. [Google Scholar] [PubMed] [Green Version]
- Reiss, T. Drug discovery of the future: The implications of the human genome project. Trends Biotechnol. 2001, 19, 496–499. [Google Scholar] [CrossRef]
- Popowicz, G.M.; Schleicher, M.; Noegel, A.A.; Holak, T.A. Filamins: Promiscuous organizers of the cytoskeleton. Trends Biochem. Sci. 2006, 31, 411–419. [Google Scholar] [CrossRef]
- Zhou, A.-X.; Hartwig, J.H.; Akyürek, L.M. Filamins in cell signaling, transcription and organ development. Trends Cell Biol. 2010, 20, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.Y.; Ridgway, L.D.; Dryer, S.E. Interactions with Filamin A Stimulate Surface Expression of Large-Conductance Ca2+-Activated K+ Channels in the Absence of Direct Actin Binding. Mol. Pharmacol. 2007, 72, 622–630. [Google Scholar] [CrossRef] [Green Version]
- Thelin, W.R.; Chen, Y.; Gentzsch, M.; Kreda, S.M.; Sallee, J.L.; Scarlett, C.O.; Borchers, C.H.; Jacobson, K.; Stutts, M.J.; Milgram, S.L. Direct interaction with filamins modulates the stability and plasma membrane expression of CFTR. J. Clin. Investig. 2007, 117, 364–374. [Google Scholar] [CrossRef]
- Spector, I.; Shochet, N.R.; Blasberger, D.; Kashman, Y. Latrunculins--novel marine macrolides that disrupt microfilament organization and affect cell growth: I. Comparison with cytochalasin D. Cell Motil. Cytoskeleton 1989, 13, 127–144. [Google Scholar] [CrossRef]
- Spector, I.; Shochet, N.R.; Kashman, Y.; Groweiss, A. Latrunculins: Novel Marine Toxins That Disrupt Microfilament Organization in Cultured Cells. Science 1983, 219, 493–495. [Google Scholar] [CrossRef]
- Cooper, J. Effects of cytochalasin and phalloidin on actin. J. Cell Biol. 1987, 105, 1473–1478. [Google Scholar] [CrossRef] [Green Version]
- Foissner, I.; Wasteneys, G. Wide-Ranging Effects of Eight Cytochalasins and Latrunculin A and B on Intracellular Motility and Actin Filament Reorganization in Characean Internodal Cells. Plant Cell Physiol. 2007, 48, 585–597. [Google Scholar] [CrossRef]
- Pacheco-Alvarez, D.; Cristóbal, P.S.; Meade, P.; Moreno, E.; Vazquez, N.; Muñoz, E.; Díaz, A.; Juárez, M.E.; Gimenez, I.; Gamba, G. The Na+:Cl– Cotransporter Is Activated and Phosphorylated at the Amino-terminal Domain upon Intracellular Chloride Depletion. J. Biol. Chem. 2006, 281, 28755–28763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moriguchi, T.; Urushiyama, S.; Hisamoto, N.; Iemura, S.-I.; Uchida, S.; Natsume, T.; Matsumoto, K.; Shibuya, H. WNK1 Regulates Phosphorylation of Cation-Chloride-coupled Cotransporters via the STE20-related Kinases, SPAK and OSR1. J. Biol. Chem. 2005, 280, 42685–42693. [Google Scholar] [CrossRef] [Green Version]
- Vadlamudi, R.K.; Li, F.; Adam, L.; Nguyen, D.; Ohta, Y.; Stossel, T.P.; Kumar, R. Filamin is essential in actin cytoskeletal assembly mediated by p21-activated kinase 1. Nature 2002, 4, 681–690. [Google Scholar] [CrossRef]
- Woo, M.S.; Ohta, Y.; Rabinovitz, I.; Stossel, T.P.; Blenis, J. Ribosomal S6 Kinase (RSK) Regulates Phosphorylation of Filamin A on an Important Regulatory Site. Mol. Cell. Biol. 2004, 24, 3025–3035. [Google Scholar] [CrossRef] [Green Version]
- Cukier, I.H.; Li, Y.; Lee, J.M. Cyclin B1/Cdk1 binds and phosphorylates Filamin A and regulates its ability to cross-link actin. FEBS Lett. 2007, 581, 1661–1672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Lu, Y.; Stemmer, P.; Chen, F. Filamin A phosphorylation by Akt promotes cell migration in response to arsenic. Oncotarget 2015, 6, 12009–12019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gholam, M.F.; Ko, B.; Ghazi, Z.M.; Hoover, R.S.; Alli, A.A. The Pharmacological Inhibition of CaMKII Regulates Sodium Chloride Cotransporter Activity in mDCT15 Cells. Biology 2021, 10, 1335. https://doi.org/10.3390/biology10121335
Gholam MF, Ko B, Ghazi ZM, Hoover RS, Alli AA. The Pharmacological Inhibition of CaMKII Regulates Sodium Chloride Cotransporter Activity in mDCT15 Cells. Biology. 2021; 10(12):1335. https://doi.org/10.3390/biology10121335
Chicago/Turabian StyleGholam, Mohammed F., Benjamin Ko, Zinah M. Ghazi, Robert S. Hoover, and Abdel A. Alli. 2021. "The Pharmacological Inhibition of CaMKII Regulates Sodium Chloride Cotransporter Activity in mDCT15 Cells" Biology 10, no. 12: 1335. https://doi.org/10.3390/biology10121335
APA StyleGholam, M. F., Ko, B., Ghazi, Z. M., Hoover, R. S., & Alli, A. A. (2021). The Pharmacological Inhibition of CaMKII Regulates Sodium Chloride Cotransporter Activity in mDCT15 Cells. Biology, 10(12), 1335. https://doi.org/10.3390/biology10121335