
Rapid and Direct Action of Lipopolysaccharide (LPS) on Skeletal Muscle of Larval Drosophila



Abstract
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
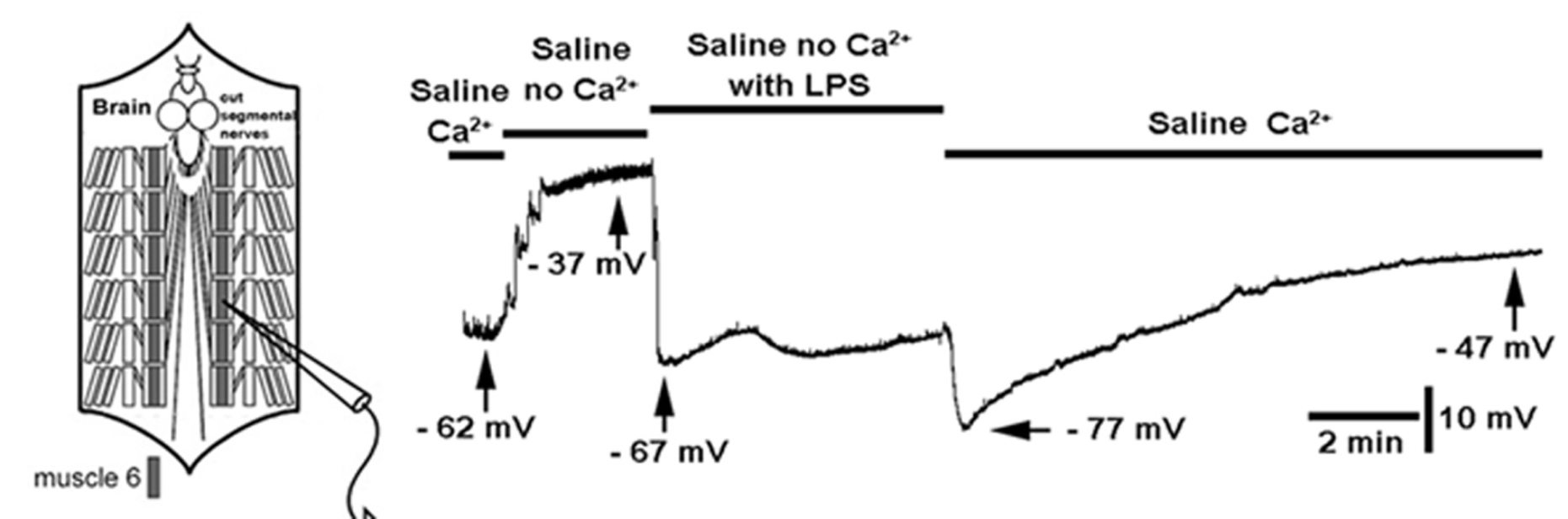
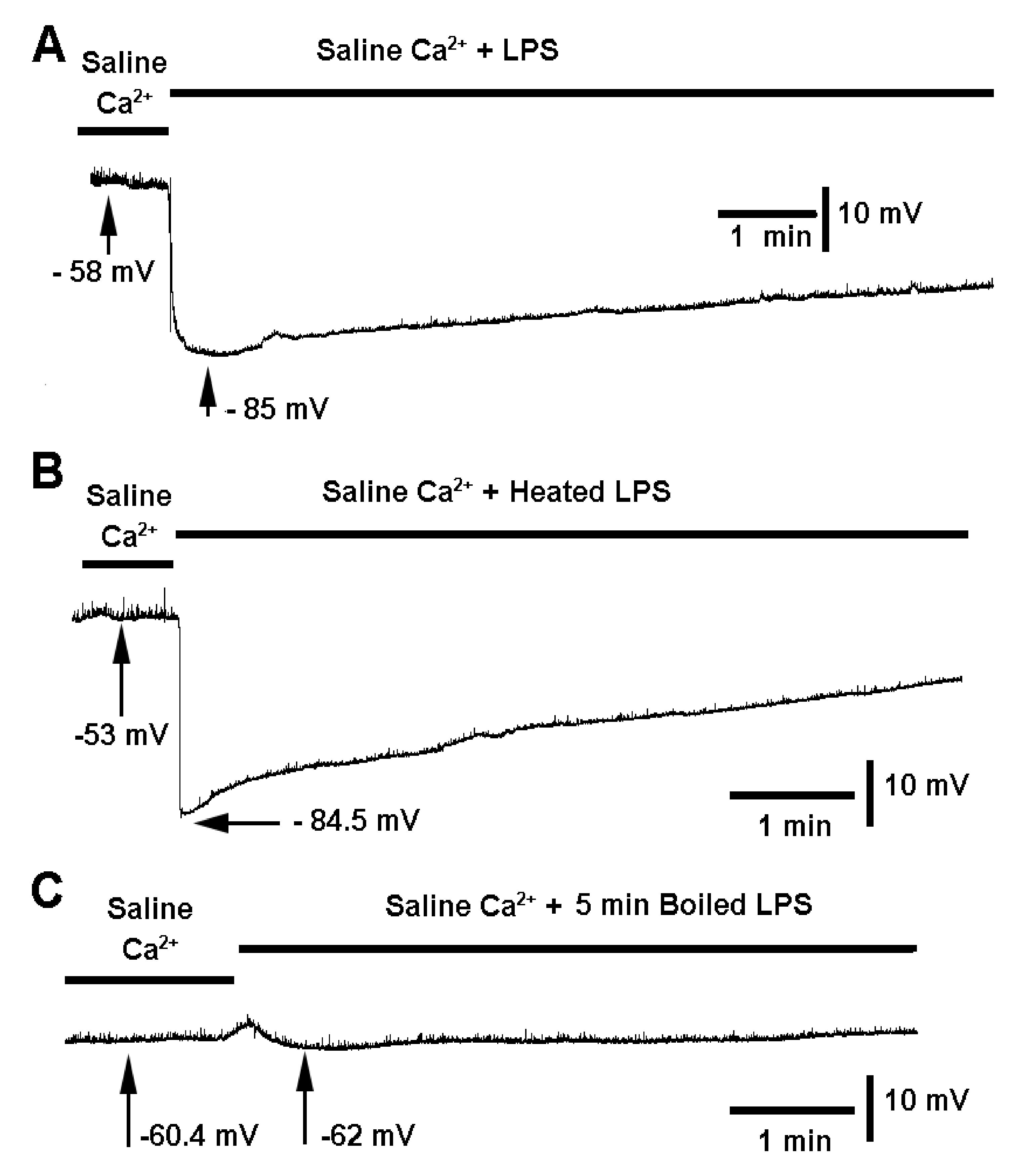
3. Results
4. Discussion
5. Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lorenzon, S.; de Guarrini, S.; Smith, V.; Ferrero, E. Effects of LPS injection on circulating haemocytes in crustaceans in vivo. Fish Shellfish Immunol. 1999, 9, 31–50. [Google Scholar] [CrossRef]
- Pien, B.C.; Sundaram, P.; Raoof, N.; Costa, S.F.; Mirrett, S.; Woods, C.W.; Reller, L.B.; Weinstein, M.P. The clinical and prognostic importance of positive blood cultures in adults. Am. J. Med. 2010, 123, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Carl, M.A.; Ndao, I.M.; Springman, A.C.; Manning, S.D.; Johnson, J.R.; Johnston, B.D.; Burnham, C.A.; Weinstock, E.S.; Weinstock, G.M.; Wylie, T.N.; et al. Sepsis from the gut: The enteric habitat of bacteria that cause late-onset neonatal bloodstream infections. Clin. Infect. Dis. 2014, 58, 1211–1218. [Google Scholar] [CrossRef] [PubMed]
- CDC Statistics. 2017. Available online: https://www.cdc.gov/nchs/fastats/leading-causes-of-death.htm (accessed on 22 July 2018).
- Kim, S.B.; Jeon, Y.D.; Kim, J.H.; Kim, J.K.; Ann, H.W.; Choi, H.; Kim, M.H.; Song, J.E.; Ahn, J.Y.; Jeong, S.J.; et al. Risk factors for mortality in patients with Serratia marcescens bacteremia. Yonsei Med. J. 2015, 56, 348–354. [Google Scholar] [CrossRef]
- Palavutitotai, N.; Jitmuang, A.; Tongsai, S.; Kiratisin, P.; Angkasekwinai, N. Epidemiology and risk factors of extensively drug-resistant Pseudomonas aeruginosa infections. PLoS ONE 2018, 13, e0193431. [Google Scholar] [CrossRef]
- Osborn, M.J.; Rosen, S.M.; Rothfield, L.; Zeleznick, L.D.; Horecker, B.L. Lipopolysaccharide of the gram-negative cell wall. Science 1964, 145, 783–789. [Google Scholar] [CrossRef]
- Linhartová, I.; Bumba, L.; Mašín, J.; Basler, M.; Osička, R.; Kamanová, J.; Procházková, K.; Adkins, I.; Hejnová-Holubová, J.; Sadílková, L.; et al. RTX proteins: A highly diverse family secreted by a common mechanism. FEMS Microbiol. Rev. 2010, 34, 1076–1112. [Google Scholar] [CrossRef]
- Eidelman, L.A.; Putterman, D.; Putterman, C.; Sprung, C.L. The spectrum of septic encephalopathy definitions, etiologies, and mortalities. JAMA 1996, 275, 470–473. [Google Scholar] [CrossRef]
- Wilson, J.X.; Young, G.B. Progress in clinical neurosciences: Sepsis-associated encephalopathy: Evolving concepts. Can. J. Neurol. Sci. 2003, 30, 98–105. [Google Scholar] [CrossRef]
- Friedrich, O.; Reid, M.B.; Van den Berghe, G.; Vanhorebeek, I.; Hermans, G.; Rich, M.M.; Larsson, L. The sick and the weak: Neuropathies/myopathies in the critically ill. Physiol. Rev. 2015, 95, 1025–1109. [Google Scholar] [CrossRef]
- Al-Nassan, S.; Fujino, H. Exercise preconditioning attenuates atrophic mediators and preserves muscle mass in acute sepsis. Gen. Physiol. Biophys. 2018, 37, 433–441. [Google Scholar] [CrossRef]
- Stefanska, B.; Człapa, W.; Pruszynska-Oszmałek, E.; Szczepankiewicz, D.; Fievez, V.; Komisarek, J.; Stajek, K.; Nowak, W. Subacute ruminal acidosis affects fermentation and endotoxin concentration in the rumen and relative expression of the CD14/TLR4/MD2 genes involved in lipopolysaccharide systemic immune response in dairy cows. J. Dairy Sci. 2018, 101, 1297–1310. [Google Scholar] [CrossRef]
- Levin, T.C.; Malik, H.S. Rapidly evolving Toll-3/4 genes encode male-specific Toll-like receptors in Drosophila. Mol. Biol. Evol. 2017, 34, 2307–2323. [Google Scholar] [CrossRef]
- Nehme, N.T.; Liégeois, S.; Kele, B.; Giammarinaro, P.; Pradel, E.; Hoffmann, J.A.; Ewbank, J.J.; Ferrandon, D. A model of bacterial intestinal infections in Drosophila melanogaster. PLoS Pathog. 2007, 3, e173. [Google Scholar] [CrossRef]
- Kreger, A.S.; Lyerly, D.M.; Hazlett, L.D.; Berk, R.S. Immunization against experimental Pseudomonas aeruginosa and Serratia marcescens keratitis. Vaccination with lipopolysaccharide endotoxins and proteases. Investig. Ophthalmol. Vis. Sci. 1986, 27, 932–939. [Google Scholar]
- Lorenzon, S.; Pasqual, P.; Ferrero, E.A. Different bacterial lipopolysaccharides as toxicants and stressors in the shrimp Palaemon elegans. Fish Shellfish Immunol. 2002, 13, 27–45. [Google Scholar] [CrossRef]
- Luchi, M.; Morrison, D.C. Comparable endotoxic properties of lipopolysaccharides are manifest in diverse clinical isolates of gram-negative bacteria. Infect. Immun. 2000, 68, 1899–1904. [Google Scholar] [CrossRef]
- Celes, M.R.; Prado, C.M.; Rossi, M.A. Sepsis: Going to the heart of the matter. Pathobiology 2013, 80, 70–86. [Google Scholar] [CrossRef]
- Rameshrad, M.; Maleki-Dizaji, N.; Vaez, H.; Soraya, H.; Nakhlband, A.; Garjani, A. Lipopolysaccharide induced activation of toll like receptor 4 in isolated rat heart suggests a local immune response in myocardium. Iran. J. Immunol. 2015, 12, 104–116. [Google Scholar]
- Volchenkov, R.; Sprater, F.; Vogelsang, P.; Appel, S. The 2011 Nobel Prize in physiology or medicine. Scand. J. Immunol. 2012, 75, 1–4. [Google Scholar] [CrossRef]
- Saelinger, C.M.; McNabb, M.C.; McNair, R.; Bierbower, S.; Cooper, R.L. Effects of bacterial endotoxin on regulation of the heart, a sensory-CNS-motor nerve circuit and neuromuscular junctions: Crustacean model. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2019, 237, 110557. [Google Scholar] [CrossRef]
- Istas, O.; Greenhalgh, A.; Cooper, R.L. The effects of a bacterial endotoxin on behavior and sensory-CNS-motor circuits in Drosophila melanogaster. Insects 2019, 10, 115. [Google Scholar] [CrossRef]
- Cooper, R.L.; McNabb, M.; Nadolski, J. The effects of a bacterial endotoxin LPS on synaptic transmission at the neuromuscular junction. Heliyon 2019, 5, e01430. [Google Scholar] [CrossRef]
- Ballinger-Boone, C.; Anyagaligbo, O.; Bernard, J.; Bierbower, S.M.; Dupont-Versteegden, E.E.; Ghoweri, A.; Greenhalgh, A.; Harrison, D.; Istas, O.; McNabb, M.; et al. The effects of bacterial endotoxin (LPS) on cardiac and synaptic function in various animal models: Larval Drosophila, crayfish, crab, and rodent. Int. J. Zool. Res. 2020, 16, 33–62. [Google Scholar] [CrossRef]
- Bhatt, D.; Cooper, R.L. The pharmacological and physiological profile of glutamate receptors at the Drosophila larval neuromuscular junction. Physiol. Entomol. 2005, 30, 205–210. [Google Scholar] [CrossRef]
- Lee, J.Y.; Bhatt, D.; Chung, W.Y.; Cooper, R.L. Furthering pharmacological and physiological assessment of the glutamatergic receptors at the Drosophila neuromuscular junction. Comp. Biochem. Physiol. C 2009, 150, 546–555. [Google Scholar] [CrossRef]
- Titlow, J.S.; Cooper, R.L. Glutamatergic Synthesis, Recycling, and Receptor Pharmacology at Drosophila and Crustacean Neuromuscular Junctions. In Biochemical Approaches for Glutamatergic Neurotransmission; Parrot, S., Denoroy, L., Eds.; Neuromethods; Humana Press: New York, NY, USA, 2018; Volume 130, pp. 263–291. [Google Scholar]
- Rose, U.; Derst, C.; Wanischeck, M.; Marinc, C.; Walther, C. Properties and possible function of a hyperpolarisation-activated chloride current in Drosophila. J. Exp. Biol. 2007, 210, 2489–2500. [Google Scholar] [CrossRef]
- Stanley, C.E.; Mauss, A.; Borst, A.; Cooper, R.L. The effects of chloride flux on Drosophila heart rate. Methods Protoc. 2019, 2, 73. [Google Scholar] [CrossRef]
- Gho, M.; Mallart, A. Two distinct calcium-activated potassium currents in larval muscle fibres of Drosophila melanogaster. Pflug. Arch. 1986, 407, 526–533. [Google Scholar] [CrossRef]
- Stewart, B.A.; Atwood, H.L.; Renger, J.J.; Wang, J.; Wu, C.F. Improved stability of Drosophila larval neuromuscular preparation in haemolymph-like physiological solutions. J. Comp. Physiol. A 1994, 175, 179–191. [Google Scholar] [CrossRef]
- De Castro, C.; Titlow, J.; Majeed, Z.R.; Cooper, R.L. Analysis of various physiological salines for heart rate, CNS function, and synaptic transmission at neuromuscular junctions in Drosophila melanogaster larvae. J. Comp. Physiol. A 2014, 200, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Henon, B.K.; Ikeda, K. Changes in membrane properties of the Drosophila dorsal longitudinal flight muscle induced by sodium pump inhibitors. J. Exp. Biol. 1981, 90, 175–183. [Google Scholar] [CrossRef]
- Istas, O.; Greenhalgh, A.; Cooper, R.L. Repetitive exposure to bacterial endotoxin LPS alters synaptic transmission. J. Pharmacol. Toxicol. 2020, 15, 65–72. [Google Scholar] [CrossRef]
- Iwaya, A.; Nakagawa, S.; Iwakura, N.; Taneike, I.; Kurihara, M.; Kuwano, T.; Gondaira, F.; Endo, M.; Hatakeyama, K.; Yamamoto, T. Rapid and quantitative detection of blood Serratia marcescens by a real-time PCR assay: Its clinical application and evaluation in a mouse infection model. FEMS Microbiol. Lett. 2005, 248, 163–170. [Google Scholar] [CrossRef][Green Version]
- Krans, J.L.; Parfitt, K.D.; Gawera, K.D.; Rivlin, P.K.; Hoy, R.R. The resting membrane potential of Drosophila melanogaster larval muscle depends strongly on external calcium concentration. J. Insect Physiol. 2010, 56, 304–313. [Google Scholar] [CrossRef]
- Armstrong, C.M. Distinguishing surface effects of calcium ion from pore-occupancy effects in Na+ channels. Proc. Natl. Acad. Sci. USA 1999, 96, 4158–4163. [Google Scholar] [CrossRef]
- Armstrong, C.M.; Cota, G. Calcium block of Na+ channels and its effect on closing rate. Proc. Natl. Acad. Sci. USA 1999, 96, 4154–4157. [Google Scholar] [CrossRef]
- Gorman, A.L.F.; Hermann, A. Internal effects of divalent cations on potassium permeability in molluscan neurons. J. Physiol. 1979, 296, 393–410. [Google Scholar] [CrossRef]
- Hermann, A.; Erxleben, C.; Armstrong, D. Small Conductance Ca Activated K Channels in Mollusks. In Calcium and Ion Channel Modulation; Grinnell, A.D., Armstrong, D., Jackson, M.B., Eds.; Springer: Boston, MA, USA, 1988. [Google Scholar] [CrossRef]
- Leinders, T.; van Kleef, R.G.D.M.; Vijverberg, H.P.M. Divalent cations activate small- (SK) and large-conductance (BK) channels in mouse neuroblastoma cells: Selective activation of SK channels by cadmium. Pflügers Arch. 1992, 422, 217–222. [Google Scholar] [CrossRef]
- Hermann, A.; Gorman, A.L.F. External and internal effects of tetraethylammonium on voltage-dependent and Ca-dependent K+ currents components in molluscan pacemaker neurons. Neurosci. Lett. 1979, 12, 87–92. [Google Scholar] [CrossRef]
- Hayabuchi, Y.; Nakaya, Y.; Mawatari, K.; Inoue, M.; Sakata, M.; Kagami, S. Cell membrane stretch activates intermediate-conductance Ca2+-activated K+ channels in arterial smooth muscle cells. Heart Vessel. 2011, 26, 91–100. [Google Scholar] [CrossRef]
- Whatley, S.A.; Powell, J.F.; Politopoulou, G.; Campbell, I.C.; Brammer, M.J.; Percy, N.S. Regulation of intracellular free calcium levels by the cellular prion protein. Neuroreport 1995, 6, 2333–2337. [Google Scholar] [CrossRef]
- Mont, M.R.; Carlson, C.G.; Geisbuhler, T.P. Resting Ca2+ influx does not contribute to anoxia-induced cell death in adult rat cardiac myocytes. Can. J. Physiol. Pharmacol. 2009, 87, 360–370. [Google Scholar] [CrossRef]
- Dayaram, V.; Malloy, C.; Martha, S.; Alvarez, B.; Chukwudolue, I.; Dabbain, N.; Mahmood, D.; Goleva, S.; Hickey, T.; Ho, A.; et al. Stretch activated channels in proprioceptive chordotonal organs of crab and crayfish are sensitive to Gd3+ but not amiloride, ruthenium red or low pH. Impluse 2017, 2017, 1–17. Available online: https://impulse.appstate.edu/issues/2017 (accessed on 1 May 2021).
- McCubbin, S.; Jeoung, A.; Waterbury, C.; Cooper, R.L. Pharmacological Profiling of Stretch Activated Channels in Proprioceptive Neuron. Comp. Biochem. Physiol. C 2020, 233, 108765. Available online: https://www.sciencedirect.com/science/article/abs/pii/S153204562030065X (accessed on 1 May 2021). [CrossRef]
- Chao, S.H.; Bu, C.H.; Cheung, W.Y. Activation of troponin C by Cd2+ and Pb2+. Arch. Toxicol. 1990, 64, 490–496. [Google Scholar] [CrossRef]
- Drakenberg, T.; Forsén, S.; Thulin, E.; Vogel, H.J. The binding of Ca2+, Mg2+ and Cd2+ to tryptic fragments of skeletal muscle troponin C. Cadmium-113 and proton NMR studies. J. Biol. Chem. 1987, 262, 672–678. [Google Scholar] [CrossRef]
- Rheuben, M. The resting potential of moth muscle fibre. J. Physiol. 1972, 225, 529–554. [Google Scholar] [CrossRef]
- Ikeda, K.; Ozawa, S.; Hagiwara, S. Synaptic transmission reversibly conditioned by single-gene mutation in Drosophila melanogaster. Nat. Lond. 1976, 259, 489–491. [Google Scholar] [CrossRef]
- Salkoff, L.B.; Wyman, R.J. Ion currents in Drosophila flight muscles. J. Physiol. 1983, 337, 687–709. [Google Scholar] [CrossRef]
- Huddart, H.; Wood, D.W. The effect of DNP on the resting potential and ionic content of some insect skeletal muscle fibres. Comp. Biochem. Physiol. 1966, 18, 681–688. [Google Scholar] [CrossRef]
- Ashcroft, F.M.; Stanfield, P.R. Calcium and potassium currents in muscle fibres of an insect (Carausius morosus). J. Physiol. 1982, 323, 93–115. [Google Scholar] [CrossRef]
- Djamgoz, M.B.A. Insect muscle: Intracellular ion concentrations and mechanisms of resting potential generation. J. Insect Physiol. 1987, 33, 287–314. [Google Scholar] [CrossRef]
- Sun, B.; Wang, W.; Salvaterra, P.M. Functional analysis and tissue-specific expression of Drosophila Na+ K+-ATPase subunits. J. Neurochem. 1998, 71, 142–151. [Google Scholar] [CrossRef]
- Bernard, J.; Greenhalgh, A.; Istas, O.; Marguerite, N.T.; Cooper, R.L. The effect of bacterial endotoxin LPS on serotonergic modulation of glutamatergic synaptic transmission. Biology 2020, 9, 210. [Google Scholar] [CrossRef]
- Greenhalgh, A.; Istas, O.; Cooper, R.L. Bacterial endotoxin lipopolysaccharide enhances synaptic transmission at low-output glutamatergic synapses. Neurosci. Res. 2020, 170, 59–65. [Google Scholar] [CrossRef]
- He, P.; Southard, R.C.; Whiteheart, S.W.; Cooper, R.L. Role of alpha-SNAP in promoting efficient neurotransmission at the crayfish neuromus-cular junction. J. Neurophysiol. 1999, 82, 3406–3416. [Google Scholar] [CrossRef]
- Cooper, A.S.; Cooper, R.L. Historical view and physiological demonstration of synaptic transmission at the crayfish opener muscle. J. Vis. Exp. 2009, 33, 1595. Available online: http://www.jove.com/video/1595/historical-view-physiology-demonstration-at-nmj-crayfish-opener (accessed on 1 May 2021). [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Paradigm Type Conditions Used |
|---|
| Paradigm 1 Std. saline/Std. saline + LPS/Std. saline |
| Paradigm 2 Std. saline/Std. saline + boiled LPS/Std. saline |
| Paradigm 3 Std. saline/Std. saline + ouabain/Std. saline + ouabain + LPS/Std. saline |
| Paradigm 4 Std. saline/No Ca2+ saline/No Ca2+ saline + LPS/Std. saline |
| Paradigm 5 Std. saline/No Ca2+ saline + Ba2+/No Ca2+ saline + Ba2+ + LPS/Std. saline |
| Paradigm 6 Std. saline/Std. saline + Cd2+/Std. saline + Cd2+ + LPS/Std. saline |
| Paradigm 7 Std. saline/No Ca2+ saline + Cd2+/No Ca2+ saline + Cd2+ + LPS/Std. saline |
| Paradigm 8 Std. saline/No Ca2+ saline + Gd3+/No Ca2+ saline+ Gd3+ + LPS/Std. saline |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Potter, R.; Meade, A.; Potter, S.; Cooper, R.L. Rapid and Direct Action of Lipopolysaccharide (LPS) on Skeletal Muscle of Larval Drosophila. Biology 2021, 10, 1235. https://doi.org/10.3390/biology10121235
Potter R, Meade A, Potter S, Cooper RL. Rapid and Direct Action of Lipopolysaccharide (LPS) on Skeletal Muscle of Larval Drosophila. Biology. 2021; 10(12):1235. https://doi.org/10.3390/biology10121235
Chicago/Turabian StylePotter, Rachel, Alexis Meade, Samuel Potter, and Robin L. Cooper. 2021. "Rapid and Direct Action of Lipopolysaccharide (LPS) on Skeletal Muscle of Larval Drosophila" Biology 10, no. 12: 1235. https://doi.org/10.3390/biology10121235
APA StylePotter, R., Meade, A., Potter, S., & Cooper, R. L. (2021). Rapid and Direct Action of Lipopolysaccharide (LPS) on Skeletal Muscle of Larval Drosophila. Biology, 10(12), 1235. https://doi.org/10.3390/biology10121235