
The Role of the Environment (Water, Air, Soil) in the Emergence and Dissemination of Antimicrobial Resistance: A One Health Perspective

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Environmental Pathways of Antimicrobial Resistance Transmission
2.1. Aquatic Ecosystems: Critical Reservoirs and Transmission Pathways for Antimicrobial Resistance
2.1.1. Key Sources of AMR Contamination in Aquatic Systems
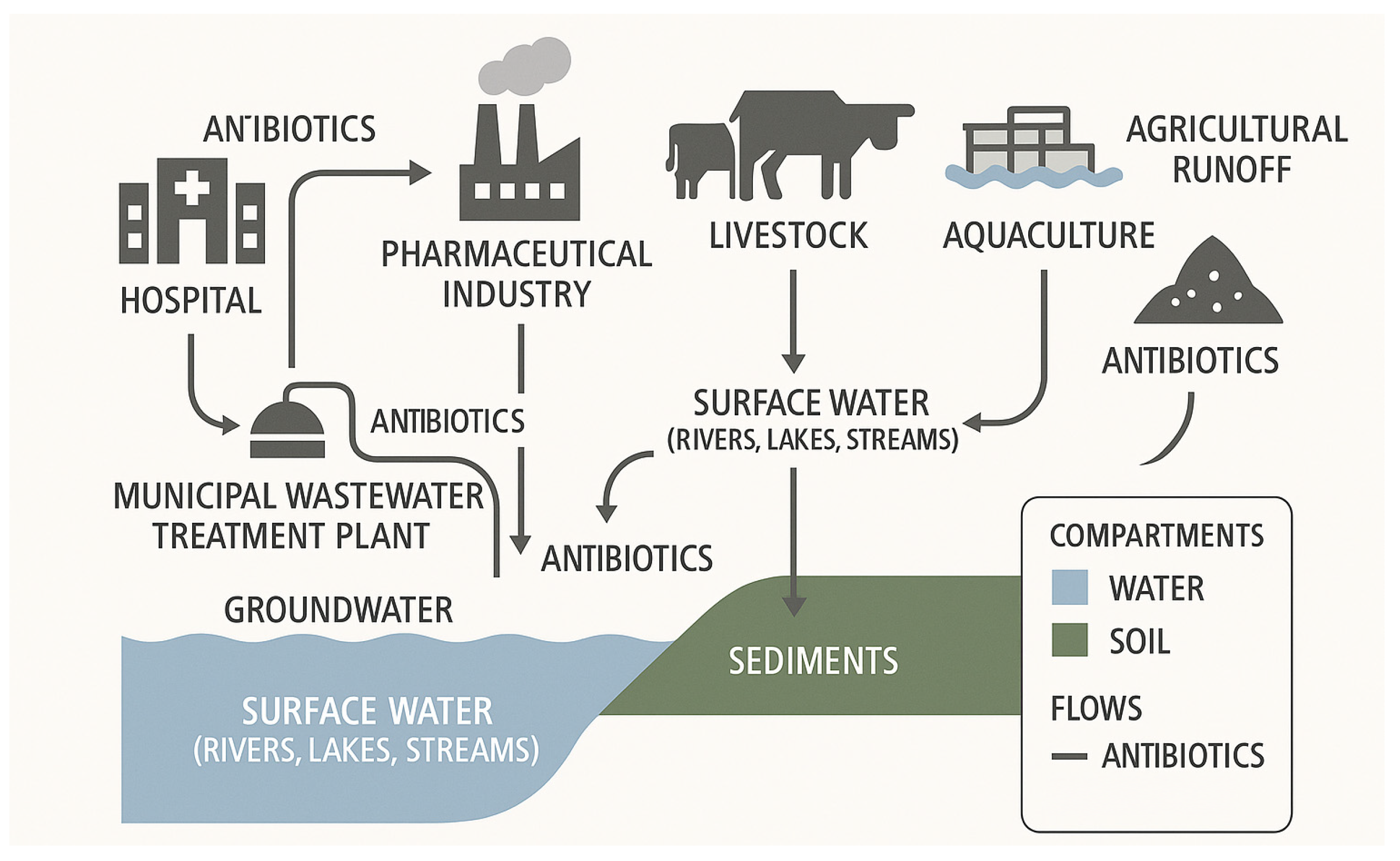
2.1.2. Mechanisms of Resistance Propagation in Aquatic Systems

2.1.3. Challenges in Wastewater Treatment and Mitigation Strategies
2.2. Terrestrial Reservoirs: Soil as a Dynamic Reservoir for Antimicrobial Resistance Evolution
2.2.1. Drivers of ARG Accumulation in Soil
2.2.2. Soil as a Hotspot for HGT
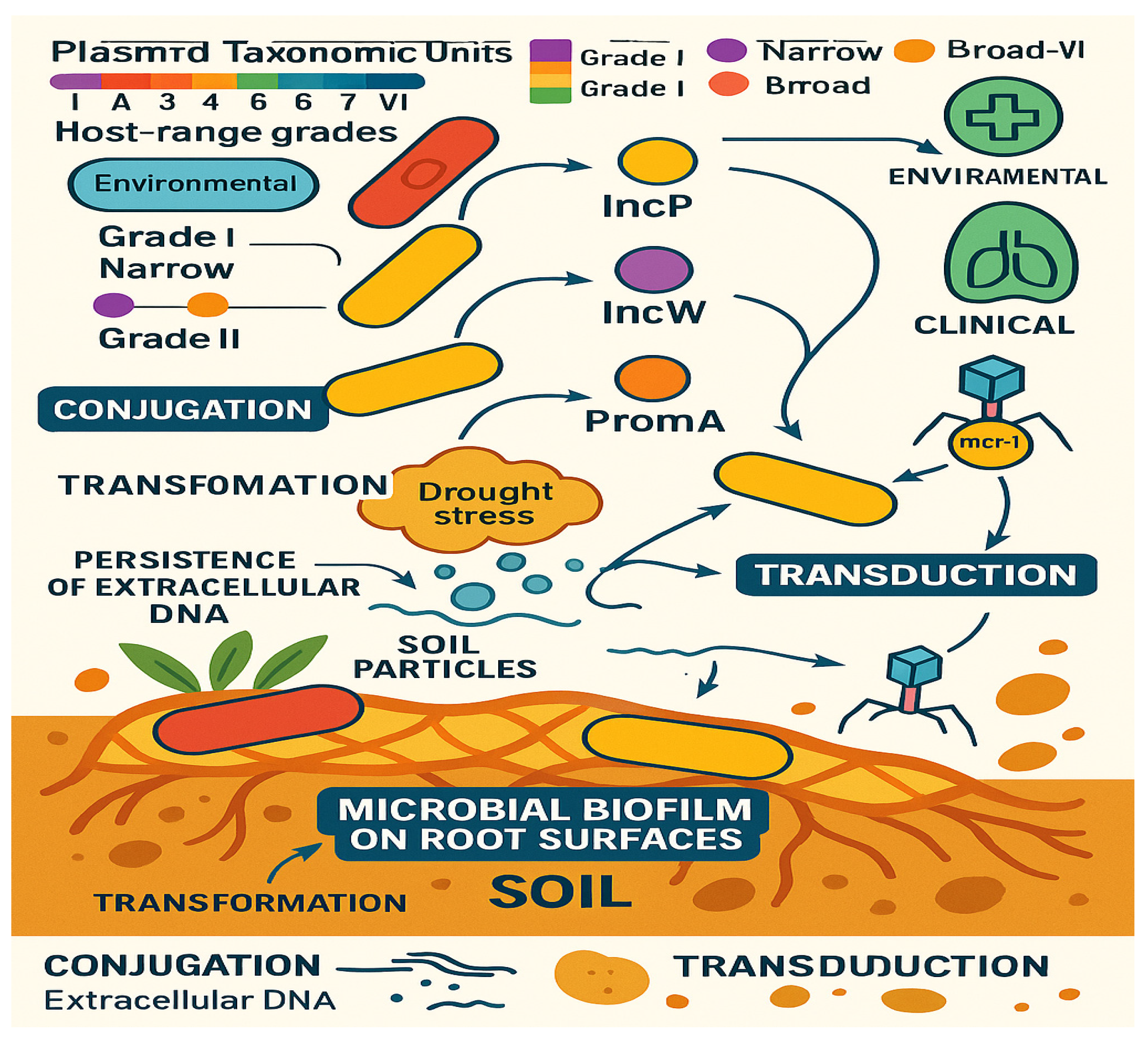
2.2.3. Metagenomic Insights into Soil Resistomes
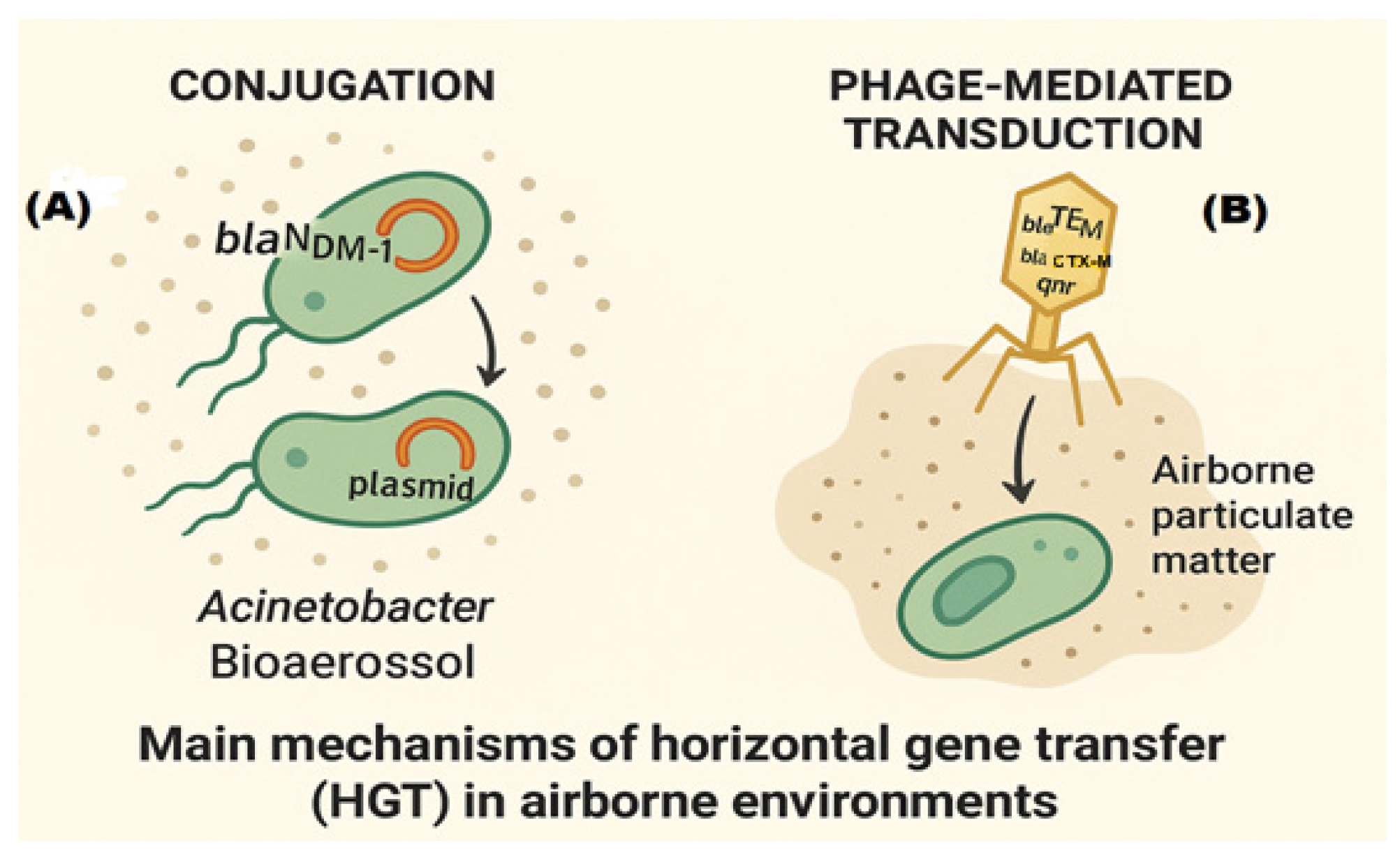
2.2.4. Atmospheric Transport Dynamics
2.2.5. Horizontal Gene Transfer in Air
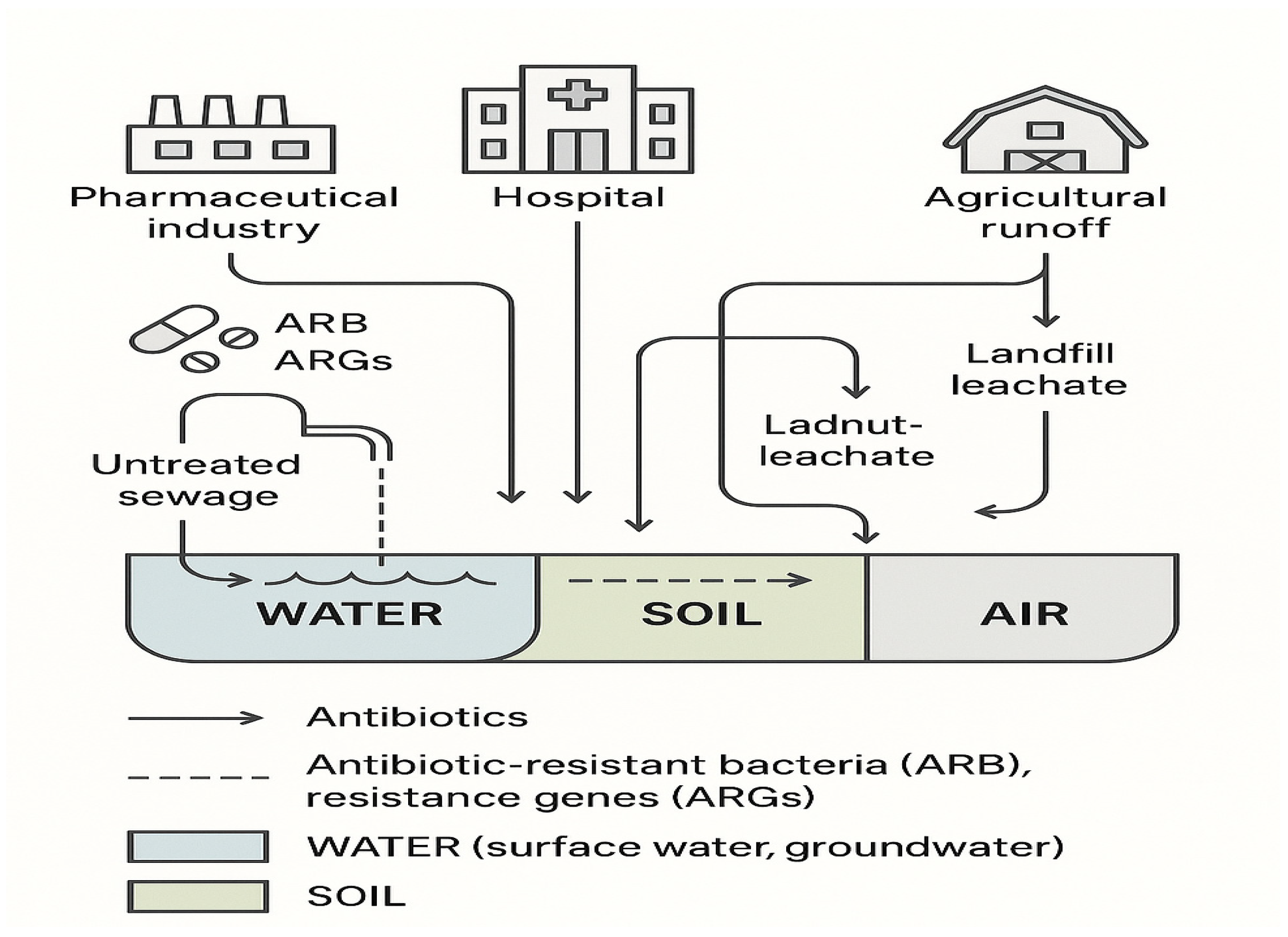
3. Impact of Environmental Pollution on AMR
3.1. Key Pollutants, Sources, Concentrations, and Global Locations
3.1.1. Pharmaceutical Waste
3.1.2. Wastewater (Treated and Untreated)
3.1.3. Agricultural Runoff
3.1.4. Industrial Waste and Microplastics
3.1.5. Air Pollution
3.2. Synergistic Effects of Combined Pollutants
3.3. Quantitative Evidence of Pollution—AMR Links
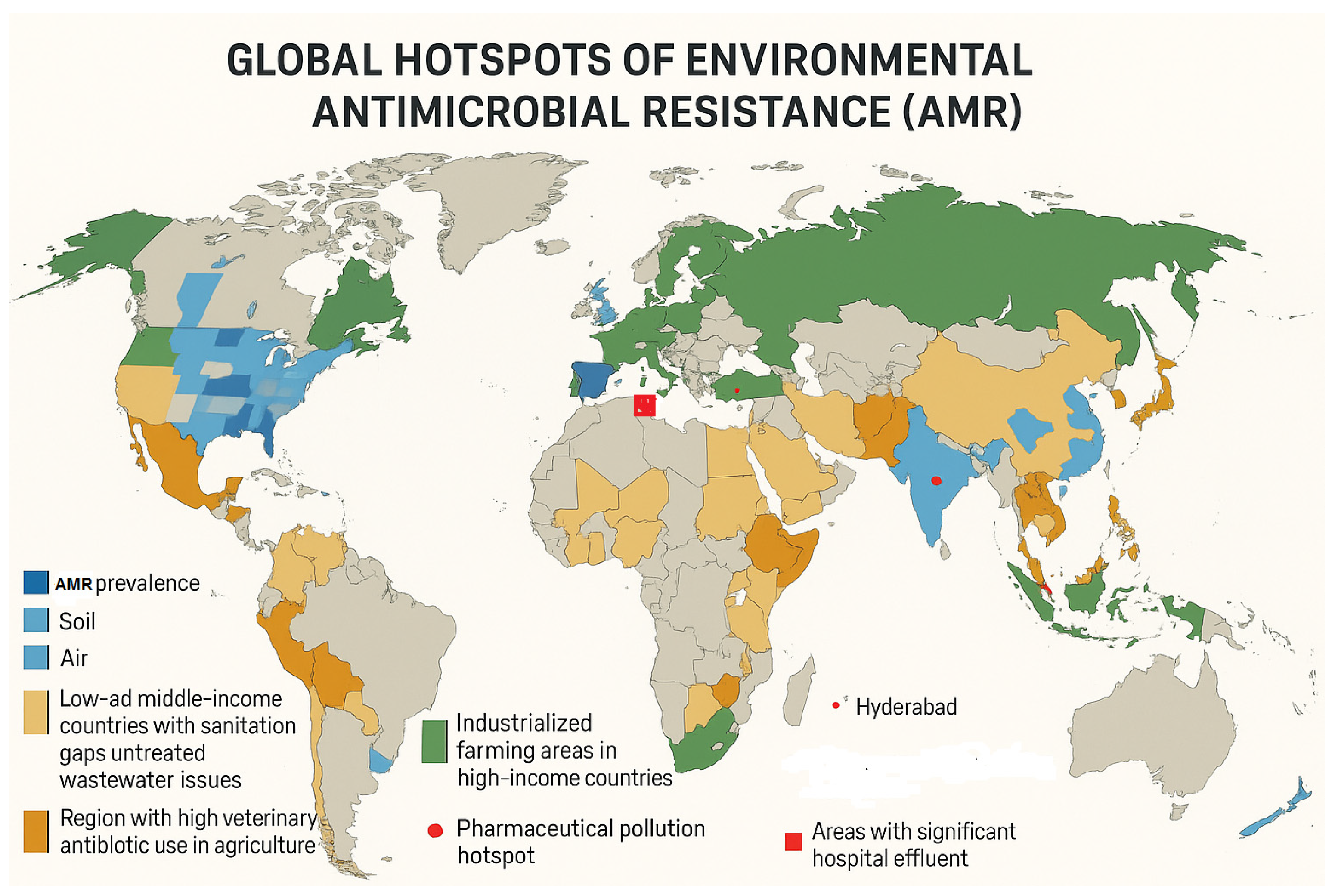
4. Antimicrobial Resistance in the Environment: A Global Perspective
4.1. Global Distribution of Environmental AMR Hotspots
4.2. Regional Case Studies
4.3. Transboundary AMR Spread
5. Strategies for Mitigating Environmental AMR: A Multidisciplinary Roadmap
5.1. Regulatory and Policy Interventions
5.2. Technological Innovations in Wastewater and Manure Treatment
5.3. One Health Integration
5.4. Global Surveillance and Monitoring Networks
5.5. Emerging Solutions and Future Directions
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. WHO List of Medically Important Antimicrobials: A Risk Management Tool for Mitigating Antimicrobial Resistance Due to Non-Human Use; World Health Organization: Geneva, Switzerland, 2024; ISBN 978-92-4-008461-2. [Google Scholar]
- Berendonk, T.U.; Manaia, C.M.; Merlin, C.; Fatta-Kassinos, D.; Cytryn, E.; Walsh, F.; Bürgmann, H.; Sørum, H.; Norström, M.; Pons, M.-N.; et al. Tackling Antibiotic Resistance: The Environmental Framework. Nat. Rev. Microbiol. 2015, 13, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Hendriksen, R.S.; Munk, P.; Njage, P.; van Bunnik, B.; McNally, L.; Lukjancenko, O.; Röder, T.; Nieuwenhuijse, D.; Pedersen, S.K.; Kjeldgaard, J.; et al. Global Monitoring of Antimicrobial Resistance Based on Metagenomics Analyses of Urban Sewage. Nat. Commun. 2019, 10, 1124. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson-Palme, J.; Kristiansson, E.; Larsson, D.G.J. Environmental Factors Influencing the Development and Spread of Antibiotic Resistance. FEMS Microbiol. Rev. 2018, 42, fux053. [Google Scholar] [CrossRef] [PubMed]
- von Wintersdorff, C.J.H.; Penders, J.; van Niekerk, J.M.; Mills, N.D.; Majumder, S.; van Alphen, L.B.; Savelkoul, P.H.M.; Wolffs, P.F.G. Dissemination of Antimicrobial Resistance in Microbial Ecosystems through Horizontal Gene Transfer. Front. Microbiol. 2016, 7, 173. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, J. Review on Antimicrobial Resistance: Tackling Drug-Resistant Infections Globally: Final Report and Recommendations; HM Government: London, UK, 2016.
- UNEP. Bracing for Superbugs: Strengthening Environmental Action in the One Health Response to Antimicrobial Resistance; UNEP: Nairobi, Kenya, 2023; ISBN 978-92-807-4006-6. [Google Scholar]
- Samreen; Ahmad, I.; Malak, H.A.; Abulreesh, H.H. Environmental Antimicrobial Resistance and Its Drivers: A Potential Threat to Public Health. J. Glob. Antimicrob. Resist. 2021, 27, 101–111. [Google Scholar] [CrossRef]
- Martinez, J.L. Environmental Pollution by Antibiotics and by Antibiotic Resistance Determinants. Environ. Pollut. Barking Essex 1987 2009, 157, 2893–2902. [Google Scholar] [CrossRef]
- Pruden, A.; Larsson, D.G.J.; Amézquita, A.; Collignon, P.; Brandt, K.K.; Graham, D.W.; Lazorchak, J.M.; Suzuki, S.; Silley, P.; Snape, J.R.; et al. Management Options for Reducing the Release of Antibiotics and Antibiotic Resistance Genes to the Environment. Environ. Health Perspect. 2013, 121, 878–885. [Google Scholar] [CrossRef]
- Rizzo, L.; Manaia, C.; Merlin, C.; Schwartz, T.; Dagot, C.; Ploy, M.C.; Michael, I.; Fatta-Kassinos, D. Urban Wastewater Treatment Plants as Hotspots for Antibiotic Resistant Bacteria and Genes Spread into the Environment: A Review. Sci. Total Environ. 2013, 447, 345–360. [Google Scholar] [CrossRef]
- La Rosa, M.C.; Maugeri, A.; Favara, G.; La Mastra, C.; Magnano San Lio, R.; Barchitta, M.; Agodi, A. The Impact of Wastewater on Antimicrobial Resistance: A Scoping Review of Transmission Pathways and Contributing Factors. Antibiotics 2025, 14, 131. [Google Scholar] [CrossRef]
- Cacace, D.; Fatta-Kassinos, D.; Manaia, C.M.; Cytryn, E.; Kreuzinger, N.; Rizzo, L.; Karaolia, P.; Schwartz, T.; Alexander, J.; Merlin, C.; et al. Antibiotic Resistance Genes in Treated Wastewater and in the Receiving Water Bodies: A Pan-European Survey of Urban Settings. Water Res. 2019, 162, 320–330. [Google Scholar] [CrossRef]
- Michael-Kordatou, I.; Karaolia, P.; Fatta-Kassinos, D. The Role of Operating Parameters and Oxidative Damage Mechanisms of Advanced Chemical Oxidation Processes in the Combat against Antibiotic-Resistant Bacteria and Resistance Genes Present in Urban Wastewater. Water Res. 2018, 129, 208–230. [Google Scholar] [CrossRef] [PubMed]
- Hosain, M.Z.; Kabir, S.M.L.; Kamal, M.M. Antimicrobial Uses for Livestock Production in Developing Countries. Vet. World 2021, 14, 210–221. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Y.; Xie, J.; Yuan, C.; Zhu, D.; Shi, X. Vertical Migration and Leaching Behavior of Antibiotic Resistance Genes in Soil during Rainfall: Impact by Long-Term Fertilization. Water Res. 2024, 267, 122508. [Google Scholar] [CrossRef] [PubMed]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global Trends in Antimicrobial Use in Food Animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef]
- Price, L.B.; Stegger, M.; Hasman, H.; Aziz, M.; Larsen, J.; Andersen, P.S.; Pearson, T.; Waters, A.E.; Foster, J.T.; Schupp, J.; et al. Staphylococcus aureus CC398: Host Adaptation and Emergence of Methicillin Resistance in Livestock. mBio 2012, 3, e00305-11. [Google Scholar] [CrossRef]
- Larsson, D.G.J.; Flach, C.-F. Antibiotic Resistance in the Environment. Nat. Rev. Microbiol. 2022, 20, 257–269. [Google Scholar] [CrossRef]
- Larsson, D.G.J.; De Pedro, C.; Paxeus, N. Effluent from Drug Manufactures Contains Extremely High Levels of Pharmaceuticals. J. Hazard. Mater. 2007, 148, 751–755. [Google Scholar] [CrossRef]
- Bhagat, C.; Kumar, M.; Tyagi, V.K.; Mohapatra, P.K. Proclivities for Prevalence and Treatment of Antibiotics in the Ambient Water: A Review. Npj Clean Water 2020, 3, 42. [Google Scholar] [CrossRef]
- MSc, P.P. Antibiotic Use by Humans Is Polluting Rivers Across the Globe, Study Shows. Available online: https://www.news-medical.net/news/20250424/Antibiotic-use-by-humans-is-polluting-rivers-across-the-globe-study-shows.aspx (accessed on 27 May 2025).
- Kenyon, C. Concentrations of Ciprofloxacin in the World’s Rivers Are Associated with the Prevalence of Fluoroquinolone Resistance in Escherichia Coli: A Global Ecological Analysis. Antibiotics 2022, 11, 417. [Google Scholar] [CrossRef]
- Fick, J.; Söderström, H.; Lindberg, R.H.; Phan, C.; Tysklind, M.; Larsson, D.G.J. Contamination of Surface, Ground, and Drinking Water from Pharmaceutical Production. Environ. Toxicol. Chem. 2009, 28, 2522–2527. [Google Scholar] [CrossRef]
- Maghsodian, Z.; Sanati, A.M.; Mashifana, T.; Sillanpää, M.; Feng, S.; Nhat, T.; Ramavandi, B. Occurrence and Distribution of Antibiotics in the Water, Sediment, and Biota of Freshwater and Marine Environments: A Review. Antibiotics 2022, 11, 1461. [Google Scholar] [CrossRef] [PubMed]
- Michaelis, C.; Grohmann, E. Horizontal Gene Transfer of Antibiotic Resistance Genes in Biofilms. Antibiotics 2023, 12, 328. [Google Scholar] [CrossRef] [PubMed]
- Abe, K.; Nomura, N.; Suzuki, S. Biofilms: Hot Spots of Horizontal Gene Transfer (HGT) in Aquatic Environments, with a Focus on a New HGT Mechanism. FEMS Microbiol. Ecol. 2020, 96, fiaa031. [Google Scholar] [CrossRef]
- Madsen, J.S.; Burmølle, M.; Hansen, L.H.; Sørensen, S.J. The Interconnection between Biofilm Formation and Horizontal Gene Transfer. FEMS Immunol. Med. Microbiol. 2012, 65, 183–195. [Google Scholar] [CrossRef]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile Genetic Elements Associated with Antimicrobial Resistance. Clin. Microbiol. Rev. 2018, 31, e00088-17. [Google Scholar] [CrossRef]
- Sanchez-Cid, C.; Guironnet, A.; Keuschnig, C.; Wiest, L.; Vulliet, E.; Vogel, T.M. Gentamicin at Sub-Inhibitory Concentrations Selects for Antibiotic Resistance in the Environment. ISME Commun. 2022, 2, 29. [Google Scholar] [CrossRef]
- Souque, C.; Escudero, J.A.; MacLean, R.C. Integron Activity Accelerates the Evolution of Antibiotic Resistance. eLife 2021, 10, e62474. [Google Scholar] [CrossRef]
- Andersson, D.I.; Hughes, D. Microbiological Effects of Sublethal Levels of Antibiotics. Nat. Rev. Microbiol. 2014, 12, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Mao, D.; Luo, Y.; Mathieu, J.; Wang, Q.; Feng, L.; Mu, Q.; Feng, C.; Alvarez, P.J.J. Persistence of Extracellular DNA in River Sediment Facilitates Antibiotic Resistance Gene Propagation. Environ. Sci. Technol. 2014, 48, 71–78. [Google Scholar] [CrossRef]
- Kunhikannan, S.; Thomas, C.J.; Franks, A.E.; Mahadevaiah, S.; Kumar, S.; Petrovski, S. Environmental Hotspots for Antibiotic Resistance Genes. MicrobiologyOpen 2021, 10, e1197. [Google Scholar] [CrossRef]
- Brovini, E.M.; Dias, M.; Teodoro, A.; Machado, E.; Santos, L.; Leal, C.; De Aquino, S.F.; De Araújo, J.C. Effects of UVC Doses on the Removal of Antimicrobial Resistance Elements from Secondary Treated Sewage. Environ. Sci. Pollut. Res. 2025, 32, 4325–4336. [Google Scholar] [CrossRef] [PubMed]
- McKinney, C.W.; Pruden, A. Ultraviolet Disinfection of Antibiotic Resistant Bacteria and Their Antibiotic Resistance Genes in Water and Wastewater. Environ. Sci. Technol. 2012, 46, 13393–13400. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, T.; Jikumaru, A.; Ueno, T.; Sei, K. Inactivation Effect of Antibiotic-Resistant Gene Using Chlorine Disinfection. Water 2017, 9, 547. [Google Scholar] [CrossRef]
- Shi, P.; Jia, S.; Zhang, X.-X.; Zhang, T.; Cheng, S.; Li, A. Metagenomic Insights into Chlorination Effects on Microbial Antibiotic Resistance in Drinking Water. Water Res. 2013, 47, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Subirats, J.; Sharpe, H.; Topp, E. Fate of Clostridia and Other Spore-Forming Firmicute Bacteria during Feedstock Anaerobic Digestion and Aerobic Composting. J. Environ. Manag. 2022, 309, 114643. [Google Scholar] [CrossRef]
- Plankaova, A.; Brajerova, M.; Capek, V.; Balikova Novotna, G.; Kinross, P.; Skalova, J.; Soltesova, A.; Drevinek, P.; Krutova, M. Clostridioides Difficile Infections Were Predominantly Driven by Fluoroquinolone-Resistant Clostridioides Difficile Ribotypes 176 and 001 in Slovakia in 2018–2019. Int. J. Antimicrob. Agents 2023, 62, 106824. [Google Scholar] [CrossRef]
- Uluseker, C.; Kaster, K.M.; Thorsen, K.; Basiry, D.; Shobana, S.; Jain, M.; Kumar, G.; Kommedal, R.; Pala-Ozkok, I. A Review on Occurrence and Spread of Antibiotic Resistance in Wastewaters and in Wastewater Treatment Plants: Mechanisms and Perspectives. Front. Microbiol. 2021, 12, 717809. [Google Scholar] [CrossRef]
- Honda, R.; Matsuura, N.; Sorn, S.; Asakura, S.; Morinaga, Y.; Van Huy, T.; Sabar, M.A.; Masakke, Y.; Hara-Yamamura, H.; Watanabe, T. Transition of Antimicrobial Resistome in Wastewater Treatment Plants: Impact of Process Configuration, Geographical Location and Season. npj Clean Water 2023, 6, 46. [Google Scholar] [CrossRef]
- Foroughi, M.; Khiadani, M.; Kakhki, S.; Kholghi, V.; Naderi, K.; Yektay, S. Effect of Ozonation-Based Disinfection Methods on the Removal of Antibiotic Resistant Bacteria and Resistance Genes (ARB/ARGs) in Water and Wastewater Treatment: A Systematic Review. Sci. Total Environ. 2022, 811, 151404. [Google Scholar] [CrossRef]
- Wang, S.; Yang, S.; Quispe, E.; Yang, H.; Sanfiorenzo, C.; Rogers, S.W.; Wang, K.; Yang, Y.; Hoffmann, M.R. Removal of Antibiotic Resistant Bacteria and Genes by UV-Assisted Electrochemical Oxidation on Degenerative TiO2 Nanotube Arrays. ACS EST Eng. 2021, 1, 612–622. [Google Scholar] [CrossRef]
- Xiao, X.; Bai, L.; Wang, S.; Liu, L.; Qu, X.; Zhang, J.; Xiao, Y.; Tang, B.; Li, Y.; Yang, H.; et al. Chlorine Tolerance and Cross-Resistance to Antibiotics in Poultry-Associated Salmonella Isolates in China. Front. Microbiol. 2022, 12, 833743. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.; Liu, F.; Tian, Y.; Zhang, J.; Liu, H.; Duan, J.; Bi, W.; Qin, J.; Xu, S. Effect of Biochar on Antibiotics and Antibiotic Resistance Genes Variations during Co-Composting of Pig Manure and Corn Straw. Front. Bioeng. Biotechnol. 2022, 10, 960476. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.-Q.; Zhang, X.-Y.; Zhang, H.-C.; Qiu, H.-B.; Li, Z.-H.; Xie, D.-H.; Yuan, L.; Sheng, G.-P. Gamma-Ray Irradiation as an Effective Method for Mitigating Antibiotic Resistant Bacteria and Antibiotic Resistance Genes in Aquatic Environments. J. Hazard. Mater. 2024, 468, 133791. [Google Scholar] [CrossRef]
- Heuer, H.; Schmitt, H.; Smalla, K. Antibiotic Resistance Gene Spread Due to Manure Application on Agricultural Fields. Curr. Opin. Microbiol. 2011, 14, 236–243. [Google Scholar] [CrossRef]
- Sarmah, A.K.; Meyer, M.T.; Boxall, A.B.A. A Global Perspective on the Use, Sales, Exposure Pathways, Occurrence, Fate and Effects of Veterinary Antibiotics (VAs) in the Environment. Chemosphere 2006, 65, 725–759. [Google Scholar] [CrossRef]
- Menz, J.; Olsson, O.; Kümmerer, K. Antibiotic Residues in Livestock Manure: Does the EU Risk Assessment Sufficiently Protect against Microbial Toxicity and Selection of Resistant Bacteria in the Environment? J. Hazard. Mater. 2019, 379, 120807. [Google Scholar] [CrossRef]
- Cycoń, M.; Mrozik, A.; Piotrowska-Seget, Z. Antibiotics in the Soil Environment—Degradation and Their Impact on Microbial Activity and Diversity. Front. Microbiol. 2019, 10, 338. [Google Scholar] [CrossRef]
- Zhu, Y.-G.; Johnson, T.A.; Su, J.-Q.; Qiao, M.; Guo, G.-X.; Stedtfeld, R.D.; Hashsham, S.A.; Tiedje, J.M. Diverse and Abundant Antibiotic Resistance Genes in Chinese Swine Farms. Proc. Natl. Acad. Sci. USA 2013, 110, 3435–3440. [Google Scholar] [CrossRef] [PubMed]
- Dharmasena, M.; Jiang, X. Isolation of Toxigenic Clostridium Difficile from Animal Manure and Composts Being Used as Biological Soil Amendments. Appl. Environ. Microbiol. 2018, 84, e00738-18. [Google Scholar] [CrossRef]
- Knapp, C.W.; Dolfing, J.; Ehlert, P.A.I.; Graham, D.W. Evidence of Increasing Antibiotic Resistance Gene Abundances in Archived Soils since 1940. Environ. Sci. Technol. 2010, 44, 580–587. [Google Scholar] [CrossRef]
- Seiler, C.; Berendonk, T.U. Heavy Metal Driven Co-Selection of Antibiotic Resistance in Soil and Water Bodies Impacted by Agriculture and Aquaculture. Front. Microbiol. 2012, 3, 399. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Chen, L.; Ye, C.; Yu, X. Co-Selection of Antibiotic Resistance via Copper Shock Loading on Bacteria from a Drinking Water Bio-Filter. Environ. Pollut. Barking Essex 1987 2018, 233, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Suprenant, M.P.; Ching, C.; Gross, N.; Sutradhar, I.; Anderson, J.E.; El Sherif, N.; Zaman, M.H. The Impact of Zinc Pre-Exposure on Ciprofloxacin Resistance Development in E. coli. Front. Microbiol. 2024, 15, 1491532. [Google Scholar] [CrossRef]
- Gillieatt, B.F.; Coleman, N.V. Unravelling the Mechanisms of Antibiotic and Heavy Metal Resistance Co-Selection in Environmental Bacteria. FEMS Microbiol. Rev. 2024, 48, fuae017. [Google Scholar] [CrossRef]
- Folvarska, V.; Marie Thomson, S.; Lu, Z.; Adelgren, M.; Schmidt, A.; Newton, R.J.; Wang, Y.; McNamara, P.J. The Effects of Lead, Copper, and Iron Corrosion Products on Antibiotic Resistant Bacteria and Antibiotic Resistance Genes. Environ. Sci. Adv. 2024, 3, 808–818. [Google Scholar] [CrossRef]
- Wales, A.D.; Davies, R.H. Co-Selection of Resistance to Antibiotics, Biocides and Heavy Metals, and Its Relevance to Foodborne Pathogens. Antibiotics 2015, 4, 567–604. [Google Scholar] [CrossRef]
- Rozman, U.; Pušnik, M.; Kmetec, S.; Duh, D.; Šostar Turk, S. Reduced Susceptibility and Increased Resistance of Bacteria against Disinfectants: A Systematic Review. Microorganisms 2021, 9, 2550. [Google Scholar] [CrossRef]
- Redondo-Salvo, S.; Fernández-López, R.; Ruiz, R.; Vielva, L.; de Toro, M.; Rocha, E.P.C.; Garcillán-Barcia, M.P.; de la Cruz, F. Pathways for Horizontal Gene Transfer in Bacteria Revealed by a Global Map of Their Plasmids. Nat. Commun. 2020, 11, 3602. [Google Scholar] [CrossRef]
- Shi, H.; Hu, X.; Xu, J.; Hu, B.; Ma, L.; Lou, L. Conjugation-Mediated Transfer of Antibiotic Resistance Genes Influenced by Primary Soil Components and Underlying Mechanisms. Sci. Total Environ. 2023, 865, 161232. [Google Scholar] [CrossRef]
- Nielsen, K.M.; van Weerelt, M.D.; Berg, T.N.; Bones, A.M.; Hagler, A.N.; van Elsas, J.D. Natural Transformation and Availability of Transforming DNA to Acinetobacter Calcoaceticus in Soil Microcosms. Appl. Environ. Microbiol. 1997, 63, 1945–1952. [Google Scholar] [CrossRef]
- Ross, J.; Topp, E. Abundance of Antibiotic Resistance Genes in Bacteriophage Following Soil Fertilization with Dairy Manure or Municipal Biosolids, and Evidence for Potential Transduction. Appl. Environ. Microbiol. 2015, 81, 7905–7913. [Google Scholar] [CrossRef] [PubMed]
- Ciach, M.A.; Pawłowska, J.; Górecki, P.; Muszewska, A. The Interkingdom Horizontal Gene Transfer in 44 Early Diverging Fungi Boosted Their Metabolic, Adaptive, and Immune Capabilities. Evol. Lett. 2024, 8, 526–538. [Google Scholar] [CrossRef]
- Liu, H.Y.; Prentice, E.L.; Webber, M.A. Mechanisms of Antimicrobial Resistance in Biofilms. npj Antimicrob. Resist. 2024, 2, 27. [Google Scholar] [CrossRef]
- Aminov, R.I. Horizontal Gene Exchange in Environmental Microbiota. Front. Microbiol. 2011, 2, 158. [Google Scholar] [CrossRef]
- Li, L.; Liu, Y.; Xiao, Q.; Xiao, Z.; Meng, D.; Yang, Z.; Deng, W.; Yin, H.; Liu, Z. Dissecting the HGT Network of Carbon Metabolic Genes in Soil-Borne Microbiota. Front. Microbiol. 2023, 14, 1173748. [Google Scholar] [CrossRef]
- Kilonzo-Nthenge, A.; Rafiqullah, I.; Netherland, M.; Nzomo, M.; Mafiz, A.; Nahashon, S.; Hasan, N.A. Comparative Metagenomics of Microbial Communities and Resistome in Southern Farming Systems: Implications for Antimicrobial Stewardship and Public Health. Front. Microbiol. 2024, 15, 1443292. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhang, N.; Li, C.; Shao, L. Diversity of Antibiotic Resistance Genes in Soils with Four Different Fertilization Treatments. Front. Microbiol. 2023, 14, 1291599. [Google Scholar] [CrossRef] [PubMed]
- Forsberg, K.J.; Patel, S.; Gibson, M.K.; Lauber, C.L.; Knight, R.; Fierer, N.; Dantas, G. Bacterial Phylogeny Structures Soil Resistomes across Habitats. Nature 2014, 509, 612–616. [Google Scholar] [CrossRef]
- Nesme, J.; Simonet, P. The Soil Resistome: A Critical Review on Antibiotic Resistance Origins, Ecology and Dissemination Potential in Telluric Bacteria. Environ. Microbiol. 2015, 17, 913–930. [Google Scholar] [CrossRef]
- Fitzpatrick, D.A. Horizontal Gene Transfer in Fungi. FEMS Microbiol. Lett. 2012, 329, 1–8. [Google Scholar] [CrossRef]
- Emamalipour, M.; Seidi, K.; Zununi Vahed, S.; Jahanban-Esfahlan, A.; Jaymand, M.; Majdi, H.; Amoozgar, Z.; Chitkushev, L.T.; Javaheri, T.; Jahanban-Esfahlan, R.; et al. Horizontal Gene Transfer: From Evolutionary Flexibility to Disease Progression. Front. Cell Dev. Biol. 2020, 8, 229. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhao, J.; Tang, N.; Sun, H.; Huang, J. Horizontal Gene Transfer from Bacteria and Plants to the Arbuscular Mycorrhizal Fungus Rhizophagus Irregularis. Front. Plant Sci. 2018, 9, 701. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Wang, S.-H.; Cheewangkoon, R.; Zhao, R.-L. Uneven Distribution of Prokaryote-Derived Horizontal Gene Transfer in Fungi: A Lifestyle-Dependent Phenomenon. mBio 2024, 16, e02855-24. [Google Scholar] [CrossRef]
- Semedo-Lemsaddek, T.; Bettencourt Cota, J.; Ribeiro, T.; Pimentel, A.; Tavares, L.; Bernando, F.; Oliveira, M. Resistance and Virulence Distribution in Enterococci Isolated from Broilers Reared in Two Farming Systems. Ir. Vet. J. 2021, 74, 22. [Google Scholar] [CrossRef]
- Kousar, S.; Rehman, N.; Javed, A.; Hussain, A.; Naeem, M.; Masood, S.; Ali, H.A.; Manzoor, A.; Khan, A.A.; Akrem, A.; et al. Intensive Poultry Farming Practices Influence Antibiotic Resistance Profiles in Pseudomonas Aeruginosa Inhabiting Nearby Soils. Infect. Drug Resist. 2021, 14, 4511–4516. [Google Scholar] [CrossRef]
- Szoke, Z.; Fauszt, P.; Mikolas, M.; David, P.; Szilagyi-Tolnai, E.; Pesti-Asboth, G.; Homoki, J.R.; Kovacs-Forgacs, I.; Gal, F.; Stundl, L.; et al. Comprehensive Analysis of Antimicrobial Resistance Dynamics among Broiler and Duck Intensive Production Systems. Sci. Rep. 2025, 15, 4673. [Google Scholar] [CrossRef] [PubMed]
- Cerqueira, F.; Matamoros, V.; Bayona, J.; Piña, B. Antibiotic Resistance Genes Distribution in Microbiomes from the Soil-Plant-Fruit Continuum in Commercial Lycopersicon Esculentum Fields under Different Agricultural Practices. Sci. Total Environ. 2019, 652, 660–670. [Google Scholar] [CrossRef]
- Baquero, F.; Martínez, J.-L.; Cantón, R. Antibiotics and Antibiotic Resistance in Water Environments. Curr. Opin. Biotechnol. 2008, 19, 260–265. [Google Scholar] [CrossRef]
- Wang, F.-H.; Qiao, M.; Su, J.-Q.; Chen, Z.; Zhou, X.; Zhu, Y.-G. High Throughput Profiling of Antibiotic Resistance Genes in Urban Park Soils with Reclaimed Water Irrigation. Environ. Sci. Technol. 2014, 48, 9079–9085. [Google Scholar] [CrossRef]
- Jones, R.M.; Brosseau, L.M. Aerosol Transmission of Infectious Disease. J. Occup. Environ. Med. 2015, 57, 501–508. [Google Scholar] [CrossRef]
- Bos, M.E.H.; Verstappen, K.M.; van Cleef, B.A.G.L.; Dohmen, W.; Dorado-García, A.; Graveland, H.; Duim, B.; Wagenaar, J.A.; Kluytmans, J.A.J.W.; Heederik, D.J.J. Transmission through Air as a Possible Route of Exposure for MRSA. J. Expo. Sci. Environ. Epidemiol. 2016, 26, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Cáliz, J.; Subirats, J.; Triadó-Margarit, X.; Borrego, C.M.; Casamayor, E.O. Global Dispersal and Potential Sources of Antibiotic Resistance Genes in Atmospheric Remote Depositions. Environ. Int. 2022, 160, 107077. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Shuai, X.; Lin, Z.; Yu, X.; Ba, X.; Holmes, M.A.; Xiao, Y.; Gu, B.; Chen, H. Association between Particulate Matter (PM)2·5 Air Pollution and Clinical Antibiotic Resistance: A Global Analysis. Lancet Planet. Health 2023, 7, e649–e659. [Google Scholar] [CrossRef]
- Zhou, X.-Y.; Li, H.; Zhou, S.-Y.-D.; Zhang, Y.-S.; Su, J.-Q. City-Scale Distribution of Airborne Antibiotic Resistance Genes. Sci. Total Environ. 2023, 856, 159176. [Google Scholar] [CrossRef]
- Becsei, Á.; Solymosi, N.; Csabai, I.; Magyar, D. Detection of Antimicrobial Resistance Genes in Urban Air. MicrobiologyOpen 2021, 10, e1248. [Google Scholar] [CrossRef] [PubMed]
- McCann, C.M.; Christgen, B.; Roberts, J.A.; Su, J.-Q.; Arnold, K.E.; Gray, N.D.; Zhu, Y.-G.; Graham, D.W. Understanding Drivers of Antibiotic Resistance Genes in High Arctic Soil Ecosystems. Environ. Int. 2019, 125, 497–504. [Google Scholar] [CrossRef]
- Habibi, N.; Uddin, S.; Behbehani, M.; Mustafa, A.S.; Al-Fouzan, W.; Al-Sarawi, H.A.; Safar, H.; Alatar, F.; Al Sawan, R.M.Z. Aerosol-Mediated Spread of Antibiotic Resistance Genes: Biomonitoring Indoor and Outdoor Environments. Int. J. Environ. Res. Public. Health 2024, 21, 983. [Google Scholar] [CrossRef]
- Zhang, K.; Zhou, X.; Zhang, X.; Huang, N.; Zhao, Z.; Zhang, X.; Zhou, Y.; Li, J.; Yu, F.; Liu, Y.; et al. Characterization of Transferable Antibiotic Resistance Plasmids in Airborne Particulate Matter from ICU Environments. iScience 2025, 28, 112254. [Google Scholar] [CrossRef]
- Mallonga, Z.; Tokuda, M.; Yamazaki, R.; Tsuruga, S.; Nogami, I.; Sato, Y.; Tarrayo, A.G.; Fuentes, R.; Parilla, R.; Kimbara, K.; et al. Emergence of Acinetobacter Towneri Harbouring a Novel Plasmid with blaNDM-1 and Tet(X7) from Hospital Wastewater in the Philippines. J. Glob. Antimicrob. Resist. 2025, 41, 287–289. [Google Scholar] [CrossRef]
- Tang, B.; Wang, C.; Sun, D.; Lin, H.; Ma, J.; Guo, H.; Yang, H.; Li, X. In Silico Characterization of BlaNDM -Harboring Conjugative Plasmids in Acinetobacter Species. Microbiol. Spectr. 2022, 10, e0210222. [Google Scholar] [CrossRef]
- Scoffone, V.C.; Trespidi, G.; Barbieri, G.; Arshad, A.; Israyilova, A.; Buroni, S. The Evolution of Antimicrobial Resistance in Acinetobacter Baumannii and New Strategies to Fight It. Antibiotics 2025, 14, 85. [Google Scholar] [CrossRef] [PubMed]
- Colomer-Lluch, M.; Jofre, J.; Muniesa, M. Antibiotic Resistance Genes in the Bacteriophage DNA Fraction of Environmental Samples. PLoS ONE 2011, 6, e17549. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Zhan, H.; Yu, X.; Li, J.; Wang, X.; Xie, Z. Detection of Organosulfates and Nitrooxy-Organosulfates in Arctic and Antarctic Atmospheric Aerosols, Using Ultra-High Resolution FT-ICR Mass Spectrometry. Sci. Total Environ. 2021, 767, 144339. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Cao, J.; Zhu, Y.-G.; Chen, Q.-L.; Shen, F.; Wu, Y.; Xu, S.; Fan, H.; Da, G.; Huang, R.-J.; et al. Global Survey of Antibiotic Resistance Genes in Air. Environ. Sci. Technol. 2018, 52, 10975–10984. [Google Scholar] [CrossRef]
- Nashwan, A.J.; Shah, H.H.; Hussain, T.; Rauf, S.A.; Ahmed, S.K. Environmental Drivers of Antimicrobial Resistance in Low and Middle-Income Countries: The Impacts of a Changing World. Environ. Health Insights 2024, 18, 11786302241246420. [Google Scholar] [CrossRef]
- New Global Guidance Aims to Curb Antibiotic Pollution from Manufacturing. Available online: https://www.who.int/news/item/03-09-2024-new-global-guidance-aims-to-curb-antibiotic-pollution-from-manufacturing (accessed on 28 May 2025).
- Sewage Study Highlights Global Antimicrobial Resistance Disparity|CIDRAP. Available online: https://www.cidrap.umn.edu/antimicrobial-stewardship/sewage-study-highlights-global-antimicrobial-resistance-disparity (accessed on 28 May 2025).
- Kelbrick, M.; Hesse, E.; O’ Brien, S. Cultivating Antimicrobial Resistance: How Intensive Agriculture Ploughs the Way for Antibiotic Resistance. Microbiology 2023, 169, 001384. [Google Scholar] [CrossRef]
- Rodgers, K.; McLellan, I.; Peshkur, T.; Williams, R.; Tonner, R.; Hursthouse, A.S.; Knapp, C.W.; Henriquez, F.L. Can the Legacy of Industrial Pollution Influence Antimicrobial Resistance in Estuarine Sediments? Environ. Chem. Lett. 2019, 17, 595–607. [Google Scholar] [CrossRef]
- Gothwal, R.; Shashidhar, T. Role of Environmental Pollution in Prevalence of Antibiotic Resistant Bacteria in Aquatic Environment of River: Case of Musi River, South India. Water Environ. J. 2017, 31, 456–462. [Google Scholar] [CrossRef]
- Asako, H.; Nakajima, H.; Kobayashi, K.; Kobayashi, M.; Aono, R. Organic Solvent Tolerance and Antibiotic Resistance Increased by Overexpression of marA in Escherichia coli. Appl. Environ. Microbiol. 1997, 63, 1428–1433. [Google Scholar] [CrossRef]
- Knapp, C.W.; McCluskey, S.M.; Singh, B.K.; Campbell, C.D.; Hudson, G.; Graham, D.W. Antibiotic Resistance Gene Abundances Correlate with Metal and Geochemical Conditions in Archived Scottish Soils. PLoS ONE 2011, 6, e27300. [Google Scholar] [CrossRef]
- Arias-Andres, M.; Klümper, U.; Rojas-Jimenez, K.; Grossart, H.-P. Microplastic Pollution Increases Gene Exchange in Aquatic Ecosystems. Environ. Pollut. Barking Essex 1987 2018, 237, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Ma, L.; Yu, Q.; Yang, J.; Su, W.; Hilal, M.G.; Li, X.; Zhang, S.; Li, H. The Source, Fate and Prospect of Antibiotic Resistance Genes in Soil: A Review. Front. Microbiol. 2022, 13, 976657. [Google Scholar] [CrossRef] [PubMed]
- Kraupner, N.; Ebmeyer, S.; Bengtsson-Palme, J.; Fick, J.; Kristiansson, E.; Flach, C.-F.; Larsson, D.G.J. Selective Concentration for Ciprofloxacin Resistance in Escherichia Coli Grown in Complex Aquatic Bacterial Biofilms. Environ. Int. 2018, 116, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Xu, Y.; Zeng, J.; Zhao, D.-H.; Li, L.; Liao, X.-P.; Liu, Y.-H.; Sun, J. Low-Concentration Ciprofloxacin Selects Plasmid-Mediated Quinolone Resistance Encoding Genes and Affects Bacterial Taxa in Soil Containing Manure. Front. Microbiol. 2016, 7, 1730. [Google Scholar] [CrossRef]
- Masclaux, F.G.; Sakwinska, O.; Charrière, N.; Semaani, E.; Oppliger, A. Concentration of Airborne Staphylococcus Aureus (MRSA and MSSA), Total Bacteria, and Endotoxins in Pig Farms. Ann. Occup. Hyg. 2013, 57, 550–557. [Google Scholar] [CrossRef]
- Progress on Household Drinking Water, Sanitation and Hygiene 2000–2022: Special Focus on Gender—UNICEF DATA. Available online: https://data.unicef.org/resources/jmp-report-2023/ (accessed on 23 April 2025).
- Van Boeckel, T.P.; Pires, J.; Silvester, R.; Zhao, C.; Song, J.; Criscuolo, N.G.; Gilbert, M.; Bonhoeffer, S.; Laxminarayan, R. Global Trends in Antimicrobial Resistance in Animals in Low- and Middle-Income Countries. Science 2019, 365, eaaw1944. [Google Scholar] [CrossRef]
- Silbergeld, E.K.; Graham, J.; Price, L.B. Industrial Food Animal Production, Antimicrobial Resistance, and Human Health. Annu. Rev. Public Health 2008, 29, 151–169. [Google Scholar] [CrossRef]
- Nasri, E.; Subirats, J.; Sànchez-Melsió, A.; Mansour, H.B.; Borrego, C.M.; Balcázar, J.L. Abundance of Carbapenemase Genes (blaKPC, blaNDM and blaOXA-48) in Wastewater Effluents from Tunisian Hospitals. Environ. Pollut. Barking Essex 1987 2017, 229, 371–374. [Google Scholar] [CrossRef]
- Falodun, O.I.; Morakinyo, Y.M.; Fagade, O.E. Determination of Water Quality and Detection of Extended Spectrum Beta-Lactamase Producing Gram-Negative Bacteria in Selected Rivers Located in Ibadan, Nigeria. Jordan J. Biol. Sci. 2018, 11, 107–112. [Google Scholar]
- Abigail, A.O.; Adebambo, M.A.; Gambo, E.D.; Ibrahim, A.M.; Ntasin, P.Y.; Opajobi, I.T.; Baba, I.M.; Abubakar, U.; Ishaku, N.M. Antibiotics Resistance Pattern and Molecular Detection of ESBL Genes in E. Coli from Both Surface and Underground Water Used for Domestic Purposes in Selected Locations in Ibadan, Oyo State, Nigeria. South Asian J. Res. Microbiol. 2024, 18, 40–57. [Google Scholar] [CrossRef]
- Calland, J.K.; Haukka, K.; Kpordze, S.W.; Brusah, A.; Corbella, M.; Merla, C.; Samuelsen, Ø.; Feil, E.J.; Sassera, D.; Karikari, A.B.; et al. Population Structure and Antimicrobial Resistance among Klebsiella Isolates Sampled from Human, Animal, and Environmental Sources in Ghana: A Cross-Sectional Genomic One Health Study. Lancet Microbe 2023, 4, e943–e952. [Google Scholar] [CrossRef] [PubMed]
- Le, T.X.; Munekage, Y. Residues of Selected Antibiotics in Water and Mud from Shrimp Ponds in Mangrove Areas in Viet Nam. Mar. Pollut. Bull. 2004, 49, 922–929. [Google Scholar] [CrossRef]
- Guenther, S.; Falgenhauer, L.; Semmler, T.; Imirzalioglu, C.; Chakraborty, T.; Roesler, U.; Roschanski, N. Environmental Emission of Multiresistant Escherichia coli Carrying the Colistin Resistance Gene Mcr-1 from German Swine Farms. J. Antimicrob. Chemother. 2017, 72, 1289–1292. [Google Scholar] [CrossRef]
- Drane, K.; Sheehan, M.; Whelan, A.; Ariel, E.; Kinobe, R. The Role of Wastewater Treatment Plants in Dissemination of Antibiotic Resistance: Source, Measurement, Removal and Risk Assessment. Antibiotics 2024, 13, 668. [Google Scholar] [CrossRef]
- Bartley, P.S.; Domitrovic, T.N.; Moretto, V.T.; Santos, C.S.; Ponce-Terashima, R.; Reis, M.G.; Barbosa, L.M.; Blanton, R.E.; Bonomo, R.A.; Perez, F. Antibiotic Resistance in Enterobacteriaceae from Surface Waters in Urban Brazil Highlights the Risks of Poor Sanitation. Am. J. Trop. Med. Hyg. 2019, 100, 1369–1377. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Kawahara, R.; Harada, K.; Teruya, S.; Nakayama, T.; Motooka, D.; Nakamura, S.; Nguyen, P.D.; Kumeda, Y.; Van Dang, C.; et al. The Presence of Colistin Resistance Gene Mcr-1 and -3 in ESBL Producing Escherichia coli Isolated from Food in Ho Chi Minh City, Vietnam. FEMS Microbiol. Lett. 2018, 365, fny100. [Google Scholar] [CrossRef]
- Litrup, E.; Kiil, K.; Hammerum, A.M.; Roer, L.; Nielsen, E.M.; Torpdahl, M. Plasmid-Borne Colistin Resistance Gene Mcr-3 in Salmonella Isolates from Human Infections, Denmark, 2009–17. Eurosurveillance 2017, 22, 30587. [Google Scholar] [CrossRef] [PubMed]
- Bai, H.; He, L.-Y.; Wu, D.-L.; Gao, F.-Z.; Zhang, M.; Zou, H.-Y.; Yao, M.-S.; Ying, G.-G. Spread of Airborne Antibiotic Resistance from Animal Farms to the Environment: Dispersal Pattern and Exposure Risk. Environ. Int. 2022, 158, 106927. [Google Scholar] [CrossRef] [PubMed]
- Regulation-2019/6-EN-EUR-Lex. Available online: https://eur-lex.europa.eu/eli/reg/2019/6/oj/eng (accessed on 23 April 2025).
- Veterinary Medicinal Products Regulation|European Medicines Agency (EMA). Available online: https://www.ema.europa.eu/en/veterinary-regulatory-overview/veterinary-medicinal-products-regulation (accessed on 23 April 2025).
- Tang, K.L.; Caffrey, N.P.; Nóbrega, D.B.; Cork, S.C.; Ronksley, P.E.; Barkema, H.W.; Polachek, A.J.; Ganshorn, H.; Sharma, N.; Kellner, J.D.; et al. Restricting the Use of Antibiotics in Food-Producing Animals and Its Associations with Antibiotic Resistance in Food-Producing Animals and Human Beings: A Systematic Review and Meta-Analysis. Lancet Planet. Health 2017, 1, e316–e327. [Google Scholar] [CrossRef]
- Implementing Decision-EU-2025/439-EN-EUR-Lex. Available online: https://eur-lex.europa.eu/eli/dec_impl/2025/439/oj/eng (accessed on 23 April 2025).
- Lüddeke, F.; Heß, S.; Gallert, C.; Winter, J.; Güde, H.; Löffler, H. Removal of Total and Antibiotic Resistant Bacteria in Advanced Wastewater Treatment by Ozonation in Combination with Different Filtering Techniques. Water Res. 2015, 69, 243–251. [Google Scholar] [CrossRef]
- NEWater|PUB, Singapore’s National Water Agency. Available online: https://www.pub.gov.sg/Public/WaterLoop/OurWaterStory/NEWater (accessed on 22 April 2025).
- Wang, J.; Ben, W.; Zhang, Y.; Yang, M.; Qiang, Z. Effects of Thermophilic Composting on Oxytetracycline, Sulfamethazine, and Their Corresponding Resistance Genes in Swine Manure. Environ. Sci. Process. Impacts 2015, 17, 1654–1660. [Google Scholar] [CrossRef] [PubMed]
- Ejileugha, C. Biochar Can Mitigate Co-Selection and Control Antibiotic Resistant Genes (ARGs) in Compost and Soil. Heliyon 2022, 8, e09543. [Google Scholar] [CrossRef] [PubMed]
- Global Antimicrobial Resistance and Use Surveillance System (GLASS) Report: 2022. Available online: https://www.who.int/publications/i/item/9789240062702 (accessed on 23 April 2025).
- JPIAMR. Available online: https://ncn.gov.pl/en/wspolpraca-zagraniczna/wspolpraca-wielostronna/konkurs-amr (accessed on 23 April 2025).
- Veldman, K.T.; Brouwer, M.S.M. MARAN 2024: Decline in Antibiotic Resistance in Animals Is Levelling off. Available online: https://www.wur.nl/en/research-results/research-institutes/bioveterinary-research/show-bvr/maran-2024-decline-in-antibiotic-resistance-in-animals-is-levelling-off.htm (accessed on 23 April 2025).
- Sumpradit, N.; Wongkongkathep, S.; Malathum, K.; Janejai, N.; Paveenkittiporn, W.; Yingyong, T.; Chuxnum, T.; Vijitleela, A.; Boonyarit, P.; Akaleephan, C.; et al. Thailand’s National Strategic Plan on Antimicrobial Resistance: Progress and Challenges. Bull. World Health Organ. 2021, 99, 661–673. [Google Scholar] [CrossRef]
- Su, H.; Hu, X.; Xu, W.; Xu, Y.; Wen, G.; Cao, Y. Diversity, Abundances and Distribution of Antibiotic Resistance Genes and Virulence Factors in the South China Sea Revealed by Metagenomic Sequencing. Sci. Total Environ. 2022, 814, 152803. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Pan, Z.; Wu, Y.; An, X.-L.; Wang, W.; Adamovich, B.; Zhu, Y.-G.; Su, J.-Q.; Huang, Q. A Database on the Abundance of Environmental Antibiotic Resistance Genes. Sci. Data 2024, 11, 250. [Google Scholar] [CrossRef]
- Resistomap—Monitor Antibiotic Resistance in the Environment. Available online: https://www.resistomap.com/ (accessed on 23 April 2025).
- Africa Knowledge Management Portal. Available online: https://khub.africacdc.org/records/resource?id=2203 (accessed on 23 April 2025).
- Kauppinen, A.; Siponen, S.; Pitkänen, T.; Holmfeldt, K.; Pursiainen, A.; Torvinen, E.; Miettinen, I.T. Phage Biocontrol of Pseudomonas Aeruginosa in Water. Viruses 2021, 13, 928. [Google Scholar] [CrossRef]
- Śmiałek, M.; Kowalczyk, J.; Koncicki, A. Influence of Vaccination of Broiler Chickens against Escherichia Coli with Live Attenuated Vaccine on General Properties of E. coli Population, IBV Vaccination Efficiency, and Production Parameters—A Field Experiment. Poult. Sci. 2020, 99, 5452–5460. [Google Scholar] [CrossRef]
- Arango-Argoty, G.; Garner, E.; Pruden, A.; Heath, L.S.; Vikesland, P.; Zhang, L. DeepARG: A Deep Learning Approach for Predicting Antibiotic Resistance Genes from Metagenomic Data. Microbiome 2018, 6, 23. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Technology | Main Advantages | Main Limitations | Potential Contribution to AMR | Reference |
|---|---|---|---|---|
| Conventional Activated Sludge | Cost-effective, widely used | Incomplete removal of antibiotics and ARGs | Residual antibiotics and ARGs in effluent | [41,42] |
| Membrane Filtration (MBR) | High removal efficiency for bacteria/particles | High cost, membrane fouling | Reduces ARB/ARGs, but not all; possible biofilm formation | [41,42] |
| Ozonation/Advanced Oxidation | Effective for many compounds | High energy cost, byproducts | Degrades some antibiotics/ARGs, but not all | [42,43] |
| UV Disinfection | Effective for bacteria, no chemical residues | Ineffective for extracellular DNA; limited by water turbidity | May not remove free ARGs; possible regrowth | [44] |
| Chlorination | Broad-spectrum disinfection, low cost | Can select for chlorine-resistant bacteria; toxic byproducts | Selection of resistant strains; incomplete ARG removal | [45] |
| Composting/Biosolids Recycling | Waste valorization, nutrient recycling | ARGs can persist in biosolids; risk if not fully sanitized | Spreads ARGs if not properly treated | [46] |
| Thermophilic Composting | High temperature degrades many ARGs | Not all ARGs eliminated; energy-intensive | Reduces but does not eliminate resistance genes | [46] |
| Biochar Amendment | Reduces extracellular DNA availability | Effectiveness varies with soil type and conditions | Limits horizontal transfer of ARGs | [46] |
| Gamma-ray Irradiation | Inactivates ARB and ARGs efficiently | High cost, limited large-scale application | Reduces ARGs and ARB in treated effluents | [47] |
| Pollutant Type | Major Sources | Key Resistance Elements | Typical Concentrations | Locations Detected | References |
|---|---|---|---|---|---|
| Pharmaceuticals | Drug manufacturing, hospitals | blaCTX-M, qnrS, mcr-1 | 10–31 mg/L (India), 1.2 mg/L (Europe), 2.5 mg/L (China) | Rivers and lakes in India, Europe, China, Africa, US | [24,32,103] |
| Wastewater (Treated and Untreated) | Urban wastewater, hospitals | vanA, blaKPC, intI1, ESBLs | Up to 108 ARG copies/mL | Coastal and inland waters, global | [10,11] |
| Agricultural Runoff | Manure, fertilizers, heavy metals | ermB, tetM, sul1, czcA | Up to 10× higher than organic soils | Europe, US, China, Africa | [55,88,104] |
| Industrial Waste and Microplastics | Textiles, plastics, dyes | sul1, ermB, marA, blaTEM | Notably high in estuaries, rivers | UK, China, Europe, Africa | [52,89,105] |
| Air Pollution | PM2.5, PM10, dust storms | blaCTX-M, mcr-1, sul1 | Up to 107 ARG copies/m2/day | China, Europe, US, Africa | [19,98] |
| Region | Key Drivers | Primary Pathways | Dominant ARB/ARGs | Human Health Impact | References |
|---|---|---|---|---|---|
| Sub-Saharan Africa | Untreated sewage | Water contamination | blaCTX-M-15, mcr-1 | Rise in pediatric MDR diarrhea | [116] |
| India and SE Asia | Pharmaceutical effluents, intensive aquaculture | River systems, aquaculture | blaNDM-1, qnrS | High rates of untreatable UTIs, aquaculture-associated resistance | [24,119] |
| Europe/N. America | Intensive farming, WWTP effluents | Agricultural runoff, WWTP effluents | erm(B), tet(M), blaCTX-M | MRSA in farmworkers; environmental dissemination of ESBLs | [120,121] |
| Latin America | Urban slums | Urban waterways | blaOXA-48, blaKPC, blaVIM-2, qnrS | Hospital CRE outbreaks, community transmission | [122] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sassi, A.; Basher, N.S.; Kirat, H.; Meradji, S.; Ibrahim, N.A.; Idres, T.; Touati, A. The Role of the Environment (Water, Air, Soil) in the Emergence and Dissemination of Antimicrobial Resistance: A One Health Perspective. Antibiotics 2025, 14, 764. https://doi.org/10.3390/antibiotics14080764
Sassi A, Basher NS, Kirat H, Meradji S, Ibrahim NA, Idres T, Touati A. The Role of the Environment (Water, Air, Soil) in the Emergence and Dissemination of Antimicrobial Resistance: A One Health Perspective. Antibiotics. 2025; 14(8):764. https://doi.org/10.3390/antibiotics14080764
Chicago/Turabian StyleSassi, Asma, Nosiba S. Basher, Hassina Kirat, Sameh Meradji, Nasir Adam Ibrahim, Takfarinas Idres, and Abdelaziz Touati. 2025. "The Role of the Environment (Water, Air, Soil) in the Emergence and Dissemination of Antimicrobial Resistance: A One Health Perspective" Antibiotics 14, no. 8: 764. https://doi.org/10.3390/antibiotics14080764
APA StyleSassi, A., Basher, N. S., Kirat, H., Meradji, S., Ibrahim, N. A., Idres, T., & Touati, A. (2025). The Role of the Environment (Water, Air, Soil) in the Emergence and Dissemination of Antimicrobial Resistance: A One Health Perspective. Antibiotics, 14(8), 764. https://doi.org/10.3390/antibiotics14080764