Abstract
Pseudomonas bacteria are renowned for their remarkable capacity to synthesize antibiotics, namely mupirocin, gluconic acid, pyrrolnitrin, and 2,4-diacetylphloroglucinol (DAPG). While these substances are extensively employed in agricultural biotechnology to safeguard plants against harmful bacteria and fungi, their potential for human medicine and healthcare remains highly promising for common science. However, the challenge of obtaining stable producers that yield higher quantities of these antibiotics continues to be a pertinent concern in modern biotechnology. Although the interest in antibiotics of Pseudomonas bacteria has persisted over the past century, many uncertainties still surround the regulation of the biosynthetic pathways of these compounds. Thus, the present review comprehensively studies the genetic organization and regulation of the biosynthesis of these antibiotics and provides a comprehensive summary of the genetic organization of antibiotic biosynthesis pathways in pseudomonas strains, appealing to both molecular biologists and biotechnologists. In addition, attention is also paid to the application of antibiotics in plant protection.
1. Introduction
Numerous soil bacteria have the ability to generate secondary metabolites that exhibit bacteriostatic and bactericidal properties. This enables them to thrive and maintain their competitiveness in demanding microbiome habitats [1,2,3]. Pseudomonas bacteria are renowned for their ability to produce a wide variety of antibiotic secondary metabolites, solidifying their reputation as some of the most prolific contributors to the field of microbial drug discovery [4,5]. The variety of compounds produced by Pseudomonas strains is mind-boggling.
Pseudomonas strains’ antibiotics play a crucial role in plant protection biotechnology by serving as antibacterial and antifungal agents [6,7]. Among them, fluorescent pseudomonads that produce phenazine co-compounds are the most commonly utilized [8]. Commercial products based on P. aureofaciens, P. chlororaphis, P. fluorescens, and P. syringae are widely recognized [9,10,11]. Various research papers offer an overview of the fluorescent Pseudomonas strain-based formulations used for plant protection in Europe, Australia, and the USA [12,13,14,15]. Typically, these strains were effective against pathogenic fungi, with some cases showing success against other pseudomonads [16]. One notable example was Pseudomonas synxantha PS54 [17], which was patented as a control agent against P. tolaasii [18], the causative agent of bacterial blotch in the mushroom Agaricus bisporus [19,20].
In the 20th century, antibiotics produced by Pseudomonas strains were utilized for human therapy, in addition to plant protection biotechnology. The strain Pseudomonas lindbergii (ATCC-31099) [21] was patented as a producer of antifungal compounds, which were claimed to be more effective than griseofulvin, an antibiotic widely used by the USA Army against fungal infections. To demonstrate the efficacy of the P. lindbergii antibiotics, they were put on filter paper and placed on the feet and infected toenails of patients. It was claimed that the manifestations of infection disappeared after 2–3 days.
Additionally, P. lindbergii and Bacillus coagulans have been combined in preparations for controlling mycoses, showing promising results [22,23]. For prophylaxis or therapeutic treatment, the clean and dry feet of patients were treated with a preparation containing supernatant or filtrate of B. coagulans or P. lindbergii. In addition to fungi, antibiotics produced by P. lindbergii were effective against Streptococcus, Staphylococcus, E. coli, Serratia, Klebsiella and Proteus [21].
Phenazine antibiotics, a group of compounds produced by Pseudomonas strains, have been extensively studied. The production of these antibiotics involved genetic and technological aspects, highlighting the importance of a complete metabolic pathway for their synthesis. The information available about them to date is summarized in reviews [24,25]. Our attention was focused on genetic and technological aspects of the production of the following antibiotics: mupirocin, gluconic acid, pyrrolnitrin (PRN), and 2,4-diacetylphloroglucinol (DAPG). All of the above antibiotics, except gluconic acid, are typical secondary metabolites, the production of which requires a separate genetic system—that is, an operon containing genes encoding the complete metabolic pathway. Gluconic acid is a product of primary metabolism, and numerous bacterial strains have the ability to synthesize it, but the key factor lies in the amount produced. Only those particular strains that generate sufficient quantities of gluconic acid to hinder the proliferation of other microbes can be identified as gluconic acid producers [26].
While the search for improved antibiotic producers from Pseudomonas strains is a common theme in the scientific literature, there is surprisingly limited discussion on their patenting and applications in biotechnology [27,28]. This gap in the research highlights the potential for innovative discoveries in this field [29].
The primary goal of this review was twofold. Firstly, to consolidate the information on the mechanisms of antibiotic synthesis by Pseudomonas strains and the control of these processes. The approaches for controlling antibiotic production were analyzed, encompassing genetic methods (such as mutagenesis of specific genes) as well as biochemical strategies, including modifying the cultivation medium composition and incorporating substances that may impact the final product yield.
The focus of the latter part of the study was on the implementation of antibiotic-producing Pseudomonas strains in the field of biotechnology. While traditional strains were commonly utilized, there were instances where genetically altered variants were employed. This study explored a range of techniques for modifying these producers in order to enhance the final product yield compared to the original strains.
Each section of this paper is focused on a separate antibiotic and contains information on the genetic organization of the biosynthesis pathway, the regulation of production, and the application of the antibiotic in medical and agricultural biotechnology.
Scientific information was searched using the Google and PubMed databases. More than 500 publications concerning antibiotic production by pseudomonads were analyzed. This review includes publications containing information on the mechanisms of biosynthesis, regulation and quantitative yields of the antibiotics. Patent searches were performed using the Google Patents, Justia Patents and PubChem services. The patent analysis focused on the efficiency of the antibiotic production and the method of obtaining the maximum yield.
2. Mupirocin
Mupirocin is an antibiotic consisting of a mixture of pseudomonic acids (A, B, C, D) [30]. Mupirocin, also known as pseudomonic acids (Figure 1), is utilized for controlling skin infections and preventing postoperative wound inflammation when used intranasally [31,32].
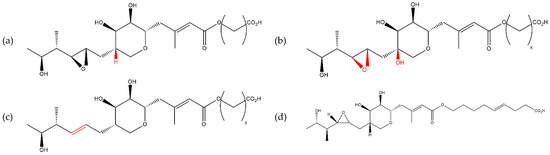
Figure 1.
Chemical structures of pseudomonic acids (a) A, (b) B, (c) C, and (d) D.
The components of the mupirocin mixture have a similar structure and consist of two basic elements: monic acid and 9-hydroxynonanoic acid connected by an ester bond [32] (Figure 2).
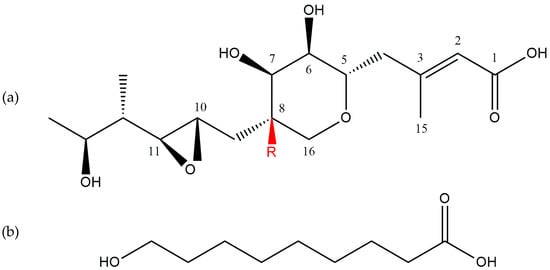
Figure 2.
The main subunits of pseudomonic acids: (a) monic acid and (b) 9-hydroxynonanoic acid [30]. R = H or OH. All the structures were taken from PubChem, an open chemistry database.
Pseudomonic acid is produced by industrial strains of P. fluorescens as part of a mixture of metabolites. At least three variants of such mixtures are known (Table 1).

Table 1.
The known mixtures of pseudomonic acids produced by industrial strains of P. fluorescens..
Since pseudomonic acid A was the dominant metabolite of the mixture (90–95%), it was it that exhibited the main antimicrobial effect on test cultures [37]. The other pseudomonic acids had the same spectrum of action, but their antimicrobial effect was weaker, possibly due to their lower quantity [36].
Pseudomonas fluorescens strain NCIMB 10586 is widely recognized as the leading producer of mupirocin [28,38,39]. Extensive research has focused on studying mupirocin production in this strain [40,41]. Besides Pseudomonas fluorescens NCIMB 10586, other pseudomonads belonging to the P. fluorescens species group have also been investigated for mupirocin production [28,42]. Haines et al. [43] demonstrated the close relationship between strain NCIMB 10586 and the species P. synxantha and P. libaniensis. Matthijs et al. [28] revealed that the closest relative of strain NCIMB 10586 is a type strain of the species P. azotoformans, with similarities also being observed with the type strains P. cedrina subsp. cedrina CFML 96-198T, P. cedrina subsp. fulgida DSM 14938T, P. libanensis CIP 105460T and P. synxantha IAM 12356. The intricate taxonomic relationships within the genus Pseudomonas make it challenging to establish clear boundaries between these species.
2.1. Mechanism of Mupirocin Biosynthesis
The pathway of mupirocin biosynthesis is regulated by a 74 kb gene cluster consisting of two transcriptional units, which can be roughly divided into two major modules [44,45] (Table 2).
Table 2.
Structure of the mupirocin biosynthesis gene cluster.
A similar gene cluster was discovered in Pseudomonas sp. strain BRG100, which is utilized in agriculture for controlling Setaria viridis (Green Foxtail) and other pests [46,47]. A similar gene cluster but with a less resemblance was discovered by Haines et al. [43] in the genome of Pseudomonas psychrotolerans strain NS383.
The mutagenesis of individual elements of the mupirocin gene cluster showed the following:
- The mupQ, mupS, mupT, and mupW genes were essential for the production of mupirocin, whereas mupO, mupU, mupV, and macpE were essential for the production of PA-A but not PA-B [45]. In this work, it was assumed for the first time that PA-B is a precursor of PA-A.
- PA-C, previously assumed to be a precursor of PA-A, was formed by a minor parallel pathway [41]. Attempting to disable this pathway at the initial stage (ΔmupW) resulted in the loss of the ability to synthesize the major product (PA-A). Moreover, all the mutagenesis operations of the mupirocin cluster elements resulted in the loss of the ability to synthesize PA-A, but ΔmupC and ΔmupF retained PA-B production.
PA-C, the simplest component of the pseudomonic acid mixture, was chemically more stable than PA-A [41]. This was due to the presence of 10,11-epoxide in the structure of PA-A, which made it susceptible to intramolecular attack by the 7-OH. PA-C, which did not contain epoxide in its structure but had the same properties as PA-A and was more promising for pharmaceutical applications as an antibiotic. PA-C was produced by chemical synthesis [48,49,50,51], but attempts to obtain a high-yield producer of PA-C alone through modifications to the metabolic pathway have not been successful [41].
2.2. Regulation of Mupirocin Production
Two proteins encoded by the mupR and mupI genes governed the regulation of mupirocin production [52] (Table 3). Upon comparing their amino acid sequences with known sequences from the databases, it was revealed that mupR and mupI exhibit significant similarities to proteins associated with quorum-sensing regulation in P. aeruginosa and Vibrio fischeri [52].
Table 3.
Role of mupI and mupR in the regulation of mupirocin production.
The mup operon demonstrated peak expression during the stationary phase, while attempts to induce its expression in the exponential phase have proven unsuccessful, indicating active repression during this stage [52].
As mentioned earlier, mupirocin is produced as a mixture of the A, B, C, and D pseudomonic acids. According to the hypothesis, overexpression of macpE, MupO, U, V, C, and F could increase the conversion of PA-B to PA-A, thus yielding a higher yield of the major metabolite [54]. However, Macioszek [53] and Gurney [55] showed that introducing these genes in trans into a wild-type strain did not increase the conversion of PA-B to PA-A, but it increased the yield of each of these compounds by two-fold proportionally.
Hothersall et al. [44] increased the efficiency of PA-A production through increased expression of the transcriptional regulator MupR. The introduction of plasmids containing (1) MupR only and (2) MupR + MupX into the wild-type strain (WT) increased the yield of PA-A by 4–5 times.
Genetic manipulation looks like a promising way to increase the proportion of pseudomonic acid A in the mixture and the overall production efficiency. However, we found no information on the patenting and use of mutant variants modified to produce only pseudomonic acid A. The only mention of such mutants was in the patent of Szell et al. [36] for Pseudomonas sp. strain No. 19/26 (NCAIM(P)B 001235). Basically, however, increasing the degree of production and the proportion of pseudomonic acid A in the mixture is currently achieved by modifying the cultivation conditions. In [56] the ratio of acids A and B in the mixture was 97:3, which was achieved by selecting the cultivation conditions of Pseudomonas sp. strain No. 19/26 (NCAIM(P)B 001235), namely pH 5.7, carbon source—dextrose, the concentration of which should be <0.5% for the entire period of production, addition of calcium chloride to a final concentration of 0.1% w/w. The authors obtained the maximum yield of pseudomonic acid A with 3.021 μg/g of culture medium, while the presence of pseudomonic acid in the product was 2.9% w/w.
2.3. Mupirocin in Biotechnology
One of its advantages compared to other antibiotics is its low toxicity to humans and animals [36]. Primarily, it was employed against Staphylococcus aureus, but it also demonstrated effectiveness against Streptococcus [57], Neisseria [58], as well as several other Gram-positive and Gram-negative bacteria [59,60,61,62].
The best-known mupirocin-based preparations are Bactroban (UK), Turixin (Germany) and Bactoderm (Israel) [40]. These preparations are available in the form of creams and ointments containing 2% of the active ingredient. Bactroban ointment, nasal agent and cream are protected by patents US4,524,075 [63], US4,790,989 [64] and US6,025,389 [65], respectively. The ointment contains mupirocin, while the nasal agent and cream contain crystalline mupirocin calcium dihydrate [66].
The original mupirocin patent [34] detailed the process of creating the sodium variant of pseudomonate. However, due to its hygroscopic nature, Curzons proposed the utilization of lithium pseudomonate instead [67,68]. Lithium pseudomonate proved to be a reliable and non-hygroscopic crystalline compound, ensuring its stability over time. The works [53,67] used the same producer, Pseudomonas fluorescens strain NCIB 10586. Lithium pseudomonate (mupirocin lithium) is available as an analytical preparation from Sigma (Burlington, MA, USA) or Thermo Fisher Scientific (Waltham, MA, USA), but it is not used for drug production, unlike mupirocin calcium. The calcium salt of mupirocin is used in ointments, as is pseudomonic acid.
Patents for crystalline [69,70,71,72] and amorphous [73] salts of pseudomonic acid are known. Previously, it has been repeatedly argued that amorphous calcium pseudomonate cannot withstand high temperatures and degrades during storage [69,71,74], whereas the crystalline form is convenient for pharmaceutical use. However, Weisman et al. [73] showed that in terms of the temperature stability, amorphous calcium pseudomonate is comparable to pseudomonic acid, which is widely used in pharmaceuticals. Calcium pseudomonate is available as an analytical preparation from Sigma.
3. Gluconic Acid
Gluconic acid (Figure 3) possesses antifungal properties and is commonly employed to control plant diseases [75,76,77,78,79]. Gram-negative bacteria utilize the oxidation of glucose to gluconic acid, which takes place in the bacterial periplasm, as a means of solubilizing inorganic phosphorus. This process provides them with a competitive advantage over other organisms, as it converts accessible carbon sources into ones that are less available to other microorganisms [80,81].
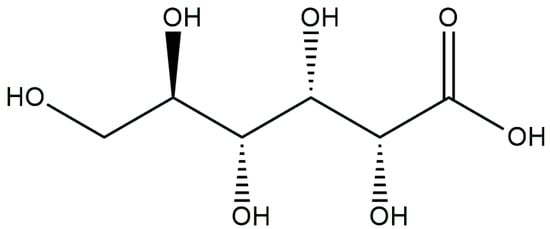
Figure 3.
Chemical structure of gluconic acid.
The first Pseudomonas strains in which gluconic acid production was observed were Pseudomonas ovalis NRRL B1486 [82,83,84] and P. ovalis NRRL B-SS [75] obtained from the US Dept of Agriculture, Peoria, USA. Gluconic acid was the major organic acid produced during phosphate solubilization in Pseudomonas sp. [85], P. fluorescens [86], P. savastanoi [87] and Pseudomonas corrugata [88].
3.1. Gluconic Acid Biosynthesis Pathway
The pathway of gluconic acid biosynthesis can be categorized as a fundamental metabolic process. Unlike fungi, where the conversion of glucose to gluconic acid is regulated by glucose oxidase (b-D-glucose: oxygen 1-oxidoreductase, E.C. 1.1.3.4) [89,90], bacteria employ glucose dehydrogenase (GDH, E.C. 1.1.99.17) to perform this reaction [91,92,93]. The glucose dehydrogenase enzyme is situated in the membrane and is induced by high concentrations of glucose (>15 mM) [94]. While gluconic acid participates in the bacterial cell’s pentose phosphate pathway, its production is repressed at elevated (>15 mM) glucose levels, leading to the accumulation of this metabolite in the environment [87].
Currently, the best-known producer of gluconic acid is the Pseudomonas strain AN5 [26,95,96]. The production of other antibiotics common for Pseudomonas strains (phenazine-1-carboxylic acid (PCA) and DAPG) was absent in strain AN5 [78].
3.2. Regulation of Gluconic Acid Production
Based on Pseudomonas AN5, two transposon mutants (AN5MN1 and AN5MN2) with no antimicrobial properties have been constructed [26]. Yu et al. [79], also through transposon mutagenesis of the strain Pseudomonas capeferrum WCS358, showed that two genes are required for gluconic acid biosynthesis: pqqF and cyoB. PqqF was a putative protease involved in pyrroloquinoline quinone (PQQ) biosynthesis [97,98]. PQQ functioned as a cofactor in many carbon utilization reactions [99]. In turn, CyoB was a subunit of the terminal ubiquinol cytochrome bo3 oxidase (CYO) complex, which was required in the aerobic respiratory chain for energy generation [100]. Both mutant variants demonstrated a reduced ability to produce organic acids, and therefore, reduced the environmental pH less efficiently than the wild-type strain [79].
Galet et al. [77] conducted Tn5 mutagenesis and found that the pqqE gene plays a critical role in the biosynthesis of gluconic acid. A mutant variant of P. fluorescens strain BBc6R8 with a defective pqqE gene was incapable of gluconic acid production, and a similar mutant of P. fluorescens strain F113 produced very small amounts of organic acid [101]. Galet et al. [77] suggested that the pQQFABCDE operon was required for gluconic acid biosynthesis in pseudomonads.
Trivedi and Sa [88] conducted a genetic modification experiment on a Pseudomonas corrugata strain (NRRL B-30409) using N’-methyl-N’-nitro-N-nitrosoguanidine (NTG), aiming to create cold-tolerant mutants capable of efficiently solubilizing phosphate at lower temperatures. Nevertheless, it is important to note that while chemical-induced mutagenesis can result in enhanced production efficiency in terms of the desired compound, it does not ensure the reproducibility of the experiment and thus cannot be considered a reliable approach for the development of biotechnologically promising producers [102].
De Werra et al. [76] investigated the engineering of Pseudomonas fluorescens strain CHA0 to enhance the production of gluconic acid [76]. They found that by disabling the gad (gluconate dehydrogenase) gene, which is responsible for the conversion reaction of gluconic acid to 2-ketogluconate, the cells accumulated a higher concentration of gluconic acid compared to the wild-type strain. Interestingly, the mutant variants of the gcd and gad genes exhibited different capabilities in terms of producing other antifungal components, namely DAPG and pyoluteorin (PLT). The mutant strain Δgcd, with glucose dehydrogenase inactivation, demonstrated a more efficient production of DAPG compared to the wild-type strain. Additionally, the Δgcd mutant produced 10 times more DAPG than the Δgad mutant (4 μM) after just 1 day of cultivation. Moreover, the Δgcd mutant showed the ability to produce PLT, while this component was absent in the wild-type strain. Thus, it could be concluded that the Δgcd mutant was more effective in producing the antifungal components DAPG and PLT, while the Δgad mutant was more efficient as a producer of gluconic acid.
In addition to the antifungal properties, Pseudomonas strains producing gluconic acid can participate in intermicrobial interactions in open ecosystems. Decreasing the pH of the cultivation medium due to gluconic acid production by P. fluorescens strain BBc6R8 in [77] prevented the production of γ-actinorhodin by S. coelicolor. Actinorhodin is a benzoisochromanequinone dimer polyketide antibiotic whose production is characteristic of Streptomyces [103,104]. All the gluconic acid producers used by Galet et al. [77] (P. fluorescens SBW25 [105], P. fluorescens Pf0-1 [106], P. protegens Pf-5 [107] and P. aeruginosa PAO1 [108]) showed similar effects on actinorhodin production by Streptomyces strains. The demonstrated effect was not due to specific properties of gluconic acid but to a general decrease in the pH of the cultivation medium.
In addition to Streptomyces, the effect of gluconic acid on the production of the siderophore pyoverdine by P. putida is also known. Ponraj et al. [109] showed that when the pH of the medium decreases, the efficiency of the pyoverdine production decreases, which may ultimately affect the efficiency of the plant root colonization. Ponraj et al. [109] noted that in the case of pyoverdine, there was no effect of gluconic acid on the expression of biosynthetic genes. A decrease in the pH of the medium affects the periplasmic transformation of pyoverdine and its secretion from the cell.
3.3. Gluconic Acid in Biotechnology
As the subject of patents in the USA and Europe [26,78], strain AN5 has emerged as a significant producer of gluconic acid. Its original purpose was to combat Gaeumannomyces graminis var. tritici (Ggt), but subsequent findings [78] revealed that the gluconic acid derived from strain AN5 possessed valuable properties against Deuteromycetes (E. pupurescens, Alternaria sp., A. oligosporus, M. fructocola, B. cinerea, V. dahlia), Basidiomycetes (F. annosus, A. mellea, B. granulatus, P. suphureus), S. cerevisiae, as well as a broad spectrum of both Gram-negative and Gram-positive bacteria.
According to the research, Pseudomonas strain AN5 exhibited a gluconic acid production rate of approximately 5.0 mg/mL. Notably, Kaur et al. [26] emphasized that there were no known reports of other pseudomonads with similar levels of gluconic acid production. Nevertheless, Vyas and Gulati [110] observed in P. trivialis, P. poae and P. fluorescens strains the production of gluconic acid at approximately 17,000–19,000 μg/mL (17–19 mg/mL) during tricalcium phosphate solubilization.
4. PRN
PRN [3-chloro-4-(2-nitro-3-chlorophenyl)-pyrrole] (Figure 4) exhibited notable antifungal properties, effectively combating a wide range of fungal infections (Alternaria sp., Botrytis cinerea, Pythium aphanidermatum, P. ultimum, Rhizoctonia solani, Rhizopus sp., Aspergillus niger, Fusarium oxysporum, Penicillium expansum, and Sclerotium rolfsii) and bacteria (Agrobacterium tumefaciens, Corynebacterium insidiousum, Pseudomonas syringae pv. syringae, and Xanthomonas campestris) [111]. It was first isolated from Pseudomonas pyrrocinia [112] in 1964. PRN producers belonging to different Pseudomonas species are now known: P. fluorescens [113,114,115], P. chlororaphis [116,117,118,119], and P. cepacia [120]. PRN is composed of a benzene ring and a pyrrole ring, each adorned with chlorine atoms. Additionally, the benzene ring harbors a nitro group.
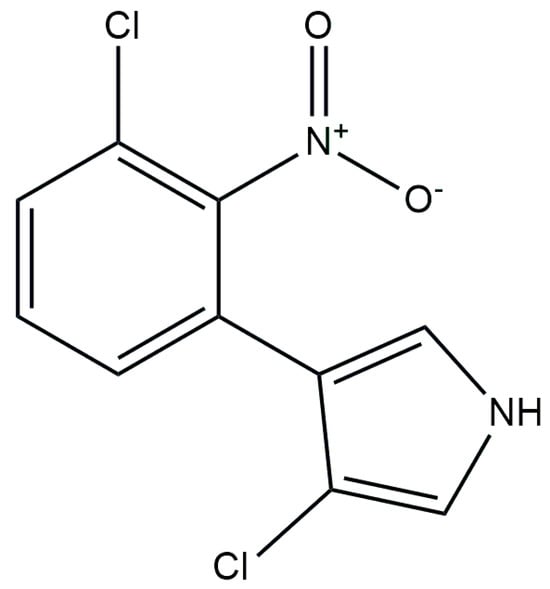
Figure 4.
Chemical structure of PRN.
4.1. Mechanism of PRN Biosynthesis
Tryptophan serves as a direct precursor of PRN, with both L- [121] and D- [122] forms being available. Microorganisms could utilize tryptophan analogs introduced into the culture medium to synthesize PRN-like compounds. However, it should be noted that these compounds exhibited lower antimicrobial activity compared to PRN [122,123]. The synthesis of PRN from tryptophan involved a series of four reactions regulated by the prn operon (Figure 5) [65,113,116,123,124].
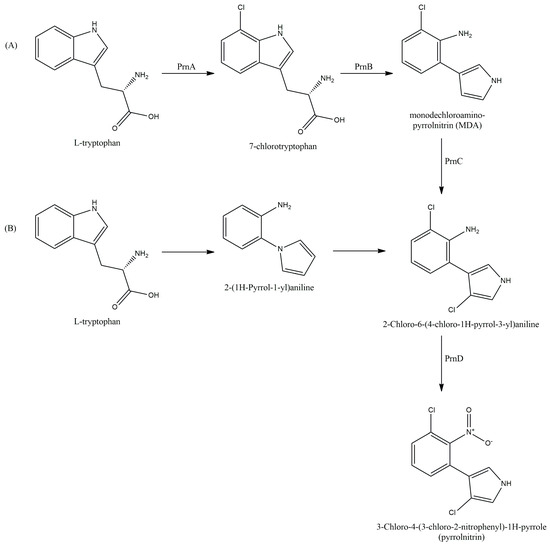
Figure 5.
PRN biosynthesis pathway according to (A) van Pée et al. [125] and (B) Chang et al. [126].
Hammer et al. [113] discovered a 5.8 kb operon containing four genes responsible for PRN biosynthesis in the genome of a P. fluorescens strain. The authors named this operon prnA-D. However, Kirner et al. [115] and van Pée [127] later revealed that chloroperoxidases, previously believed to be involved in PRN biosynthesis [128], were not actually involved in this process. Using the prn operon genes as a reference for cloning and sequencing homologous genes, Hammer et al. [116] demonstrated that the prn gene cluster was highly conserved and could be found in other Pseudomonas strains, Burkholderia cepacia and Myxococcus fulvus.
PrnA and PrnB proteins have been isolated and thoroughly characterized [124,129], while prnD has been 3-D modeled to determine its protein structure [130]. De Souza et al. [131] developed a method to detect prnD in soils to assess the potential diversity of producers in ecosystems.
In contrast to previous findings by van Pée regarding the prn metabolic pathway [125], Chang et al. [126] proposed that the product of the first reaction of the PRN biosynthesis pathway in P. aureofaciens may be aminophenylpyrrole rather than 7-chlorotryptophan.
4.2. Regulation of PRN Production
The synthesis of numerous secondary metabolites is regulated by global regulators. For example, mutations in the gacA gene in P. fluorescens strain CHA0 [132] impede the production of DAPG, hydrogen cyanide (HCN), and pyoluteorin. Similarly, in the Pseudomonas fluorescens strain Pf-5 that failed to produce PRN, a mutation was detected in the rpoS gene, the product of which was involved in the biosynthesis of secondary metabolites, particularly PRN [133,134]. By introducing a plasmid containing a functional copy of rpoS, the researchers managed to restore the PRN production ability in Pseudomonas fluorescens strain Pf-5 [133]. In a study conducted by Park et al. [119] using the P. chlororaphis strain, it was demonstrated that functional (not defective) rpoS, gacS (a component of the two-component GacS/GacA system), and prnA genes are necessary for PRN production. The highest expression level of prnA was observed during the stationary phase of growth.
In contrast with Saringuet et al. [133], Manuel et al. [135] showed that a mutation in the rpoS gene of Pseudomonas chlororaphis strain PA23 conferred the increased antifungal properties of the strain against Sclerotinia sclerotiorum and elevated the PRN, lipase and protease production. When rpoS was added in trans, the production levels of the above compounds returned to those of the wild-type strain. Given the difference in the effect of mutant rpoS on PRN production by Pseudomonas fluorescens Pf-5 [133] and Pseudomonas chlororaphis PA23 strains (negative in the former and positive in the latter), Manuel et al. [135] suggested that the difference may be due to the different spectrum of metabolites produced by the strains, or that RpoS may affect PRN expression not directly but through regulatory elements different in these bacteria.
Another regulator of PRN biosynthesis is the vfr gene [136]. Vfr is a cAMP-dependent global regulator of virulence factor expression in Pseudomonas [137,138,139]. By knockout mutagenesis, it was shown that when the vfr gene is switched off, the prn operon expression critically drops and the amount of PRN produced is quite negligible. At the same time, the level of PCA production was unchanged in vfr-defective variants. Wu et al. [136] concluded that Vfr is a regulator of only PRN production, not of phenazines.
Huang et al. [117] used knockout mutant variants of Pseudomonas chlororaphis strain G05 (Δphz, Δprn and ΔphzΔprn) to prove that disabling the phenazine biosynthesis cluster does not affect the antifungal capabilities of the strain too much. This work showed for the first time that it is not phenazines but PRN that plays a key role in the control of F. graminearum and the bioprotection of cereal crops against diseases.
Park et al. [119] showed that the presence of glucose in the culture medium inhibited the production of PRN by the Pseudomonas chlororaphis strain. On medium without glucose, the product yield was 1.7 µg/mL, while when glucose was added, the yield was 0.2 µg/mL. Interestingly, the effect on PCA production was reversed: 2.5 µg/mL without glucose and 27.4 µg/mL with glucose. In this work, as well as in [117], it was demonstrated that the antifungal effect on F. graminearum and R. solani is exerted more by PRN than by phenazines.
Keum et al. [140] demonstrated that the production of PRN by pseudomonads was influenced by the presence of xenobiotics, particularly pesticides. In the case of Pseudomonas fluorescens strain Pf-5, accumulation of PRN (up to 6 mg/L) or amino-PRN (up to 7 mg/L), respectively, was observed when cultured in the presence of 10 μm fenpiclonil or fludioxonil. Additionally, when cultured with fludioxonil, the culture medium contained approximately 2.5 mg/L of PRN. As for the control group without pesticides, the accumulation of approximately 2 mg/L PRN and 7.5 mg/L amino-PRN was observed. Thus, fenpiclonil was found to increase the production of PRN by three times, while fludioxonil did not affect the yield of PRN but influenced the production of the intermediate metabolite, amino-PRN.
4.3. Modifications of PRN Producers
The first attempt to modify a PRN producer was made by Salcher and Lingens in 1979 [141]. They conducted a study to enhance the production efficiency of PRN production in Pseudomonas aureofaciens strain ATCC 15926 using variations in the culture conditions [142] and chemical mutagenesis. The original strain yielded 0.3 μg/mL PRN, and efforts to increase the yield through media selection proved futile. To induce mutations, the authors employed N-methyl-N′-nitro-N-nitrosoguanidine (MNNG) as a mutagen. Remarkably, the P. aureofaciens ACN mutant variant exhibited the best outcome, producing 9 μg/mL (9 mg/L) of PRN. Notably, the ACN variant colonies lacked color, contrary to the orange colonies of the parental strain. However, research by Salcher and Lingens in 1979 [141] revealed that both strains produced PCA, and the lack of color in the mutant may be due to an inability to synthesize 2-hydroxyphenazine-1-carboxylic acid [143]. Encouragingly, the ACN mutant variant remained stable over an extended period without reverting to the wild-type state.
In a groundbreaking study, Zhang et al. [144] successfully transferred the prn operon from P. protegens (formerly P. fluorescens [145]) Pf-5 into P. synxantha 2-79. The recipient strain initially possessed the ability to produce phenazines and displayed effectiveness against both take-all and Rhizoctonia root rot of wheat. This remarkable research marked the first documented instance where a prn operon introduced through transfer into a phenazine-producing recipient strain demonstrated consistent maintenance and expression. Moreover, the resulting recombinants exhibited significantly enhanced biocontrol properties when compared to the wild-type strain. Of notable importance, these recombinants displayed remarkable effectiveness against various pathogens, including Rhizoctonia solani, Gaeumannomyces graminis var. tritici, Sclerotinia sclerotiorum, Fusarium culmorum, and F. pseudograminearum significantly, surpassing the capabilities of P. synxantha 2-79.
4.4. PRN in Biotechnology
A series of P. fluorescens strains producing PRN have been patented in the USA for the purpose of managing diseases caused by Rhizoctonia and Pythium [146]. Additionally, P. cepacia, which has been isolated from the surface of apple leaves, has been patented as a method to control postharvest diseases in pome fruits [147]. While PRN is the only antibiotic produced by the aforementioned strains in the examples provided, there are instances where PRN was produced in conjunction with other compounds. Pseudomonas sp. strain DSM21663 [148] served as a producer of PRN, DAPG, and indole acetic acid. The aim of the patent was to utilize this particular strain for the management of the soil-borne root and foliar pathogens originating from both fungal and bacterial sources. Strain DSM21663 was classified as a fluorescent pseudomonad, with the 16S rRNA gene sequence demonstrating a similarity of 98.6% with P. congelans and 97.3% similarity with P. fluorescens.
Commercial preparations utilizing PRN derivatives (fludioxonil and fenpiclonil) have been registered in France and Switzerland (Table 4). The drugs are intended for use as fungicides and in agriculture [123].
Table 4.
Commercial preparations based on PRN derivatives.
The products have a long history of use (since 1993), since they are not microbial preparations but combinations of chemical compounds. No commercial bacterial preparations where PRN-producing pseudomonads have been used for plant defense have been found. Purified (≥98% purity) PRN from P. cepacia is sold by Sigma as a standard in the detection of PRN in Serratia marcescens cell culture extract [149].
5. DAPG
DAPG, as shown in Figure 6, is a polyketide antibiotic. It was previously believed that its production was specific to bacteria of the P. fluorescens species group (including P. protegens, P. brassicacearum, P. kilonensis, and P. thivervalensis). The genes responsible for DAPG synthesis were thought to have a monophyletic distribution, meaning they originated from a common ancestor [150]. However, a recent study by Almario et al. [151], which involved analyzing the genomes of various Pseudomonas strains, challenged this view. The researchers discovered that the genes involved in the biosynthesis pathway of DAPG biosynthesis (phl) were acquired independently through multiple events rather than being inherited vertically from a single ancestor. Almario et al. [151] found phl genes in pseudomonads, beyond P. fluorescens, as well as in two genera of Betaproteobacteria.
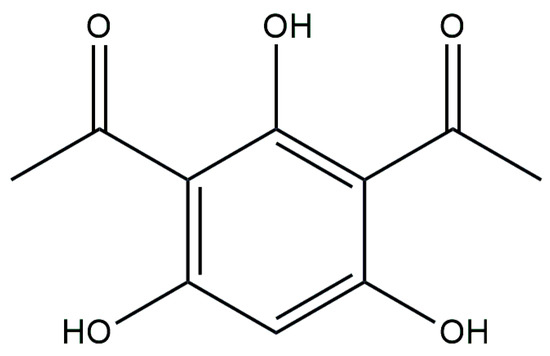
Figure 6.
Chemical structure of 1,1′-(2,4,6-Trihydroxy-1,3-phenylene)di(ethan-1-one) (DAPG).
In 1993, using the example of the strain Pseudomonas sp. F113, it was shown that the precursor of DAPG is MAPG, and an acyltransferase responsible for the conversion of MAPG to DAPG was described [152]. The genes for the DAPG biosynthesis pathway were first identified by Bangera and Thomashow [153] using Pseudomonas fluorescens strain Q2-87 as an example. The DAPG biosynthesis pathway has now been described in detail [151,154].
P. fluorescens strain Pf-5, originally identified as the best-known DAPG producer [155], was later reidentified as P. protegens [156]. Nowak-Thompson et al. [155] revealed that the composition and quantity of compounds produced by the strain (DAPG/pyoluteorin/PRN), as well as the absolute amount of each, varies depending on the culture medium. In particular, when 2% glucose was added as a carbon source, DAPG was the primary compound produced and was synthesized at 16.6 mg/L. However, when glucose was replaced by glycerol, the amount of synthesized DAPG dropped sharply (to 0.19 mg/L), but pyoluteorin was synthesized in the amount of 5.75 mg/L. These findings obtained by Nowak-Thompson et al. [155] for P. fluorescens (protegens) Pf-5 contrast with those obtained earlier by Shanahan et al. [157] for Pseudomonas sp. F113. The strain Pseudomonas sp. F113 actively produced DAPG when growing on sucrose, mannitol, and fructose, but no DAPG synthesis occurred on glucose despite the high numbers of growing cells.
5.1. Mechanism of DAPG Biosynthesis
Bangera and Thomashow [153] identified a 6.5 kb region consisting of six reading frames in strain Q2-87. Five of these (phlA, phlC, phlB, phlD, and phlE) were transcribed in one direction, phlF in the opposite direction. The authors demonstrated through knockout mutagenesis of the region components that (1) phlD was required for MAPG synthesis, and the mutant strain with a defective phlD gene was able to convert MAPG to DAPG but not to perform MAPG synthesis, and (2) phlA, B and C were required for MAPG to DAPG conversion. The protein products of these genes acted as a single complex: knockout of A, B, or C, even with functional D, resulted in the inability to synthesize MAPG and DAPG. Bangera and Thomashow [153] initially proposed that the reaction controlled by phlD used acetoacetylCoA as the starting unit, yielding MAPG. However, later, Achkar et al. [158] discovered that this reaction involves the formation of phloroglucinol from malonylCoA. It is suggested that phlD is responsible for the formation and cyclization of an activated 3,5,7-triketooctanoate [153,158] to synthesize MAPG, which is then acetylated to provide DAPG. Bangera and Thomashow [153] noted the relatedness of phlD to plant chalcone/stilbene synthases (CHS/STS). This is unusual, since antibiotic biosynthesis by bacteria is mainly controlled by type I and type II polyketide synthases [155,159,160]. Furthermore, Gupta et al. [161] confirmed the relationship between the phl operon and the CHS/STS plant gene family by observing the absence of the acyl carrier protein gene.
Inactivation of phlA [162,163] and phlC [164] resulted in loss of the ability to produce DAPG. Generally, at the sequence level, components of the DAPG operon were conserved within the genus Pseudomonas [165]. However, Dash et al. [165], in their study of a strain of P. fluorescens from India, identified a unique phlB gene in the strain that differs in the nucleotide sequence from the phlB of other pseudomonads (2P24 [166], P12 [151] and Q37-87 [167,168]). Interestingly, the phlB strain from Dash et al.’s study [165] showed high affinity with the phlB strain of Pseudomonas fluorescens Q2-87 [153] in terms of the amino acid sequences.
Bangera and Thomashow [153] suggested that phlA, phlC and phlB have dual roles: they are required for the conversion of MAPG to DAPG and they participate in MAPG biosynthesis.
The phl operon is flanked downstream by phlE and upstream by phlF, phlG, and phlH genes [169]. The phlE gene product is responsible for the release of DAPG outside the cell [153,170]. More recently, the role of phlE as a permease was confirmed by Abbas et al. [171] in P. fluorescens strain F113. They also suggested that the phlE gene product was involved in the resistance of the cells themselves to DAPG. A mutant variant of strain F113 defective in the phlE gene showed greater sensitivity to osmotic, oxidative and heat-shock stresses. The authors also noted that phlE knockout led to a decrease in DAPG production because the cells became sensitive to extracellular DAPG.
PhlG was a crucial hydrolase responsible for the degradation of DAPG back to MAPG [172,173]. This process played a vital role in controlling the extracellular DAPG levels. Excessive amounts of DAPG were toxic to the cells, so when its concentration exceeded the acceptable level, a process was triggered to convert DAPG into the less toxic MAPG [174] by cleavage of a carbon–carbon bond linking an acetyl group to the phenolic ring [175]. Zhao et al. [174] demonstrated that the deletion of phlG in Pseudomonas fluorescens strain 2P24 did not affect its antifungal activity and ability to colonize plants. The authors noted that the opposite would be logical, since a variant strain with defective phlG should have more DAPG. The structure of the PhlG protein and its catalytic features have been extensively described by Saitou et al. [176] and He et al. [175].
Both phlF and phlH are tetR-like repressors. PhlF inhibited transcription of the phl operon by binding to the inverted phO repeat, which was located downstream of the phlA transcriptional start site [177,178]. PhlH inhibited transcription of the phlG gene by binding to a motif overlapping with the −35 site recognized by σ70 factors [173,179,180].
There is also phlI, but its function is still unknown [181]. Yu et al. [182] studied the effect of flavonoids on DAPG production by Pseudomonas fluorescens strain 2P24 and found that exposure to apigenin and phloretin resulted in decreased expression of the phl operon, including phlE. At the same time, there was no effect of apigenin and phloretin on the expression of phlI, which the authors suggested is cotranscribed with phlE. This suggested that phlI was not directly involved in DAPG biosynthesis [182].
5.2. Regulation of DAPG Biosynthesis
DAPG biosynthesis in pseudomonads is regulated by a number of factors:
1. By the TetR family transcriptional repressors phlF and phlH [173,177,183].
2. At the post-transcriptional level using the Gac/Rsm system [174,184].
Both of these mechanisms, as well as the co-regulation of DAPG and pioluteorin production, are described in detail in a review [185]. We will consider factors that are of a different nature.
3. PsrA
Similar to phlF, psrA is a transcriptional repressor that affects the DAPG biosynthesis genes by directly binding to a binding box localized on the phlA promoter region. Wu et al. [178], using P. fluorescens strain 2P24 as an example, revealed why two regulators, seemingly identical in their mechanism of action, are maintained in the genome. The phlF- mutant produced approximately 20-fold more DAPG than the psrA mutant; hence, regulation by phlF is dominant for DAPG production [178]. PsrA affected DAPG biosynthesis by activating the expression of the rsmA gene through the sigma factor RpoS.
4. OprF/SigX
Bangera and Thomashow [153] demonstrated that a Pseudomonas strain, which had a transposon insertion in the oprF gene, exhibited an increased quantity of red pigment of an unknown nature in comparison to the original strain. In P. fluorescens strain 2P24, the sigX gene was found to be positioned upstream of the oprF gene and acted as a positive regulator of oprF. Li et al. [186] showed that sigX did not affect the expression of the phl operon components but intensified the expression of the acetyl CoA carboxylases involved in malonyl-CoA synthesis. Malonyl-CoA served as the initial substrate in the DAPG biosynthesis pathway. The authors also revealed that the regulation of DAPG production by oprF is sigX-dependent and this process is unrelated to the Gac/Rsm system. Furthermore, the transcription of sigX was intensified by salt starvation or the addition of glycine to the medium [186].
5. Hfq
In their study, Xiao-Gang et al. [187] utilized transposon mutagenesis to discover yet another key regulator of the production of DAPG in P. fluorescens strain 2P24, the hfq gene. Inserting the transposon into the hfq gene led to a noteworthy decrease in phlA expression and the amount of DAPG in the medium. Furthermore, the hfq gene was revealed to play a vital role in the expression of pcoI synthase, a component of the quorum-sensing system in pseudomonads. As a consequence of hfq inactivation, not only was DAPG production affected, but the ability to establish biofilms and successfully colonize plants was also significantly diminished.
6. GrxD
Dong et al. [188] showed the effect of the monothiol glutaredoxin GrxD on DAPG production by P. fluorescens strain 2P24. The level of DAPG production was dramatically decreased when grxD was inactivated. Furthermore, site-directed mutagenesis of the CGFS motif in GrxD demonstrated that this motif is directly involved in the regulation of DAPG synthesis. The authors hypothesized that the positive regulation of DAPG production by GrxD was mediated by the Gac/Rsm signaling cascade. In addition, GrxD negatively affected the expression of phlF, which intensified the DAPG production in cells.
7. DsbA1
Zhang et al. [154] demonstrated the negative regulation of DAPG production by the disulfide oxidoreductase dsbA1. DsbA1 affected DAPG production not through the Gac/Rsm system but through the membrane-bound glucose dehydrogenase gcd. DsbA1 interacted with Gcd, which modulated the DAPG production. Site-directed mutagenesis of the cysteine residues in DsbA1 resulted in increased Gcd activity and intensified DAPG production.
When the gcd gene was deactivated, it led to a significant reduction of approximately 60% in DAPG production. However, in a study conducted by De Werra et al. [76], mutagenesis of gcd in P. protegens strain CHA0 resulted in the opposite outcome, with a strong accumulation of DAPG in the culture medium. Zhang et al. [154] suggested that the possible reasons for this were (1) the difference in the mechanisms of glucose catabolism in the strains, and (2) the different spectrum of antibiotics produced by the strains. Here, 2P24 produced only DAPG, while CHA0 produced DAPG, pyoluteorin (Plt), and PRN.
8. RetS
RetS is a regulator of exopolysaccharide and type III secretion [189]. Liu et al. [190] showed that RetS regulates DAPG synthesis through the Gac/Rsm signaling cascade, and the inactivation of RetS leads to increased production of DAPG and a red pigment of an unknown nature, which was also detected by Bangera and Thomashow [153] during mutagenesis of P. fluorescens strain 2P24 by the oprF gene.
He et al. [191] reported two additional genes in P. fluorescens strain HP72 that are involved in the regulation of DAPG biosynthesis. The authors designated them as ORFA and ORFB and showed that their inactivation resulted in the inability of the strain to synthesize DAPG. ORFA was related to a putative transport protein from P. putida, and ORFB—to a GNTR-like regulator (possibly phlH).
5.3. Producer Modifications
A study conducted by Zhou et al. [192] utilized Pseudomonas fluorescens strain J2 as a model to investigate the impact of phlF gene inactivation on DAPG production of almost 3-fold relative to the parental strain (3.14 mg/L vs. 1.12 mg/L). Despite this significant increase, the growth characteristics of the cultures remained largely unchanged. Similar results were obtained by Schnider-Keel et al. [180] for P. fluorescens strain CHA0, in which the amount of DAPG produced increased 4-fold upon phlF inactivation. Furthermore, Wu et al. [178] observed that P. fluorescens strain 2P24 with an inactivated phlF gene produced 5 mg/L DAPG (vs. 0.05 mg/L in the parental strain), i.e., the authors demonstrated a 100-fold enhancement of production. Notably, the authors also discovered that overexpression of psrA in inactivated phlF results in a 2-fold decrease in DAPG.
At the same time, in their study, Delany et al. [183] observed that inactivating phlF in Pseudomonas fluorescens strain F113 resulted in reduced DAPG production compared to only the parental strain during the initial stages of culture growth. DAPG was present in the mutant culture medium as early as 6 h of growth, whereas in the parental strain appeared at 12 h. The authors explained the slower growth rate of the mutant strain by the early presence of DAPG in the medium at the early stages of culture growth, and they also noted that both cultures had similar amounts of DAPG in the medium by the end of the exponential phase.
Further experimentation with Pseudomonas fluorescens strain F113 revealed that overexpression of the prrB gene encoding a regulatory RNA molecule increased the production of DAPG in the parental strain and also restored the ability to produce DAPG in mutant variants defective in the gacS (formerly lemA) and gacA genes [177,193].
Wu et al. [178] revealed that the psrA gene functions in regulating DAPG production in the strain P. fluorescens strain 2P24. PsrA bound to an operator in the promoter region of phlA and thus negatively controlled phlA function at the transcriptional level. In addition, PsrA affected the expression of RpoS and RsmA at the post-transcriptional level. Inactivation of the psrA gene resulted in a 5-fold increase in the amount of DAPG produced (250 mg/L vs. 50 mg/L). The rpoS mutant of the same strain was able to produce ~3-fold more DAPG than the parental strain (~175 mg/L vs. 50 mg/L). The data obtained were consistent with the experimental materials of Sarniguet et al. [133] for the P. protegens strain Pf-5. The parental strain P. protegens Pf-5 produced PRN (2.2 mg/L), pioluteorin (6.6 mg/L), and negligible (<0.1 mg/L) amounts of DAPG. The rpoS mutant variant lost the ability to produce PRN, but pyoluteorin was synthesized in amounts of 37.7 mg/L and DAPG in amounts of 52.2 mg/L.
Mutagenesis of the retS gene in P. protegens strain Pf-5 [156] resulted in a 20–30-fold increase in the amount of DAPG produced.
Achkar et al. [158] conducted a study to investigate the enhanced production of phloroglucinols (phloroglucinol, MAPG and DAPG) in the P. protegens strain Pf-5 transformed with the integration of the plasmid pJA2.232. The plasmid pJA2.232 was derived from pME6031 [194] and contained the phl operon as an additional component. The objective of introducing another copy of the phl operon into the strain was to bypass the regulation of DAPG synthesis by phlF. Consequently, the levels of MAPG and DAPG synthesis rose approximately 20-fold compared to the control, while phloroglucinol saw a remarkable 47-fold increase. The parental strain yielded 35 mg/L of DAPG, whereas the variant with PJA2.232 produced a significantly higher amount of 790 mg/L.
Mutagenesis of the disulfide oxidoreductase dsbA1 gene plays a crucial role in regulating DAPG production by fine-tuning the function of glucose dehydrogenase (Gcd). A recent study [154] revealed that the overexpression of the dsbA1 gene led to a significant increase in DAPG production in P. fluorescens 2P24, approximately doubling the amount from 10 mg/L to around 18 mg/L. Additionally, the authors also performed site-directed mutagenesis of the cysteine residues (C)-235, C275, and C578 of the gcd gene, which made the interaction with dsbA1 less pronounced and affected the amount of DAPG produced to some extent.
Li et al. [186], also using P. fluorescens 2P24 as an example, demonstrated the influence of two other factors, the outer membrane protein gene oprF and the sigma factor sigX, on DAPG production. SigX was located in the genome of the strain directly upstream of the oprF gene and was a positive regulator of oprF transcription. The ΔoprF mutant produced approximately 1.5-fold more DAPG than the control, and ΔsigX produced approximately 30% of the amount of DAPG produced by the parental strain.
Zhang et al. [184] and Yan et al. [173], using the strain P. fluorescens strain 2P24, demonstrated that modified (mutant) variants that produce large amounts of DAPG grow more slowly due to the increased metabolic costs in the cells.
5.4. DAPG in Biotechnology
DAPG is a broad-spectrum antibiotic that is effective against yeast (Saccharomyces [195,196], Candida [197]), fungi (Fusarium [198], Rhizoctonia solani, Sclerotium rolfsii, Macrophomina phaseolina [199], Pythium [131], Gaeumannomyces graminis [195,200], Thielaviopsis [201]), bacteria (Staphylococcus aureus [202,203], Ralstonia [199], Xanthomonas [204]), and nematodes (Meloidogyne incognita [205,206] and M. javanica [207], Globodera rostochiensis [208]).
Natural producers of DAPG include Pseudomonas protegens CL145A (ATCC 55799) [209,210], Pf-5 [211], P. fluorescens NRRL B-21806 and B-21807 [212], and P. congelans DSM 21663 [148].
The Phl gene cluster and a set of constructs using these sequences have been patented [213]. The sequence donor was P. fluorescens Q2-87, and the purpose of these constructs was twofold: (1) Pseudomonas strains lacking the ability to produce DAPG to acquire this ability, and (2) Pseudomonas strains already capable of producing DAPG to enhance their properties as producers. The modified strains were supposed to be used for plant defense against fungal pathogens.
The use of genetically modified DAPG producers in biotechnology has garnered significant attention, with various studies conducted to evaluate their impact on the biodiversity of natural microbial communities. For instance, research has shown that DAPG overproducer based on P. fluorescens F113 could positively affect mycorrhizal symbiosis and stimulate the development of micelles of the symbiotic fungus Glomus mosseae on maize roots [214]. On the other hand, it has also been demonstrated that this overproducer has no discernible effect on the indigenous microbial population in the rhizosphere of sugar beet [215].
“Phl” and “Plt” overproducers derived from P. fluorescens CHA0 impacted the composition of the microbial communities in cucumber, albeit temporary [216,217]. Niemann et al. [218] demonstrated that the addition of P. fluorescens CHA0 and its modified variant decreased the population of the nitrogen-fixing symbiotic Sinorhizobium meliloti within the soil microbiome.
Modifying Pseudomonas strains to enhance their properties as biocontrol strains is a well-known practice. Huang et al. [219] genetically modified a natural DAPG-producing strain of Pseudomonas fluorescens Q8r1-96 by integrating a plasmid containing the phz operon, which is responsible for phenazine production. This engineered strain is anticipated to be effective against Rhizoctonia, Gaeumannomyces graminis or Pythium.
Zhang et al. [29] incorporated a genomic island containing NiF (nitrogen fixation) genes into P. protegens Pf-5. The donor of the genomic island was Pseudomonas stutzeri strain DSM4166. Through markerless mutagenesis of the retS gene, the resulting Pf-5-NiF strain exhibited an increased level of DAPG production [156].
6. Conclusions
This article focuses on antibiotics produced by Pseudomonas strains, namely mupirocin, gluconic acid, PRN, and DAPG. Each section on an antibiotic covers aspects like the genetic structure of the biosynthesis pathway, the regulation of this process, and the application of the antibiotic in medical or agricultural biotechnology.
Even with the great progress in understanding the genetics of antibiotic-producing pseudomonads, a number of issues remain challenging for the future generations. For instance, it would be interesting to develop a method for the production of mupirocin consisting of only pseudomonic acid C. Due to its chemical structure, this pseudomonic acid is more stable than the others (A, B, D) and has the same antibiotic properties as PA-A. To date, attempts to obtain high yields of PA-C alone have not been successful.
Although the interest in antibiotics of Pseudomonas bacteria has persisted over the past century, many uncertainties still surround the regulation of the biosynthetic pathways of these compounds. Furthermore, the challenge of obtaining stable producers that yield higher quantities of these antibiotics continues to be a pertinent concern in modern biotechnology.
In the field of basic science, there are documented instances where producers have been successfully modified to provide high yields of the target product. However, only a few of them have been patented for biotechnological purposes. It is of interest to use modified producers in practical settings, not only in the laboratory, because in many cases genetic constructs show instability when experiments are scaled up.
Future research on antibiotics of pseudomonads can follow several main directions. First, there is biotechnological interest in improving the efficiency of antibiotic production using already-known producers and searching for new ones [123,199]. Second, there are proposals to combine antibiotic-producing strains with other useful strains, creating consortia [162]. Such work would involve studying the compatibility of strains and working out the conditions that enhance their joint productivity. Third, in both medicine and plant protection technologies, deciphering the molecular basis of pathogen sensitivity to pseudomonad antibiotics is necessary [162,220]. Since the problem of pathogen resistance to antibiotics remains relevant, special attention should be paid to the development of drug-delivery strategies, combining pseudomonad antibiotics with other agents such as anesthetics and other antibiotics [220].
This review provides a comprehensive summary of the genetic organization of the antibiotic biosynthesis pathways in Pseudomonas strains, appealing to both molecular biologists and biotechnologists.
Author Contributions
Conceptualization, Y.D., A.B. (Alexandra Baukova) and A.B. (Alexander Bogun); formal analysis, S.S., V.D.R., I.A., T.M., H.S.J., V.K. and S.M.; writing—original draft preparation, Y.D., S.S. and A.B. (Alexandra Baukova); writing—review and editing, Y.D., V.D.R., I.A., A.B. (Alexandra Baukova), S.S., T.M., A.B. (Alexander Bogun), H.S.J., V.K. and S.M.; validation, S.M. and V.K.; project administration, Y.D.; funding acquisition, S.S. and T.M. All authors have read and agreed to the published version of the manuscript.
Funding
This study was carried out in the “Soil Health” laboratory of the Southern Federal University with the financial support from the Ministry of Science and Higher Education of the Russian Federation, agreement no. 075-15-2022-1122. The research was supported by the Strategic Academic Leadership Program of the Southern Federal University (“Priority 2030”).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Last 25 Years. J. Nat. Prod. 2007, 70, 461–477. [Google Scholar] [CrossRef] [PubMed]
- Saadoun, I.; Gharaibeh, R. The Streptomyces flora of Badia region of Jordan and its potential as a source of antibiotics active against antibiotic-resistant bacteria. J. Arid Environ. 2003, 53, 365–371. [Google Scholar] [CrossRef]
- Zetola, N.; Francis, J.S.; Nuermberger, E.L.; Bishai, W.R. Community-acquired meticillin-resistant Staphylococcus aureus: An emerging threat. Lancet Infect. Dis. 2005, 5, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Haas, D.; Keel, C. Regulation of antibiotic production in root-colonizing Pseudomonas spp. and relevance for biological control of plant disease. Annu. Rev. Phytopathol. 2003, 41, 117–153. [Google Scholar] [CrossRef]
- Del Rio, L.A.; Olivares, J.; Blesa, M.C.; Mayor, F. Antibiotics from Pseudomonas reptilivora I. Taxonomic Classification and Optimal Conditions of Fermentation for Antibiotic Production. Antimicrob. Agents Chemother. 1972, 2, 186–188. [Google Scholar] [CrossRef] [PubMed]
- Castro Tapia, M.P.; Madariaga Burrows, R.P.; Ruiz Sepúlveda, B.; Vargas Concha, M.; Vera Palma, C.; Moya-Elizondo, E.A. Antagonistic Activity of Chilean Strains of Pseudomonas protegens Against Fungi Causing Crown and Root Rot of Wheat (Triticum aestivum L.). Front. Plant Sci. 2020, 11, 951. [Google Scholar] [CrossRef] [PubMed]
- Furuya, N.; Yamasaki, S.; Nishioka, M.; Shiraishi, I.; Iiyama, K.; Matsuyama, N. Antimicrobial Activities of Pseudomonads against Plant Pathogenic Organisms and Efficacy of Pseudomonas aeruginosa ATCC7700 against Bacterial Wilt of Tomato. Jpn. J. Phytopathol. 1997, 63, 417–424. [Google Scholar] [CrossRef]
- Das, K.; Abrol, S.; Verma, R.; Annapragada, H.; Katiyar, N.; Senthilkumar, M. Pseudomonas. In Beneficial Microbes in Agro-Ecology; Elsevier: Amsterdam, The Netherlands, 2020; pp. 133–148. [Google Scholar] [CrossRef]
- Arrebola, E.; Tienda, S.; Vida, C.; de Vicente, A.; Cazorla, F.M. Fitness Features Involved in the Biocontrol Interaction of Pseudomonas chlororaphis with Host Plants: The Case Study of PcPCL1606. Front. Microbiol. 2019, 10, 719. [Google Scholar] [CrossRef]
- O’Callaghan, M. Microbial inoculation of seed for improved crop performance: Issues and opportunities. Appl. Microbiol. Biotechnol. 2016, 100, 5729–5746. [Google Scholar] [CrossRef]
- Stockwell, V.O.; Johnson, K.B.; Sugar, D.; Loper, J.E. Control of Fire Blight by Pseudomonas fluorescens A506 and Pantoea vagans C9-1 Applied as Single Strains and Mixed Inocula. Phytopathology® 2010, 100, 1330–1339. [Google Scholar] [CrossRef]
- Stockwell, V.O.; Stack, J.P. Using Pseudomonas spp. for Integrated Biological Control. Phytopathology® 2007, 97, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Sahayaraj, K. (Ed.) Basic and Applied Aspects of Biopesticides; Springer: New Delhi, India, 2014. [Google Scholar] [CrossRef]
- Kumar, A. (Ed.) Microbial Biocontrol: Sustainable Agriculture and Phytopathogen Management; Springer International Publishing: Berlin/Heidelberg, Germany, 2022. [Google Scholar] [CrossRef]
- Anderson, J.A.; Staley, J.; Challender, M.; Heuton, J. Safety of Pseudomonas chlororaphis as a gene source for genetically modified crops. Transgenic Res. 2018, 27, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Osdaghi, E.; Martins, S.J.; Ramos-Sepulveda, L.; Vieira, F.R.; Pecchia, J.A.; Beyer, D.M.; Bell, T.H.; Yang, Y.; Hockett, K.L.; Bull, C.T. 100 Years Since Tolaas: Bacterial Blotch of Mushrooms in the 21st Century. Plant Dis. 2019, 103, 2714–2732. [Google Scholar] [CrossRef]
- Sajben, E.; Manczinger, L.; Nagy, A.; Kredics, L.; Vágvölgyi, C. Characterization of pseudomonads isolated from decaying sporocarps of oyster mushroom. Microbiol. Res. 2011, 166, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Manczinger, L.; Vágvölgyi, C.; Sajben, E.; Nagy, A.; Szöke-Kis, Z.; Nagy, A.; Turóczi, G.; Kovács, A. Wirkstoffe Gegen Pseudomona-Spezies als Verursacher von Fäulnisbefall bei der Pilzproduktion, Ihre Verwendung und Diese Enthaltende Zusammensetzungen. European Patent No. EP2,753,182B1, 30 August 2012. [Google Scholar]
- Fermor, T.R.; Henry, M.B.; Fenlon, J.S.; Glenister, M.J.; Lincoln, S.P.; Lynch, J.M. Development and application of a biocontrol system for bacterial blotch of the cultivated mushroom. Crop Prot. 1991, 10, 271–278. [Google Scholar] [CrossRef]
- Singh, M.; Srivastava, M.; Kumar, A.; Singh, A.K.; Pandey, K.D. Endophytic Bacteria in Plant Disease Management. In Microbial Endophytes; Elsevier: Amsterdam, The Netherlands, 2020; pp. 61–89. [Google Scholar] [CrossRef]
- Lindberg, G.D. Antibiotic Produced from the Microorganism (Pseudomonas lindbergii), Its Preparation and Method of Use. U.S. Patent No. US4,062,943, 1 February 1977. [Google Scholar]
- Farmer, S. Topical Compositions Containing Extracellular Products of Pseudomonas lindbergii and Emu oil. U.S. Patent No. US 6,645,506 B1, 26 August 1999. [Google Scholar]
- Farmer, S. Verbesserte Topische Zusammensetzungen mit Extrazellulären Produkten von Probiotischen Bakterien und Verwendungen Davon. European Patent No. EP 1 212 069 B1, 25 August 2000. [Google Scholar]
- Che, Y.; Qi, X.; Qu, W.; Shi, B.; Lin, Q.; Yao, H.; Zhang, Y.; Wei, T. Synthetic strategies of phenazine derivatives: A review. J. Heterocycl. Chem. 2022, 59, 969–996. [Google Scholar] [CrossRef]
- Yan, J.; Liu, W.; Cai, J.; Wang, Y.; Li, D.; Hua, H.; Cao, H. Advances in Phenazines over the Past Decade: Review of Their Pharmacological Activities, Mechanisms of Action, Biosynthetic Pathways and Synthetic Strategies. Mar. Drugs 2021, 19, 610. [Google Scholar] [CrossRef] [PubMed]
- Kaur, R.; Macleod, J.; Foley, W.; Nayudu, M. Gluconic acid: An antifungal agent produced by Pseudomonas species in biological control of take-all. Phytochemistry 2006, 67, 595–604. [Google Scholar] [CrossRef] [PubMed]
- Carruthers, F.L.; Shum-Thomas, T.; Conner, A.J.; Mahanty, H.K. The significance of antibiotic production by Pseudomonas aureofaciens PA 147-2 for biological control of Phytophthora megasperma root rot of asparagus. Plant Soil 1995, 170, 339–344. [Google Scholar] [CrossRef]
- Matthijs, S.; Vander Wauven, C.; Cornu, B.; Ye, L.; Cornelis, P.; Thomas, C.M.; Ongena, M. Antimicrobial properties of Pseudomonas strains producing the antibiotic mupirocin. Res. Microbiol. 2014, 165, 695–704. [Google Scholar] [CrossRef]
- Zhang, Y.; Tu, Q.; Yu, F.; Jing, X.; Bian, X.; Chen, H. Method for Screening Pseudomonas Protegens Mutant Strain, and Application Thereof in Biological Control. International Patent No. WO2018192507A1, 18 April 2018. [Google Scholar]
- Khoshnood, S.; Heidary, M.; Asadi, A.; Soleimani, S.; Motahar, M.; Savari, M.; Saki, M.; Abdi, M. A review on mechanism of action, resistance, synergism, and clinical implications of mupirocin against Staphylococcus aureus. Biomed. Pharmacother. 2019, 109, 1809–1818. [Google Scholar] [CrossRef] [PubMed]
- Troeman, D.P.R.; Van Hout, D.; Kluytmans, J.A.J.W. Antimicrobial approaches in the prevention of Staphylococcus aureus infections: A review. J. Antimicrob. Chemother. 2019, 74, 281–294. [Google Scholar] [CrossRef] [PubMed]
- Fuller, A.T.; Mellows, G.; Woolford, M.; Banks, G.T.; Barrow, K.D.; chain, e.b. Pseudomonic Acid: An Antibiotic produced by Pseudomonas fluorescens. Nature 1971, 234, 416–417. [Google Scholar] [CrossRef] [PubMed]
- Barrow, K.D.; Mellows, G. Antibiotics. U.S. Patent No. US3,977,943, 7 July 1975. [Google Scholar]
- Beecham Group PLC. The Antibiotic Pseudomonic Acid. United Kingdom Patent No. UK1,395,907, 9 June 1972.
- Class, Y.J.; DeShong, P. The Pseudomonic Acids. Chem. Rev. 1995, 95, 1843–1857. [Google Scholar] [CrossRef]
- Szell, V.; Lang, I.; Barta, I.; Tedges, A.; Albrecht, K.; Mozes, J.; Suto, N.; Szabo, I.; Petroczki, M.; Erdei, J.; et al. Process for the Preparation of Pseudomonic Acid A Antibiotic by Microbiological Method. U.S. Patent No. US20030100083A1, 12 November 2002. [Google Scholar]
- O’Donnell, J.A.; Gelone, S.P.; Safdar, A. Topical Antibacterials. In Mandell, Douglas, and Bennett’s Principles and Practice of Infectious Diseases; Elsevier: Amsterdam, The Netherlands, 2015; pp. 452–462.e2. [Google Scholar] [CrossRef]
- Hothersall, J.; Murphy, A.C.; Iqbal, Z.; Campbell, G.; Stephens, E.R.; Wu, J.; Cooper, H.; Atkinson, S.; Williams, P.; Crosby, J.; et al. Manipulation of quorum sensing regulation in Pseudomonas fluorescens NCIMB 10586 to increase mupirocin production. Appl. Microbiol. Biotechnol. 2011, 90, 1017–1026. [Google Scholar] [CrossRef] [PubMed]
- Whatling, C.A.; Hodgson, J.E.; Burnham, M.K.R.; Clarke, N.J.; Franklin, F.C.H.; Thomas, C.M. Identification of a 60 kb region of the chromosome of Pseudomonas fluorescens NCIB 10586 required for the biosynthesis of pseudomonic acid (mupirocin). Microbiology 1995, 141, 973–982. [Google Scholar] [CrossRef][Green Version]
- Tucaliuc, A.; Blaga, A.C.; Galaction, A.I.; Cascaval, D. Mupirocin: Applications and production. Biotechnol. Lett. 2019, 41, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.-S.; Hothersall, J.; Wu, J.; Murphy, A.C.; Song, Z.; Stephens, E.R.; Thomas, C.M.; Crump, M.P.; Cox, R.J.; Simpson, T.J.; et al. Biosynthesis of Mupirocin by Pseudomonas fluorescens NCIMB 10586 Involves Parallel Pathways. J. Am. Chem. Soc. 2014, 136, 5501–5507. [Google Scholar] [CrossRef]
- Fritz, E.; Fekete, A.; Lintelmann, J.; Schmitt-Kopplin, P.; Meckenstock, R.U. Isolation of two Pseudomonas strains producing pseudomonic acid A. Syst. Appl. Microbiol. 2009, 32, 56–64. [Google Scholar] [CrossRef]
- Haines, A.S.; Kendrew, S.G.; Crowhurst, N.; Stephens, E.R.; Connolly, J.; Hothersall, J.; Miller, C.E.; Collis, A.J.; Huckle, B.D.; Thomas, C.M. High quality genome annotation and expression visualisation of a mupirocin-producing bacterium. PLoS ONE 2022, 17, e0268072. [Google Scholar] [CrossRef]
- Hothersall, J.; Wu, J.; Rahman, A.S.; Shields, J.A.; Haddock, J.; Johnson, N.; Cooper, S.M.; Stephens, E.R.; Cox, R.J.; Crosby, J.; et al. Mutational Analysis Reveals That All Tailoring Region Genes Are Required for Production of Polyketide Antibiotic Mupirocin by Pseudomonas fluorescens. J. Biol. Chem. 2007, 282, 15451–15461. [Google Scholar] [CrossRef]
- Cooper, S.M.; Laosripaiboon, W.; Rahman, A.S.; Hothersall, J.; El-Sayed, A.K.; Winfield, C.; Crosby, J.; Cox, R.J.; Simpson, T.J.; Thomas, C.M. Shift to Pseudomonic Acid B Production in P. fluorescens NCIMB10586 by Mutation of Mupirocin Tailoring Genes mupO, mupU, mupV, and macpE. Chem. Biol. 2005, 12, 825–833. [Google Scholar] [CrossRef]
- Caldwell, C.J.; Hynes, R.K.; Boyetchko, S.M.; Korber, D.R. Colonization and bioherbicidal activity on green foxtail by Pseudomonas fluorescens BRG100 in a pesta formulation. Can. J. Microbiol. 2012, 58, 1–9. [Google Scholar] [CrossRef]
- Dumonceaux, T.J.; Town, J.; Links, M.G.; Boyetchko, S. High-Quality Draft Genome Sequence of Pseudomonas sp. BRG100, a Strain with Bioherbicidal Properties against Setaria viridis (Green Foxtail) and Other Pests of Agricultural Signifi-cance. Genome Announc. 2014, 2, e00995-14. [Google Scholar] [CrossRef]
- Barrish, J.C.; Lee, H.L.; Mitt, T.; Pizzolato, G.; Baggiolini, E.G.; Uskokovic, M.R. Total synthesis of pseudomonic acid C. J. Org. Chem. 1988, 53, 4282–4295. [Google Scholar] [CrossRef]
- Lazar, R.G.; Blaga, A.C.; Dragoi, E.N.; Galaction, A.I.; Cascaval, D. Application of reactive extraction for the separation of pseudomonic acids: Influencing factors, interfacial mechanism, and process modelling. Can. J. Chem. Eng. 2022, 100, S246–S257. [Google Scholar] [CrossRef]
- Snider, B.B.; Phillips, G.B. Total synthesis of (+-)-pseudomonic acids A and C. J. Am. Chem. Soc. 1982, 104, 1113–1114. [Google Scholar] [CrossRef]
- Sridhar, Y.; Srihari, P. A unified strategy for the synthesis of the C1–C14 fragment of marinolic acids, mupirocins, pseudomonic acids and thiomarinols: Total synthesis of pseudomonic acid methyl monate C. Org. Biomol. Chem. 2014, 12, 2950. [Google Scholar] [CrossRef]
- El-Sayed, A.K.; Hothersall, J.; Thomas, C.M. Quorum-sensing-dependent regulation of biosynthesis of the polyketide antibiotic mupirocin in Pseudomonas fluorescens NCIMB 10586 The GenBank accession numbers for the sequences determined in this work are AF318063 (mupA), AF318064 (mupR) and AF318065 (mupI). Microbiology 2001, 147, 2127–2139. [Google Scholar] [CrossRef] [PubMed]
- Macioszek, M. Biosynthesis of Mupirocin by Pseudomonas fluorescens NCIMB 10586. Ph.D. Thesis, The University of Birmingham, Birmingham, UK, 2009. [Google Scholar]
- Dwivedi, D.; Johri, B.N. Antifungals from fluorescent pseudomonads: Biosynthesis and regulation. Curr. Sci. 2003, 85, 1693–1703. [Google Scholar]
- Gurney, R. Biosynthesis of the Antibiotic Mupirocin by Pseudomonas fluorescens NCIMB 10586. Ph.D. Thesis, The University of Birmingham, Birmingham, UK, 2012. [Google Scholar]
- Gulyas, E.; Balogh, G.; Erdei, J.; Seress, P. PH Controlled Fermentation Process for Pseudomonic Acid Production. U.S. Patent No. 7439,045 B2, 21 June 2002. [Google Scholar]
- Conly, J.M.; Johnston, B.L. Mupirocin—Are We in Danger of Losing It? Can. J. Infect. Dis. 2002, 13, 157–159. [Google Scholar] [CrossRef]
- Cern, A.; Connolly, K.L.; Jerse, A.E.; Barenholz, Y. In Vitro Susceptibility of Neisseria gonorrhoeae Strains to Mupirocin, an Antibiotic Reformulated for Parenteral Administration in Nanoliposomes. Antimicrob. Agents Chemother. 2018, 62, e02377-17. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.; Mellows, G. Inhibition of isoleucyl-transfer ribonucleic acid synthetase in Echerichia coli by pseudomonic acid. Biochem. J. 1978, 176, 305–318. [Google Scholar] [CrossRef]
- Lamb, Y.J. Overview of the role of mupirocin. J. Hosp. Infect. 1991, 19, 27–30. [Google Scholar] [CrossRef]
- Parenti, M.A.; Hatfield, S.M.; Leyden, J.J. Mupirocin: A topical antibiotic with a unique structure and mechanism of action. Clin. Pharm. 1987, 6, 761–770. [Google Scholar]
- Savage, P.; Milner, S.M. Expanding the spectrum of activity of mupirocin to include gram-negative bacteria using cationic steroid antibiotics. J. Am. Acad. Dermatol. 2005, 52, P7. [Google Scholar] [CrossRef]
- Oduro-Yeboah, J. Pharmaceutical Formulations Containing Pseudomonic Acid. U.S. Patent No. 4,524,075, 26 May 1983. [Google Scholar]
- Hunter, P.; Berry, V.; Oduro-Yeboah, J.; Orr, N. Treatment of Fungal Infections. U.S. Patent No. 4,790,989, 10 April 1987. [Google Scholar]
- Zimmerman, H.L. Pharmaceutical and Veterinary Compositions of Mupirocin and Methods for Their Preparation. U.S. Patent No. 6,025,389, 25 July 1997. [Google Scholar]
- Lavon, I.; Zeevi, A.; Cherkez, S.; Arkin, M.; Abu-Gnim, C.; Raechav, Y.; Kaspi, J. Pharmaceutical Compositions Con-Taining Amorphous Mupirocin. European Patent Application No. 1 174 133 A1, 23 January 2002. [Google Scholar]
- Curzons, A.D. Lithium Pseudomonate and Its Preparation. Canada Patent No. CA1,115,699A, 4 May 1979. [Google Scholar]
- Curzons, A.D. Crystalline Lithium Pseudomonete. U.S. Patent No. US4,786,742, 8 August 1986. [Google Scholar]
- Baker, G.H.; Beal, M. Crystalline Calcium Pseudomonate. U.S. Patent No. US4,916,155, 13 April 1989. [Google Scholar]
- Baker, G.H.; Beal, M. Process for Preparing Crystalline Calcium Pseudomonate. U.S. Patent No. US5,191,093, 12 March 1990. [Google Scholar]
- Baker, G.H.; Beal, M. Compounds. U.S. Patent No. US5,436,266, 28 February 1994. [Google Scholar]
- Orr, N.A.; Greenway, M.J. Pharmaceutical Composition. U.S. Patent No. US4,879,287, 21 December 1988. [Google Scholar]
- Weisman, A.; Kaspi, J.; Cherkez, S. Stable Amorphous Calcium Pseudomonate and Processes for the Preparation Thereof. European Patent No. EP 1 384 721 A1, 23 July 2003. [Google Scholar]
- Pappa, K.A. The clinical development of mupirocin. J. Am. Acad. Dermatol. 1990, 22, 873–879. [Google Scholar] [CrossRef]
- Bull, D.N.; Kempe, L.L. Kinetics of the conversion of glucose to gluconic acid by Pseudomonas ovalis. Biotechnol. Bioeng. 1970, 12, 273–290. [Google Scholar] [CrossRef][Green Version]
- de Werra, P.; Péchy-Tarr, M.; Keel, C.; Maurhofer, M. Role of Gluconic Acid Production in the Regulation of Biocontrol Traits of Pseudomonas fluorescens CHA0. Appl. Environ. Microbiol. 2009, 75, 4162–4174. [Google Scholar] [CrossRef] [PubMed]
- Galet, J.; Deveau, A.; Hôtel, L.; Leblond, P.; Frey-Klett, P.; Aigle, B. Gluconic acid-producing Pseudomonas sp. prevent γ-actinorhodin biosynthesis by Streptomyces coelicolor A3(2). Arch. Microbiol. 2014, 196, 619–627. [Google Scholar] [CrossRef][Green Version]
- Kaur, R.; Nayudu, M. A Method of Controlling Fungal Pathogens, and Agents Useful for Same. U.S. Patent No. US 2007/0274973 A1, 7 August 2006. [Google Scholar]
- Yu, K.; Liu, Y.; Tichelaar, R.; Savant, N.; Lagendijk, E.; van Kuijk, S.J.L.; Stringlis, I.A.; van Dijken, A.J.H.; Pieterse, C.M.J.; Bakker, P.A.H.M.; et al. Rhizosphere-Associated Pseudomonas Suppress Local Root Immune Responses by Gluconic Acid-Mediated Lowering of Environmental pH. Curr. Biol. 2019, 29, 3913–3920.e4. [Google Scholar] [CrossRef] [PubMed]
- Whiting, P.H.; Midgley, M.; Dawes, E.A. The regulation of transport of glucose, gluconate and 2-oxogluconate and of glucose catabolism in Pseudomonas aeruginosa. Biochem. J. 1976, 154, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, A.H. Recent Progress in Understanding the Molecular Genetics and Biochemistry of Calcium Phosphate Solubilization by Gram Negative Bacteria. Biol. Agric. Hortic. 1995, 12, 185–193. [Google Scholar] [CrossRef]
- Ghose, T.K.; Ghosh, P. Kinetic analysis of gluconic acid production by Pseudomonas ovalis. J. Appl. Chem. Biotechnol. 2007, 26, 768–777. [Google Scholar] [CrossRef]
- Constantinides, A.; Rai, V.R. Mathematical Modeling and Optimization of Gluconic Acid Fermentation. AIChE Symp. Ser. 1973, 69, 114. [Google Scholar]
- Nyiri, L.K.; Toth, G.M. Space Biosynthesis Systems; Report No. 102110176FD. 1976. Available online: https://ntrs.nasa.gov/api/citations/19770009736/downloads/19770009736.pdf (accessed on 20 April 2024).
- Illmer, P.; Schinner, F. Solubilization of inorganic phosphates by microorganisms isolated from forest soils. Soil Biol. Biochem. 1992, 24, 389–395. [Google Scholar] [CrossRef]
- Di Simine, C.D.; Sayer, J.A.; Gadd, G.M. Solubilization of zinc phosphate by a strain of Pseudomonas fluorescens isolated from a forest soil. Biol. Fertil. Soils 1998, 28, 87–94. [Google Scholar] [CrossRef]
- Ramachandran, S.; Fontanille, P.; Pandey, A.; Larroche, C. Gluconic Acid: Properties, Applications and Microbial Production. Food Technol. Biotechnol. 2006, 44, 185–195. [Google Scholar]
- Trivedi, P.; Sa, T. Pseudomonas corrugata (NRRL B-30409) Mutants Increased Phosphate Solubilization, Organic Acid Production, and Plant Growth at Lower Temperatures. Curr. Microbiol. 2008, 56, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Zia, M.A.; Khalil-ur-Rahman; Saeed, M.K.; Andaleeb, F.; Rajoka, M.I.; Sheikh, M.A.; Khan, I.A.; Khan, A.I. Thermal Characterization of Purified Glucose Oxidase from A Newly Isolated Aspergillus Niger UAF-1. J. Clin. Biochem. Nutr. 2007, 41, 132–138. [Google Scholar] [CrossRef]
- Frederick, K.R.; Tung, J.; Emerick, R.S.; Masiarz, F.R.; Chamberlain, S.H.; Vasavada, A.; Rosenberg, S.; Chakraborty, S.; Schopfer, L.M.; Schopter, L.M. Glucose oxidase from Aspergillus niger. Cloning, gene sequence, secretion from Saccharomyces cerevisiae and kinetic analysis of a yeast-derived enzyme. J. Biol. Chem. 1990, 265, 3793–3802. [Google Scholar] [CrossRef]
- Dokter, P.; Frank, J.; Duine, J.A. Purification and characterization of quinoprotein glucose dehydrogenase from Acinetobacter calcoaceticus L.M.D. 79.41. Biochem. J. 1986, 239, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Pronk, J.T.; van Schie, B.J.; van Dijken, J.P.; Kuenen, J.G. Energization of solute transport by PQQ-dependent glucose dehydrogenase in membrane vesicles of Acinetobacter species. Antonie Van Leeuwenhoek 1985, 51, 560. [Google Scholar] [CrossRef]
- Schie, B.J.; Dijken, J.P.; Kuenen, J.G. Non-coordinated synthesis of glucose dehydrogenase and its prosthetic group PQQ in Acinetobacter and Pseudomonas species. FEMS Microbiol. Lett. 1984, 24, 133–138. [Google Scholar] [CrossRef]
- Velizarov, S.; Beschkov, V. Production of free gluconic acid by cells of Gluconobacter oxydans. Biotechnol. Lett. 1994, 16, 715–720. [Google Scholar] [CrossRef]
- Khan, S.; Somerville, D.; Frese, M.; Nayudu, M. Environmental gut bacteria in European honey bees (Apis mellifera) from Australia and their relationship to the chalkbrood disease. PLoS ONE 2020, 15, e0238252. [Google Scholar] [CrossRef]
- Kremmydas, G.F.; Tampakaki, A.P.; Georgakopoulos, D.G. Characterization of the Biocontrol Activity of Pseudomonas fluorescens Strain X Reveals Novel Genes Regulated by Glucose. PLoS ONE 2013, 8, e61808. [Google Scholar] [CrossRef] [PubMed]
- An, R.; Moe, L.A. Regulation of Pyrroloquinoline Quinone-Dependent Glucose Dehydrogenase Activity in the Model Rhizosphere-Dwelling Bacterium Pseudomonas putida KT2440. Appl. Environ. Microbiol. 2016, 82, 4955–4964. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Ran, T.; Ma, C.; He, J.; Xu, D.; Wang, W. Crystal Structure and Function of PqqF Protein in the Pyrroloquinoline Quinone Biosynthetic Pathway. J. Biol. Chem. 2016, 291, 15575–15587. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.-Q.; Bonnot, F.; Imsand, E.M.; RoseFigura, J.M.; Sjölander, K.; Klinman, J.P. Distribution and Properties of the Genes Encoding the Biosynthesis of the Bacterial Cofactor, Pyrroloquinoline Quinone. Biochemistry 2012, 51, 2265–2275. [Google Scholar] [CrossRef]
- Morales, G.; Ugidos, A.; Rojo, F. Inactivation of the Pseudomonas putida cytochrome o ubiquinol oxidase leads to a significant change in the transcriptome and to increased expression of the CIO and cbb3-1 terminal oxidases. Environ. Microbiol. 2006, 8, 1764–1774. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.H.; Browne, P.; Prigent-Combaret, C.; Combes-Meynet, E.; Morrissey, J.P.; O’Gara, F. Biochemical and genomic comparison of inorganic phosphate solubilization in Pseudomonas species. Environ. Microbiol. Rep. 2010, 2, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Foster, P.L. In Vivo Mutagenesis; Elsevier: Amsterdam, The Netherlands, 1991; pp. 114–125. [Google Scholar] [CrossRef]
- Brian, P.; Riggle, P.J.; Santos, R.A.; Champness, W.C. Global negative regulation of Streptomyces coelicolor antibiotic synthesis mediated by an absA-encoded putative signal transduction system. J. Bacteriol. 1996, 178, 3221–3231. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Magnolo, S.K.; Leenutaphong, D.L.; DeModena, J.A.; Curtis, J.E.; Bailey, J.E.; Galazzo, J.L.; Hughes, D.E. Actinorhodin Production by Streptomyces coelicolor and Growth of Streptomyces lividans Are Improved by the Expression of a Bacterial Hemoglobin. Nat. Biotechnol. 1991, 9, 473–476. [Google Scholar] [CrossRef] [PubMed]
- Thompson, I.P.; Lilley, A.K.; Ellis, R.J.; Bramwell, P.A.; Bailey, M.J. Survival, Colonization and Dispersal of Genetically Modified Pseudomonas fluorescens SBW25 in the Phytosphere of Field Grown Sugar Beet. Nat. Biotechnol. 1995, 13, 1493–1497. [Google Scholar] [CrossRef]
- Compeau, G.; Al-Achi, B.J.; Platsouka, E.; Levy, S.B. Survival of rifampin-resistant mutants of Pseudomonas fluorescens and Pseudomonas putida in soil systems. Appl. Environ. Microbiol. 1988, 54, 2432–2438. [Google Scholar] [CrossRef] [PubMed]
- Ramette, A.; Frapolli, M.; Saux, M.F.-L.; Gruffaz, C.; Meyer, J.-M.; Défago, G.; Sutra, L.; Moënne-Loccoz, Y. Pseudomonas protegens sp. nov.; widespread plant-protecting bacteria producing the biocontrol compounds 2,4-diacetylphloroglucinol and pyoluteorin. Syst. Appl. Microbiol. 2011, 34, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Holloway, B.W.; Romling, U.; Tummler, B. Genomic mapping of Pseudomonas aeruginosa PAO. Microbiology 1994, 140, 2907–2929. [Google Scholar] [CrossRef] [PubMed]
- Ponraj, P.; Shankar, M.; Ilakkiam, D.; Rajendhran, J.; Gunasekaran, P. Influence of periplasmic oxidation of glucose on pyoverdine synthesis in Pseudomonas putida S11. Appl. Microbiol. Biotechnol. 2013, 97, 5027–5041. [Google Scholar] [CrossRef]
- Vyas, P.; Gulati, A. Organic acid production in vitro and plant growth promotion in maize under controlled environment by phosphate-solubilizing fluorescent Pseudomonas. BMC Microbiol. 2009, 9, 174. [Google Scholar] [CrossRef]
- Chernin, L.; Brandis, A.; Ismailov, Z.; Chet, I. Pyrrolnitrin Production by an Enterobacter agglomerans Strain with a Broad Spectrum of Antagonistic Activity Towards Fungal and Bacterial Phytopathogens. Curr. Microbiol. 1996, 32, 208–212. [Google Scholar] [CrossRef]
- Arima, K.; Imanaka, H.; Kousaka, M.; Fukuta, A.; Tamura, G. Pyrrolnitrin, a New Antibiotic Substance, Produced by Pseudomonas. Agric. Biol. Chem. 1964, 28, 575–576. [Google Scholar] [CrossRef]
- Hammer, P.E.; Hill, D.S.; Lam, S.T.; Van Pée, K.H.; Ligon, J.M. Four genes from Pseudomonas fluorescens that encode the biosynthesis of pyrrolnitrin. Appl. Environ. Microbiol. 1997, 63, 2147–2154. [Google Scholar] [CrossRef] [PubMed]
- Kirner, S.; Hammer, P.E.; Hill, D.S.; Altmann, A.; Fischer, I.; Weislo, L.J.; Lanahan, M.; van Pée, K.-H.; Ligon, J.M. Functions Encoded by Pyrrolnitrin Biosynthetic Genes from Pseudomonas fluorescens. J. Bacteriol. 1998, 180, 1939–1943. [Google Scholar] [CrossRef] [PubMed]
- Kirner, S.; Krauss, S.; Sury, G.; Lam, S.T.; Ligon, J.M.; van Pee, K.-H. The non-haem chloroperoxidase from Pseudomonas fluorescens and its relationship to pyrrolnitrin biosynthesis. Microbiology 1996, 142, 2129–2135. [Google Scholar] [CrossRef]
- Hammer, P.E.; Burd, W.; Hill, D.S.; Ligon, J.M.; Pée, K.-H. Conservation of the pyrrolnitrin biosynthetic gene cluster among six pyrrolnitrin-producing strains. FEMS Microbiol. Lett. 1999, 180, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Feng, Z.; Chi, X.; Sun, X.; Lu, Y.; Zhang, B.; Lu, R.; Luo, W.; Wang, Y.; Miao, J.; et al. Pyrrolnitrin is more essential than phenazines for Pseudomonas chlororaphis G05 in its suppression of Fusarium graminearum. Microbiol. Res. 2018, 215, 55–64. [Google Scholar] [CrossRef]
- Nandi, M.; Selin, C.; Brassinga, A.K.C.; Belmonte, M.F.; Fernando, W.G.D.; Loewen, P.C.; de Kievit, T.R. Pyrrolnitrin and Hydrogen Cyanide Production by Pseudomonas chlororaphis Strain PA23 Exhibits Nematicidal and Repellent Activity against Caenorhabditis elegans. PLoS ONE 2015, 10, e0123184. [Google Scholar] [CrossRef] [PubMed]
- Park, J.Y.; Oh, S.A.; Anderson, A.J.; Neiswender, J.; Kim, J.-C.; Kim, Y.C. Production of the antifungal compounds phenazine and pyrrolnitrin from Pseudomonas chlororaphis O6 is differentially regulated by glucose. Lett. Appl. Microbiol. 2011, 52, 532–537. [Google Scholar] [CrossRef]
- Cartwright, D.K.; Chilton, W.S.; Benson, D.M. Pyrrolnitrin and phenazine production by Pseudomonas cepacia, strain 5.5B, a biocontrol agent of Rhizoctonia solani. Appl. Microbiol. Biotechnol. 1995, 43, 211–216. [Google Scholar] [CrossRef]
- Lively, D.H.; Gorman, M.; Haney, M.E.; Mabe, J.A. Metabolism of tryptophans by Pseudomonas aureofaciens. I. Biosynthesis of pyrrolnitrin. Antimicrob. Agents Chemother. 1966, 6, 462–469. [Google Scholar]
- Hamill, R.; Elander, R.; Mabe, J.; Gorman, M. Metabolism of tryptophans by Pseudomonas aureofaciens. V. Conversion of tryptophan to pyrrolnitrin. Antimicrob. Agents Chemother. 1967, 7, 388–396. [Google Scholar]
- Pawar, S.; Chaudhari, A.; Prabha, R.; Shukla, R.; Singh, D.P. Microbial Pyrrolnitrin: Natural Metabolite with Immense Practical Utility. Biomolecules 2019, 9, 443. [Google Scholar] [CrossRef]
- De Laurentis, W.; Khim, L.; Anderson, J.L.R.; Adam, A.; Phillips, R.S.; Chapman, S.K.; van Pee, K.-H.; Naismith, J.H. The Second Enzyme in Pyrrolnitrin Biosynthetic Pathway Is Related to the Heme-Dependent Dioxygenase Superfamily. Biochemistry 2007, 46, 12393–12404. [Google Scholar] [CrossRef]
- van Pée, K.-H.; Salcher, O.; Lingens, F. Formation of Pyrrolnitrin and 3-(2-Amino-3-chlorophenyl)pyrrole from 7-Chlorotryptophan. Angew. Chem. Int. Ed. Engl. 1980, 19, 828–829. [Google Scholar] [CrossRef]
- Chang, C.J.; Floss, H.G.; Hook, D.J.; Mabe, J.A.; Manni, P.E.; Martin, L.L.; Schröder, K.; Shieh, T.L. The biosynthesis of the antibiotic pyrrolnitrin by Pseudomonas aureofaciens. J. Antibiot. 1981, 34, 555–566. [Google Scholar] [CrossRef]
- van Pée, K.-H. Biosynthesis of halogenated metabolites by bacteria. Annu. Rev. Microbiol. 1996, 50, 375–399. [Google Scholar] [CrossRef]
- Wiesner, W.; van Pée, K.H.; Lingens, F. Purification and characterization of a novel bacterial non-heme chloroperoxidase from Pseudomonas pyrrocinia. J. Biol. Chem. 1988, 263, 13725–13732. [Google Scholar] [CrossRef]
- Keller, S.; Wage, T.; Hohaus, K.; Hölzer, M.; Eichhorn, E.; van Pée, K.-H. Purification and Partial Characterization of Tryptophan 7-Halogenase (PrnA) from Pseudomonas fluorescens. Angew. Chem. Int. Ed. 2000, 39, 2300–2302. [Google Scholar] [CrossRef]
- Singh, R.N.; Singh, R.P.; Sharma, A.; Saxena, A.K. Modeling of PrnD protein from Pseudomonas fluorescens RajNB11 and its comparative structural analysis with PrnD proteins expressed in Burkholderia and Serratia. Turk. J. Biol. 2016, 40, 623–633. [Google Scholar] [CrossRef]
- de Souza, J.T.; Arnould, C.; Deulvot, C.; Lemanceau, P.; Gianinazzi-Pearson, V.; Raaijmakers, J.M. Effect of 2,4-Diacetylphloroglucinol on Pythium: Cellular Responses and Variation in Sensitivity Among Propagules and Species. Phytopathology 2003, 93, 966–975. [Google Scholar] [CrossRef]
- Laville, J.; Voisard, C.; Keel, C.; Maurhofer, M.; Défago, G.; Haas, D. Global control in Pseudomonas fluorescens mediating antibiotic synthesis and suppression of black root rot of tobacco. Proc. Natl. Acad. Sci. USA 1992, 89, 1562–1566. [Google Scholar] [CrossRef]
- Sarniguet, A.; Kraus, J.; Henkels, M.D.; Muehlchen, A.M.; Loper, J.E. The sigma factor sigma s affects antibiotic production and biological control activity of Pseudomonas fluorescens Pf-5. Proc. Natl. Acad. Sci. USA 1995, 92, 12255–12259. [Google Scholar] [CrossRef]
- Pfender, W.F. A Genomic Region from Pseudomonas fluorescens Pf-5 Required for Pyrrolnitrin Production and Inhibition of Pyrenophora tritici-repentis in Wheat Straw. Phytopathology 1993, 83, 1223. [Google Scholar] [CrossRef]
- Manuel, J.; Selin, C.; Fernando, W.G.D.; de Kievit, T. Stringent response mutants of Pseudomonas chlororaphis PA23 exhibit enhanced antifungal activity against Sclerotinia sclerotiorum in vitro. Microbiology 2012, 158, 207–216. [Google Scholar] [CrossRef]
- Wu, X.; Chi, X.; Wang, Y.; Zhang, K.; Kai, L.; He, Q.; Tang, J.; Wang, K.; Sun, L.; Hao, X.; et al. vfr, A Global Regulatory Gene, is Required for Pyrrolnitrin but not for Phenazine-1-carboxylic Acid Biosynthesis in Pseudomonas chlororaphis G05. Plant Pathol. J. 2019, 35, 351–361. [Google Scholar] [CrossRef]
- West, S.E.; Sample, A.K.; Runyen-Janecky, L.J. The vfr gene product, required for Pseudomonas aeruginosa exotoxin A and protease production, belongs to the cyclic AMP receptor protein family. J. Bacteriol. 1994, 176, 7532–7542. [Google Scholar] [CrossRef][Green Version]
- Ogura, K.; Matsui, H.; Yamamoto, M.; Noutoshi, Y.; Toyoda, K.; Taguchi, F.; Ichinose, Y. Vfr targets promoter of genes encoding methyl-accepting chemotaxis protein in Pseudomonas syringae pv. tabaci 6605. Biochem. Biophys. Rep. 2021, 26, 100944. [Google Scholar] [CrossRef]
- Fuchs, E.L.; Brutinel, E.D.; Jones, A.K.; Fulcher, N.B.; Urbanowski, M.L.; Yahr, T.L.; Wolfgang, M.C. The Pseudomonas aeruginosa Vfr regulator controls global virulence factor expression through cyclic AMP-dependent and -independent mechanisms. J. Bacteriol. 2010, 192, 3553–3564. [Google Scholar] [CrossRef]
- Keum, Y.S.; Lee, H.R.; Kim, J.-H. Effects of Pesticides on the Bacterial Production of Pyrrolnitrin. J. Agric. Food Chem. 2010, 58, 5531–5537. [Google Scholar] [CrossRef]
- Salcher, O.; Lingens, F. Isolation and Characterization of a Mutant of Pseudomonas aureofaciens ATCC 15926 with an Increased Capacity for Synthesis of Pyrrolnitrin. Microbiology 1980, 118, 509–513. [Google Scholar] [CrossRef]
- Auden, J.; Gruner, J.; Nüesch, J.; Knüsel, F. Some Statistical Methods in Nutrient Medium Optimalisation. Pathobiology 1967, 30, 858–866. [Google Scholar] [CrossRef]
- Olson, E.S.; Richards, J.H. Structure of the orange pigment from Pseudomonas aureofaciens. J. Org. Chem. 1967, 32, 2887–2890. [Google Scholar] [CrossRef]
- Zhang, J.; Mavrodi, D.V.; Yang, M.; Thomashow, L.S.; Mavrodi, O.V.; Kelton, J.; Weller, D.M. Pseudomonas synxantha 2-79 Transformed with Pyrrolnitrin Biosynthesis Genes Has Improved Biocontrol Activity Against Soilborne Pathogens of Wheat and Canola. Phytopathology 2020, 110, 1010–1017. [Google Scholar] [CrossRef]
- Loper, J.E.; Hassan, K.A.; Mavrodi, D.V.; Davis, E.W.; Lim, C.K.; Shaffer, B.T.; Elbourne, L.D.H.; Stockwell, V.O.; Hartney, S.L.; Breakwell, K.; et al. Comparative Genomics of Plant-Associated Pseudomonas spp.: Insights into Diversity and Inheritance of Traits Involved in Multitrophic Interactions. PLoS Genet. 2012, 8, e1002784. [Google Scholar] [CrossRef]
- Ligon, J.; Hill, D.; Lam, S.; Gaffney, T.; Torkewitz, N. Genetically Modified Pseudomonas Strans with Enhanced Biocontrol Activity. U.S. Patent No. US 5,756,087, 6 December 1996. [Google Scholar]
- Janisiewicz, W.; Roitman, J. Biological Control of Postharvest Rots in Fruits Using Pseudomonas cepacia and Pyrrolnitrin Produced Therefrom. U.S. Patent No. US 4,975,277, 2 August 1988. [Google Scholar]
- Gnanamanickam, S. Pseudomonas Bacterium. U.S. Patent Application No. US 2010/0093538A1, 29 September 2009. [Google Scholar]
- Roberts, D.P.; McKenna, L.F.; Lakshman, D.K.; Meyer, S.L.F.; Kong, H.; de Souza, J.T.; Lydon, J.; Baker, C.J.; Buyer, J.S.; Chung, S. Suppression of damping-off of cucumber caused by Pythium ultimum with live cells and extracts of Serratia marcescens N4-5. Soil Biol. Biochem. 2007, 39, 2275–2288. [Google Scholar] [CrossRef]
- Envall, M. On the difference between mono-, holo-, and paraphyletic groups: A consistent distinction of process and pattern. Biol. J. Linn. Soc. 2008, 94, 217–220. [Google Scholar] [CrossRef]
- Almario, J.; Bruto, M.; Vacheron, J.; Prigent-Combaret, C.; Moënne-Loccoz, Y.; Muller, D. Distribution of 2,4-Diacetylphloroglucinol Biosynthetic Genes among the Pseudomonas spp. Reveals Unexpected Polyphyletism. Front. Microbiol. 2017, 8, 1218. [Google Scholar] [CrossRef]
- Shanahan, P.; Glennon, J.D.; Crowley, J.J.; Donnelly, D.F.; O’Gara, F. Liquid chromatographic assay of microbially derived phloroglucinol antibiotics for establishing the biosynthetic route to production, and the factors affecting their regulation. Anal. Chim. Acta 1993, 272, 271–277. [Google Scholar] [CrossRef]
- Bangera, M.G.; Thomashow, L.S. Identification and Characterization of a Gene Cluster for Synthesis of the Polyketide Antibiotic 2,4-Diacetylphloroglucinol from Pseudomonas fluorescens Q2-87. J. Bacteriol. 1999, 181, 3155–3163. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zhao, H.; Wu, X.; Zhang, L.-Q. The Oxidoreductase DsbA1 negatively influences 2,4-diacetylphloroglucinol biosynthesis by interfering the function of Gcd in Pseudomonas fluorescens 2P24. BMC Microbiol. 2020, 20, 39. [Google Scholar] [CrossRef] [PubMed]
- Nowak-Thompson, B.; Gould, S.J.; Kraus, J.; Loper, J.E. Production of 2,4-diacetylphloroglucinol by the biocontrol agent Pseudomonas fluorescens Pf-5. Can. J. Microbiol. 1994, 40, 1064–1066. [Google Scholar] [CrossRef]
- Jing, X.; Cui, Q.; Li, X.; Yin, J.; Ravichandran, V.; Pan, D.; Fu, J.; Tu, Q.; Wang, H.; Bian, X.; et al. Engineering Pseudomonas protegens Pf-5 to improve its antifungal activity and nitrogen fixation. Microb. Biotechnol. 2020, 13, 118–133. [Google Scholar] [CrossRef] [PubMed]
- Shanahan, P.; O’Sullivan, D.J.; Simpson, P.; Glennon, J.D.; O’Gara, F. Isolation of 2,4-Diacetylphloroglucinol from a Fluorescent Pseudomonad and Investigation of Physiological Parameters Influencing Its Production. Appl. Environ. Microbiol. 1992, 58, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Achkar, J.; Xian, M.; Zhao, H.; Frost, J.W. Biosynthesis of Phloroglucinol. J. Am. Chem. Soc. 2005, 127, 5332–5333. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, C.R.; Fujii, I. Polyketide synthase gene manipulation: A Structure-Function Approach in Engineering Novel Antibiotics. Annu. Rev. Microbiol. 1995, 49, 201–238. [Google Scholar] [CrossRef]
- McDaniel, R.; Ebert-Khosla, S.; Hopwood, D.A.; Khosla, C. Engineered Biosynthesis of Novel Polyketides. Science 1993, 262, 1546–1550. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Dash, P.K.; Sanjay, T.D.; Pradhan, S.K.; Sreevathsa, R.; Rai, R. Cloning and Molecular Characterization of the phlD Gene Involved in the Biosynthesis of “Phloroglucinol”, a Compound with Antibiotic Properties from Plant Growth Promoting Bacteria Pseudomonas spp. Antibiotics 2023, 12, 260. [Google Scholar] [CrossRef]
- Balthazar, C.; St-Onge, R.; Léger, G.; Lamarre, S.G.; Joly, D.L.; Filion, M. Pyoluteorin and 2,4-diacetylphloroglucinol are major contributors to Pseudomonas protegens Pf-5 biocontrol against Botrytis cinerea in cannabis. Front. Microbiol. 2022, 13, 945498. [Google Scholar] [CrossRef] [PubMed]
- Kidarsa, T.A.; Goebel, N.C.; Zabriskie, T.M.; Loper, J.E. Phloroglucinol mediates cross-talk between the pyoluteorin and 2,4-diacetylphloroglucinol biosynthetic pathways in Pseudomonas fluorescens Pf-5. Mol. Microbiol. 2011, 81, 395–414. [Google Scholar] [CrossRef]
- Zhang, Q.X.; Kong, X.W.; Li, S.Y.; Chen, X.J.; Chen, X.J. Antibiotics of Pseudomonas protegens FD6 are essential for biocontrol activity. Australas. Plant Pathol. 2020, 49, 307–317. [Google Scholar] [CrossRef]
- Dash, P.K.; Gupta, P.; Panwar, B.S.; Rai, R. Isolation, cloning and characterization of phlB gene from an Indian strain of Gram negative soil bacteria Pseudomonas fluorescens. Indian J. Exp. Biol. 2020, 58, 412–419. [Google Scholar] [CrossRef]
- Liang, F.; Zhang, B.; Yang, Q.; Zhang, Y.; Zheng, D.; Zhang, L.; Yan, Q.; Wu, X. Cyclic-di-GMP Regulates the Quorum-Sensing System and Biocontrol Activity of Pseudomonas fluorescens 2P24 through the RsmA and RsmE Proteins. Appl. Environ. Microbiol. 2020, 86, e02016-20. [Google Scholar] [CrossRef] [PubMed]
- Landa, B.B.; Mavrodi, O.V.; Raaijmakers, J.M.; McSpadden Gardener, B.B.; Thomashow, L.S.; Weller, D.M. Differential Ability of Genotypes of 2,4-Diacetylphloroglucinol-Producing Pseudomonas fluorescens Strains to Colonize the Roots of Pea Plants. Appl. Environ. Microbiol. 2002, 68, 3226–3237. [Google Scholar] [CrossRef] [PubMed]
- Mavrodi, O.V.; McSpadden Gardener, B.B.; Mavrodi, D.V.; Bonsall, R.F.; Weller, D.M.; Thomashow, L.S. Genetic Diversity of phlD from 2,4-Diacetylphloroglucinol-Producing Fluorescent Pseudomonas spp. Phytopathology 2001, 91, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Weller, D.M.; Landa, B.B.; Mavrodi, O.V.; Schroeder, K.L.; De La Fuente, L.; Blouin Bankhead, S.; Allende Molar, R.; Bonsall, R.F.; Mavrodi, D.V.; Thomashow, L.S. Role of 2,4-Diacetylphloroglucinol-Producing Fluorescent Pseudomonas spp. in the Defense of Plant Roots. Plant Biol. 2007, 9, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Bangera, G.M. Characterization of a Genomic Locus Required for Synthesis of the Antibiotic 2,4-diacetylphloroglucinol by the Biological Control Agent Pseudomonas fluorescens Q2-87. Mol. Plant-Microbe Interact. 1996, 9, 083. [Google Scholar] [CrossRef] [PubMed]
- Abbas, A.; McGuire, J.E.; Crowley, D.; Baysse, C.; Dow, M.; O’Gara, F. The putative permease PhlE of Pseudomonas fluorescens F113 has a role in 2,4-diacetylphloroglucinol resistance and in general stress tolerance. Microbiology 2004, 150, 2443–2450. [Google Scholar] [CrossRef]
- Bottiglieri, M.; Keel, C. Characterization of PhlG, a Hydrolase That Specifically Degrades the Antifungal Compound 2,4-Diacetylphloroglucinol in the Biocontrol Agent Pseudomonas fluorescens CHA0. Appl. Environ. Microbiol. 2006, 72, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Yang, R.; Zhao, R.-X.; Han, J.-T.; Jia, W.-J.; Li, D.-Y.; Wang, Y.; Zhang, N.; Wu, Y.; Zhang, L.-Q.; et al. Transcriptional Regulator PhlH Modulates 2,4-Diacetylphloroglucinol Biosynthesis in Response to the Biosynthetic Intermediate and End Product. Appl. Environ. Microbiol. 2017, 83, e01419-17. [Google Scholar] [CrossRef]
- Zhao, M.-M.; Lyu, N.; Wang, D.; Wu, X.-G.; Zhao, Y.-Z.; Zhang, L.-Q.; Zhou, H.-Y. PhlG mediates the conversion of DAPG to MAPG in Pseudomonas fluorescens 2P24. Sci. Rep. 2020, 10, 4296. [Google Scholar] [CrossRef]
- He, Y.-X.; Huang, L.; Xue, Y.; Fei, X.; Teng, Y.-B.; Rubin-Pitel, S.B.; Zhao, H.; Zhou, C.-Z. Crystal Structure and Computational Analyses Provide Insights into the Catalytic Mechanism of 2,4-Diacetylphloroglucinol Hydrolase PhlG from Pseudomonas fluorescens. J. Biol. Chem. 2010, 285, 4603–4611. [Google Scholar] [CrossRef] [PubMed]
- Saitou, H.; Watanabe, M.; Maruyama, K. Molecular and Catalytic Properties of 2,4-Diacetylphloroglucinol Hydrolase (PhlG) from Pseudomonas sp. YGJ3. Biosci. Biotechnol. Biochem. 2012, 76, 1239–1241. [Google Scholar] [CrossRef]
- Abbas, A.; Morrissey, J.P.; Marquez, P.C.; Sheehan, M.M.; Delany, I.R.; O’Gara, F. Characterization of Interactions between the Transcriptional Repressor PhlF and Its Binding Site at the phlA Promoter in Pseudomonas fluorescens F113. J. Bacteriol. 2002, 184, 3008–3016. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Liu, J.; Zhang, W.; Zhang, L. Multiple-Level Regulation of 2,4-Diacetylphloroglucinol Production by the Sigma Regulator PsrA in Pseudomonas fluorescens 2P24. PLoS ONE 2012, 7, e50149. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.L.; He, Z.; Wibowo, M.; Jelsbak, L. A Whole-Cell Biosensor for Detection of 2,4-Diacetylphloroglucinol (DAPG)-Producing Bacteria from Grassland Soil. Appl. Environ. Microbiol. 2021, 87, e01400-20. [Google Scholar] [CrossRef] [PubMed]
- Schnider-Keel, U.; Seematter, A.; Maurhofer, M.; Blumer, C.; Duffy, B.; Gigot-Bonnefoy, C.; Reimmann, C.; Notz, R.; Défago, G.; Haas, D.; et al. Autoinduction of 2,4-Diacetylphloroglucinol Biosynthesis in the Biocontrol Agent Pseudomonas fluorescens CHA0 and Repression by the Bacterial Metabolites Salicylate and Pyoluteorin. J. Bacteriol. 2000, 182, 1215–1225. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, A.; Saitou, H.; Mori, T.; Matano, I.; Sugisaki, H.; Maruyama, K. Molecular and Catalytic Properties of Monoacetylphloroglucinol Acetyltransferase from Pseudomonas sp. YGJ3. Biosci. Biotechnol. Biochem. 2012, 76, 559–566. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yu, X.; Yan, X.; Zhang, M.; Zhang, L.; He, Y. Flavonoids repress the production of antifungal 2,4-DAPG but potentially facilitate root colonization of the rhizobacterium Pseudomonas fluorescens. Environ. Microbiol. 2020, 22, 5073–5089. [Google Scholar] [CrossRef]
- Delany, I.; Sheehan, M.M.; Fenton, A.; Bardin, S.; Aarons, S.; O’Gara, F. Regulation of production of the antifungal metabolite 2,4-diacetylphloroglucinol in Pseudomonas fluorescens F113: Genetic analysis of phlF as a transcriptional repressor The GenBank accession number for the sequence reported in this paper is AF129856. Microbiology 2000, 146, 537–546. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, B.; Wu, H.; Wu, X.; Yan, Q.; Zhang, L.-Q. Pleiotropic effects of RsmA and RsmE proteins in Pseudomonas fluorescens 2P24. BMC Microbiol. 2020, 20, 191. [Google Scholar] [CrossRef] [PubMed]
- Biessy, A.; Filion, M. Phloroglucinol Derivatives in Plant-Beneficial Pseudomonas spp.: Biosynthesis, Regulation, and Functions. Metabolites 2021, 11, 182. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Gu, G.-Q.; Chen, W.; Gao, L.-J.; Wu, X.-H.; Zhang, L.-Q. The outer membrane protein OprF and the sigma factor SigX regulate antibiotic production in Pseudomonas fluorescens 2P24. Microbiol. Res. 2018, 206, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Xiao-Gang, W.; Hui-Mei, D.; Tao, T.; Nan, Y.; Hong-You, Z.; Li-Qun, Z. Effect of the hfq gene on 2,4-diacetylphloroglucinol production and the PcoI/PcoR quorum-sensing system in Pseudomonas fluorescens 2P24. FEMS Microbiol. Lett. 2010, 309, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.; Yan, Q.; Zhang, B.; Zhang, L.; Wu, X. Effect of the Monothiol Glutaredoxin GrxD on 2,4-Diacetylphloroglucinol Biosynthesis and Biocontrol Activity of Pseudomonas fluorescens 2P24. Front. Microbiol. 2022, 13, 920793. [Google Scholar] [CrossRef] [PubMed]
- Humair, B.; González, N.; Mossialos, D.; Reimmann, C.; Haas, D. Temperature-responsive sensing regulates biocontrol factor expression in Pseudomonas fluorescens CHA0. ISME J. 2009, 3, 955–965. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, W.; Wu, X.; Zhang, L. [Effect of retS gene on biosynthesis of 2, 4-diacetyl-phloroglucinol in Pseudomonas fluorescens 2P24]. Wei Sheng Wu Xue Bao = Acta Microbiol. Sin. 2013, 53, 118–126. [Google Scholar]
- He, Y.; Suzuki, S.; Aono, T.; Oyaizu, H. Importance of 2,4-DAPG in the biological control of brown patch by Pseudomonas fluorescens HP72 and newly identified genes involved in 2,4-DAPG biosynthesis. Soil Sci. Plant Nutr. 2004, 50, 1287–1293. [Google Scholar] [CrossRef]
- Zhou, T.-T.; Li, C.-Y.; Chen, D.; Wu, K.; Shen, Q.-R.; Shen, B. phlF− mutant of Pseudomonas fluorescens J2 improved 2,4-DAPG biosynthesis and biocontrol efficacy against tomato bacterial wilt. Biol. Control 2014, 78, 1–8. [Google Scholar] [CrossRef]
- Aarons, S.; Abbas, A.; Adams, C.; Fenton, A.; O’Gara, F. A Regulatory RNA (PrrB RNA) Modulates Expression of Secondary Metabolite Genes in Pseudomonas fluorescens F113. J. Bacteriol. 2000, 182, 3913–3919. [Google Scholar] [CrossRef]
- Heeb, S.; Itoh, Y.; Nishijyo, T.; Schnider, U.; Keel, C.; Wade, J.; Walsh, U.; O’Gara, F.; Haas, D. Small, Stable Shuttle Vectors Based on the Minimal pVS1 Replicon for Use in Gram-Negative, Plant-Associated Bacteria. Mol. Plant-Microbe Interact. 2000, 13, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Kwak, Y.-S.; Han, S.; Thomashow, L.S.; Rice, J.T.; Paulitz, T.C.; Kim, D.; Weller, D.M. Saccharomyces cerevisiae Genome-Wide Mutant Screen for Sensitivity to 2,4-Diacetylphloroglucinol, an Antibiotic Produced by Pseudomonas fluorescens. Appl. Environ. Microbiol. 2011, 77, 1770–1776. [Google Scholar] [CrossRef] [PubMed]
- Troppens, D.M.; Dmitriev, R.I.; Papkovsky, D.B.; O’Gara, F.; Morrissey, J.P. Genome-wide investigation of cellular targets and mode of action of the antifungal bacterial metabolite 2,4-diacetylphloroglucinol in Saccharomyces cerevisiae. FEMS Yeast Res. 2013, 13, 322–334. [Google Scholar] [CrossRef] [PubMed]
- Stepanov, A.A.; Vasilchenko, A.S. 2,4-Diacetylphloroglucinol against Candida albicans: Biofilm Formation, Aspartyl Protease Production and Ultrastructure Changes. bioRxiv 2022. [Google Scholar] [CrossRef]
- Schouten, A.; van den Berg, G.; Edel-Hermann, V.; Steinberg, C.; Gautheron, N.; Alabouvette, C.; de Vos, C.H. (Ric), Lemanceau, P.; Raaijmakers, J.M. Defense Responses of Fusarium oxysporum to 2,4-Diacetylphloroglucinol, a Broad-Spectrum Antibiotic Produced by Pseudomonas fluorescens. Mol. Plant-Microbe Interact. 2004, 17, 1201–1211. [Google Scholar] [CrossRef] [PubMed]
- Suresh, P.; Varathraju, G.; Shanmugaiah, V.; Almaary, K.S.; Elbadawi, Y.B.; Mubarak, A. Partial purification and characterization of 2, 4-diacetylphloroglucinol producing Pseudomonas fluorescens VSMKU3054 against bacterial wilt disease of tomato. Saudi J. Biol. Sci. 2021, 28, 2155–2167. [Google Scholar] [CrossRef] [PubMed]
- Mazzola, M.; Fujimoto, D.K.; Thomashow, L.S.; Cook, R.J. Variation in Sensitivity of Gaeumannomyces graminis to Antibiotics Produced by Fluorescent Pseudomonas spp. and Effect on Biological Control of Take-All of Wheat. Appl. Environ. Microbiol. 1995, 61, 2554–2559. [Google Scholar] [CrossRef] [PubMed]
- Keel, C. Suppression of Root Diseases by Pseudomonas fluorescens CHA0: Importance of the Bacterial Secondary Metabolite 2,4-Diacetylphloroglucinol. Mol. Plant-Microbe Interact. 1992, 5, 4. [Google Scholar] [CrossRef]
- Isnansetyo, A.; Horikawa, M.; Kamei, Y. In vitro anti-methicillin-resistant Staphylococcus aureus activity of 2,4-diacetylphloroglucinol produced by Pseudomonas sp. AMSN isolated from a marine alga. J. Antimicrob. Chemother. 2001, 47, 724–725. [Google Scholar] [CrossRef]
- Julian, W.T.; Vasilchenko, A.V.; Shpindyuk, D.D.; Poshvina, D.V.; Vasilchenko, A.S. Bacterial-Derived Plant Protection Metabolite 2,4-Diacetylphloroglucinol: Effects on Bacterial Cells at Inhibitory and Subinhibitory Concentrations. Biomolecules 2020, 11, 13. [Google Scholar] [CrossRef]
- Velusamy, P.; Immanuel, J.E.; Gnanamanickam, S.S.; Thomashow, L. Biological control of rice bacterial blight by plant-associated bacteria producing 2,4-diacetylphloroglucinol. Can. J. Microbiol. 2006, 52, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Meyer, S.L.F.; Everts, K.L.; Gardener, B.M.; Masler, E.P.; Abdelnabby, H.M.E.; Skantar, A.M. Assessment of DAPG-producing Pseudomonas fluorescens for Management of Meloidogyne incognita and Fusarium oxysporum on Watermelon. J. Nematol. 2016, 48, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, I.A.; Shaukat, S.S. Plant Species, Host Age and Host Genotype Effects on Meloidogyne incognita Biocontrol by Pseudomonas fluorescens Strain CHA0 and its Genetically-Modified Derivatives. J. Phytopathol. 2003, 151, 231–238. [Google Scholar] [CrossRef]
- Siddiqui, I.A.; Shaukat, S.S. Suppression of root-knot disease by Pseudomonas fluorescens CHA0 in tomato: Importance of bacterial secondary metabolite, 2,4-diacetylpholoroglucinol. Soil Biol. Biochem. 2003, 35, 1615–1623. [Google Scholar] [CrossRef]
- Cronin, D.; Moenne-Loccoz, Y.; Fenton, A.; Dunne, C.; Dowling, D.N.; O’gara, F. Role of 2,4-Diacetylphloroglucinol in the Interactions of the Biocontrol Pseudomonad Strain F113 with the Potato Cyst Nematode Globodera rostochiensis. Appl. Environ. Microbiol. 1997, 63, 1357–1361. [Google Scholar] [CrossRef] [PubMed]
- Asolkar, R.; Cordova-Kreylos, A.L.; Todd, C. Control of Phytopathogenic Microorganisms with Pseudomonas sp. and Substances and Compositions Derived Therefrom. Canadian Patent No. CA2865237C, Patent of PRC No. CN104254611A, 21 August 2014. [Google Scholar]
- Huang, H.; Asolkar, R.; Marrone, P.; Cordova-Kreylos, A.L. Use of Proteins Isolated from Pseudomonas to Control Molluscs. U.S. Patent No. US 8,728,754 B1, 15 March 2013. [Google Scholar]
- Frost, J.W. Biosynthesis of Phloroglucinol and Preparation of 1,3-Dihydroxybenzene Therefrom. Patent of PRC No. CN101084311A, 10 November 2005. [Google Scholar]
- Raaijmakers, J.M.; Weller, D.M.; Thomashow, L.S.; Cook, R.J. Biocontrol Agents for Take-All. U.S. Patent No. US 6,447,770 B1, 14 September 1999. [Google Scholar]
- Thomashow, L.S.; Bangera, M.; Weller, D.M.; Cook, J.R. Sequences for Production of 2,4-Diacetylphloroglucinol and Methods. International Patent No. WO1997001572A1, 26 June 1996. [Google Scholar]
- Barea, J.M.; Andrade, G.; Bianciotto, V.; Dowling, D.; Lohrke, S.; Bonfante, P.; O’Gara, F.; Azcon-Aguilar, C. Impact on Arbuscular Mycorrhiza Formation of Pseudomonas Strains Used as Inoculants for Biocontrol of Soil-Borne Fungal Plant Pathogens. Appl. Environ. Microbiol. 1998, 64, 2304–2307. [Google Scholar] [CrossRef] [PubMed]
- Resca, R.; Basaglia, M.; Poggiolini, S.; Vian, P.; Bardin, S.; Walsh, U.F.; Enriquez Barreiros, C.M.; O’Gara, F.; Nuti, M.P.; Casella, S.; et al. An integrated approach for the evaluation of biological control of the complex Polymyxa betae/Beet Necrotic Yellow Vein Virus, by means of seed inoculants. Plant Soil 2001, 232, 215–226. [Google Scholar] [CrossRef]
- Girlanda, M.; Perotto, S.; Moenne-Loccoz, Y.; Bergero, R.; Lazzari, A.; Defago, G.; Bonfante, P.; Luppi, A.M. Impact of Biocontrol Pseudomonas fluorescens CHA0 and a Genetically Modified Derivative on the Diversity of Culturable Fungi in the Cucumber Rhizosphere. Appl. Environ. Microbiol. 2001, 67, 1851–1864. [Google Scholar] [CrossRef]
- Natsch, A.; Keel, C.; Hebecker, N.; Laasik, E.; Défago, G. Impact of Pseudomonas fluorescens strain CHA0 and a derivative with improved biocontrol activity on the culturable resident bacterial community on cucumber roots. FEMS Microbiol. Ecol. 1998, 27, 365–380. [Google Scholar] [CrossRef]
- Niemann, S.; Keel, C.; Pühler, A.; Selbitschka, W. Biocontrol strain Pseudomonas fluorescens CHA0 and its genetically modified derivative with enhanced biocontrol capability exert comparable effects on the structure of a Sinorhizobium meliloti population in gnotobiotic systems. Biol. Fertil. Soils 1997, 25, 240–244. [Google Scholar] [CrossRef]
- Huang, Z.; Thomashow, L.S.; Mavrodi, D.V.; Raaijmakers, J.M.; Weller, D.M.; Cook, J.R. Transgenic Strains of Pseudomonas for Biocontrol of Plant Root Diseases. U.S. Patent No. US 6,277,625 B1, 18 December 1997. [Google Scholar]
- Gangwar, A.; Kumar, P.; Singh, R.; Kush, P. Recent Advances in Mupirocin Delivery Strategies for the Treatment of Bacterial Skin and Soft Tissue Infection. Future Pharmacol. 2021, 1, 80–103. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).