Abstract
Antibiotic resistance emerged shortly after the discovery of the first antibiotic and has remained a critical public health issue ever since. Managing antibiotic resistance in clinical settings continues to be challenging, particularly with the rise of superbugs, or bacteria resistant to multiple antibiotics, known as multidrug-resistant (MDR) bacteria. This rapid development of resistance has compelled researchers to continuously seek new antimicrobial agents to curb resistance, despite a shrinking pipeline of new drugs. Recently, the focus of antimicrobial discovery has shifted to plants, fungi, lichens, endophytes, and various marine sources, such as seaweeds, corals, and other microorganisms, due to their promising properties. For this review, an extensive search was conducted across multiple scientific databases, including PubMed, Elsevier, ResearchGate, Scopus, and Google Scholar, encompassing publications from 1929 to 2024. This review provides a concise overview of the mechanisms employed by bacteria to develop antibiotic resistance, followed by an in-depth exploration of plant secondary metabolites as a potential solution to MDR pathogens. In recent years, the interest in plant-based medicines has surged, driven by their advantageous properties. However, additional research is essential to fully understand the mechanisms of action and verify the safety of antimicrobial phytochemicals. Future prospects for enhancing the use of plant secondary metabolites in combating antibiotic-resistant pathogens will also be discussed.
1. Introduction
In the early 20th century, pharmacologist Alexander Fleming serendipitously discovered penicillin, the first natural antibiotic compound [1]. Interestingly, antibiotic resistance was identified in the same year, 1928 [2]. This groundbreaking discovery revolutionized the medical field, paving the way for the development of other antibiotics such as streptomycin, chloramphenicol, erythromycin, and chlortetracycline. Between 1960 and 1980, the pharmaceutical industry experienced a “golden period” in antibiotic development, producing numerous new antimicrobials [3].
After this period, the development of new antibiotics significantly declined as the pharmaceutical industry shifted its focus towards more profitable drug categories. This shift, combined with the rise in antimicrobial resistance, restricted the development of new antimicrobial agents [4,5]. Consequently, the increasing incidence of bacterial infections, coupled with their evolving multidrug resistance, has transformed them into major global health threats. The Comprehensive Antibiotic Resistance Database (CARD) reveals the existence of 5159 reference sequences. Although 381 of these are linked to relatively few significant pathogens, this offers little relief [6]. The situation is further exacerbated by the slow rate at which new generations of therapeutically effective antibiotics become available [7].
Antimicrobial resistance (AMR) refers to the ability of microorganisms, including bacteria, viruses, fungi, and parasites, to survive and multiply despite drug treatment [8]. Antimicrobial agents, such a antibiotics, antivirals, and antifungals, are designed to inhibit or eliminate pathogens. Among these, antibiotics are the most commonly used, particularly for bacterial infections. AMR develops naturally through genetic mutations, allowing organisms to adapt and survive [8]. In response to environmental pressures, bacteria evolve mechanisms to resist antimicrobial drugs, diminishing the effectiveness of these treatments [9]. The widespread use of antibiotics, especially in low-resource settings, provides bacteria with numerous opportunities to develop resistance, leading to significant health consequences, such as increased morbidity and mortality [10,11,12,13].
The rapid global rise of “superbugs” (microorganisms resistant to most available antimicrobials) underscores the urgent issue of drug-resistant pathogens. The World Health Organization (WHO) has identified AMR as one of the top three critical threats to global public health [14]. A report from 2019 estimated that antimicrobial-resistant infections were directly responsible for approximately 1.27 million deaths, and a study published in January 2022 indicated that nearly 5 million additional deaths were associated with drug-resistant infections [15]. Projections suggest that by 2050, the annual death toll from such infections could rise to 10 million, far surpassing cancer fatalities [13].
Methicillin-resistant Staphylococcus aureus (MRSA) exemplifies an early “superbug”, significantly contributing to global mortality rates from drug-resistant infections [16]. Bacteria, present in both domestic and professional environments, are among the earliest life forms on Earth [17]. While most bacterial species are harmless to human health, specific strains like Staphylococcus aureus, Helicobacter pylori, Escherichia coli, and Bacillus anthracis can breach host defenses and cause severe diseases such as pneumonia, endocarditis, septicemia, and osteomyelitis [18].
Healthcare-associated infections continue to pose a significant threat to patient safety and public health, often leading to severe complications and imposing considerable societal burdens [19]. Traditional preventive strategies for clinical infections primarily rely on aseptic techniques and systemic antibiotic therapies, but these methods frequently prove inadequate [20]. For instance, systemic antibiotic therapy for infections linked to medical devices, such as catheters, artificial prosthetics, subcutaneous sensors, and orthopedic implants, has an effectiveness rate of only 22–37% [21]. Moreover, high doses of antibiotics required for localized infections can result in cytotoxicity and adverse effects on surrounding tissues [22], further accelerating the emergence of bacterial drug resistance [23,24].
To combat infections caused by drug-resistant bacteria, scientists have developed various antimicrobial agents, including antibacterial peptides, amphiphiles, and antimicrobial materials such as nanoparticles, hydrogels, engineered surfaces, and coatings [25,26]. Despite these advancements, bacterial resistance remains a significant challenge. Current research is focused on discovering methods to eliminate bacteria without promoting resistance.
2. Methodology
To conduct this review, a comprehensive search was performed across several scientific databases, including PubMed, Elsevier, ResearchGate, Scopus, and Google Scholar. The search covered publications from 1929 to 2024, ensuring both a broad historical perspective and contemporary insights. A total of 278 publications were collected and thoroughly assessed.
The search strategy was designed to encompass a wide range of relevant literature, utilizing English MeSH (Medical Subject Headings) descriptors to ensure consistency and comprehensiveness. To maintain the quality and relevance of the selected studies, several criteria were applied. Only articles published in English that focused on the antibacterial properties of medicinal plant extracts were included. Duplicate articles, review articles, and studies evaluating isolated, commercially acquired molecules were excluded. This ensured that the review concentrated solely on the effects of plant extracts.
By implementing these methods, this review aimed to provide a thorough and detailed analysis of the antibacterial properties of medicinal plants, particularly their efficacy against species identified as priorities by the World Health Organization.
3. Strategies for Overcoming Antibiotic Resistance
The misuse and overuse of antibiotics has led to the emergence of multidrug-resistant bacteria, or superbugs, complicating infection treatment in clinical settings. This escalating issue necessitates innovative treatment strategies, which in turn require a comprehensive understanding of the mechanisms pathogenic bacteria employ to resist antibiotics.
3.1. Antibiotic Modification
Antibiotics can be categorized based on their targets: the bacterial cell envelope, the cytoplasm during protein synthesis, or the bacterial genome. To counteract antibiotics, bacteria produce modifying enzymes. For instance, bacteria resistant to β-lactam antibiotics produce β-lactamase, which hydrolyzes the β-lactam ring, rendering antibiotics like penicillins, carbapenems, monobactams, and cephalosporins ineffective.
Based on primary sequence homology, β-lactamase enzymes are grouped into four classes: A, B, C, and D. While classes A, C, and D are serine-based, class B is metal-based and includes a Zn2+ ion. Serine-based β-lactamases form a covalent acyl-enzyme intermediate with the antibiotic, which is then hydrolyzed. In contrast, class B β-lactamases use water molecules for hydrolysis without forming an intermediate [27].
To combat β-lactamase activity, one strategy involves combining β-lactam antibiotics with β-lactamase inhibitors like clavulanic acid and tazobactam. These inhibitors bind to the enzyme with higher affinity, allowing the antibiotic to remain effective. Another approach is modifying the antibiotic structure, such as removing the aminoadipoyl sidechain from cephalosporins to form 7-aminocephalosporanic acid, making them significantly more effective and resistant to β-lactamase hydrolysis.
3.2. Modification of Antibiotic Target Sites
Microorganisms employ various strategies to withstand antibiotic treatment, with one of the most effective being the modification of the antibiotic’s target site. This approach involves a minor gene mutation that alters the enzyme’s target site, reducing the binding affinity of antibiotics while maintaining the enzyme’s functionality. For example, β-lactam antibiotics like penicillin target and inactivate penicillin-binding protein (PBP), a transpeptidase that assists in cross-linking the peptidoglycan cell wall by acylating its active site. A mutation in the PBP gene can result in a slightly altered but still functional protein, preventing β-lactam antibiotics from binding effectively. Drug-resistant strains of Clostridium difficile, Enterococcus faecium, and Streptococcus pneumoniae exhibit this ability [28,29]. Microbial resistance to ciprofloxacin, which interferes with cellular division by targeting DNA gyrase and topoisomerase, also falls under this mechanism [30,31].
Another method of conferring resistance is through post-transcriptional and post-translational modifications of antibiotic target sites. Post-transcriptional modifications involve altering the primary RNA transcript to form mature RNA, which can then be translated into functional proteins. For instance, methylation of the tRNA anticodon stem loop prevents frame-shifting during translation. Methylation of specific 16S ribosomal RNA has been reported to prevent the binding of antibiotics like spectinomycin and streptomycin, thus allowing normal protein translation [32,33]. Post-translational modifications involve enzymatic alterations of proteins after synthesis. Common modifications in bacteria include phosphorylation and succinylation. For example, cysteine phosphorylation in eukaryotic-like kinase-phosphatases has been shown to confer resistance to vancomycin and ceftriaxone in methicillin-resistant Staphylococcus aureus (MRSA) [34]. Additionally, lysine succinylation in isocitrate lyase has been linked to resistance to rifampicin in Mycobacterium tuberculosis.
3.3. Antibiotic Resistence: Efflux Pump and Reduced Permeability
Bacteria have developed various strategies to counteract antibiotics by preventing these drugs from accumulating sufficiently within their cytoplasm. These strategies include overproducing efflux pumps in the bacterial cytoplasmic membrane and reducing membrane permeability.
Efflux pumps are active transport proteins that expel toxic compounds, including antibiotics, from the bacterial cytoplasm. They use energy derived from adenosine triphosphate (ATP) or an electrochemical potential gradient. For instance, bacteria such as Escherichia coli and Pseudomonas aeruginosa show significant resistance to ciprofloxacin and fluoroquinolones due to the overexpression of efflux pump proteins powered by the hydrogen ion gradient [35,36]. Instead of having pumps specific to certain antibiotics, bacteria like MRSA and P. aeruginosa produce non-specific multidrug resistance pumps that expel a wide range of antibiotics, including β-lactams [37,38,39].
Another resistance mechanism involves reducing the permeability of the bacterial outer membrane. Bacteria transport small polar molecules, amino acids, and nutrients through water-filled channels called porins [40]. These channels also permit the entry of antibiotics like penicillin and cephalosporins. To counteract this, bacteria can either decrease or silence the expression of porin proteins or alter their structure to reduce permeability. For example, drug-resistant strains of E. coli and Enterobacter aerogenes modify the structure of porins by narrowing the channels, thereby limiting antibiotic entry. Conversely, Klebsiella pneumoniae reduces antibiotic uptake by minimizing porin expression [41,42,43].
3.4. Enhancing Resistance through Target/Substrate Overproduction
The discovery of antibiotic modifiers like the β-lactamase enzyme paved the way for combinatorial treatments that pair antibiotics with β-lactamase inhibitors. However, it is not surprising that bacteria eventually become resistant to these inhibitors, especially with the frequent use of such combinations. This resistance arises in addition to the high mutation rates bacteria experience due to the direct effects of both the antibiotic and the inhibitor. Consequently, bacteria can develop resistance to β-lactamase inhibitors through the overproduction of the target enzyme, β-lactamase. This phenomenon has been observed in drug-resistant strains of E. coli and K. pneumoniae [44,45].
Another example is the overexpression of the enzyme dihydrofolate reductase, which is essential for nucleic acid precursor synthesis in M. tuberculosis and E. coli. Overproduction of this enzyme confers resistance to antimicrobial drugs such as para-aminosalicylic acid and trimethoprim [46,47].
3.5. Cell Wall Remodeling
Under typical physiological conditions, bacterial cell walls undergo constant remodeleling, with old peptidoglycan layers being degraded and replaced by new ones. Glycopeptide antibiotics primarily disrupt this cell wall construction process. For instance, teicoplanin and vancomycin inhibit the actions of glycosyltransferases and transpeptidases by binding to the peptidyl-D-Ala-D-Ala ends of peptidoglycan precursors [48,49]. This binding blocks transpeptidation and transglycosylation, preventing the bacterial cell from renewing its peptidoglycan wall. As the old wall breaks down due to treatment, the cell ultimately undergoes lysis and dies.
However, vancomycin-resistant Enterococci species can produce altered peptidoglycan precursors, such as peptidyl-D-Ala-D-Lac or peptidyl-D-Ala-D-Ser, which impede the binding of glycopeptide antibiotics [50,51]. These modifications arise through either gene acquisition or inherent genetic mutations. Specifically, the vanA gene cluster encodes VanH dehydrogenase, which converts pyruvate into D-Lac, and VanA ligase, which esterifies D-Ala with D-Lac to form peptidyl-D-Ala-D-Lac [52]. Such changes prevent the binding of glycopeptide antibiotics, while allowing glycosyltransferases and transpeptidases to continue synthesizing the cell wall.
Moreover, a single amino acid alteration in the Dd1 ligase, responsible for ligating two D-Ala molecules, can enable it to connect D-Ala to D-Lac instead, thereby conferring resistance to glycopeptide antibiotics. This mechanism has also been observed in vancomycin-resistant strains of E. coli.
4. Plant-Derived Antibiotics: A Solution to Multidrug-Resistant Microbes
Exploring innovative antibiotics derived from natural products is crucial for addressing the socio-economic and health impacts of multidrug-resistant microbes [53]. Plant-derived therapeutic agents have gained importance due to the emergence of new diseases and the growing scientific understanding of herbal medicines as alternative or complementary treatments [54] (Figure 1). Several plants and plant parts known for their antimicrobial properties and available on the market are enumerated in Table 1.
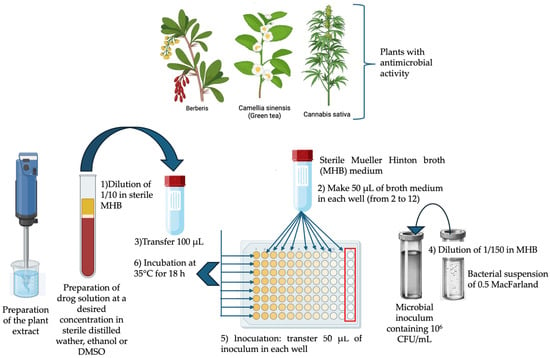
Figure 1.
Broth microdilution for antibacterial testing as recommended by the Clinical and Laboratory Standards Institute (CLSI) M07-A9: Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standard—Ninth Edition [55].
Research shows that medicinal plants contain bioactive compounds such as coumarins, flavonoids, phenolics, alkaloids, terpenoids, tannins, essential oils, lectins, polypeptides, and polyacetylenes [56,57,58], which serve as foundations for antibiotic development [58]. For instance, crude extracts from Polygonum persicaria, P. plebejum, Rumex hastatus, R. dentatus, R. nepalensis, and Rheum australe exhibit antibacterial and antifungal properties, inhibiting bacteria such as C. frundii, E. coli, E. aerogenes, and S. aureus [59]. Additionally, Calotropis gigantea extracts have shown significant antifungal activity against pathogenic fungi like Candida albicans and Aspergillus species in Asia [57]. Ethanolic extracts from Plumbago zeylanica roots exhibit strong antimicrobial effects against V. cholerae, E. coli, P. aeruginosa, Curvularia lunata, Colletotrichum corchori, and Fusarium equiseti [58]. The aqueous leaf extracts of Euphorbia hirta and Erythrophleum suaveolens, along with the methanolic leaf extract of Thevetia peruviana, show antibacterial effects against extended-spectrum beta-lactamase (ESBL)-producing bacteria, including E. coli, Pseudomonas, K. pneumonia, methicillin-resistant Staphylococcus aureus (MRSA), Salmonella, and Proteus [60,61,62,63]. Limited research on aqueous and hydro-alcoholic extracts from various plants has revealed antibacterial effects on multidrug-resistant bacteria, including MRSA and ESBL producers [64].
Methanolic and ethyl acetate extracts from Anacyclus maroccanus Ball and A. radiatus Loisel have been evaluated for their antimicrobial activity against a variety of bacterial, fungal, and dermatophyte species. Notably, E. coli and T. rubrum exhibited the highest sensitivity to these extracts [65]. The antimicrobial characteristics of aqueous extracts derived from the roots and leaves of Lactuca longidentata have been analyzed in relation to various microorganisms, including Gram-positive and Gram-negative bacteria, yeast strains from the Candida genus, dermatophytes such as species from the Trichophyton and Arthroderma genera, and fungi commonly found as contaminants in public swimming pools. Within the limits of biocompatibility (concentration < 200 µg/mL), the leaf extract demonstrated notable efficacy in inhibiting the growth of Escherichia coli and Trichophyton tonsurans, with a minimum inhibitory concentration (MIC) of less than 10 µg/mL [66]. The root and leaf extracts, both aqueous and hydroalcoholic, from female Cannabis sativa cv. Strawberry exhibit potent antimicrobial activity against Bacillus subtilis (PeruMycA 6). Most tested bacterial strains were highly susceptible to both hydroalcoholic extracts, which demonstrated MIC values below 62.99 µg/mL. These extracts also showed significant efficacy in inhibiting dermatophyte growth, with Arthroderma currey (CCF 5207) being the most sensitive fungal species, exhibiting MIC values less than 6.25 µg/mL [67]. These findings align with previous studies on the antifungal effects of water extracts from the inflorescences of the industrial hemp (Cannabis sativa L.) cultivar “Futura 75” [68].
Despite the approval of synthetic antimicrobial agents in numerous countries, natural compounds derived from plants [69,70], fungi [71], lichens [72], endophytes [73], and various marine sources, such as plants [74,75,76,77], seaweeds [78], corals [79], and other microorganisms [80,81], remain a focal point of substantial research interest [70]. These natural compounds have shown promise in combating antibiotic resistance in bacterial pathogens [82]. Among the various options, plant-derived compounds are notable for their potential in fighting bacterial infections. These naturally occurring plant chemicals have demonstrated significant benefits, including antioxidant, antibacterial, and antifungal activities. They can enhance the effectiveness of existing antibiotics, helping to prevent the development of resistance [83].
Based on their chemical structures, these compounds can be categorized into major groups such as alkaloids, sulfur-containing compounds, terpenoids, and polyphenols.

Table 1.
Some of the plant products with antimicrobial activity. This table is adapted from Khameneh et al. [84].
Table 1.
Some of the plant products with antimicrobial activity. This table is adapted from Khameneh et al. [84].
| Family | Scientific Name (Common Name) | Compound | Effective in Combating | Drug Delivery System |
|---|---|---|---|---|
| Berberidaceae | Berberis vulgaris (Barberry) | Berberine | Bacteria, protozoa | Soft gel 1000 mg |
| Piperaceae | Piper nigrum (Black pepper) | Piperine | Fungi, Lactobacillus, Micrococcus | |
| Asteraceae | Arctium lappa (Burdock) | Bacteria, fungi, virus | Capsule 475 mg | |
| Apiaceae | Carum carvi (Caraway) | Bacteria, fungi, virus | Capsule 1000 mg | |
| Rhamnaceae | Rhamnus purshiana (Cascara sagrada) | Tannins | Bacteria, fungi, virus | Capsule 425, 450 mg |
| Asteraceae | Matricaria chamomilla (Chamomille) | Anthemic acid | M. tuberulosis, S. typhimurium, S. aureus | |
| Apiaceae | Syzygium aromaticum (Clove) | Eugenol | General | Capsule 500 mg |
| Ericaceae | Vaccinium spp. (Cranberry) | Fructose | Bacteria | Capsule 500 mg |
| Myrtaceae | Eucalyptus globulus (Eucalyptus) | Tannins | Bacteria, virus | Inhaler and tablet |
| Amaryllidaceae | Allium sativum (Garlic) | Allicin, ajoene | General | Tablet |
| Asteraceae | Hydrastis canadensis (Goldenseal) | Berberine, hydrastine | Bacteria, Giarda duodenale, Trypanosomes | Solution, 500 mg per dosage |
| Theaceae | Camellia sinensis (Green tea) | Catechin | General | |
| Fabaceae | Glycyrrhiza glabra (Licorice) | Glabrol | S. aureus, M. tuberculosis | Capsule 450 mg |
| Fagaceae | Quercus rubra (Oak) | Tannins, Quercetin | Capsule 500, 650 mg | |
| Amaryllidaceae | Allium cepa (Onion) | Allcin | Bacteria, Candida | |
| Berberidaceae | Mahonia aquifolia (Oregon grape) | Berberine | Plasmodium, Trypansomes, general | Capsule 500 mg |
| Hypericaceae | Hypericum perforatum (Senna St. John’s wort) | Hypericin, others | General | Capsule 450 mg |
| Lamiaceae | Thymus vulgaris (Thyme) | Caffeic acid, Thymol, Tannins | Viruses, bacteria, fungi | Capsule 450 mg |
| Zingiberaceae | Curcuma longa (Turmeric) | Curcumin, Turmeric oil | Bacteria, protozoa |
5. Plant Secondary Metabolites as Antimicrobial Agent
5.1. Alkaloids
The term “alkaloid” denotes “similar to alkalis”, indicating the basic or alkaline nature of these substances. To date, around 12,000 alkaloid compounds have been extracted from plants and categorized. These compounds exhibit diverse medicinal properties, including antitumor, analgesic (such as morphine and codeine), and antimicrobial effects [85]. Their chemical structures feature heterocyclic rings with N-heterocyclic nitrogen and can be classified based on their carbon precursors and structural characteristics [86]. Their antibacterial activity is well-documented, and many studies suggest they play a crucial role in treating infectious diseases [87]. Most alkaloids function as efflux pump inhibitors (EPIs), which represents a key antibacterial mechanism [88].
Piperine, a piperidine-type alkaloid from Piper nigrum and Piper longum, when combined with ciprofloxacin, has been shown to inhibit the growth of a mutant S. aureus and significantly reduce its MIC values. Additionally, the co-administration of piperine and gentamicin proved effective against MRSA infections. Studies have shown that piperine impacts the NorA efflux pump activity in both S. aureus and MRSA [88,89].
Berberine, an isoquinoline alkaloid, is found in the roots and stem-bark of Berberis species. It is the primary active ingredient in Rhizoma coptidis and Cortex phellodendri and has long been utilized in traditional medicine. This compound has demonstrated effectiveness against a variety of pathogens, including bacteria, fungi, protozoa, and viruses. It intercalates DNA, targets RNA polymerase, gyrase, and topoisomerase IV, and inhibits cell division. Berberine also inhibits the FtsZ protein, essential for bacterial cell division, and disrupts cell structure [90,91,92,93].
Ungeremine, an isoquinoline alkaloid extracted from Pancratium illyricum L. bulbs, possesses significant antibacterial properties. It enhances DNA cleavage by targeting and inhibiting bacterial topoisomerase IA [94,95].
Quinoline alkaloids like dictamnine, koku-sagine, and maculine, extracted from the stem bark of Teclea afzeli, have shown significant antibacterial properties. These compounds inhibit type II topoisomerase enzymes, DNA replication, and act as respiratory inhibitors [96,97].
Reserpine, an indole alkaloid obtained from Rauwolfia serpentina, is a well-known natural compound with strong EPI activity. When combined with reserpine, various bacterial species, including Staphylococcus, Streptococcus, and Micrococcus, have demonstrated increased antibiotic susceptibility [98,99]. It enhances antibiotic susceptibility in various bacteria by inhibiting efflux pumps and is effective against MDR Acinetobacter baumannii and Stenotrophomonas maltophilia [100,101,102].
Sanguinarine, extracted from specific plants such as Chelidonium majus, Sanguinaria canadensis, and Macleaya cordata, disrupts bacterial membranes, acts as a DNA intercalator, and inhibits replication and transcription. It also shows antimycobacterial activity [103,104,105].
Tomatidine, a steroidal alkaloid found in solanaceous plants like tomatoes, potatoes, and eggplants, boosts the effectiveness of antibiotics against S. aureus and other bacteria, particularly when used with aminoglycosides [106].
Chanoclavine, a tricyclic ergot alkaloid derived from Ipomoea muricata, acts synergistically with tetracycline to combat MDR E. coli by inhibiting ATPase-dependent efflux pumps [107].
Conessine, a steroidal alkaloid derived from Holarrhena antidysenterica barks, is effective against both Gram-positive and Gram-negative bacteria, showing synergistic effects with conventional antibiotics. It acts as an EPI against A. baumannii [108,109,110].
Squalamine, a natural steroid-polyamine compound first isolated from the dogfish shark, is distinct from many other compounds, as it is not primarily plant-derived. It disrupts microbial membranes by interacting with their components, leading to cell death in both Gram-negative and Gram-positive bacteria. These alkaloids not only boost the effectiveness of conventional antibiotics, but also offer potential solutions to combat antibiotic resistance [111].
5.2. Organosulfur Compounds
A significant body of literature discusses the antibacterial and antifungal properties of plant-derived sulfur-containing compounds [82,112,113]. Various compounds, including allicin, ajoene, dialkenyl and dialkyl sulfides, S-allyl cysteine, S-allyl-mercapto cysteine, and isothiocyanates, have demonstrated significant antibacterial properties against a wide range of bacteria, both Gram-positive and Gram-negative [114,115]. Research indicates that plants with high polysulfide content exhibit broad-spectrum antimicrobial properties [116,117].
Allicin (allyl 2-propenethiosulfinate or diallyl thiosulfinate or S-allyl cysteine sulfoxide), derived from garlic (Allium sativum) and other Allium species, has been found to have antimicrobial effects against bacteria such as Staphylococcus epidermidis, Pseudomonas aeruginosa, Streptococcus agalactiae, MRSA, and oral pathogens responsible for periodontitis [118]. Allicin has been shown to boost the effectiveness of antibiotics like cefoperazone, tobramycin, and ciprofloxacin against P. aeruginosa [119]. Its antimicrobial properties arise from the inhibition of sulfhydryl-dependent enzymes, such as alcohol dehydrogenase, thioredoxin reductase, and RNA polymerase [120]. The presence of cysteine and glutathione diminishes allicin’s inhibitory impact, suggesting that these compounds interact with allicin to mitigate microbial damage. Additionally, allicin partially inhibits DNA and protein synthesis and may also affect RNA [120].
Ajoene, another compound from garlic, is composed of the E- and Z-ajoene stereoisomers. It demonstrates broad-spectrum antimicrobial activity, effective against bacteria, fungi, and protozoa, and possesses even stronger antiviral properties compared to allicin [121]. Like allicin, the inhibitory effect of ajoene is diminished in the presence of cysteine, suggesting that both compounds share a mechanism that targets thiol-dependent enzyme systems [122].
Isothiocyanates (ITCs) are volatile sulfur compounds produced from plant glucosinolates and the enzyme myrosinase, found in Brassicaceae plants like cauliflower, cabbage, mustard, and broccoli. ITCs have potent antibacterial effects and are considered promising candidates. For instance, ITCs derived from horseradish (Armoracia rusticana) roots demonstrate potent antimicrobial effects against oral pathogens [123,124]. They act as highly effective bactericides against Helicobacter pylori, functioning by inhibiting urease and reducing inflammation [125]. The antimicrobial action of ITCs is likely attributed to their interaction with proteins, thereby disrupting essential biochemical processes. Their primary mode of action involves targeting sulfhydryl groups by reacting with amines, thiols, and hydroxyls. Additionally, ITCs inhibit bacterial ATP binding sites by targeting cysteine residues [125].
Sulforaphane, present in various plants like Diplotaxis harra, is derived from 4-methyl sulfinyl butyl glucosinolate. It exhibits strong anticarcinogenic and antibacterial properties, particularly against H. pylori, a known risk factor for stomach cancer. Additionally, sulforaphane is effective against S. aureus and L. monocytogenes, highlighting its potential as a natural antibacterial agent [126,127].
Allyl isothiocyanates (AITCs), found in Brassicaceae plants such as Armoracia rusticana and Eutrema japonicum, possess significant antibacterial properties against E. coli and S. aureus, demonstrating both bacteriostatic and bactericidal activities [128]. AITCs lower the minimum inhibitory concentration (MIC) values of erythromycin against S. pyogenes and show synergistic effects with streptomycin against E. coli and P. aeruginosa [129,130]. However, AITCs exhibit low inhibitory effects against certain Gram-positive bacteria [131,132]. Their antimicrobial mechanisms include compromising cell wall integrity and inducing internal structural changes, as observed through electron microscopy [133]. AITCs can additionally deactivate crucial intracellular enzymes by oxidatively cleaving disulfide bonds [134]. They form pores in bacterial cell membranes, causing leakage of intracellular substances [135].
Benzyl isothiocyanate (BITC), found in Alliaria petiolate [136], has demonstrated effectiveness against 15 MRSA isolates, exhibiting bactericidal effects on 11 of them. This suggests BITC’s potential in combating MRSA strains [137]. The antibacterial action of BITC is attributed to its lipophilic and electrophilic properties, which enable it to penetrate bacterial membranes and disrupt their integrity [138].
Phenethyl isothiocyanate (PEITC), present in Brassica vegetables such as Brassica campestris and Brassica rapa [136], exhibits antimicrobial activity primarily against Gram-positive bacteria, with lower efficacy against Gram-negative ones [137,138]. Additionally, PEITC has antifungal properties against Alternaria brassicicola [139], possibly due to mechanisms involving reduced oxygen consumption, intracellular ROS accumulation, and mitochondrial membrane depolarization [140].
Berteroin, a compound present in broccoli (Brassica oleracea L.), exhibits the lowest minimum inhibitory concentration (MIC) values against both extracellular and intracellular bacterial strains, highlighting it as a highly potent bactericidal compound. It also proves effective against H. pylori [141,142].
5.3. Phenolic Compounds
Phenolic compounds encompass a wide array of bioactive natural molecules extensively employed in medical applications. These compounds enhance the effectiveness of antibiotics against resistant pathogens by employing multiple mechanisms [143,144,145]. One notable mechanisms is the reduction of efflux pump (EP) activity, acting as efflux pump inhibitors (EPIs) and demonstrating significant EPI activity against pathogenic bacteria. Table 1 lists important plant-derived EPIs.
Resveratrol, a natural phenolic compound, inhibits the CmeABC efflux pumps of Campylobacter jejuni and the efflux pumps of Mycobacterium smegmatis [146,147]. Ferreira et al. [147] reported that resveratrol increases ethidium bromide accumulation in Arcobacter butzleri and Arcobacter cryaerophilus [148].
Baicalein, a flavone from Thymus vulgaris, Scutellaria baicalensis, and Scutellaria lateriflora, enhances the effectiveness of β-lactam antibiotics, tetracycline, and ciprofloxacin against MRSA by inhibiting the NorA efflux pump [149]. Combining baicalein with tetracycline exhibits synergistic effects against E. coli by inhibiting efflux pumps [150,151].
Biochanin A, an isoflavone, inhibits the efflux system of MRSA by reducing NorA protein expression and shows potent activity against Chlamydia spp. and Mycobacterium strains [152,153,154]. Other flavonoids, such as chrysosplenol-D and chrysoplenetin from Artemisia annua, inhibit the NorA efflux pump in the presence of subinhibitory berberine concentrations [155]. Isoflavonoids and flavonolignans also inhibit NorA, enhancing the potency of norfloxacin and berberine [156].
Silybin, biochanin A, genistein, and orobol augment the activity of S. aureus against NorA substrates [156,157]. The hybridization of antibiotics with flavonoids diminishes efflux pump activity, thereby boosting antibiotic accumulation and efficacy.
Kaempferol has emerged as a promising candidate against various pathogens, effective against fluconazole-resistant C. albicans and MRSA by inhibiting the NorA pump [158,159]. Kaempferol rhamnoside from Persea lingue increases ciprofloxacin’s antimicrobial activity in NorA-overexpressing S. aureus strains [160]. Brown et al. [159] identified efflux pump inhibitors using an LC-MS method, finding that rhamentin and kaempferol significantly inhibit efflux pumps [161].
Quercetin exhibits moderate efflux pump inhibition, more effectively identified with LC-MS methods than fluorescence-based techniques [159].
Chalcones, such as 4′,6′-Dihydroxy-3′,5′-dimethyl-2′-methoxychalcone from Dalea versicolor, inhibit the NorA efflux pump and reduce erythromycin MIC [159]. Holler et al. [161] identified two synthetic chalcones with potent NorA efflux pump inhibitory activity.
Catechin gallates, like epigallocatechin gallate (EGCG), provide health benefits and potent antimicrobial activity against resistant pathogens, including MRSA, by weakly inhibiting the NorA efflux pump [162]. Several phenolic compounds inhibit DNA gyrase, such as novobiocin [163,164], and natural products like green tea polyphenols, chebulinic acid, and anthraquinones exhibit similar activity [165,166,167]. EGCG inhibits the B subunit of DNA gyrase and efflux pumps, making it a significant focus for future research.
Chebulinic acid from Terminalia chebula effectively inhibits quinolone-resistant M. tuberculosis DNA gyrase [163]. Haloemodins, which are semisynthetic anthraquinone derivatives, inhibit DNA gyrase in MRSA and vancomycin-resistant Enterococcus faecium [163].
3-p-trans-Coumaroyl-2-hydroxyquinic acid (CHQA) from Cedrus deodara demonstrates potent antibacterial activity against food-borne pathogens by damaging the cytoplasmic membrane and inducing intracellular leakage [168].
Hydroxycinnamic acids like p-coumaric acid interfere with membrane integrity, displaying significant activity [169].
Flavonoids engage with critical enzymes involved in the production of bacterial cell membrane precursors, including beta-ketoacyl acyl carrier protein synthase (KAS) II and III. Flavanones like naringenin, eriodictyol, and taxifolin inhibit KAS III, exhibiting moderate antibacterial activity [170]. 3,6-Dihydroxyflavone binds to KAS I and III, indicating that many flavonoids can inhibit these enzymes. EGCG covalently binds to and inactivates beta-ketoacyl[acyl carrier protein] reductase (FabG) [171].
Sakuranetin exhibits potent antibacterial activity by inhibiting FabZ in Helicobacter pylori. Quercetin and apigenin also inhibit FabZ, though less effectively [172]. Curcumin damages the cell membranes of S. aureus and E. coli due to its amphipathic and lipophilic structure [173].
Quercetin and apigenin inhibit d-alanine ligase in H. pylori and E. coli, albeit with high MIC values, indicating low inhibitory activity [174].
Sophoraflavanone B directly interacts with peptidoglycan, inhibiting MRSA growth [175]. Other phenolic compounds inhibit enzymes like dihydrofolate reductase, urease, and sortase [176,177,178].
Curcumin inhibits sortase A in S. aureus [179,180]. Morin from Rhus verniciflua inhibits sortase A and B, showing fibrinogen cell-clumping activity [180,181,182]. Flavonoids like 4′,7,8-trihydroxy-2-isoflavene inhibit urease, offering potential as natural inhibitors [168].
Phenolic compounds exhibit diverse mechanisms against bacteria, from efflux pump inhibition and cell membrane interaction to enzyme inhibition. Their significant activities make them promising candidates for future studies and clinical trials. EGCG and curcumin, with their multiple modes of action, exemplify compounds that bacteria cannot easily resist.
5.4. Coumarins
Coumarins are naturally produced by many plants and microorganisms [181,182]. They possess a wide range of bioactivities, including vasodilator, estrogenic, anticoagulant, analgesic, anti-inflammatory, sedative, hypothermic, anti-helminthic, anticancer, antioxidant, and dermal photosensitizing properties [183,184]. Numerous studies have highlighted the antimicrobial activity of both natural and synthetic coumarin derivatives [185,186]. For example, Basile et al. [187] identified several coumarins and pyranocoumarins from Ferulago campestris roots, such as agasyllin, grandivittin, and aegelinol benzoate, which showed antibacterial and antioxidant activities against both Gram-negative and Gram-positive bacteria.
Aegelinol and agasyllin showed significant effectiveness against Salmonella enterica serovar typhi, Enterobacter aerogenes, E. cloacae, and S. aureus, with minimum inhibitory concentrations (MIC) of 16 μg/mL for aegelinol and 32 μg/mL for agasyllin. Additionally, they exhibited dose-dependent activity against Helicobacter pylori at concentrations ranging from 5 to 25 μg/mL [185,188,189].
Tan et al. [186] identified one new and nine previously known prenylated coumarins in extracts from the roots of Prangos hulusii and assessed their antimicrobial efficacy against both standard strains and clinical isolates. The new coumarin, 4′-senecioiloxyosthol, was highly active against Bacillus subtilis (MIC = 5 μg/mL), while osthole was effective against B. subtilis, S. aureus, Klebsiella pneumonia, and methicillin-sensitive Staphylococcus aureus (MSSA) (all MICs = 125 μg/mL).
El-Seedi [190] identified a new aryl coumarin glucoside, asphodelin A 4′-O-β-D-glucoside, and its aglycon, asphodelin A, from Asphodelus microcarpus. Asphodelin A exhibited potent activity against S. aureus, E. coli, P. aeruginosa, C. albicans, and Botrytis cinerea, with MIC values ranging from 4 to 128 μg/mL.
Maxwell [191] investigated the structure-activity relationship (SAR) of coumarins such as clorobiocin, novobiocin, and coumermycin A1, which are derived from different Streptomyces species. His findings indicated that the noviosyl sugar moiety, in conjunction with the coumarin structure, is essential for biological activity. These coumarins are highly effective inhibitors of DNA topoisomerase type II, commonly referred to as DNA gyrase [192].
Structure-activity relationship (SAR) studies have demonstrated that lipophilicity and a planar configuration are crucial for potent antibacterial activity [193]. The antimicrobial efficacy of coumarins is primarily due to passive diffusion, which enhances cellular penetration, especially in Gram-positive bacteria. Sardari et al. [194] suggested that the presence of a free 6-OH group is important for antifungal activity, while a free 7-OH group is vital for antibacterial activity. Further systematic SAR analysis revealed that coumarins possessing a methoxy group at C-7 and a hydroxyl group at C-6 or C-8 exhibit broad-spectrum antibacterial properties. An aromatic dimethoxy configuration boosts effectiveness against certain pathogens, such as Haemophilus influenzae, beta-hemolytic Streptococcus, and Streptococcus pneumoniae [193]. Recent studies also suggest that coumarins can suppress bacterial quorum-sensing networks, affecting biofilm formation and virulence factor production [183,195,196,197,198,199].
Some coumarin derivatives can inhibit EP in MRSA strains. Bergamottin epoxide, a furanocoumarin from grapefruit (Citrus paradisi), reduced the MIC value of norfloxacin against MRSA by 20-fold via EP inhibition [198]. Another study on EP inhibition by coumarins from Mesua ferrea found two compounds that reduced the MIC of norfloxacin by 8-fold in MRSA and clinical S. aureus isolates [200].
Coumarins can bind to isoprene units in plant cells to form more complex structures. 6-Geranyl coumarin and galbanic acid are terpenoid coumarins that significantly inhibit EP in S. aureus. Galbanic acid reduced the MIC of ciprofloxacin by up to 8-fold, with a mode of action comparable to verapamil, a known EP inhibitor [201].
5.5. Terpenes
Terpenes, also known as isoprenoids, constitute the most diverse group of natural compounds and are present in almost every form of life. They serve various functions, ranging from contributing to cell structure (such as cholesterol and steroids in membranes) to facilitating cellular processes (such as retinal in vision, carotenoids in photosynthesis, and quinones in electron transport) [202,203].
Terpenes are plentiful in flowers, fruits, and vegetables, particularly in plant reproductive structures and foliage during and after the flowering phase. They constitute a significant part of herbal resins and are responsible for the distinctive scents of many plants [203]. Numerous terpenes and their derivatives function as protective agents against herbivores and pathogens [204]. Gram-positive bacteria are generally more susceptible to terpenes than Gram-negative ones due to terpenes’ lipophilic properties. Monoterpenes increase membrane fluidity and permeability, alter protein topology, and disrupt the respiration chain [205].
Togashi et al. [206] studied the inhibitory effects of various terpene alcohols on S. aureus, finding that farnesol and nerolidol had strong antibacterial effects with minimum bactericidal concentration (MBC) values of 20 and 40 μg/mL, respectively. They further investigated how these terpene alcohols interact with bacterial cell membranes by evaluating intracellular K+ ion leakage, indicating that potassium leakage serves as an indicator of the antibacterial strength of compounds that disrupt cell membranes. Farnesol and nerolidol were the most effective, with the hydrocarbon chain length between C10 and C12 being optimal for antibacterial and membrane-disrupting activity [207]. Dehydroabietic acid, a resin acid, also shows antibacterial activity against S. aureus [208,209,210,211].
Carvone, particularly (4R)-(−)-carvone, is effective against Campylobacter jejuni, E. faecium, and E. coli, while (4S)-(+)-carvone is active against L. monocytogenes. Both isomers demonstrate efficacy against a range of pathogenic fungi, inhibiting the transformation of C. albicans from yeast to its pathogenic filamentous form [212,213,214].
Thymol shows strong efficacy against Candida albicans, C. glabrata, and C. krusei, both independently and when combined with fluconazole. MIC values are 49.37, 51.25, and 70 μg/mL, respectively. Thymol shows synergistic effects with fluconazole against Candida species and, along with carvacrol, eugenol, and menthol, is effective against various food-decaying fungi, making them good alternatives to synthetic fungicides [215].
Thymol and carvacrol, major components of Thymus capitatus, also exhibit antibacterial effects against E. coli, Enterobacter aerogenes, S. aureus, and P. aeruginosa, with MIC values of 0.005–0.008 mg/mL for thymol and 0.007–0.008 mg/mL for carvacrol [216,217]. These compounds reduce bacterial counts on polypropylene surfaces during the biofilm formation of Salmonella spp. (S. typhimurium, S. enteritidis, and S. saintpaul) and disrupt established biofilms at MIC or 2 × MIC [218]. Thymol’s mechanism against S. typhimurium involves “disruption of membrane integrity”, while thymol and carvacrol inhibit EP in a concentration-dependent manner, enhancing ethidium bromide accumulation in foodborne pathogens [219].
Ursolic acid and α-amyrin, two pentacyclic triterpenes, show broad-spectrum antibacterial activity by disorganizing the E. coli inner membrane model [220]. Eugenol and cinnamaldehyde, found in essential oils, prevent H. pylori growth and exhibit bioactivity against MRSA and MSSA biofilms, disrupting cell-to-cell communication and biofilm construction [221]. Cinnamaldehyde damages bacterial membrane structure, reduces membrane potential, and affects metabolic activity, inhibiting E. coli and S. aureus growth [222,223,224]. Various terpenoid derivatives exhibit antimycobacterial properties against pathogens like M. tuberculosis.
Copp [222] documented that a range of terpenoids, such as sandaracopimaric acid, (+)-totarol, agelasine F, elisapterosin B, costunolide, parthenolide, 1,10-epoxycostunolide, santamarine, reynosin, alantolactone, puupehenone, elatol, deschloroelatol, debromolaurinterol, allolaurinterol, and aureol, are effective against M. tuberculosis. This activity is linked to their lipophilic structures, aiding in the penetration of the mycobacterial cell wall.
5.6. Antimicrobial Peptides from Plants
Plants are abundant sources of biologically active compounds with a wide range of properties, many of which find applications in medicine and agriculture [224,225,226,227]. Among these, antimicrobial peptides (AMPs) stand out. AMPs are small, amphiphilic molecules that generally weigh between 2 and 10 kDa, and typically carry a positive charge at neutral and physiological pH levels [228,229]. Despite significant variations in their primary and secondary structures, most plant-derived AMPs share a compact spatial structure stabilized by intramolecular disulfide bonds, which provide stability against temperature fluctuations, enzymatic activity, and chemical agents [228,230].
Plant AMPs are grouped into various families based on their amino acid sequences, cysteine motifs, disulfide bond locations, and secondary structural elements [228,229]. The main AMP families include defensins, thionins, α-hairpinins (hairpin-like peptides), hevein-like peptides, knottins, snakins, lipid-transfer proteins, and cyclotides. There are also peptides that do not fit into these categories, such as those with atypical cysteine motifs, those without disulfide bonds, cyclic peptides lacking a cysteine knot, and peptides that are rich in glycine, histidine, and alanine [228,230,231,232,233,234,235]. According to the Data Repository of Antimicrobial Peptides (DRAMP), more than 800 plant peptides have been identified so far [236].
Antimicrobial peptides (AMPs) are essential components of plant innate immunity, particularly in defense against biotic stressors [230]. These peptides demonstrate a variety of functions, such as antibacterial, antifungal, insecticidal, and antiviral activities. Additionally, certain AMPs are known to inhibit hydrolases and protein synthesis [237]. Owing to their chemical characteristics, plant AMPs also possess antiproliferative properties [238,239]. These properties make AMPs promising candidates for the development of novel pharmaceuticals or biological agents for plant protection [240].
AMPs are present in all plants, and each species within a specific taxon (such as a genus or family) displays a distinct molecular diversity of peptides from various structural families [241,242,243,244]. They can be extracted from various parts of plants, including vegetative [245] and generative parts [242,246,247,248,249,250], as well as from aboveground [242,248,249] and underground tissues [250,251,252,253]. Seeds, in particular, are a rich source of diverse AMPs [234].
Advancements in transcriptomic and proteomic methods have significantly enhanced plant AMP research [254,255,256]. However, the isolation of AMPs remains crucial for studying their structure–function relationships at the cellular and organism levels [257,258,259]. Classical extraction methods continue to be relevant, especially for isolating new peptides that are not yet classified within existing plant AMP families [229,232,251].
Summarizing the accumulated knowledge and developing comprehensive isolation schemes that allow for the extraction of a wide variety of peptides will provide new opportunities in AMP research.
6. The Utility of Artificial Intelligence (AI)
Numerous pathogenic bacteria have increasingly developed resistance to existing antibiotics, while the pace of new antibiotic development has significantly slowed. This escalating resistance poses a severe threat to global health, underscoring the urgent need for innovative solutions.
Leveraging AI algorithms offers a novel approach to accelerating the drug discovery process. In a recent study, researchers trained a neural network to screen approximately 7500 molecules, resulting in the identification of a compound named abaucin. This compound proved effective in controlling an Acinetobacter baumannii infection [260]. This success highlights AI’s potential to identify promising antibiotic candidates more efficiently than traditional methods.
AI, especially through machine learning and deep learning techniques, is instrumental not only in designing new antibiotics but also in optimizing the effectiveness of existing drugs through synergistic combinations [261]. Machine learning algorithms analyze patterns in antimicrobial resistance (AMR) data, providing critical support to healthcare providers and policymakers by predicting which bacteria and fungi may develop resistance to specific drugs or compounds [185,262]. Furthermore, machine learning models are valuable tools for AMR surveillance. By examining data on antimicrobial usage and the presence of resistant microorganisms, these models allow public health authorities to make well-informed decisions, prepare for, and respond swiftly to resistance outbreaks by identifying emerging resistance patterns and at-risk populations and areas [11]. Collectively, these applications play a crucial role in mitigating the overall impact of AMR [263].
7. Other Strategies to Overcome Antibiotic Resistance
Researchers are actively exploring natural resources to find viable alternatives to traditional antibiotics. Plants, in particular, are considered a valuable source of antimicrobial compounds [264,265]. Despite identifying numerous phytochemicals, many remain unexplored. The primary challenge lies not only in discovering these compounds but also in translating laboratory findings into practical applications within clinical and hospital settings. Bridging the gap between research and real-world healthcare implementation of these natural antimicrobial agents is a significant obstacle that scientists are striving to overcome [266,267].
One promising strategy focuses on increasing the intracellular concentration of antimicrobials within bacterial cells. This can be achieved through potentiation by targeting non-essential bacterial components, such as efflux pump inhibitors [268].
Another approach involves using membrane transporters, like iron transporters, by conjugating antibiotics to an iron-binding siderophore mimetic group, facilitating their entry and enhancing their biological activity [269].
A recent study identified di-berberine conjugates that demonstrate enhanced synergistic effects with aminoglycosides, highlighting their potential as lead compounds in the development of efflux pump inhibitors (EPIs) [270].
8. Future Perspectives and Conclusions
The increasing evidence supporting the efficacy of medicinal plants in treating infectious diseases underscores their potential as sources of novel antimicrobial agents [271]. These natural compounds, especially phytochemicals, exhibit significant antimicrobial activity against various human pathogens. Their benefits include accessibility, cost-effectiveness, and minimal side effects. Additionally, some plant-derived compounds can enhance the effectiveness of traditional antibiotics, offering a promising strategy to combact bacterial resistance [272].
A key approach involves the co-administration of plant compounds with conventional antibiotics, which can reduce minimum inhibitory concentration (MIC) values and produce synergistic effects. This method parallels successful antibiotic combinations like amoxicillin-clavulanate and isoniazid-rifampicin-pyrazinamide-ethambutol, which target different bacterial sites to enhance efficacy and prevent resistance development [273].
Understanding the specific molecular mechanisms of antimicrobial plants is crucial for developing new therapeutic strategies [274]. Proposed mechanisms include disrupting bacterial cell membranes, inhibiting efflux pumps (EPs), and blocking DNA and protein synthesis [275]. For instance, the combination of EGCG and tetracycline has shown synergistic effects by inhibiting bacterial efflux pumps and restoring antibiotic efficacy in resistant strains [276].
The rapid evolution of resistance among bacteria and other microbes poses a significant threat to modern medicine, challenging the effectiveness of existing antimicrobial therapies [277]. The overuse of antibiotics in healthcare and agriculture has exacerbated this issue, leading to the proliferation of multidrug-resistant pathogens. As antibiotic discovery lags behind the pace of resistance development, we risk entering a post-antibiotic era where common infections could become untreatable [278].
Addressing antimicrobial resistance (AMR) requires comprehensive and coordinated global action. This includes implementing stewardship programs to limit inappropriate antibiotic use, enhancing infection control measures, and fostering international cooperation [279]. A One Health approach, integrating human, animal, and environmental health, is essential for curbing resistance transmission and preserving the efficacy of antimicrobials [280].
Future research should focus on identifying and developing more potential treatments from plant secondary metabolites. Combining these natural compounds with existing antibiotics holds promise for effectively treating infections caused by resistant pathogens. With continued innovation and global collaboration, we can mitigate the impact of AMR and safeguard public health.
Funding
This research received no external funding.
Data Availability Statement
No new data were created or analyzed in this study.
Acknowledgments
The author sincerely thanks Giancarlo Angeles Flores (University of Chieti, Italy), Gaia Cusumano, and Roberto Venanzoni (University of Perugia, Italy) for their valuable advice provided during the drafting of the manuscript.
Conflicts of Interest
The author declares no conflicts of interest.
References
- Fleming, A. On the antibacterial action of cultures of a penicillium with special reference to their use in the isolation of B. influenza. Bull. World Health Org. 1929, 79, 780–790. [Google Scholar] [CrossRef]
- Uddin, T.M.; Chakraborty, A.I.; Khusro, A.; Matin Zidan, B.M.R.; Mitra, S.; Emran, T.B.; Dhama, K.; Ripon, M.K.H.; Gajdács, M.; Sahibzada, M.U.K.; et al. Antibiotic resistance in microbes: History, mechanisms, therapeutic strategies and future prospects. J. Infect. Public Health 2021, 14, 1750–1766. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, E.M.; Alhatlani, B.Y.; de Paula Menezes, R.; Martins, C.H.G. Back to Nature: Medicinal Plants as Promising Sources for Antibacterial Drugs in the Post-Antibiotic Era. Plants 2023, 12, 3077. [Google Scholar] [CrossRef] [PubMed]
- Podolsky, S.H. The Evolving Response to Antibiotic Resistance (1945–2018). Palgrave Commun. 2023, 12, 3077. [Google Scholar] [CrossRef]
- Parmar, A.; Lakshminarayanan, R.; Iyer, A.; Mayandi, V.; Leng Goh, E.T.; Lloyd, D.G.; Chalasani, M.L.S.; Verma, N.K.; Prior, S.H.; Beuerman, R.W.; et al. Design and Syntheses of Highly Potent Teixobactin Analogues against Staphylococcus aureus methicillin-resistant Staphylococcus aureus (MRSA), and Vancomycin-Resistant Enterococci (VRE) In Vitro and In Vivo. J. Med. Chem. 2018, 61, 2009–2017. [Google Scholar] [CrossRef]
- Alcock, B.P.; Huynh, W.; Chalil, R.; Smith, K.W.; Raphenya, A.R.; Wlodarski, M.A.; Edalatmand, A.; Petkau, A.; Syed, S.A.; Tsang, K.K.; et al. CARD 2023: Expanded curation support for machine learning, and resistome prediction at the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2023, 51, D690–D699. [Google Scholar] [CrossRef] [PubMed]
- Muteeb, G.; Rehman, M.T.; Shahwan, M.; Aatif, M. Origin of Antibiotics and Antibiotic Resistance, and Their Impacts on Drug Development: A Narrative Review. Pharmaceuticals 2023, 16, 1615. [Google Scholar] [CrossRef] [PubMed]
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Fair, R.J.; Tor, Y. Antibiotics and Bacterial Resistance in the 21st century. Perspect. Medicin. Chem. 2014, 6, 25–64. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.K.; Hussein, S.; Qurbani, K.; Ibrahim, R.H.; Fareeq, A.; Mahmood, K.A.; Mohamed, M.G. Antimicrobial resistance: Impacts, challenges, and future prospects. J. Med. Surg. Public Health 2024, 2, 100081. [Google Scholar] [CrossRef]
- Salam, M.A.; Al-Amin, M.Y.; Salam, M.T.; Pawar, J.S.; Akhter, N.; Rabaan, A.A.; Alqumber, M.A.A. Antimicrobial Resistance: A Growing Serious Threat for Global Public Health. Healthcare 2023, 11, 1946. [Google Scholar] [CrossRef] [PubMed]
- Khameneh, B.; Diab, R.; Ghazvini, K.; Fazly Bazzaz, B.S. Breakthroughs in bacterial resistance mechanisms and the potential ways to combat them. Microb. Pathog. 2016, 95, 32–42. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations; HM Government and Wellcome Trust: London, UK, 2016; Available online: https://amr-review.org/sites/default/files/160518_Final%20paper_with%20cover.pdf (accessed on 15 June 2024).
- Dheda, K.; Mirzayev, F.; Cirillo, D.M.; Udwadia, Z.; Dooley, K.E.; Chang, K.C.; Omar, S.V.; Reuter, A.; Perumal, T.; Horsburgh, C.R., Jr.; et al. Multidrug-resistant tuberculosis. Nat. Rev. Dis. Primers 2024, 10, 22. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.J.L. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 10325, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Dreisbach, A.; Wang, M.; van der Kooi-Pol, M.M.; Reilman, E.; Koedijk, D.G.A.M.; Mars, R.A.T.; Duipmans, J.; Jonkman, M.; Benschop, J.J.; Bonarius, H.P.J.; et al. Tryptic Shaving of Staphylococcus aureus Unveils Immunodominant Epitopes on the Bacterial Cell Surface. J. Proteome Res. 2020, 19, 2997–3010. [Google Scholar] [CrossRef] [PubMed]
- Staley, J.T. The bacterial species dilemma and the genomic–phylogenetic species concept. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 1899–1909. [Google Scholar] [CrossRef] [PubMed]
- Soni, J.; Sinha, S.; Pandey, R. Understanding bacterial pathogenicity: A closer look at the journey of harmful microbes. Front. Microbiol. 2024, 15, 1370818. [Google Scholar] [CrossRef] [PubMed]
- Gidey, K.; Gidey, M.T.; Hailu, B.Y.; Gebreamlak, Z.B.; Niriayo, Y.L. Clinical and economic burden of healthcare-associated infections: A prospective cohort study. PLoS ONE 2023, 18, e0282141. [Google Scholar] [CrossRef] [PubMed]
- Clare, S.; Rowley, S. Implementing the Aseptic Non Touch Technique (ANTT) clinical practice framework for aseptic technique: A pragmatic evaluation using a mixed methods approach in two London hospitals. J. Infect. Prev. 2018, 19, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Tettey, F.; Parupelli, S.K.; Desai, S. A Review of Biomedical Devices: Classification Regulatory Guidelines Human Factors Software as a Medical Device and Cybersecurity. Biomed. Mater. Devices 2024, 2, 316–341. [Google Scholar] [CrossRef]
- Braun, J.; Eckes, S.; Rommens, P.M.; Schmitz, K.; Nickel, D.; Ritz, U. Toxic Effect of Vancomycin on Viability and Functionality of Different Cells Involved in Tissue Regeneration. Antibiotics 2020, 9, 238. [Google Scholar] [CrossRef] [PubMed]
- Walsh, T.R.; Gales, A.C.; Laxminarayan, R.; Dodd, P.C. Antimicrobial Resistance: Addressing a Global Threat to Humanity. PLoS Med. 2023, 20, e1004264. [Google Scholar] [CrossRef] [PubMed]
- Miklasinska-Majdanik, M.; Kepa, M.; Wojtyczka, R.D.; Idzil, D. Phenolic Compounds Diminish Antibiotic Resistance of Staphylococcus aureus Clinical Strains. Int. J. Environ. Res. Public Health 2018, 15, 2321. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Ahmad, R.; Zeyaullah, M.; Khare, S.K. Microbial Nano-Factories: Synthesis and Biomedical Applications. Front. Chem. 2021, 9, 626834. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.K.; Low, L.Y.; Yap, P.S.X.; Yusoff, K.; Mai, C.W.; Lai, K.S.; Lim, S.H.E. Plant-Derived Antimicrobials: Insights into Mitigation of Antimicrobial Resistance. Rec. Nat. Prod. 2018, 12, 295–396. [Google Scholar] [CrossRef]
- Macheboeuf, P.; Di Guilmi, A.M.; Vernet, T.; Dideberg, O.; Dessen, A. Active site restructuring regulates ligand recognition in class A penicillin-binding proteins. Proc. Natl. Acad. Sci. USA 2005, 102, 577–582. [Google Scholar] [CrossRef] [PubMed]
- Sacco, E.; Cortes, M.; Josseaume, N.; Rice, L.B.; Mainardi, J.L.; Arthur, M. Serine/threonine protein phosphatase-mediated control of the peptidoglycan cross-linking LD-transpeptidase pathway in Enterococcus faecium. MBio 2014, 5, e01446. [Google Scholar] [CrossRef] [PubMed]
- Morais-Cabral, J.H.; Jackson, A.P.; Smith, C.V.; Shikotra, A.; Maxwell, A.; Liddington, R.C. Crystal structure of the breakage-reunion domain of DNA gyrase. Nature 1997, 388, 903–906. [Google Scholar] [CrossRef] [PubMed]
- Willmott, C.J.; Maxwell, A. A single point mutation in the DNA gyrase A protein greatly reduces binding of fluoroquinolones to the gyrase-DNA complex. Antimicrob. Agents Chemother. 1993, 37, 126–127. [Google Scholar] [CrossRef] [PubMed]
- Grise-Miron, L.; Brakier-Gingras, L. Effect of neomycin and protein S1 on the binding of streptomycin to the ribosome. Eur. J. Biochem. 1982, 123, 643–646. [Google Scholar] [CrossRef] [PubMed]
- Moazed, D.; Noller, H.F. Interaction of antibiotics with functional sites in 16S ribosomal RNA. Nature 1987, 327, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Ding, Q.; Ji, Z.; Liang, X.; Deng, C.C.; Wong, C.; Yi, L.; Zhang, S.; Xie, S.; Alvarez, L.M.; et al. Protein cysteine phosphorylation of SarA/MgrA family transcriptional regulators mediates bacterial virulence and antibiotic resistance. Proc. Natl. Acad. Sci. USA 2012, 109, 15461–15466. [Google Scholar] [CrossRef] [PubMed]
- Goswami, M.; Subramanian, R.; Kumar, J.; Jass, J.; Jawali, N. Involvement of antibiotic efflux machinery in glutathione-mediated decreased ciprofloxacin activity in Escherichia coli. Antimicrob. Agents Chemother. 2016, 60, 4369–4374. [Google Scholar] [CrossRef] [PubMed]
- Kohler, T.; Michea-Hamzehpour, M.; Plesiat, P.L.; Kahr, A.; Pechere, J.C. Differential selection of multidrug efflux systems by quinolones in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 1997, 41, 2540–2543. [Google Scholar] [CrossRef] [PubMed]
- Masuda, N.; Sakagawa, E.; Ohya, S.; Gotoh, H.; Tsujimoto, N.; Nishino, T. Substrate specificities of MexAB-OprM MexCD-OprJ and MexXY-OprM efflux pumps in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2000, 44, 3322–3327. [Google Scholar] [CrossRef] [PubMed]
- Morita, Y.; Tomida, J.; Kawamura, Y. MexXY multidrug efflux system of Pseudomonas aeruginosa. Front. Microbiol. 2012, 3, 408. [Google Scholar] [CrossRef] [PubMed]
- Santos, S.; Costa, M.; Viveiros, A.E.; Rosato, J.; Melo-Cristino, J.; Couto, I. Impact of efflux in the development of multidrug resistance phenotypes in Staphylococcus aureus. BMC Microbiol. 2015, 15, 232. [Google Scholar]
- Blair, J.M.A.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J.V. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef] [PubMed]
- De, E.; Basle, A.; Jaquinod, M.; Saint, N.; Mallea, G.; Molle, G.; Pages, J.M. A new mechanism of antibiotic resistance in Enterobacteriaceae induced by a structural modification of the major porin. Mol. Microbiol. 2001, 41, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Gayet, S.; Chollet, R.; Molle, G.; Pages, J.M.; Chevalier, J. Modification of outer membrane protein profile and evidence suggesting an active drug pump in Enterobacter aerogenes clinical strains. Antimicrob. Agents Chemother. 2003, 47, 1555–1559. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Melano, R.; Corso, A.; Petroni, D.; Centron, B.; Orman, A.; Pereyra, N.; Moreno, M.; Galas, M. Multiple antibiotic-resistance mechanisms including a novel combination of extended-spectrum betalactamases in a Klebsiella pneumoniae clinical strain isolated in Argentina. J. Antimicrob. Chemother. 2003, 52, 36–42. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gheorghiu, R.; Yuan, M.; Hall, L.M.; Livermore, D.M. Bases of variation in resistance to beta lactams in Klebsiella oxytoca isolates hyperproducing K1 beta-lactamase. J. Antimicrob. Chemother. 1997, 40, 533–541. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wu, P.J.; Shannon, K.; Phillips, I. Mechanisms of hyperproduction of TEM-1 beta-lactamase by clinical isolates of Escherichia coli. J. Antimicrob. Chemother. 1995, 36, 927–939. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, J.V.; Bershtein, S.; Li, A.; Lozovsky, E.R.; Hartl, D.L.; Shakhnovich, E.I. Biophysical principles predict fitness landscapes of drug resistance. Proc. Natl. Acad. Sci. USA 2016, 113, 11. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Rubin, E.J.; Bifani, P.; Mathys, V.; Lim, M.; Au, J.; Jang, J.; Nam, T.; Dick, J.R.; Walker, K.; et al. Para-Aminosalicylic acid is a prodrug targeting dihydrofolate reductase in Mycobacterium tuberculosis. J. Biol. Chem. 2013, 288, 23447–23456. [Google Scholar] [CrossRef] [PubMed]
- Frere, J.M.; Joris, B. Penicillin-sensitive enzymes in peptidoglycan biosynthesis. Crit. Rev. Microbiol. 1985, 11, 299–396. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, P.E. Structure, biochemistry and mechanism of action of glycopeptide antibiotics. Eur. J. Clin. Microbiol. Infect. Dis. 1989, 8, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Arthur, M.; Molinas, C.; Depardieu, F.; Courvalin, P. Characterization of Tn1546 a Tn3-related transposon conferring glycopeptide resistance by synthesis of depsipeptide peptidoglycan precursors in Enterococcus faecium BM4147. J. Bacteriol. 1993, 175, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Meziane-Cherif, D.; Stogios, P.J.; Evdokimova, E.; Egorova, O.; Savchenko, A.; Courvalin, P. Structural and functional adaptation of vancomycin resistance VanT Serine racemases. MBio 2015, 6, e00806. [Google Scholar] [CrossRef] [PubMed]
- Bugg, T.D.; Wright, G.D.; Dutka-Malen, S.; Arthur, M.; Courvalin, P.; Walsh, C.T. Molecular basis for vancomycin resistance in Enterococcus faecium BM4147: Biosynthesis of a depsipeptide peptidoglycan precursor by vancomycin resistance proteins VanH and VanA. Biochemistry 1991, 30, 10408–10415. [Google Scholar] [CrossRef] [PubMed]
- Bakal, S.N.; Bereswill, S.; Heimesaat, M.M. Finding novel antibiotic substances from medicinal plants—Antimicrobial properties of Nigella sativa directed against multidrug-resistant bacteria. Eur. J. Microbiol. Immunol. 2017, 7, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Sasidharan, S.; Chen, Y.; Saravanan, D.; Sundram, L.Y.; Yoga, L. Extraction isolation and characterization of bioactive compounds from plants’ extracts. Afr. J. Tradit. Complement. Altern. Med. 2011, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Valli, M.; Pivatto, M.; Danuello, A.; Castro-Gamboa, I.; Silva, D.H.S.; Cavalheiro, A.J.; Araújo, Â.R.; Furlan, M.; Lopes, M.N.; Bolzani, V.D.S. Tropical biodiversity: Has it been a potential source of secondary metabolites useful for medicinal chemistry? Trop. Biodivers. 2012, 35, 2278–2287. [Google Scholar] [CrossRef]
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests f or Bacteria That Grow Aerobically; Approved Standard—Ninth Edition. CLSI document M07-A9; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012. [Google Scholar]
- Edeoga, H.O.; Okwu, D.E.; Mbaebie, B.O. Phytochemical constituents of some Nigerian medicinal plants. Afr. J. Biotechnol. 2005, 4, 685–688. [Google Scholar] [CrossRef]
- Parvin, S.; Kader, M.A.; Chouduri, A.U.; Rafshanjani, M.A.S.; Haque, M.E. Antibacterial antifungal and insecticidal activities of the n-hexane and ethyl-acetate fractions of methanolic extract of the leaves of Calotropis gigantea Linn. J. Pharmacogn. Phytochem. 2014, 2, 47–51. [Google Scholar]
- Rahman, M.S.; Anwar, M.N. Antimicrobial Activity of Crude Extract Obtained from the Root of Plumbago zeylanica. Bangladesh J. Microbiol. 2007, 24, 73–75. [Google Scholar] [CrossRef]
- Batool, K.; Sultana, S.; Akhtar, N.; Muhammad, H.; Naheed, A. Medicinal plants combating against human pathogens: A review. Int. J. Biotechnol. Food Sci. 2018, 6, 42–51. [Google Scholar]
- Chuah, E.; Zakaria, Z.; Suhaili, Z.; Abu Bakar Jamaludin, S.; Mohd Desa, M. Antimicrobial activities of plant extracts against methicillin-susceptible and methicillin-resistant Staphylococcus aureus. J. Microbiol. Res. 2014, 4, 6–13. [Google Scholar]
- Sharifi-Rad, J.; Mnayer, D.; Roointan, A.; Shahri, F.; Ayatollahi, S.A.M.; Sharifi-Rad, M.; Molaee, N. Antibacterial activities of essential oils from Iranian medicinal plants on extended-spectrum β-lactamase-producing Escherichia coli. Cell. Mol. Biol. 2016, 62, 75–82. [Google Scholar] [PubMed]
- Pacheco, A.G.; Alcântara, A.F.C.; Abreu, V.G.C.; Corrêa, G.M. Relationships Between Chemical Structure and Activity of Triterpenes against Gram-Positive and Gram-Negative Bacteria. In A Search for Antibact Agents; Bobbarala, V., Ed.; InTech: Rijeka, Croatia, 2012; Chapter 1; pp. 1–25. [Google Scholar]
- Niranjan, P.S.; Kaushal, C.; Jain, S.K. Pharmacological Investigation of Leaves of Polypodium decumanum for Antidiabetic Activity. J. Drug Deliv. Ther. 2017, 7, 2685–2688. [Google Scholar] [CrossRef]
- Manso, T.; Lores, M.; de Miguel, T. Antimicrobial Activity of Polyphenols and Natural Polyphenolic Extracts on Clinical Isolates. Antibiotics 2021, 11, 46. [Google Scholar] [CrossRef] [PubMed]
- Sissi, S.; Di Giacomo, S.; Ferrante, C.; Angelini, P.; Macone, A.; Giusti, A.M.; Toniolo, C.; Vitalone, A.; Abdellah, A.; Larhsini, M.; et al. Characterization of the Phytochemical Composition and Bioactivities of Anacyclus maroccanus Ball. and Anacyclus radiatus Loisel Aerial Parts: Preliminary Evidence for the Possible Development of Moroccan Plants. Molecules 2022, 27, 692. [Google Scholar] [CrossRef]
- Di Simone, S.C.; Angeles Flores, G.; Acquaviva, A.; Nilofar, M.; Libero, M.L.; Venanzoni, R.; Tirillini, B.; Orlando, G.; Zengin, G.; Lai, F.; et al. Phytochemical and biological properties of the water extract from roots and leaves of Lactuca longidentata an endemic phytoalimurgic (food) species of Central Sardinia (Italy). Plant Biosyst. 2023, 157, 594–604. [Google Scholar] [CrossRef]
- Serventi, L.; Flores, G.A.; Cusumano, G.; Barbaro, D.; Tirillini, B.; Venanzoni, R.; Angelini, P.; Acquaviva, A.; Di Simone, S.C.; Orlando, G.; et al. Comparative Investigation of Antimicrobial and Antioxidant Effects of the Extracts from the Inflorescences and Leaves of the Cannabis sativa L. cv. strawberry. Antioxidants 2023, 12, 219. [Google Scholar] [CrossRef] [PubMed]
- Orlando, G.; Adorisio, S.; Delfino, D.; Chiavaroli, A.; Brunetti, L.; Recinella, L.; Leone, S.; D’Antonio, M.; Zengin, G.; Acquaviva, A.; et al. Comparative Investigation of Composition Antifungal and Anti-Inflammatory Effects of the Essential Oil from Three Industrial Hemp Varieties from Italian Cultivation. Antibiotics 2021, 10, 334. [Google Scholar] [CrossRef] [PubMed]
- Kauffmann, A.C.; Castro, V.S. Phenolic Compounds in Bacterial Inactivation: A Perspective from Brazil. Antibiotics 2023, 12, 645. [Google Scholar] [CrossRef] [PubMed]
- Gechev, T.S.; Van Breusegem, F.; Stone, J.M.; Denev, I.; Laloi, C. Natural products from resurrection plants: Potential for medical applications. Biotechnol. Adv. 2014, 32, 1091–1101. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Kim, H.K.; van Wezel, G.P.; Choi, Y.H. Expanding the chemical space for natural products by Aspergillus–Streptomyces co-cultivation and biotransformation. Sci. Rep. 2015, 5, 10868. [Google Scholar] [CrossRef] [PubMed]
- Youssef, S.; Hassan, M.; Ahmed, S. Lichens: Chemistry and biological activities. In Studies in Natural Products Chemistry; Atta-ur-Rahman, Ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 223–259. [Google Scholar]
- Deshmukh, S.K.; Verekar, S.A.; Bhave, S.V. Endophytic fungi: A reservoir of antibacterials. Front. Microbiol. 2015, 5, 715. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.B.; Cheung, R.C.F.; Wong, J.H.; Wang, Y.; Ip, D.T.M.; Wan, D.C.C.; Xia, J.; Chan, W.Y.; Fang, E.F. Antibacterial products of marine organisms. Appl. Microbiol. Biotechnol. 2015, 99, 4145–4173. [Google Scholar] [CrossRef] [PubMed]
- Kiuru, P.; D’Auria, M.V.; Muller, C.D.; Tammela, P.; Vuorela, H.; Yli-Kauhaluoma, J. Exploring marine resources for bioactive compounds. Planta Med. 2014, 80, 1234–1246. [Google Scholar] [CrossRef] [PubMed]
- Mondol, M.A.M.; Shin, H.J.; Rahman, M.A. Antibacterial and antiyeast compounds from marine-derived bacteria. Mar. Drugs 2014, 12, 2913–2921. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M.S.; Glaser, K.B.; Cuevas, C.; Jacobs, R.S.; Kem, W.; Little, R.D.; McIntosh, J.M.; Newman, D.J.; Potts, B.C.; Shuster, D.E. Marine pharmacology in 2009–2011: Marine compounds with antibacterial antidiabetic antifungal anti-inflammatory antiprotozoal antituberculosis and antiviral activities; affecting the immune and nervous systems and other miscellaneous mechanisms of action. Mar. Drugs 2013, 11, 2510–2573. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Singh, A.; Arya, M.; Maheshwari, A.; Bhadouria, R.; Pandey, S.; Kharwar, R.N.; Stal, L.J.; Dubey, N.K. Antimicrobial compounds from seaweeds-associated bacteria and fungi. Appl. Microbiol. Biotechnol. 2015, 99, 1571–1586. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.A.; Park, H.C.; Park, S.W.; Park, S.C.; Seo, M.G.; Her, M.; Kang, J. Synergism of the Combination of Traditional Antibiotics and Novel Phenolic Compounds against Escherichia coli. Pathogens 2020, 9, 811. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Zhang, Z.; Guo, L. Antibacterial Molecules from Marine Microorganisms against Aquatic Pathogens: A Concise Review. Mar. Drugs 2022, 20, 230. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Zha, J.; Koffas, M.A.G. Microbial production of bioactive chemicals for human health. Curr. Opin. Food Sci. 2020, 32, 9–16. [Google Scholar] [CrossRef]
- Barbieri, R.; Coppo, E.; Marchese, A.; Daglia, M.; Sobarzo-Sanchez, E.; Nabavi, S.F.; Nabavi, S.M. Phytochemicals for human disease: An update on plant-derived compounds antibacterial activity. Microbiol. Res. 2017, 196, 44–68. [Google Scholar] [CrossRef] [PubMed]
- Bribi, N. Pharmacological activity of alkaloids: A review. Asian J. Bot. 2018, 1, 1–6. [Google Scholar]
- Khameneh, B.; Iranshahi, M.; Soheili, V.; Bazzaz, B.S.F. Review on plant antimicrobials: A mechanistic viewpoint. Antimicrob. Resist. Infect. Control 2019, 8, 118. [Google Scholar] [CrossRef] [PubMed]
- Jadimurthy, R.; Jagadish, S.; Nayak, S.C.; Kumar, S.; Mohan, C.D.; Rangappa, K.S. Phytochemicals as Invaluable Sources of Potent Antimicrobial Agents to Combat Antibiotic Resistance. Life 2023, 13, 948. [Google Scholar] [CrossRef] [PubMed]
- Otmann, L.; Sleiman, A.; Abdel-Massih, R.M. Antimicrobial Activity of Polyphenols and Alkaloids in Middle Eastern Plants. Front. Microbiol. 2019, 10, 911. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Li, X.; Zhang, C.; Lv, L.; Gao, B.; Li, M. Research Progress on Antibacterial Activities and Mechanisms of Natural Alkaloids: A Review. Antibiotics 2021, 10, 318. [Google Scholar] [CrossRef] [PubMed]
- Stojanovic-Radic, Z.; Pejcic, M.; Dimitrijevic, M.; Aleksic, A.; Anil Kumar, V.N.; Salehi, B.; Cho, C.W.; Sharifi-Rad, J. Piperine-A major principle of black pepper: A review of its bioactivity and studies. Appl. Sci. 2019, 9, 4270. [Google Scholar] [CrossRef]
- Zahin, M.; Bokhari, N.A.; Ahmad, I.; Husain, F.M.; Althubiani, A.S.; Alruways, M.W.; Perveen, K.; Shalawi, M. Antioxidant antibacterial and antimutagenic activity of Piper nigrum seeds extracts. Saudi J. Biol. Sci. 2021, 28, 5094–5105. [Google Scholar] [CrossRef] [PubMed]
- Yi, Z.B.; Yan, Y.; Liang, Y.Z.; Bao, Z. Evaluation of the antimicrobial mode of berberine by LC/ESI-MS combined with principal component analysis. J. Pharm. Biomed. Anal. 2007, 44, 301–304. [Google Scholar] [CrossRef] [PubMed]
- Domadia, P.N.; Bhunia, A.; Sivaraman, J.; Swarup, S.; Dasgupta, D. Berberine targets assembly of Escherichia coli cell division protein FtsZ. Biochemistry 2008, 47, 3225–3234. [Google Scholar] [CrossRef] [PubMed]
- Boberek, J.M.; Stach, J.; Good, L. Genetic evidence for inhibition of bacterial division protein FtsZ by berberine. PLoS ONE 2010, 5, e13745. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, M.; Jannat, K.; Nissapatorn, V.; Rahmatullah, M.; Paul, A.K.; de Lourdes Pereira, M.; Rajagopal, M.; Suleiman, M.; Butler, M.S.; Break, M.K.B.; et al. Antibacterial and Antifungal Alkaloids from Asian Angiosperms: Distribution, Mechanisms of Action, Structure-Activity, and Clinical Potentials. Antibiotics 2022, 11, 1146. [Google Scholar] [CrossRef] [PubMed]
- Casu, L.; Cottiglia, F.; Leonti, M.; De Logu, A.; Agus, E.; Tse-Dinh, Y.C.; Lombardo, V.; Sissi, C. Ungeremine effectively targets mammalian as well as bacterial type I and type II topoisomerases. Bioorg. Med. Chem. Lett. 2011, 23, 7041–7044. [Google Scholar] [CrossRef] [PubMed]
- Romeo, L.; Iori, R.; Rollin, P.; Bramanti, P.; Mazzon, E. Isothiocyanates: An Overview of Their Antimicrobial Activity against Human Infections. Molecules 2018, 23, 624. [Google Scholar] [CrossRef] [PubMed]
- Kuete, V.; Wansi, J.D.; Mbaveng, A.T.; Kana Sop, M.M.; Tadjong, A.T.; Beng, V.P. Antimicrobial activity of the methanolic extract and compounds from Teclea afzelii (Rutaceae). S. Afr. J. Bot. 2008, 74, 572–576. [Google Scholar] [CrossRef]
- Heeb, S.; Fletcher, M.P.; Chhabra, S.R.; Diggle, S.P.; Williams, P.; Camara, M. Quinolones: From antibiotics to autoinducers. FEMS Microbiol. Rev. 2011, 35, 247–274. [Google Scholar] [CrossRef] [PubMed]
- Parai, D.; Banerjee, M.; Dey, P.; Mukherjee, S.M. Reserpine attenuates biofilm formation and virulence of Staphylococcus aureus. Microb. Pathog. 2020, 138, 103790. [Google Scholar] [CrossRef] [PubMed]
- Abdelfatah, S.A.; Efferth, T. Cytotoxicity of the indole alkaloid reserpine from Rauwolfia serpentina against drug-resistant tumor cells. Phytomedicine 2015, 22, 308–318. [Google Scholar] [CrossRef] [PubMed]
- Sridevi, D.; Shankar, C.; Prakash, P.; Park, J.H.; Thamaraiselvi, K. Inhibitory effects of reserpine against efflux pump activity of antibiotic resistance bacteria. Chem. Biol. Lett. 2017, 4, 69–72. [Google Scholar]
- Jia, W.; Li, C.; Zhang, H.; Li, G.; Liu, X.; Wei, J. Prevalence of genes of OXA-23 carbapenemase and AdeABC efflux pump associated with multidrug resistance of Acinetobacter baumannii isolates in the ICU of a comprehensive hospital of northwestern China. Int. J. Environ. Res. Public Health 2015, 12, 10079–10092. [Google Scholar] [CrossRef] [PubMed]
- Obiang-Obounou, B.W.; Kang, O.H.; Choi, J.G.; Keum, J.H.; Kim, S.B.; Mun, S.H.; Shin, D.W.; Kim, K.W.; Park, C.B.; Kim, Y.G.; et al. The mechanism of action of sanguinarine against methicillin-resistant Staphylococcus aureus. J. Toxicol. Sci. 2011, 36, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Al-Ani, I.; Zimmermann, S.; Reichling, J.; Wink, M. Pharmacological synergism of bee venom and melittin with antibiotics and plant secondary metabolites against multi-drug resistant microbial pathogens. Phytomedicine 2015, 22, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Newton, S.M.; Lau, C.; Gurcha, S.S.; Besra, G.S.; Wright, C.W. The evaluation of fortythree plant species for in vitro antimycobacterial activities; isolation of active constituents from Psoralea corylifolia and Sanguinaria canadensis. J. Ethnopharmacol. 2002, 79, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, G.; Lafrance, M.; Boulanger, S.; Séguin, D.L.; Guay, I.; Gattuso, M.; Marsault, E.; Bouarab, K.; Malouin, F. Tomatidine acts in synergy with aminoglycoside antibiotics against multiresistant Staphylococcus aureus and prevents virulence gene expression. J. Antimicrob. Chemother. 2012, 67, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Langlois, J.P.; Millette, G.; Guay, I.; Dubé-Duquette, A.; Chamberland, S.; Jacques, P.É.; Rodrigue, S.; Bouarab, K.; Marsault, É.; Malouin, F. Bactericidal Activity of the Bacterial ATP Synthase Inhibitor Tomatidine and the Combination of Tomatidine and Aminoglycoside against Persistent and Virulent Forms of Staphylococcus aureus. Front. Microbiol. 2020, 11, 805. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, G.R.; Maurya, A.; Yadav, D.K.; Singh, V.; Khan, F.; Gupta, M.K.; Singh, M.; Darokar, M.P.; Srivastava, S.K. Synergy of clavine alkaloid “chanoclavine” with tetracycline against multi-drug-resistant E. coli. J. Biomol. Struct. Dyn. 2019, 37, 1307–1325. [Google Scholar] [CrossRef] [PubMed]
- Siriyong, T.; Srimanote, P.; Chusri, S.; Yingyongnarongkul, B.E.; Suaisom, C.; Tipmanee, V.; Voravuthikunchai, S.P. Conessine as a novel inhibitor of multidrug efflux pump systems in Pseudomonas aeruginosa. BMC Complement. Altern. Med. 2017, 17, 405. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Singh, B.; Bhandari, P.; Gupta, A.P.; Kaul, V.K. Steroidal alkaloids from Holarrhena antidysenterica (L.) WALL. Chem. Pharm. Bull. 2007, 55, 912–914. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.N.; Ge, X.L.; Dong, T.T.; Gao, H.Y.; Sun, B.H. Antibacterial steroidal alkaloids from Holarrhena antidysenterica. Chin. J. Nat. Med. 2017, 15, 540–545. [Google Scholar] [PubMed]
- Alhanout, K.; Malesinki, S.; Vidal, N.; Peyrot, V.; Rolain, J.M.; Brunel, J.M. New insights into the antibacterial mechanism of action of squalamine. J. Antimicrob. Chemother. 2010, 65, 1688–1693. [Google Scholar] [CrossRef] [PubMed]
- Miekus, N.; Marszałek, K.; Podlacha, M.; Iqbal, A.; Puchalski, C.; Swiergiel, H. Health Benefits of Plant-Derived Sulfur Compounds, Glucosinolates, and Organosulfur Compounds. Molecules 2020, 25, 3804. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.R.; Shafaei, A.; Balmer, L.; Lewis, J.R.; Hodgson, J.M.; Millar, A.H.; Blekkenhorst, L.C. Sulfur compounds: From plants to humans and their role in chronic disease prevention. Crit. Rev. Food Sci. Nutr. 2022, 63, 8616–8638. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.V.; Pant, S.; Khan, M.A.H.; Shah, A.A.; Siddiqui, S.; Jeridi, M.; Alhamdi, H.W.S.; Ahmad, S. Phytochemicals as Antimicrobials: Prospecting Himalayan Medicinal Plants as Source of Alternate Medicine to Combat Antimicrobial Resistance. Pharmaceuticals 2023, 16, 881. [Google Scholar] [CrossRef] [PubMed]
- Boghrati, Z.; Iranshahi, M. Ferula species: A rich source of antimicrobial compounds. J. Herb. Med. 2019, 16, 100244. [Google Scholar] [CrossRef]
- Reiter, J.; Levina, N.; van der Linden, M.; Gruhlke, M.; Martin, C.; Slusarenko, A.J. Diallylthiosulfinate (Allicin), a Volatile Antimicrobial from Garlic (Allium sativum), Kills Human Lung Pathogenic Bacteria, Including MDR Strains, as a Vapor. Molecules 2017, 22, 1711. [Google Scholar] [CrossRef] [PubMed]
- Bhatwalkar, S.B.; Mondal, R.; Krishna, S.B.N.; Adam, J.K.; Govender, P.; Anupam, R. Antibacterial Properties of Organosulfur Compounds of Garlic (Allium sativum). Front. Microbiol. 2021, 12, 613077. [Google Scholar] [CrossRef] [PubMed]
- Lanzotti, V.; Scala, F.; Bonanomi, G. Compounds from allium species with cytotoxic and antimicrobial activity. Phytochem. Rev. 2014, 13, 769–791. [Google Scholar] [CrossRef]
- Davis, S.R. An overview of the antifungal properties of allicin and its breakdown products—The possibility of a safe and effective antifungal prophylactic. Mycoses 2005, 48, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Sen, D.; Bhattacharjee, C. Inhibition mechanism study for diallyl thiosulfinate (allicin) against crucial bacterial proteins through in silico molecular docking simulation. Process Biochem. 2022, 122, 110–119. [Google Scholar] [CrossRef]
- Rehman, F.; Mairaj, S. Antimicrobial studies of allicin and ajoene. Int. J. Pharm. Bio Sci. 2013, 4, 1095–1105. [Google Scholar]
- Nguyen, N.M.; Gonda, S.; Vasas, G. A Review on the Phytochemical Composition and Potential Medicinal Uses of Horseradish (Armoracia rusticana) Root. Food Rev. Int. 2013, 29, 261–275. [Google Scholar] [CrossRef]
- Kim, H.Y.; Pha-a-god, S.; Shin, I.S. Antibacterial Activities of Isothiocyanates Extracted from Horseradish (Armoracia rusticana) Root against Antibiotic-resistant Bacteria. Food Sci. Biotechnol. 2015, 24, 1029–1034. [Google Scholar] [CrossRef]
- Dufour, V.; Stahl, M.; Baysse, C. The antibacterial properties of isothiocyanates. Microbiology 2015, 161, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Breier, A.; Zieoelhofpt, A. “Lysine is the Lord”, thought some scientists in regard to the group interacting with fluorescein isothiocyanate in ATP-binding sites of P-type ATPases but, is it not cysteine? Gen. Physiol. Biophys. 2000, 19, 253–263. [Google Scholar] [PubMed]
- Benzekri, R.; Bouslama, L.; Papetti, A.; Snoussi, M.; Benslimene, I.; Hamami, M.; Limam, F. Isolation and identification of an antibacterial compound from Diplotaxis harra (Forssk.) Boiss. Ind. Crop. Prod. 2016, 80, 228–234. [Google Scholar] [CrossRef]
- Palaniappan, K.; Holley, R.A. Use of natural antimicrobials to increase antibiotic susceptibility of drug resistant bacteria. Int. J. Food Microbiol. 2010, 140, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Dockery, C.R.; Crosby, M.; Chavarria, K.; Patterson, B.; Giedd, M. Antibacterial activities of wasabi against Escherichia coli O157 and Staphylococcus aureus. Front. Microbiol. 2016, 7, 1403. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Massih, R.M.; Debs, E.; Othman, L.; Attieh, J.; Cabrerizo, F.M. Glucosinolates, a natural chemical arsenal: More to tell than the myrosinase story. Front. Microbiol. 2023, 14, 1130208. [Google Scholar] [CrossRef] [PubMed]
- Nowicki, D.; Maciąg-Dorszyńska, M.; Bogucka, K.; Szalewska-Pałasz, A.; Herman-Antosiewicz, A. Various modes of action of dietary phytochemicals, sulforaphane and phenethyl isothiocyanate, on pathogenic bacteria. Sci. Rep. 2019, 9, 13677. [Google Scholar] [CrossRef] [PubMed]
- Luciano, F.B.; Holley, R.A. Enzymatic inhibition by allyl isothiocyanate and factors affecting its antimicrobial action against Escherichia coli O157. Int. J. Food Microbiol. 2009, 131, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Gao, H. Isothiocyanates in Food. In Handbook of Dietary Phytochemicals; Xiao, J., Sarker, S.D., Asakawa, Y., Eds.; Springer: Singapore, 2021. [Google Scholar]
- Takahashi, H.; Nakamura, A.; Fujino, N.; Sawaguchi, Y.; Sato, M.; Kuda, T.; Kimura, B. Evaluation of the antibacterial activity of allyl isothiocyanate, clove oil, eugenol and carvacrol against spoilage lactic acid bacteria. LWT 2021, 145, 111263. [Google Scholar] [CrossRef]
- Cipollini, D.; Cipollini, K. A review of garlic mustard (Alliaria petiolata, Brassicaceae) as an allelopathic plant. J. Torrey Bot. Soc. 2016, 143, 339–348. [Google Scholar] [CrossRef]
- Dias, C.; Aires, A.; Saavedra, M.J. Antimicrobial activity of isothiocyanates from cruciferous plants against methicillin-resistant Staphylococcus aureus (MRSA). Int. J. Mol. Sci. 2014, 15, 19552–19561. [Google Scholar] [CrossRef] [PubMed]
- Sofrata, A.; Santangelo, E.M.; Azeem, M.; Borg-Karlson, A.K.; Gustafsson, A.; Putsep, K. Benzyl isothiocyanate, a major component from the roots of Salvadora persica is highly active against gram-negative bacteria. PLoS ONE 2011, 6, e23045. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Hong, E.; Kim, G.H. Evaluation of antibacterial activity of 3-butenyl, 4-pentenyl, 2-phenylethyl, and benzyl isothiocyanate in Brassica vegetables. J. Food Sci. 2010, 75, M412–M416. [Google Scholar] [CrossRef] [PubMed]
- Saladino, F.; Bordin, K.; Luciano, F.B.; Franzón, M.F.; Mañes, J.; Meca, G. Antimicrobial Activity of the Glucosinolates. In Glucosinolates. Reference Series in Phytochemistry; Mérillon, J.M., Ramawat, K., Eds.; Springer: Cham, Switzerland, 2017. [Google Scholar] [CrossRef]
- Yu, H.; Jia, W.; Zhao, M.; Li, L.; Liu, J.; Chen, J.; Pan, H.; Zhang, X. Antifungal mechanism of isothiocyanates against Cochliobolus heterostrophus. Pest Manag. Sci. 2022, 12, 5133–5141. [Google Scholar] [CrossRef] [PubMed]
- Calmes, B.; N’Guyen, G.; Dumur, J.; Brisach, C.A.; Campion, C.; Iacomi, B.; Pigné, S.; Dias, E.; Macherel, D.; Guillemette, T.; et al. Glucosinolate-derived isothiocyanates impact mitochondrial function in fungal cells and elicit an oxidative stress response necessary for growth recovery. Front. Plant Sci. 2015, 6, 414. [Google Scholar] [CrossRef]
- Moon, J.K.; Kim, J.R.; Ahn, Y.J.; Shibamoto, T. Analysis and anti-helicobacter activity of sulforaphane and related compounds present in broccoli (Brassica oleracea L.) sprouts. J. Agric. Food Chem. 2010, 58, 6672–6677. [Google Scholar] [CrossRef] [PubMed]
- Sathianarayanan, S.; Ammanath, A.V.; Biswas, R.; Anita, B.; Sukumaran, S.; Venkidasamy, B. A new approach against Helicobacter pylory using plants and its constituents: A review study. Microb. Pathog. 2022, 168, 105594. [Google Scholar] [CrossRef] [PubMed]
- Farhadi, F.; Khameneh, B.; Iranshahi, M.; Iranshahy, M. Antibacterial activity of flavonoids and their structure–activity relationship: An update review. Phytother. Res. 2019, 33, 13–40. [Google Scholar] [CrossRef] [PubMed]
- Hrytsyk, R.A.; Kutsyk, R.V.; Yurchyshyn, O.I.; Struk, O.A.; Kireev, I.V.; Grytsyk, A.R. The investigation of antimicrobial and antifungal activity of some Artemisia L. species. Pharmacia 2021, 68, 93–100. [Google Scholar] [CrossRef]
- Górniak, I.; Bartoszewski, R.; Króliczewski, J. Comprehensive review of antimicrobial activities of plant flavonoids. Phytochem. Rev. 2019, 18, 241–272. [Google Scholar] [CrossRef]
- Klancnik, A.; Sikic Pogacar, M.; Trost, K.; Tusek Znidaric, M.; Mozetic Vodopivec, B.; Smole, M.S. Anti-campylobacter activity of resveratrol and an extract from waste pinot noir grape skins and seeds, and resistance of camp. Jejuni planktonic and biofilm cells, mediated via the CmeABC efflux pump. J. Appl. Microbiol. 2017, 122, 65–77. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, S.; Silva, F.; Queiroz, J.A.; Oleastro, M.; Domingues, F.C. Resveratrol against Arcobacter butzleri and Arcobacter cryaerophilus: Activity and effect on cellular functions. Int. J. Food Microbiol. 2014, 180, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Lechner, D.; Gibbons, S.; Bucar, F. Plant phenolic compounds as ethidium bromide efflux inhibitors in mycobacterium smegmatis. J. Antimicrob. Chemother. 2008, 62, 345–348. [Google Scholar] [CrossRef]
- Chan, B.C.; Ip, M.; Lau, C.B.; Lui, S.L.; Jolivalt, C.; Ganem-Elbaz, C.; Litaudon, M.; Reiner, N.E.; Gong, H.; See, R.H.; et al. Synergistic effects of baicalein with ciprofloxacin against NorA over-expressed methicillin-resistant Staphylococcus aureus (MRSA) and inhibition of MRSA pyruvate kinase. J. Ethnopharmacol. 2011, 137, 767–773. [Google Scholar] [CrossRef]
- Fujita, M.; Shiota, S.; Kuroda, T.; Hatano, T.; Yoshida, T.; Mizushima, T.; Tsuchiya, T. Remarkable synergies between baicalein and tetracycline, and baicalein and beta-lactams against methicillin-resistant Staphylococcus aureus. Microbiol. Immunol. 2005, 49, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Zou, D.; Xie, K.; Wang, H.; Chen, Y.; Xie, M. Inhibitory effects of biochanin a on the efflux pump of methicillin-resistant Staphylococcus aureus (MRSA). Wei Sheng Wu Xue Bao 2014, 54, 1204–1211. [Google Scholar] [PubMed]
- Hanski, L.; Genina, N.; Uvell, H.; Malinovskaja, K.; Gylfe, Å.; Laaksonen, T.; Kolakovic, R.; Mäkilä, E.; Salonen, J.; Hirvonen, J.; et al. Inhibitory activity of the isoflavone biochanin A on intracellular bacteria of genus Chlamydia and initial development of a buccal formulation. PLoS ONE 2014, 9, e115115. [Google Scholar] [CrossRef] [PubMed]
- Cannalire, R.; Machado, D.; Felicetti, T.; Costa, S.S.; Massari, S.; Manfroni, G.; Barreca, M.L.; Tabarrini, O.; Couto, I.; Viveiros, M.; et al. Natural isoflavone biochanin a as a template for the design of new and potent 3-phenylquinolone efflux inhibitors against Mycobacterium avium. Eur. J. Med. Chem. 2017, 140, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Stermitz, F.R.; Cashman, K.K.; Halligan, K.M.; Morel, C.; Tegos, G.P.; Lewis, K. Polyacylated neohesperidosides from Geranium caespitosum: Bacterial multidrug resistance pump inhibitors. Bioorg. Med. Chem. Lett. 2003, 13, 1915–1918. [Google Scholar] [CrossRef] [PubMed]
- Morel, C.; Stermitz, F.R.; Tegos, G.; Lewis, K. Isoflavones as potentiators of antibacterial activity. J. Agric. Food Chem. 2003, 51, 5677–5679. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.R.; Ettefagh, K.A.; Todd, D.A.; Cole, P.S.; Egan, J.M.; Foil, D.H.; Lacey, E.P.; Cech, N.B. Bacterial efflux inhibitors are widely distributed in land plants. J. Ethnopharmacol. 2021, 267, 113533. [Google Scholar] [CrossRef]
- Randhawa, H.K.; Hundal, K.K.; Ahirrao, P.N.; Jachak, S.M.; Nandanwar, H.S. Efflux pump inhibitory activity of flavonoids isolated from Alpinia calcarata against methicillin-resistant Staphylococcus aureus. Biologia 2016, 71, 484–493. [Google Scholar] [CrossRef]
- Shao, J.; Zhang, M.; Wang, T.; Li, Y.; Wang, C. The roles of CDR1, CDR2, and MDR1 in kaempferol-induced suppression with fluconazole-resistant Candida albicans. Pharm. Biol. 2016, 54, 984–992. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.R.; Ettefagh, K.A.; Todd, D.; Cole, P.S.; Egan, J.M.; Foil, D.H.; Graf, T.N.; Schindler, B.D.; Kaatz, G.W.; Cech, N.B. A mass spectrometry-based assay for improved quantitative measurements of efflux pump inhibition. PLoS ONE 2015, 10, e0124814. [Google Scholar] [CrossRef] [PubMed]
- Holler, J.G.; Christensen, S.B.; Slotved, H.C.; Rasmussen, H.B.; Gúzman, A.; Olsen, C.E.; Petersen, B.; Mølgaard, P. Novel inhibitory activity of the Staphylococcus aureus NorA efflux pump by a kaempferol rhamnoside isolated from Persea lingue Nees. J. Antimicrob. Chemother. 2012, 67, 1138–1144. [Google Scholar] [CrossRef] [PubMed]
- Holler, J.G.; Slotved, H.C.; Mølgaard, P.; Olsen, C.E.; Christensen, S.B. Chalcone inhibitors of the NorA efflux pump in Staphylococcus aureus whole cells and enriched everted membrane vesicles. Bioorg. Med. Chem. 2012, 20, 4514–4521. [Google Scholar] [CrossRef] [PubMed]
- Singhal, D.; Saxena, S. Catechin gallate a promising resistance modifying candidate to potentiate β-lactam antibiotics to overcome resistance in Staphylococcus aureus. Curr. Med. Res. Pract. 2017, 7, 224–228. [Google Scholar] [CrossRef]
- Anderle, C.; Stieger, M.; Burrell, M.; Reinelt, S.; Maxwell, A.; Page, M.; Heide, L. Biological activities of novel gyrase inhibitors of the aminocoumarin class. Antimicrob. Agents Chemother. 2008, 52, 1982–1990. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.; Tyagi, C.; Goyal, S.; Jamal, S.; Wahi, D.; Jain, R.; Bharadvaja, N.; Grover, A. Identification of chebulinic acid as potent natural inhibitor of M. tuberculosis DNA gyrase and molecular insights into its binding mode of action. Comput. Biol. Chem. 2015, 59, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Duan, F.; Li, X.; Cai, S.; Xin, G.; Wang, Y.; Du, D.; He, S.; Huang, B.; Guo, X.; Zhao, H.; et al. Haloemodin as novel antibacterial agent inhibiting DNA gyrase and bacterial topoisomerase I. J. Med. Chem. 2014, 57, 3707–3714. [Google Scholar] [CrossRef]
- Wu, Y.; Bai, J.; Zhong, K.; Huang, Y.; Qi, H.; Jiang, Y.; Gao, H. Antibacterial Activity and Membrane-Disruptive Mechanism of 3-p-trans-Coumaroyl-2-hydroxyquinic Acid, a Novel Phenolic Compound from Pine Needles of Cedrus deodara, against Staphylococcus aureus. Molecules 2016, 21, 1084. [Google Scholar] [CrossRef] [PubMed]
- Campos, F.M.; Couto, J.A.; Figueiredo, A.R.; Toth, I.V.; Rangel, A.O.; Hogg, T.A. Cell membrane damage induced by phenolic acids on wine lactic acid bacteria. Int. J. Food Microbiol. 2009, 135, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Dikdik, K.; Eti, A.; Cut, S.; Mieke, H.S. Antibacterial Flavonoids against Oral Bacteria of Enterococcus Faecalis ATCC 29212 from Sarang Semut (Myrmecodia pendans) and Its Inhibitor Activity against Enzyme MurA. Curr. Drug Discov. Technol. 2019, 16, 290–296. [Google Scholar]
- Li, B.H.; Zhang, R.; Du, Y.T.; Sun, Y.H.; Tian, W.X. Inactivation mechanism of the beta-ketoacyl-[acyl carrier protein] reductase of bacterial type-II fatty acid synthase by epigallocatechin gallate. Biochem. Cell Biol. 2006, 84, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Kong, Y.; Wu, D.; Zhang, H.; Wu, J.; Chen, J.; Ding, J.; Hu, L.; Jiang, H.; Shen, X. Three flavonoids targeting the beta-hydroxyacyl-acyl carrier protein dehydratase from Helicobacter pylori: Crystal structure characterization with enzymatic inhibition assay. Protein Sci. 2008, 17, 1971–1978. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, P.; Singh, M.; Kumari, H.; Kumari, A.; Mukhopadhyay, K. Bactericidal activity of curcumin I is associated with damaging of bacterial membrane. PLoS ONE 2015, 10, e0121313. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Kong, Y.; Han, C.; Chen, J.; Hu, L.; Jiang, H.; Shen, X. D-Alanine ligase as a new target for the flavonoids quercetin and apigenin. Int. J. Antimicrob. Agents 2008, 32, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Mun, S.H.; Joung, D.K.; Kim, S.B.; Park, S.J.; Seo, Y.S.; Gong, R.; Choi, J.G.; Shin, D.W.; Rho, J.R.; Kang, O.H.; et al. The mechanism of antimicrobial activity of sophoraflavanone B against methicillin-resistant Staphylococcus aureus. Foodborne Pathog. Dis. 2014, 11, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Maresso, A.W.; Schneewind, O. Sortase as a target of anti-infective therapy. Pharmacol. Rev. 2008, 60, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.P.; Peng, Z.Y.; Dong, J.J.; He, J.; Ouyang, H.; Feng, Y.T.; Lu, C.L.; Lin, W.Q.; Wang, J.X.; Xiang, Y.P.; et al. Synthesis, structure-activity relationship analysis and kinetics study of reductive derivatives of flavonoids as Helicobacter pylori urease inhibitors. Eur. J. Med. Chem. 2013, 63, 685–695. [Google Scholar] [CrossRef] [PubMed]
- Vidigal, P.G.; Müsken, M.; Becker, K.A.; Häussler, S.; Wingender, J.; Steinmann, E.; Kehrmann, J.; Gulbins, E.; Buer, J.; Rath, P.M.; et al. Effects of Green Tea Compound Epigallocatechin-3-Gallate against Stenotrophomonas maltophilia Infection and Biofilm. PLoS ONE 2014, 9, e92876. [Google Scholar] [CrossRef] [PubMed]
- Budhathoki, R.; Timilsina, A.P.; Regmi, B.P.; Sharma, K.R.; Aryal, N.; Parajuli, N. Metabolome Mining of Curcuma longa L. Using HPLC-MS/MS and Molecular Networking. Metabolites 2023, 13, 898. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wang, X.; Sun, L.; Gao, Y.; Niu, X.; Wang, H. Novel Inhibitor Discovery of Staphylococcus aureus Sortase B and the Mechanism Confirmation via Molecular Modeling. Molecules 2018, 23, 977. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.I.; Syed, Q.A.; Khattak, M.N.K.; Hafez, B.; Reigosa, M.J.; El-Keblawy, A. Natural product coumarins: Biological and pharmacological perspectives. Biologia 2019, 74, 863–888. [Google Scholar] [CrossRef]
- Tsivileva, O.M.; Koftin, O.V. Fungal coumarins: Biotechnological and pharmaceutical aspects. Stud. Nat. Prod. Chem. 2023, 78, 441–479. [Google Scholar]
- Ismael, R.N.; Mustafa, Y.F.; Al-Qazaz, H.K. Coumarin-based products: Their biodiversity and pharmacology. Iraq J. Pharm. 2021, 18, 162–179. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, H.-S.; Cheng, M.; Xia, P.; Qian, K.; Wu, P.-C.; Lai, C.-Y.; Xia, Y.; Yang, Z.-Y.; Morris-Natschke, S.L.; et al. Antitumor agents 292. Design, synthesis and pharmacological study of S- and O-substituted 7-mercapto- or hydroxy-coumarins and chromones as potent cytotoxic agents. Eur. J. Med. Chem. 2012, 49, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.; Agrawal, R.; Ubaidullah, M.; Hassan, M.I.; Tarannum, N. Design, synthesis and validation of anti-microbial coumarin derivatives: An efficient green approach. Heliyon 2019, 5, e02615. [Google Scholar] [CrossRef] [PubMed]
- Cheke, R.S.; Patel, H.M.; Patil, V.M.; Ansari, I.A.; Ambhore, J.P.; Shinde, S.D.; Kadri, A.; Snoussi, M.; Adnan, M.; Kharkar, P.S.; et al. Molecular Insights into Coumarin Analogues as Antimicrobial Agents: Recent Developments in Drug Discovery. Antibiotics 2022, 11, 566. [Google Scholar] [CrossRef] [PubMed]
- Vaou, N.; Stavropoulou, E.; Voidarou, C.; Tsigalou, C.; Bezirtzoglou, E. Towards Advances in Medicinal Plant Antimicrobial Activity: A Review Study on Challenges and Future Perspectives. Microorganisms 2021, 9, 2041. [Google Scholar] [CrossRef] [PubMed]
- Tan, N.; Yazıcı-Tütüniş, S.; Bilgin, M.; Tan, E.; Miski, M. Antibacterial Activities of Pyrenylated Coumarins from the Roots of Prangos hulusii. Molecules 2017, 22, 1098. [Google Scholar] [CrossRef] [PubMed]
- El-Seedi, H.R. Antimicrobial arylcoumarins from Asphodelus microcarpus. J. Nat. Prod. 2007, 70, 118–120. [Google Scholar] [CrossRef] [PubMed]
- Basile, A.; Sorbo, S.; Spadaro, V.; Bruno, M.; Maggio, A.; Faraone, N.; Rosselli, S. Antimicrobial and antioxidant activities of coumarins from the roots of Ferulago campestris (Apiaceae). Molecules 2009, 14, 939–952. [Google Scholar] [CrossRef] [PubMed]
- Spencer, A.C.; Panda, S.S. DNA Gyrase as a Target for Quinolones. Biomedicines 2023, 11, 371. [Google Scholar] [CrossRef] [PubMed]
- Leyn, S.A.; Kent, J.E.; Zlamal, J.E.; Elane, M.L.; Vercruysse, M.; Osterman, A.L. Two classes of DNA gyrase inhibitors elicit distinct evolutionary trajectories toward resistance in gram-negative pathogens. NPJ Antimicrob. Resist. 2024, 2, 5. [Google Scholar] [CrossRef]
- Maxwell, A. The interaction between coumarin drugs and DNA gyrase. Mol. Microbiol. 1993, 9, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Wen, Y.; Liang, C.G.; Liu, J.; Ding, Y.B.; Zhang, W.H. Design, Synthesis and Antifungal Activity of Psoralen Derivatives. Molecules 2017, 22, 1672. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sass, A.; Van Acker, H.; Wille, J.; Verhasselt, B.; Van Nieuwerburgh, F.; Kaever, V.; Crabbé, A.; Coenye, T. Coumarin reduces virulence and biofilm formation in Pseudomonas aeruginosa by affecting quorum sensing, type III secretion and c-di-GMP levels. Front. Microbiol. 2018, 9, 1952. [Google Scholar]
- Reen, F.J.; Gutiérrez-Barranquero, J.A.; Parages, M.L. Coumarin: A novel player in microbial quorum sensing and biofilm formation inhibition. Appl. Microbiol. Biotechnol. 2018, 102, 2063–2073. [Google Scholar] [CrossRef]
- Sardari, S.; Mori, Y.; Horita, K.; Micetich, R.G.; Nishibe, S.; Daneshtalab, M. Synthesis and antifungal activity of coumarins and angular furanocoumarins. Bioorg Med Chem. 1999, 7, 1933–1940. [Google Scholar] [CrossRef] [PubMed]
- da SM Forezi, L.; Froes, T.Q.; Cardoso, M.F.; de Oliveira Maciel, C.A.; Nicastro, G.G.; Baldini, R.L.; Costa, D.; Ferreira, V.F.; Castilho, M.S.; de C da Silva, F. Synthesis and biological evaluation of Coumarins derivatives as potential inhibitors of the production of Pseudomonas aeruginosa virulence factor Pyocyanin. Curr. Top. Med. Chem. 2018, 18, 149–156. [Google Scholar]
- Mikulášová, M.; Chovanová, R.; Vaverková, Š. Synergism between antibiotics and plant extracts or essential oils with efflux pump inhibitory activity in coping with multidrug-resistant staphylococci. Phytochem. Rev. 2016, 15, 651–662. [Google Scholar] [CrossRef]
- Araújo-Neto, J.B.d.; Oliveira-Tintino, C.D.d.M.; de Araújo, G.A.; Alves, D.S.; Ribeiro, F.R.; Brancaglion, G.A.; Carvalho, D.T.; Lima, C.M.G.; Mohammed Ali, H.S.H.; Rather, I.A.; et al. 3-Substituted Coumarins Inhibit NorA and MepA Efflux Pumps of Staphylococcus aureus. Antibiotics 2023, 12, 1739. [Google Scholar] [CrossRef] [PubMed]
- de Araújo, R.S.A.; Barbosa-Filho, J.M.; Scotti, M.T.; Scotti, L.; Cruz, R.M.D.D.; Falcão-Silva, V.D.S.; Siqueira-Júnior, J.P.D.; Mendonça-Junior, F.J.B. Modulation of drug resistance in Staphylococcus aureus with Coumarin derivatives. Scientifica 2016, 2016, 6894758. [Google Scholar] [CrossRef] [PubMed]
- Oldfield, E.; Lin, F.Y. Terpene biosynthesis: Modularity rules. Angew. Chem. Int. Ed. 2012, 51, 1124–1137. [Google Scholar] [CrossRef] [PubMed]
- Masyita, A.; Sari, R.M.; Astuti, A.D.; Yasir, B.; Rumata, N.R.; Emran, T.B.; Nainu, F.; Simal-Gandara, J. Terpenes and terpenoids as main bioactive compounds of essential oils, their roles in human health and potential application as natural food presenvatives. Food Chem. 2022, 13, 100217. [Google Scholar] [CrossRef] [PubMed]
- Toffolatti, S.L.; Maddalena, G.; Passera, A.; Casati, P.; Bianco, P.A.; Quaglino, F. Role of terpenes in plant defense to biotic stress. In Biocontrol Agents and Secondary Metabolites; Sudisha, J., Ed.; Woodhead Publishing: Sawston, UK, 2021; pp. 401–417. [Google Scholar]
- Khameneh, B.; Eskin, N.A.M.; Iranshahi, M.; Fazly Bazzaz, B.S. Phytochemicals: A Promising Weapon in the Arsenal against Antibiotic-Resistant Bacteria. Antibiotics 2021, 10, 1044. [Google Scholar] [CrossRef] [PubMed]
- Togashi, N.; Hamashima, H.; Shiraishi, A.; Inoue, Y.; Takano, A. Antibacterial activities against Staphylococcus aureus of terpene alcohols with aliphatic carbon chains. J. Essent. Oil Res. 2010, 22, 263–269. [Google Scholar] [CrossRef]
- Ephrem, E.; Najjar, A.; Charcosset, C.; Greige-Gerges, H. Selection of nerolidol among a series of terpenic and phenolic compounds for its potent activity against Lactobacillus fermentum ATCC 9338. Process Biochem. 2019, 80, 146–156. [Google Scholar] [CrossRef]
- El Fannassi, Y.; Gharsallaoui, A.; Khelissa, S.; El Amrani, M.A.; Suisse, I.; Sauthier, M.; Jama, C.; Boudra, S.; Chihib, N.E. Complexation of Terpenes for the Production of New Antimicrobial and Antibiofilm Molecules and Their Encapsulation in Order to Improve Their Activities. Appl. Sci. 2023, 13, 9854. [Google Scholar] [CrossRef]
- Catteau, L.; Zhu, L.; Van Bambeke, F.; Quetin-Leclercq, J. Natural and hemi-synthetic pentacyclic triterpenes as antimicrobials and resistance modifying agents against Staphylococcus aureus: A review. Phytochem. Rev. 2018, 17, 1129–1163. [Google Scholar] [CrossRef]
- Shakeri, A.; Akhtari, J.; Soheili, V.; Taghizadeh, S.F.; Sahebkar, A.; Shaddel, R.; Asili, J. Identification and biological activity of the volatile compounds of Glycyrrhiza triphylla Fisch. & C.A. Mey. Microb. Pathog. 2017, 109, 39–44. [Google Scholar] [PubMed]
- Tretyakova, E.V.; Salimova, E.V.; Parfenova, L.V. Synthesis and Antimicrobial and Antifungal Activity of Resin Acid Acetylene Derivatives. Russ. J. Bioorg. Chem. 2019, 45, 545–551. [Google Scholar] [CrossRef]
- Bouyahya, A.; Mechchate, H.; Benali, T.; Ghchime, R.; Charfi, S.; Balahbib, A.; Burkov, P.; Shariati, M.A.; Lorenzo, J.M.; Omari, N.E. Health Benefits and Pharmacological Properties of Carvone. Biomolecules 2021, 11, 1803. [Google Scholar] [CrossRef] [PubMed]
- Sharifzadeh, A.; Khosravi, A.R.; Shokri, H.; Shirzadi, H. Potential effect of 2-isopropyl-5-methylphenol (thymol) alone and in combination with fluconazole against clinical isolates of Candida albicans, C. glabrata and C. krusei. J. Mycol. Med. 2018, 28, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Cox-Georgian, D.; Ramadoss, N.; Dona, C.; Basu, C. Therapeutic and Medicinal Uses of Terpenes. In Medicinal Plants; Springer: Berlin/Heidelberg, Germany, 2019; pp. 333–359. [Google Scholar]
- Ben Miri, Y.; Nouasri, A.; Herrera, M.; Djenane, D.; Ariño, A. Antifungal Activity of Menthol, Eugenol and Their Combination against Aspergillus ochraceus and Aspergillus niger In Vitro and in Stored Cereals. Foods 2023, 12, 2108. [Google Scholar] [CrossRef] [PubMed]
- Marinelli, L.; Di Stefano, A.; Cacciatore, I. Carvacrol and its derivatives as antibacterial agents. Phytochem. Rev. 2018, 17, 903–921. [Google Scholar] [CrossRef]
- Althunibat, O.Y.; Qaralleh, H.; Ahmed Al-Dalin, S.Y.; Abboud, M.; Khleifat, K.; Majali, I.S.; Aldal’in, H.K.; Rayyan, W.A.; Jaafraa, A. Effect of thymol and carvacrol, the major components of Thymus capitatus on the growth of Pseudomonas aeruginosa. J. Pure Appl. Microbiol. 2016, 10, 367–374. [Google Scholar]
- Amaral, V.C.S.; Santos, P.R.; da Silva, A.F.; dos Santos, A.R.; Machinski, M.; Mikcha, J.M.G. Effect of carvacrol and thymol on Salmonella spp. biofilms on polypropylene. Int. J. Food Sci. Technol. 2015, 50, 2639–2643. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Luo, J.; Jie, J.; Deng, X.; Song, L. The Herbal Compound Thymol Targets Multiple Salmonella Typhimurium Virulence Factors for Lon Protease Degradation. Front. Pharmacol. 2021, 12, 674955. [Google Scholar] [CrossRef] [PubMed]
- Broniatowski, M.; Mastalerz, P.; Flasiński, M. Studies of the interactions of ursane-type bioactive terpenes with the model of Escherichia coli inner membrane—Langmuir monolayer approach. Biochim. Biophys. Acta Biomembr. 2015, 1848, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Abdelhamid, S.M.; Edris, A.E.; Sadek, Z. Novel approach for the inhibition of Helicobacter pylori contamination in yogurt using selected probiotics combined with eugenol and cinnamaldehyde nanoemulsions. Food Chem. 2023, 417, 135877. [Google Scholar] [CrossRef] [PubMed]
- Yadav, M.K.; Chae, S.W.; Im, G.J.; Chung, J.W.; Song, J.J. Eugenol: A phyto-compound effective against methicillin-resistant and methicillin-sensitive Staphylococcus aureus clinical strain biofilms. PLoS ONE 2015, 10, e0119564. [Google Scholar] [CrossRef] [PubMed]
- Rathinam, P.; Vijay Kumar, H.S.; Viswanathan, P. Eugenol exhibits anti-virulence properties by competitively binding to quorum sensing receptors. Biofouling 2017, 33, 624–639. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.B.; Liu, X.Y.; Jiang, P.P.; Li, W.D.; Wang, Y.F. Mechanism and antibacterial activity of cinnamaldehyde against Escherichia coli and Staphylococcus aureus. Mod. Food Sci. Technol. 2015, 31, 31–35. [Google Scholar]
- Copp, B.R. Anti-mycobacterial natural products and mechanisms of action. Nat. Prod. Rep. 2022, 39, 77–89. [Google Scholar]
- Godlewska, K.; Ronga, D.; Michalak, I. Plant extracts—Importance in sustainable agriculture. Ital. J. Agron. 2021, 16, 1851. [Google Scholar] [CrossRef]
- Kulaeva, O.; Kliukova, M.; Afonin, A.; Sulima, A.; Zhukov, V.; Tikhonovich, I. The role of plant antimicrobial peptides (AMPs) in response to biotic and abiotic environmental factors. Bio. Comm. 2020, 65, 187–199. [Google Scholar] [CrossRef]
- Chen, N.; Jiang, C. Antimicrobial peptides: Structure, mechanism, and modification. Eur. J. Med. Chem. 2023, 255, 115377. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Z.; Yang, P.; Lei, J.; Zhao, J. Biological Function of Antimicrobial Peptides on Suppressing Pathogens and Improving Host Immunity. Antibiotics 2023, 12, 1037. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos-Silva, C.A.; Zupin, L.; Oliveira-Lima, M.; Vilela, L.M.B.; Bezerra-Neto, J.P.; Ferreira-Neto, J.R.; Ferreira, J.D.C.; de Oliveira-Silva, R.L.; Pires, C.d.J.; Aburjaile, F.F.; et al. Plant Antimicrobial Peptides: State of the Art, In Silico Prediction and Perspectives in the Omics Era. Bioinform. Biol. Insights 2020, 14, 1177932220952739. [Google Scholar] [CrossRef] [PubMed]
- Slavokhotova, A.A.; Rogozhin, E.A. Defense Peptides From the α-Hairpinin Family Are Components of Plant Innate Immunity. Front. Plant Sci. 2020, 11, 465. [Google Scholar] [CrossRef] [PubMed]
- Ryazantsev, D.Y.; Rogozhin, E.A.; Dimitrieva, T.V.; Drobyazina, P.E.; Khadeeva, N.V.; Egorov, T.A.; Grishin, E.V.; Zavriev, S.K. A novel hairpin-like antimicrobial peptide from barnyard grass (Echinochloa crusgalli L.) seeds: Structure–functional and molecular-genetics characterization. Biochimie 2014, 99, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Lima, A.M.; Azevedo, M.I.G.; Sousa, L.M.; Oliveira, N.S.; Andrade, C.R.; Freitas, C.D.T.; Souza, P.F.N. Plant antimicrobial peptides: An overview about classification, toxicity and clinical applications. Int. J. Biol. Macromol. 2022, 214, 10–21. [Google Scholar] [CrossRef] [PubMed]
- de Souza Cândido, E.; Silva Cardoso, M.H.; Sousa, D.A.; Viana, J.C.; Gomes de Oliveira-Júnior, N.; Miranda, V.; Luiz Franco, O. The use of versatile plant antimicrobial peptides in agribusiness and human health. Peptides 2014, 55, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Elshafie, H.S.; Camele, I.; Mohamed, A.A. A Comprehensive Review on the Biological, Agricultural and Pharmaceutical Properties of Secondary Metabolites Based-Plant Origin. Int. J. Mol. Sci. 2023, 24, 3266. [Google Scholar] [CrossRef] [PubMed]
- Panwar, S.; Thapliyal, M.; Kuriyal, V.; Tripathi, V.; Thapliyal, A. GEU-AMP50: Enhanced antimicrobial peptide prediction using a machine learning approach. Mater. Today Proc. 2023, 73, 81–87. [Google Scholar] [CrossRef]
- Barashkova, A.S.; Rogozhin, E.A. Isolation of antimicrobial peptides from different plant sources: Does a general extraction method exist? Plant Methods 2020, 16, 143. [Google Scholar] [CrossRef] [PubMed]
- Bakare, O.O.; Gokul, A.; Fadaka, A.O.; Wu, R.; Niekerk, L.A.; Barker, A.M.; Keyster, M.; Klein, A. Plant Antimicrobial Peptides (PAMPs): Features, Applications, Production, Expression, and Challenges. Molecules 2022, 27, 3703. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Liu, S.; Zhang, C. Antimicrobial peptides with antiviral and anticancer properties and their modification and nanodelivery systems. Curr. Res. Biotechnol. 2023, 5, 100121. [Google Scholar] [CrossRef]
- Büyükkiraz, E.M.; Kesmen, Z. Antimicrobial peptides (AMPs): A promising class of antimicrobial compounds. J. Appl. Microbiol. 2022, 132, 1573–1596. [Google Scholar] [CrossRef] [PubMed]
- Luong, H.X.; Thanh, T.T.; Tran, T.H. Antimicrobial peptides—Advances in development of therapeutic applications. Life Sci. 2020, 260, 118407. [Google Scholar] [CrossRef] [PubMed]
- Koehbach, J.; Craik, D.J. The Vast Structural Diversity of Antimicrobial Peptides. Trends Pharmacol. Sci. 2019, 40, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Bin Hafeez, A.; Jiang, X.; Bergen, P.J.; Zhu, Y. Antimicrobial Peptides: An Update on Classifications and Databases. Int. J. Mol. Sci. 2021, 22, 11691. [Google Scholar] [CrossRef]
- Barashkova, A.S.; Sadykova, V.S.; Salo, V.A.; Zavriev, S.K.; Rogozhin, E.A. Nigellothionins from Black Cumin (Nigella sativa L.) Seeds Demonstrate Strong Antifungal and Cytotoxic Activity. Antibiotics 2021, 10, 166. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Culebras, P.V.; Gandía, M.; Garrigues, S.; Marcos, J.F.; Manzanares, P. Antifungal Peptides and Proteins to Control Toxigenic Fungi and Mycotoxin Biosynthesis. Int. J. Mol. Sci. 2021, 22, 13261. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Li, W.; Fang, C.; Zheng, X.; Liu, C.; Huang, Q. Extraction and identification of new flavonoid compounds in dandelion Taraxacum mongolicum Hand.-Mazz. with evaluation of antioxidant activities. Sci. Rep. 2023, 13, 2166. [Google Scholar] [CrossRef] [PubMed]
- Odintsova, T.I.; Korostyleva, T.V.; Odintsova, M.S.; Pukhalsky, V.A.; Grishin, E.V.; Egorov, T.A. Analysis of Triticum boeoticum and Triticum urartu seed defensins: To the problem of the origin of polyploid wheat genomes. Biochimie 2008, 90, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Chan, L.Y.; Gilding, E.K.; Henriques, S.T.; Condon, N.D.; Ravipati, A.S.; Kaas, Q.; Huang, Y.H.; Craik, D.J. Discovery and mechanistic studies of cytotoxic cyclotides from the medicinal herb Hybanthus enneaspermus. J. Biol. Chem. 2020, 295, 10911–10925. [Google Scholar] [CrossRef] [PubMed]
- Slazak, B.; Haugmo, T.; Badyra, B.; Göransson, U. The life cycle of cyclotides: Biosynthesis and turnover in plant cells. Plant Cell Rep. 2020, 3, 1359–1367. [Google Scholar] [CrossRef] [PubMed]
- Shalovylo, Y.I.; Yusypovych, Y.M.; Hrunyk, N.I.; Roman, I.I.; Zaika, V.K.; Krynytskyy, H.T.; Nesmelova, I.V.; Kovaleva, V.A. Seed-derived defensins from Scots pine: Structural and functional features. Planta 2021, 254, 129. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Xu, L.; Huang, J.; Serra, A.; Yang, H.; Tam, J.P. Potentides: New Cysteine-Rich Peptides with Unusual Disulfide Connectivity from Potentilla anserina. Chembiochem 2019, 20, 1995–2004. [Google Scholar] [CrossRef] [PubMed]
- Govindarajan, D.K.; Kandaswamy, K. Antimicrobial peptides: A small molecule for sustainable healthcare applications. Med. Microecol. 2023, 18, 100090. [Google Scholar] [CrossRef]
- Pires, Á.S.; Rigueiras, P.O.; Dohms, S.M.; Porto, W.F.; Franco, O.L. Structure-guided identification of antimicrobial peptides in the spathe transcriptome of the non-model plant, arum lily (Zantedeschia aethiopica). Chem. Biol. Drug Des. 2019, 93, 1265–1275. [Google Scholar] [CrossRef] [PubMed]
- Shelenkov, A.; Slavokhotova, A.; Odintsova, T. Predicting antimicrobial and other cysteine-rich peptides in 1267 plant transcriptomes. Antibiotics 2020, 9, 60. [Google Scholar] [CrossRef] [PubMed]
- Tsekouras, V.; Mavrikou, S.; Vlachakis, D.; Makridakis, M.; Stroggilos, R.; Zoidakis, J.; Termentzi, A.; Moschopoulou, G.; Kintzios, S. Proteome analysis of leaf, stem and callus in Viscum album and identification of lectins and viscotoxins with bioactive properties. Plant Cell Tissue Organ. Cult. 2020, 141, 167–178. [Google Scholar] [CrossRef]
- Hayes, B.M.E.; Bleackley, M.R.; Anderson, M.A.; van der Weerden, N.L. The plant defensin NaD1 enters the cytoplasm of Candida albicans via endocytosis. J. Fungi 2018, 4, 20. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.; Arendt, E.K.; Thery, T.L.C. Isolation and characterisation of the antifungal activity of the cowpea defensin Cp-thionin II. Food Microbiol. 2019, 82, 504–514. [Google Scholar] [CrossRef] [PubMed]
- Dancewicz, K.; Slazak, B.; Kiełkiewicz, M.; Kapusta, M. Behavioral and physiological effects of Viola spp. cyclotides on Myzus persicae (Sulz.). J. Insect Physiol. 2020, 122, 104025. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Catacutan, D.B.; Rathod, K.; Swanson, K.; Jin, W.; Mohammed, J.C.; Chiappino-Pepe, A.; Syed, S.A.; Fragis, M.; Rachwalski, K.; et al. Deep learning-guided discovery of an antibiotic targeting Acinetobacter baumannii. Nat. Chem. Biol. 2023, 19, 1342–1350. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.-L.; Akash, K.; Reid, T.; Jain, N. Computational modeling of the dynamics of human trust during human-machine interactions. IEEE Trans. Hum.-Mach. Syst. 2019, 49, 485–497. [Google Scholar] [CrossRef]
- Rabaan, A.A.; Alhumaid, S.; Mutair, A.A.; Garout, M.; Abulhamayel, Y.; Halwani, M.A.; Alestad, J.H.; Bshabshe, A.A.; Sulaiman, T.; AlFonaisan, M.K.; et al. Application of artificial intelligence in combating high antimicrobial resistance rates. Antibiotics 2022, 11, 784. [Google Scholar] [CrossRef] [PubMed]
- Amsterdam, D. Perspective: Limiting antimicrobial resistance with artificial intelligence/machine learning. BME Front. 2023, 4, 0033. [Google Scholar] [CrossRef] [PubMed]
- Ali, T.; Ahmed, S.; Aslam, M. Artificial Intelligence for Antimicrobial Resistance Prediction: Challenges and Opportunities towards Practical Implementation. Antibiotics 2023, 12, 523. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hack, M.E.; El-Saadony, M.T.; Salem, H.M.; El-Tahan, A.M.; Soliman, M.M.; Youssef, G.B.A.; Ayman, E.T.; Soliman, S.M.; Ahmed, A.E.; El-kott, A.F.; et al. Alternatives to antibiotics for organic poultry production: Types, modes of action and impacts on bird’s health and production. Poult. Sci. 2022, 101, 101696. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Sharma, S. Role of alternatives to antibiotics in mitigating the antimicrobial resistance crisis. Indian J. Med. Res. 2022, 156, 464–477. [Google Scholar] [CrossRef] [PubMed]
- Lamut, A.; Peterlin Masic, L.; Kikelj, D.; Tomasic, T. Efflux pump inhibitors of clinically relevant multidrug resistant bacteria. Med. Res. Rev. 2019, 39, 2460–2504. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, D.; González-Bello, C. Siderophores: Chemical tools for precise antibiotic delivery. Bioorganic Med. Chem. Lett. 2023, 87, 129282. [Google Scholar] [CrossRef] [PubMed]
- Morita, Y.; Nakashima, K.-I.; Nishino, K.; Kotani, K.; Tomida, J.; Inoue, M.; Kawamura, Y. Berberine Is a Novel Type Efflux Inhibitor Which Attenuates the MexXY-Mediated Aminoglycoside Resistance in Pseudomonas aeruginosa. Front. Microbiol. 2016, 7, 1223. [Google Scholar] [CrossRef] [PubMed]
- Seukep, A.J.; Mbuntcha, H.G.; Zeuko’o, E.M.; Woquan, L.S.; Nembu, N.E.; Bomba, F.T.; Watching, D.; Kuete, V. Chapter Five—Established Antibacterial Drugs from Plants; Kuete, V., Ed.; Advances in Botanical Research; Academic Press: Cambridge, MA, USA, 2023; Volume 106, pp. 81–149. [Google Scholar]
- Jubair, N.; Rajagopal, M.; Chinnappan, S.; Abdullah, N.B.; Fatima, A. Review on the Antibacterial Mechanism of Plant-Derived Compounds against Multidrug-Resistant Bacteria (MDR). Evid. Based Complement. Alternat. Med. 2021, 2021, 3663315. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.; Li, J.; Sun, Z. The Combination of Antibiotic and Non-Antibiotic Compounds Improves Antibiotic Efficacy against Multidrug-Resistant Bacteria. Int. J. Mol. Sci. 2023, 24, 15493. [Google Scholar] [CrossRef] [PubMed]
- Shang, D.; Liu, Y.; Jiang, F.; Ji, F.; Wang, H.; Han, X. Synergistic Antibacterial Activity of Designed Trp-Containing Antibacterial Peptides in Combination with Antibiotics against Multidrug-Resistant Staphylococcus epidermidis. Front. Microbiol. 2019, 25, 2719. [Google Scholar] [CrossRef] [PubMed]
- Si, Z.; Pethe, K.; Chan-Park, M.B. Chemical Basis of Combination Therapy to Combat Antibiotic Resistance. JACS 2023, 3, 276–292. [Google Scholar] [CrossRef] [PubMed]
- Miethke, M.; Pieroni, M.; Weber, T.; Brönstrup, M.; Hammann, P.; Halby, L.; Arimondo, P.B.; Glaser, P.; Aigle, B.; Bode, H.B.; et al. Towards the sustainable discovery and development of new antibiotics. Nat. Rev. Chem. 2021, 5, 726–749. [Google Scholar] [CrossRef] [PubMed]
- Siriphap, A.; Kiddee, A.; Duangjai, A.; Yosboonruang, A.; Pook-In, G.; Saokaew, S.; Sutheinkul, O.; Rawangkan, A. Antimicrobial Activity of the Green Tea Polyphenol (−)-Epigallocatechin-3-Gallate (EGCG) against Clinical Isolates of Multidrug-Resistant Vibrio cholerae. Antibiotics 2022, 11, 518. [Google Scholar] [CrossRef] [PubMed]
- Bonincontro, G.; Scuderi, S.A.; Marino, A.; Simonetti, G. Synergistic Effect of Plant Compounds in Combination with Conventional Antimicrobials against Biofilm of Staphylococcus aureus, Pseudomonas aeruginosa, and Candida spp. Pharmaceuticals 2023, 16, 1531. [Google Scholar] [CrossRef] [PubMed]
- Selvarajan, R.; Obize, C.; Sibanda, T.; Abia, A.L.K.; Long, H. Evolution and Emergence of Antibiotic Resistance in Given Ecosystems: Possible Strategies for Addressing the Challenge of Antibiotic Resistance. Antibiotics 2023, 12, 28. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Chauhan, A.; Ranjan, A.; Mathkor, D.M.; Haque, S.; Ramniwas, S.; Tuli, H.S.; Jindal, T.; Yadav, V. Emerging challenges in antimicrobial resistance: Implications for pathogenic microorganisms, novel antibiotics, and their impact on sustainability. Front. Microbiol. 2024, 15, 1403168. [Google Scholar] [CrossRef] [PubMed]
- Bobate, S.; Mahalle, S.; Dafale, N.D.; Bajaj, A. Emergence of environmental antibiotic resistance: Mechanism, monitoring and management. Environ. Adv. 2023, 13, 100409. [Google Scholar] [CrossRef]
- Samreen, A.I.; Malak, H.A.; Abulreesh, H.H. Environmental antimicrobial resistance and its drivers: A potential threat to public health. J. Glob. Antimicrob. Resist. 2021, 27, 101–111. [Google Scholar]
- Zhang, L.; Tian, X.; Sun, L.; Mi, K.; Wang, R.; Gong, F.; Huang, L. Bacterial Efflux Pump Inhibitors Reduce Antibiotic Resistance. Pharmaceutics 2024, 16, 170. [Google Scholar] [CrossRef] [PubMed]
- Walsh, T.R. A one-health approach to antimicrobial resistance. Nat. Microbiol. 2018, 3, 854–855. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).