
Rational Engineering of a Brevinin-2 Peptide: Decoupling Potency from Toxicity Through C-Terminal Truncation and N-Terminal Chiral Substitution

 , ,
, ,  ,
,  , and
, and
Abstract
1. Introduction
2. Results
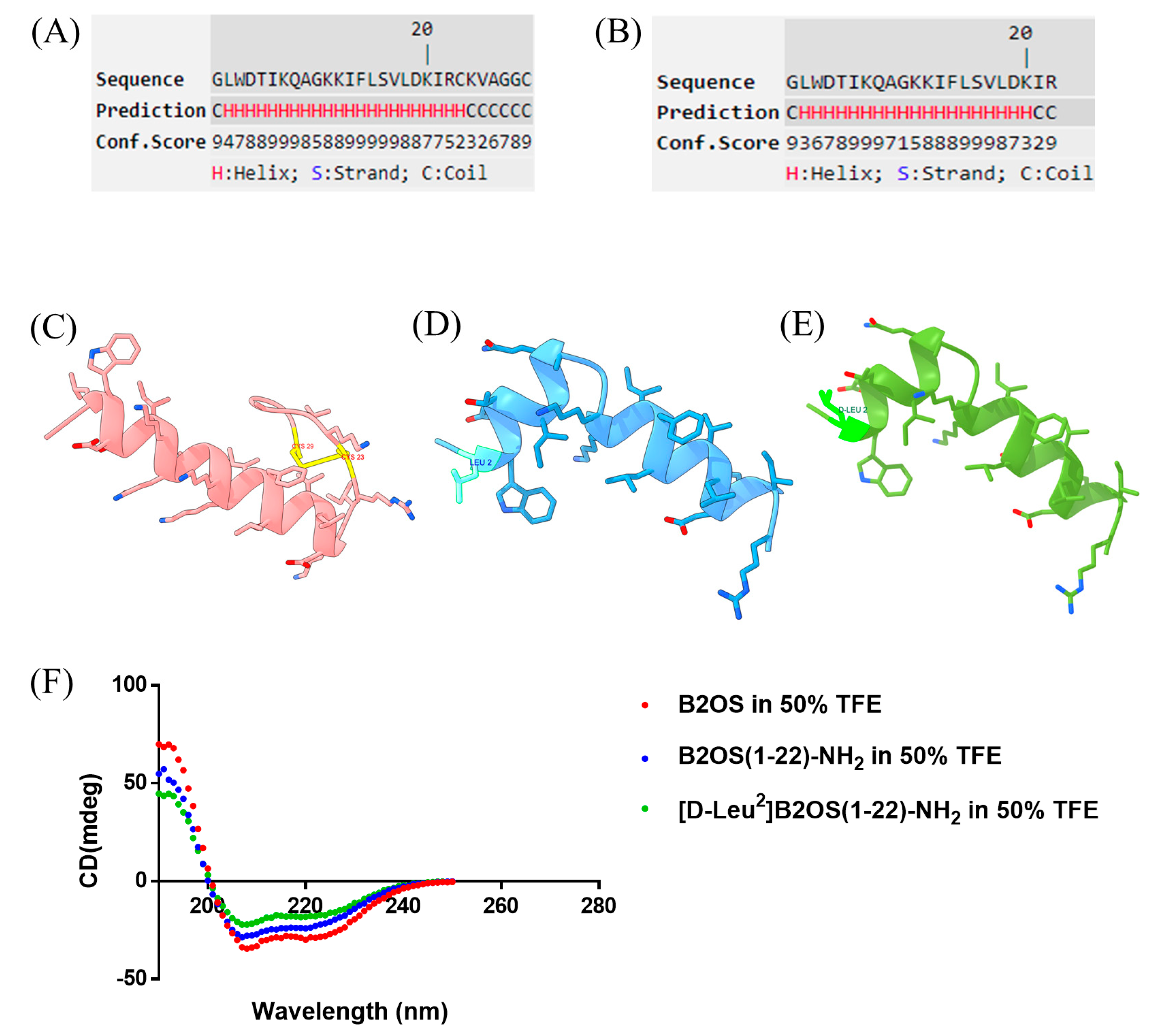
2.1. Peptide Conformational Analysis of B2OS and Its Analogues
2.2. Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC) of B2OS and Its Two Designed Analogues
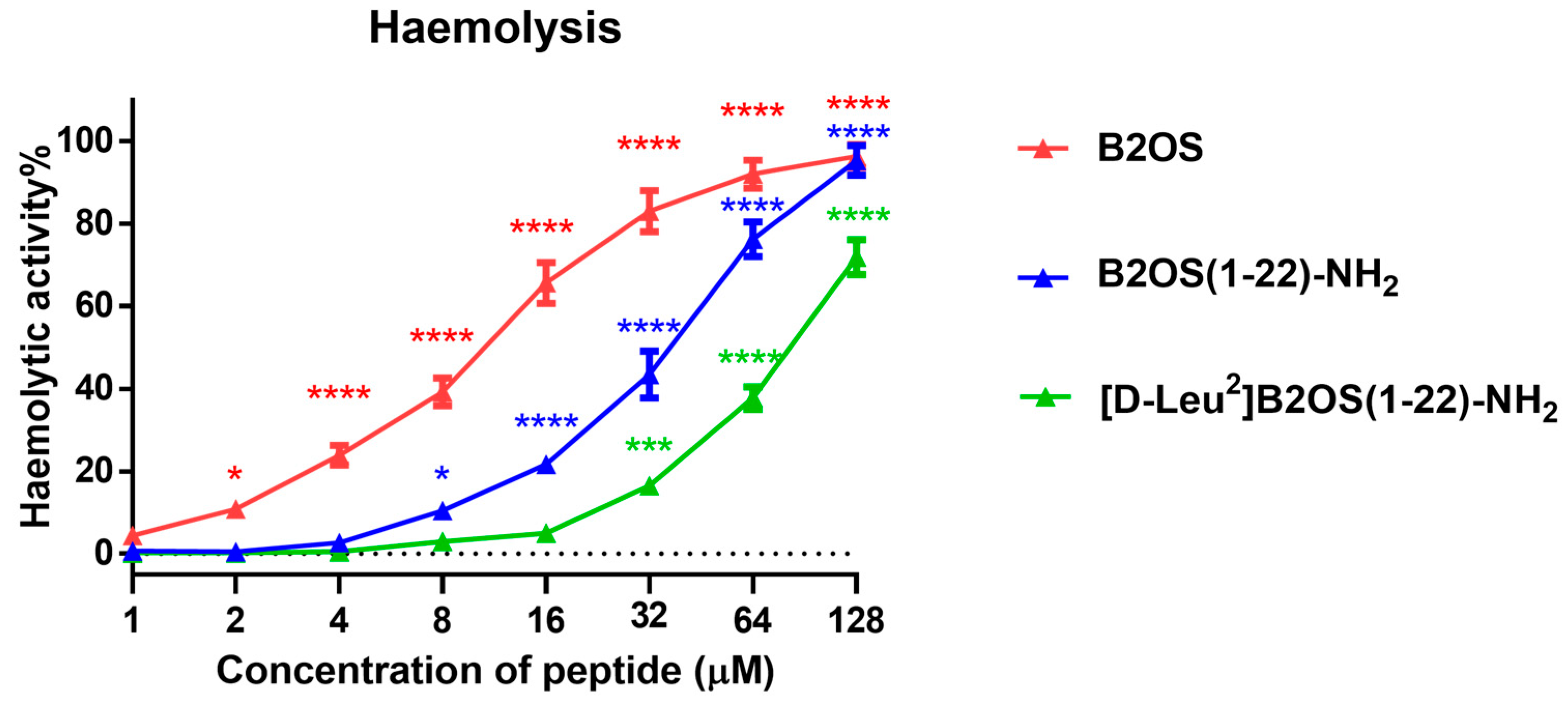
2.3. Haemolytic Activity of B2OS and Its Analogues
2.4. Prevention and Eradication of Biofilm by B2OS and Its Two Designed Analogues
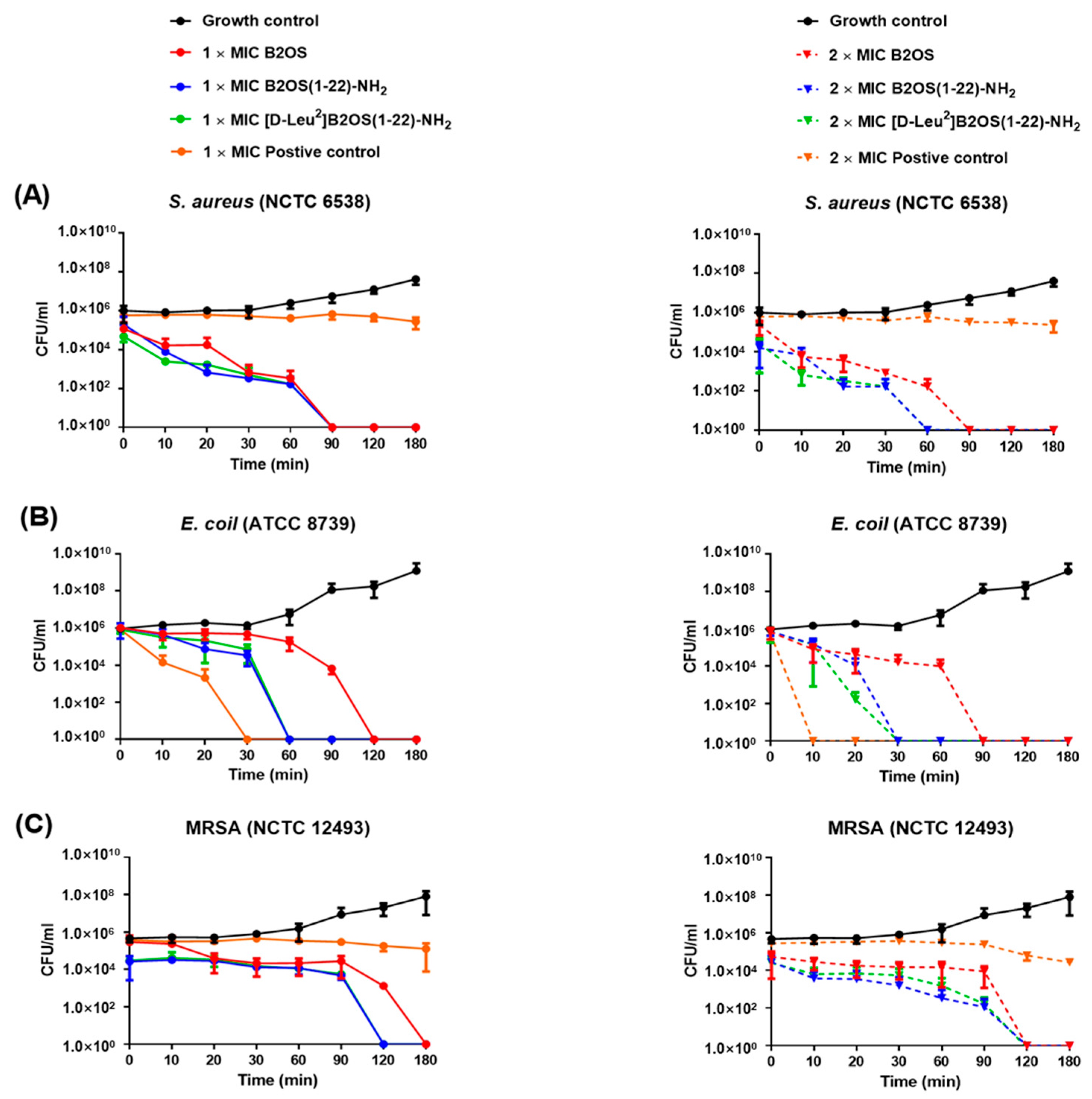
2.5. Kinetic Killing Effect of B2OS and Its Analogues
2.6. Membrane Permeabilisation on MRSA
2.7. In Vivo Waxworm Larvae Model
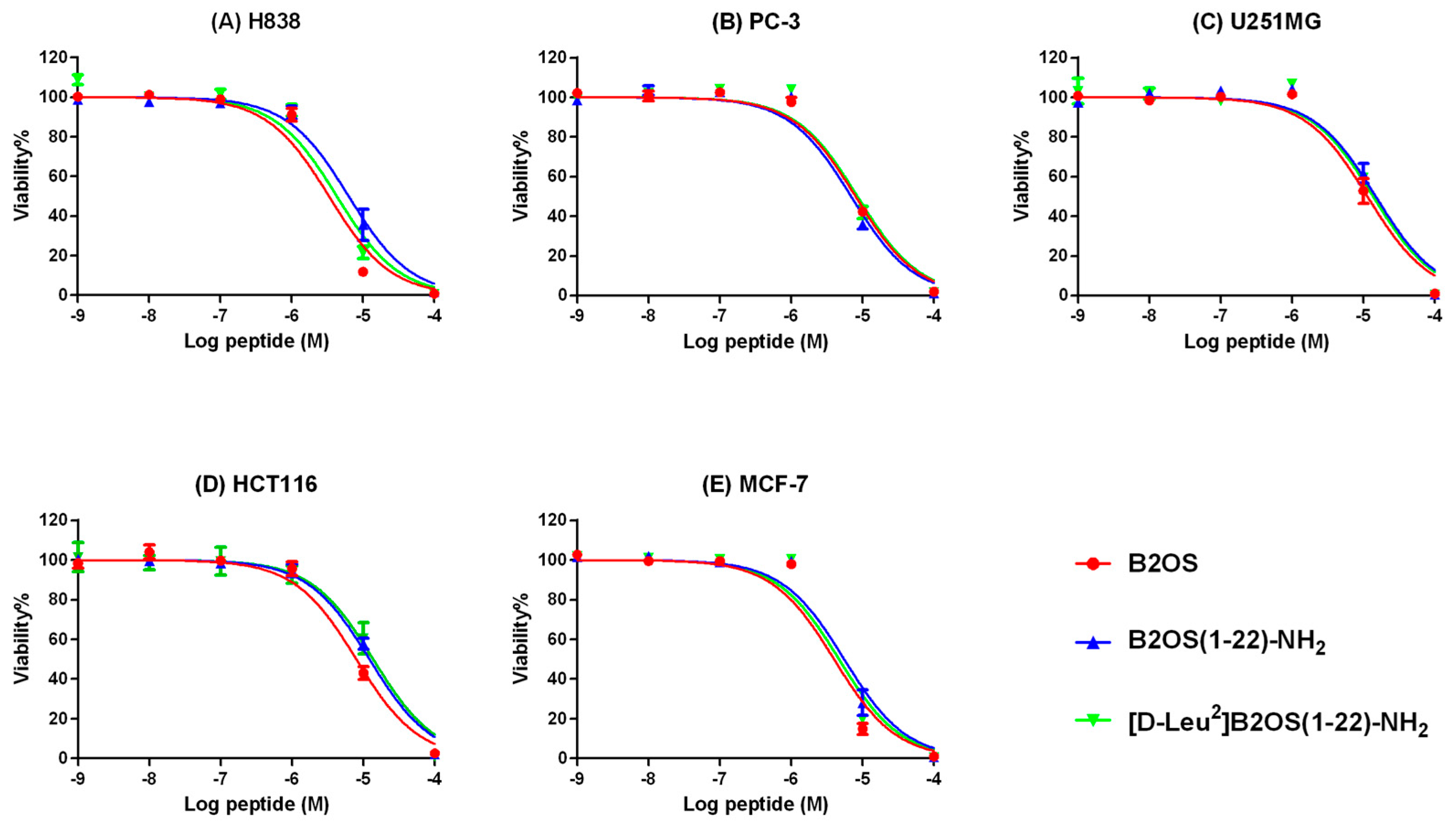
2.8. Anti-Cancer Proliferation Activity of B2OS and Its Analogues
3. Discussion
4. Materials and Methods
4.1. Peptide Synthesis and Purification
4.2. Bioinformatics Analysis
4.3. Circular Dichroism Spectra
4.4. Antibacterial Assay
4.5. Haemolysis Assays
4.6. Biofilm Inhibition Assay
4.7. Time-Killing Assays
4.8. Bacterial Membrane Permeability Assays
4.9. Efficacy Evaluation of Peptides Against MRSA in Larvae
4.10. Anti-Cancer Cell Proliferation Assays
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Antimicrobial Resistance Collaborators. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer statistics, 2023. CA Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef] [PubMed]
- Vasan, N.; Baselga, J.; Hyman, D.M. A view on drug resistance in cancer. Nature 2019, 575, 299–309. [Google Scholar] [CrossRef]
- Moghnieh, R.A.; Kanafani, Z.A.; Tabaja, H.Z.; Sharara, S.L.; Awad, L.S.; Kanj, S.S. Epidemiology of common resistant bacterial pathogens in the countries of the Arab League. Lancet Infect. Dis. 2018, 18, e379–e394. [Google Scholar] [CrossRef]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef]
- Hancock, R.E.; Sahl, H.G. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Mahlapuu, M.; Håkansson, J.; Ringstad, L.; Björn, C. Antimicrobial Peptides: An Emerging Category of Therapeutic Agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial Peptides: Classification, Design, Application and Research Progress in Multiple Fields. Front. Microbiol. 2020, 11, 582779. [Google Scholar] [CrossRef]
- Zohrab, F.; Askarian, S.; Jalili, A.; Kazemi Oskuee, R. Biological Properties, Current Applications and Potential Therapeautic Applications of Brevinin Peptide Superfamily. Int. J. Pept. Res. Ther. 2019, 25, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Hassanvand Jamadi, R.; Khalili, S.; Mirzapour, T.; Yaghoubi, H.; Hashemi, Z.S.; Mard-Soltani, M.; Jafarisani, M. Anticancer Activity of Brevinin-2R Peptide and its Two Analogues Against Myelogenous Leukemia Cell Line as Natural Treatments: An In Vitro Study. Int. J. Pept. Res. Ther. 2020, 26, 1013–1020. [Google Scholar] [CrossRef]
- Abdel-Wahab, Y.H.; Patterson, S.; Flatt, P.R.; Conlon, J.M. Brevinin-2-related peptide and its [D4K] analogue stimulate insulin release in vitro and improve glucose tolerance in mice fed a high fat diet. Horm. Metab. Res. 2010, 42, 652–656. [Google Scholar] [CrossRef] [PubMed]
- Quan, Z.; Zhou, M.; Chen, W.; Chen, T.; Walker, B.; Shaw, C. Novel brevinins from Chinese piebald odorous frog (Huia schmackeri) skin deduced from cloned biosynthetic precursors. Peptides 2008, 29, 1456–1460. [Google Scholar] [CrossRef]
- Conlon, J.M.; Kolodziejek, J.; Nowotny, N. Antimicrobial peptides from ranid frogs: Taxonomic and phylogenetic markers and a potential source of new therapeutic agents. Biochim. Biophys. Acta 2004, 1696, 1–14. [Google Scholar] [CrossRef]
- Yang, B.; Gelfanov, V.M.; Perez-Tilve, D.; DuBois, B.; Rohlfs, R.; Levy, J.; Douros, J.D.; Finan, B.; Mayer, J.P.; DiMarchi, R.D. Optimization of Truncated Glucagon Peptides to Achieve Selective, High Potency, Full Antagonists. J. Med. Chem. 2021, 64, 4697–4708. [Google Scholar] [CrossRef]
- Sumiyoshi, S.; Suyama, K.; Tanaka, N.; Andoh, T.; Nagata, A.; Tomohara, K.; Taniguchi, S.; Maeda, I.; Nose, T. Development of truncated elastin-like peptide analogues with improved temperature-response and self-assembling properties. Sci. Rep. 2022, 12, 19414. [Google Scholar] [CrossRef]
- Tan, Y.X.; Chen, C.; Wang, Y.L.; Lin, S.; Wang, Y.; Li, S.B.; Jin, X.P.; Gao, H.W.; Du, F.S.; Gong, F.; et al. Truncated peptides from melittin and its analog with high lytic activity at endosomal pH enhance branched polyethylenimine-mediated gene transfection. J. Gene Med. 2012, 14, 241–250. [Google Scholar] [CrossRef]
- Vieregg, J.R.; Lueckheide, M.; Marciel, A.B.; Leon, L.; Bologna, A.J.; Rivera, J.R.; Tirrell, M.V. Oligonucleotide–Peptide Complexes: Phase Control by Hybridization. J. Am. Chem. Soc. 2018, 140, 1632–1638. [Google Scholar] [CrossRef]
- Mohamed, M.F.; Brezden, A.; Mohammad, H.; Chmielewski, J.; Seleem, M.N. A short D-enantiomeric antimicrobial peptide with potent immunomodulatory and antibiofilm activity against multidrug-resistant Pseudomonas aeruginosa and Acinetobacter baumannii. Sci. Rep. 2017, 7, 6953. [Google Scholar] [CrossRef]
- Scudiero, O.; D’Amora, M.; Galdiero, M.; Galdiero, S. Non-Canonical Amino Acids as Building Blocks for Peptidomimetics: Structure, Function, and Applications. Molecules 2023, 28, 5275. [Google Scholar]
- Stephani, J.C.; Gerhards, L.; Khairalla, B.; Solov’yov, I.A.; Brand, I. How do Antimicrobial Peptides Interact with the Outer Membrane of Gram-Negative Bacteria? Role of Lipopolysaccharides in Peptide Binding, Anchoring, and Penetration. ACS Infect. Dis. 2024, 10, 763–778. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Liu, S.; Xi, X.; Ma, C.; Wang, L.; Chen, X.; Shi, Z.; Chen, T.; Shaw, C.; Zhou, M. Study on the Structure-Activity Relationship of an Antimicrobial Peptide, Brevinin-2GUb, from the Skin Secretion of Hylarana guentheri. Antibiotics 2021, 10, 895. [Google Scholar] [CrossRef]
- Xie, H.; Zhan, Y.; Chen, X.; Zeng, Q.; Chen, D.; Liang, J. Brevinin-2 Drug Family—New Applied Peptide Candidates Against Methicillin-Resistant Staphylococcus aureus and Their Effects on Lys-7 Expression of Innate Immune Pathway DAF-2/DAF-16 in Caenorhabditis elegans. Appl. Sci. 2018, 8, 2627. [Google Scholar] [CrossRef]
- Sierra, J.M.; Viñas, M. Future prospects for Antimicrobial peptide development: Peptidomimetics and antimicrobial combinations. Expert Opin. Drug Discov. 2021, 16, 601–604. [Google Scholar] [CrossRef]
- Conlon, J.M.; Kolodziejek, J.; Nowotny, N.; Leprince, J.; Vaudry, H.; Coquet, L.; Jouenne, T.; Iwamuro, S. Cytolytic peptides belonging to the brevinin-1 and brevinin-2 families isolated from the skin of the Japanese brown frog, Rana dybowskii. Toxicon 2007, 50, 746–756. [Google Scholar] [CrossRef]
- Liu, S.; Long, Q.; Xu, Y.; Wang, J.; Xu, Z.; Wang, L.; Zhou, M.; Wu, Y.; Chen, T.; Shaw, C. Assessment of antimicrobial and wound healing effects of Brevinin-2Ta against the bacterium Klebsiella pneumoniae in dermally-wounded rats. Oncotarget 2017, 8, 111369–111385. [Google Scholar] [CrossRef]
- Chen, G.; Miao, Y.; Ma, C.; Zhou, M.; Shi, Z.; Chen, X.; Burrows, J.F.; Xi, X.; Chen, T.; Wang, L. Brevinin-2GHk from Sylvirana guentheri and the Design of Truncated Analogs Exhibiting the Enhancement of Antimicrobial Activity. Antibiotics 2020, 9, 85. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Guarnieri, M.T.; Vasil, A.I.; Vasil, M.L.; Mant, C.T.; Hodges, R.S. Role of peptide hydrophobicity in the mechanism of action of α-helical antimicrobial peptides. Antimicrob. Agents Chemother. 2007, 51, 1398–1406. [Google Scholar] [CrossRef]
- Dathe, M.; Wieprecht, T.; Nikolenko, H.; Handel, L.; Maloy, W.L.; MacDonald, D.L.; Beyermann, M.; Bienert, M. Hydrophobicity, hydrophobic moment and angle subtended by charged residues modulate antibacterial and haemolytic activity of amphipathic helical peptides. FEBS Lett. 1997, 403, 208–212. [Google Scholar] [CrossRef]
- Eisenberg, D. Three-dimensional structure of membrane and surface proteins. Annu. Rev. Biochem. 1984, 53, 595–623. [Google Scholar] [CrossRef] [PubMed]
- Wieprecht, T.; Dathe, M.; Beyermann, M.; Krause, E.; Maloy, W.L.; MacDonald, D.L.; Bienert, M. Peptide hydrophobicity controls the activity and selectivity of magainin 2 amide in interaction with membranes. Biochemistry 1997, 36, 6124–6132. [Google Scholar] [CrossRef] [PubMed]
- Won, H.-S.; Kang, S.-J.; Lee, B.-J. Action mechanism and structural requirements of the antimicrobial peptides, gaegurins. Biochim. Biophys. Acta Biomembr. 2009, 1788, 1620–1629. [Google Scholar] [CrossRef]
- Teixeira, V.; Feio, M.J.; Bastos, M. Role of lipids in the interaction of antimicrobial peptides with membranes. Prog. Lipid. Res. 2012, 51, 149–177. [Google Scholar] [CrossRef]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Ding, L.; Yang, L.; Weiss, T.M.; Waring, A.J.; Lehrer, R.I.; Huang, H.W. Interaction of Antimicrobial Peptides with Lipopolysaccharides. Biochemistry 2003, 42, 12251–12259. [Google Scholar] [CrossRef]
- Shang, D.; Zhang, Q.; Dong, W.; Liang, H.; Bi, X. The effects of LPS on the activity of Trp-containing antimicrobial peptides against Gram-negative bacteria and endotoxin neutralization. Acta Biomater. 2016, 33, 153–165. [Google Scholar] [CrossRef]
- Xiao, W.; Sun, R.; Lou, J.; Xu, Y.; Li, X.; Xin, K.; Lu, W.; Sun, C.; Chen, T.; Gao, Y.; et al. LPS-enriched interaction drives spectrum conversion in antimicrobial peptides: Design and optimization of AA16 derivatives for targeting gram-negative bacteria. Eur. J. Med. Chem. 2025, 289, 117462. [Google Scholar] [CrossRef]
- Jia, F.; Wang, J.; Peng, J.; Zhao, P.; Kong, Z.; Wang, K.; Yan, W.; Wang, R. D-amino acid substitution enhances the stability of antimicrobial peptide polybia-CP. Acta Biochim. Biophys. Sin. 2017, 49, 916–925. [Google Scholar] [CrossRef]
- Kapil, S.; Sharma, V. d-Amino acids in antimicrobial peptides: A potential approach to treat and combat antimicrobial resistance. Can. J. Microbiol. 2021, 67, 119–137. [Google Scholar] [CrossRef]
- Garton, M.; Nim, S.; Stone, T.A.; Wang, K.E.; Deber, C.M.; Kim, P.M. Method to generate highly stable D-amino acid analogs of bioactive helical peptides using a mirror image of the entire PDB. Proc. Natl. Acad. Sci. USA 2018, 115, 1505–1510. [Google Scholar] [CrossRef]
- Lu, J.; Xu, H.; Xia, J.; Ma, J.; Xu, J.; Li, Y.; Feng, J. D-and unnatural amino acid substituted antimicrobial peptides with improved proteolytic resistance and their proteolytic degradation characteristics. Front. Microbiol. 2020, 11, 563030. [Google Scholar] [CrossRef]
- Liang, Y.; Zhang, X.; Yuan, Y.; Bao, Y.; Xiong, M. Role and modulation of the secondary structure of antimicrobial peptides to improve selectivity. Biomater. Sci. 2020, 8, 6858–6866. [Google Scholar] [CrossRef]
- Di Grazia, A.; Cappiello, F.; Cohen, H.; Casciaro, B.; Luca, V.; Pini, A.; Di, Y.P.; Shai, Y.; Mangoni, M.L. D-Amino acids incorporation in the frog skin-derived peptide esculentin-1a (1-21) NH 2 is beneficial for its multiple functions. Amino Acids 2015, 47, 2505–2519. [Google Scholar] [CrossRef]
- Ghaly, G.; Tallima, H.; Dabbish, E.; Badr ElDin, N.; Abd El-Rahman, M.K.; Ibrahim, M.A.A.; Shoeib, T. Anti-Cancer Peptides: Status and Future Prospects. Molecules 2023, 28, 1148. [Google Scholar] [CrossRef]
- Ran, S.; Downes, A.; Thorpe, P.E. Increased exposure of anionic phospholipids on the surface of tumor blood vessels. Cancer Res. 2002, 62, 6132–6140. [Google Scholar]
- Birge, R.B.; Boeltz, S.; Kumar, S.; Carlson, J.; Wanderley, J.; Calianese, D.; Barcinski, M.; Brekken, R.A.; Huang, X.; Hutchins, J.T.; et al. Phosphatidylserine is a global immunosuppressive signal in efferocytosis, infectious disease, and cancer. Cell Death Differ. 2016, 23, 962–978. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Wu, Q.; Sun, A.; Liu, X.; Fan, Y.; Deng, X. Cancer Cell Glycocalyx and Its Significance in Cancer Progression. Int. J. Mol. Sci. 2018, 19, 2484. [Google Scholar] [CrossRef]
- Ju, X.; Fan, D.; Kong, L.; Yang, Q.; Zhu, Y.; Zhang, S.; Su, G.; Li, Y. Antimicrobial Peptide Brevinin-1RL1 from Frog Skin Secretion Induces Apoptosis and Necrosis of Tumor Cells. Molecules 2021, 26, 2059. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, L.; Ma, C.; Zhang, Y.; Xi, X.; Wang, L.; Zhou, M.; Burrows, J.F.; Chen, T. A novel antimicrobial peptide, Ranatuerin-2PLx, showing therapeutic potential in inhibiting proliferation of cancer cells. Biosci. Rep. 2018, 38, BSR20180710. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y. I-TASSER server for protein 3D structure prediction. BMC Bioinform. 2008, 9, 40. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A unified platform for automated protein structure and function prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Meng, E.C.; Goddard, T.D.; Pettersen, E.F.; Couch, G.S.; Pearson, Z.J.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Tools for structure building and analysis. Protein Sci. 2023, 32, e4792. [Google Scholar] [CrossRef] [PubMed]
- Desbois, A.P.; Coote, P.J. Wax moth larva (Galleria mellonella): An in vivo model for assessing the efficacy of antistaphylococcal agents. J. Antimicrob. Chemother. 2011, 66, 1785–1790. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence | Hydrophobicity | Hydrophobic Moment | Net Charge |
|---|---|---|---|---|
| B2OS | GLWDTIKQAGKKIFLSVLDKIRCKVAGGC | 0.454 | 0.278 | +4 |
| B2OS(1-22)-NH2 | GLWDTIKQAGKKIFLSVLDKIR-NH2 | 0.435 | 0.462 | +3 |
| [D-Leu2]B2OS(1-22)-NH2 | G(D-Leu)WDTIKQAGKKIFLSVLDKIR-NH2 | 0.435 | 0.462 | +3 |
| Microorganisms | MICs/MBCs | ||||
|---|---|---|---|---|---|
| B2OS | B2OS(1-22)-NH2 | [D-Leu2] B2OS(1-22)-NH2 | Vancomycin | Colistin | |
| S. aureus (NCTC 6538) | 8/16 | 2/4 | 2/2 | 0.5/0.5 | N/A * |
| MRSA (NCTC 12493) | 32/64 | 8/8 | 8/8 | 0.5/0.5 | NA |
| E. faecalis (NCTC 12697) | 8/16 | 8/16 | 8/16 | 0.5/>64 | NA |
| E. coli (ATCC 8739) | 16/16 | 16/16 | 8/8 | N/A | 0.125/0.25 |
| K. pneumoniae (ATCC 43816) | 32/64 | 16/32 | 16/16 | N/A | 16/16 |
| P. aeruginosa (ATCC 9027) | 64/128 | 32/64 | 32/32 | N/A | 1/2 |
| Peptides | HC50 ± 95% CI (μM) 1 | TI 2 | ||
|---|---|---|---|---|
| TI+ | TI− | TI (all) | ||
| B2OS | 10.44 (8.428–12.93) | 0.82 | 0.33 | 0.52 |
| B2OS(1-22)-NH2 | 41.88 (31.03–56.52) | 8.31 | 2.08 | 4.15 |
| [D-Leu2]B2OS(1-22)-NH2 | 118.1 (48.36–288.1) | 23.48 | 7.381 | 13.15 |
| Microorganisms | MBICs/MBECs | ||
|---|---|---|---|
| B2OS | B2OS(1-22)-NH2 | [D-Leu2]B2OS(1-22)-NH2 | |
| S. aureus (NCTC 6538) | 32/512 | 4/256 | 4/128 |
| MRSA (NCTC 12493) | 64/>512 | 8/128 | 8/128 |
| E. faecalis (NCTC 12697) | 32/>512 | 16/>512 | 8/512 |
| E. coli (ATCC 8739) | 32/256 | 16/256 | 16/128 |
| K. pneumoniae (ATCC 43816) | 64/512 | 32/256 | 32/256 |
| P. aeruginosa (ATCC 9027) | 128/>512 | 64/>512 | 32/512 |
| Cell Line | IC50 ± 95% CI (µM) 1 | ||
|---|---|---|---|
| B2OS | B2OS(1-22)-NH2 | [D-Leu2]B2OS(1-22)-NH2 | |
| H838 | 3.624 (2.756–4.101) | 6.363 (5.013–8.078) | 4.339 (3.242–5.808) |
| PC-3 | 8.065 (6.777–9.598) | 7.004 (5.685–8.630) | 8.551 (6.932–10.55) |
| U251MG | 11.52 (9.103–14.57) | 15.05 (11.90–19.02) | 13.88 (9.305–20.69) |
| MCF-7 | 4.022 (3.158–5.122) | 5.571 (4.238–7.323) | 4.719 (3.682–6.049) |
| HCT116 | 8.052 (6.622–9.790) | 12.29 (10.44–14.47) | 13.86 (11.92–16.11) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, A.; Zhang, Z.; Song, Z.; Yuan, Y.; Chen, X.; Ma, C.; Chen, T.; Shaw, C.; Zhou, M.; Wang, L. Rational Engineering of a Brevinin-2 Peptide: Decoupling Potency from Toxicity Through C-Terminal Truncation and N-Terminal Chiral Substitution. Antibiotics 2025, 14, 784. https://doi.org/10.3390/antibiotics14080784
Yao A, Zhang Z, Song Z, Yuan Y, Chen X, Ma C, Chen T, Shaw C, Zhou M, Wang L. Rational Engineering of a Brevinin-2 Peptide: Decoupling Potency from Toxicity Through C-Terminal Truncation and N-Terminal Chiral Substitution. Antibiotics. 2025; 14(8):784. https://doi.org/10.3390/antibiotics14080784
Chicago/Turabian StyleYao, Aifang, Zeyu Zhang, Zhengmin Song, Yi Yuan, Xiaoling Chen, Chengbang Ma, Tianbao Chen, Chris Shaw, Mei Zhou, and Lei Wang. 2025. "Rational Engineering of a Brevinin-2 Peptide: Decoupling Potency from Toxicity Through C-Terminal Truncation and N-Terminal Chiral Substitution" Antibiotics 14, no. 8: 784. https://doi.org/10.3390/antibiotics14080784
APA StyleYao, A., Zhang, Z., Song, Z., Yuan, Y., Chen, X., Ma, C., Chen, T., Shaw, C., Zhou, M., & Wang, L. (2025). Rational Engineering of a Brevinin-2 Peptide: Decoupling Potency from Toxicity Through C-Terminal Truncation and N-Terminal Chiral Substitution. Antibiotics, 14(8), 784. https://doi.org/10.3390/antibiotics14080784